
Evidence for Anti-Pseudogymnoascus destructans (Pd) Activity of Propolis
by
 and
and
Soumya Ghosh
1,†, 
Robyn McArthur
1,†,
Zhi Chao Guo
1,
Rory McKerchar
1,
Kingsley Donkor
1,
Jianping Xu
2 and
and
Naowarat Cheeptham
1,* 1
Department of Biological Sciences, Thompson Rivers University, Kamloops, BC V2C 0C8, Canada
2
Department of Biology, McMaster University, Hamilton, ON L8S 4K1 Canada
*
Author to whom correspondence should be addressed.
†
Equally contributed author.
Antibiotics 2018, 7(1), 2; https://doi.org/10.3390/antibiotics7010002
Submission received: 16 October 2017
/
Revised: 5 December 2017
/
Accepted: 20 December 2017
/
Published: 21 December 2017
(This article belongs to the Special Issue Top 35 of Antibiotics Travel Awards 2017)
Abstract
:White-nose syndrome (WNS) in bats, caused by Pseudogymnoascus destructans (Pd), is a cutaneous infection that has devastated North American bat populations since 2007. At present, there is no effective method for controlling this disease. Here, we evaluated the effect of propolis against Pd in vitro. Using Sabouraud dextrose agar (SDA) medium, approximately 1.7 × 107 conidia spores of the Pd strain M3906-2/mL were spread on each plate and grown to form a consistent lawn. A Kirby–Bauer disk diffusion assay was employed using different concentrations of propolis (1%, 2%, 3%, 4%, 5%, 10%, 15%, 20%, 25%), in plates incubated at 8 °C and 15 °C. At 8 °C and 15 °C, as the concentration of propolis increased, there was an increasing zone of inhibition (ZOI), reaching the highest degree at 10% and 25% concentrations, respectively. A germule suppression assay showed a similar effect on Pd conidia germination. A MALDI-TOF-MS analysis of propolis revealed multiple constituents with a potential anti-Pd activity, including cinnamic acid, p-coumaric acid, and dihydrochalcones, which could be further tested for their individual effects. Our study suggests that propolis or its individual constituents might be suitable products against Pd.
1. Introduction
White-nose syndrome (WNS) has devastated many eastern North American bat populations since 2007, killing more than six million bats [1]. Since the first observations of mortality at a cave near Albany, New York in 2007, WNS has spread to 31 US states and 5 eastern Canadian provinces [2], most recently appearing in 2017 in the states of Mississippi, Texas, and Washington, USA [2]. The etiological agent of WNS is Pseudogymnoascus destructans (Pd), a psychrophilic fungus that grows optimally on hibernating bats at temperatures between 12–16 °C [3].
Pseudogymnoascus destructans (Pd) preferentially infects thinly haired regions on the skin of hibernating bats, and is able to degrade collagen and invade living tissues [4]. Hibernating bats lower their body temperature to near-ambient temperature during torpor bouts [5]. During hibernation, bats generally seek microclimates that remain above freezing and can be as warm as 15 °C or more for some species [5,6,7], and are relatively high in humidity. Bats generally adapt to the temperature of their surroundings to maximize their energy budget [8]. The similarity between the preferred hibernating environment of bats and the optimal growth condition of the Pd pathogen is a major contributor to the WNS epidemic. Pd infection disrupts the normal torpor and arousal cycles of hibernating bats [9], causing premature depletion of fat reserves, in addition to electrolyte imbalances and dehydration, resulting in mortality [10]. Bats are essential components of both the natural, agricultural, and other human ecosystems [11]. They play important roles in maintaining ecosystem stability, consume insects that are human or animal pests, and redistribute nutrients through their guano [11,12]. To reduce bat mortality and eliminate the likelihood of species extinction by Pd, it is essential to identify effective methods to control Pd.
In regard to investigations of anti-Pd agents, several recent studies have identified putative anti-Pd agents, including (i) volatile compounds produced by the bacteria Rhodococcus rhodochrous DAP96253 [13] and by Pseudomonas spp. isolated from bat wings [14]; (ii) cold-pressed, terpeneless orange oil (CPT) [15]; and (iii) sesquiterpene trans, trans-farnesol (Candida albicans quorum-sensing compound) [13]. Our goal was to identify alternative or additional potent treatments against Pd using substances likely to be unharmful to cave environments and to bats.
Propolis is a resinous substance produced by honey bees in beehives throughout the year [16,17]. Stingless bees are widely spread, especially in the tropical and subtropical areas of the world. Propolis produced from such bees possesses therapeutic properties [18], including antimicrobial, antitumor [18], antioxidant [19], anti-stimulant [20], anti-inflammatory [21,22,23,24], antiulcer [22,23,24,25], and anti-HIV activities [25]. For instance, two compounds, cardanol and cardol, isolated from a Thai propolis, possessed antiproliferation and cytotoxicity against carcinomas originated from the lungs, the liver, and the colon [26]. Khacha-ananda et al. 2016 [19] found that ethanolic extracts of propolis (EEP) obtained from Chiang Mai, Thailand, exhibited higher antioxidant activity than EEP from other sources. In 2005, Hu et al. [23] showed that both the ethanol and the water extracts of propolis had anti-inflammatory activities in mice and rats [21]. Significant anti-HIV activities [EC(50) < 0.1 µg/mL, TI > 186] resulted from moronic acid (triterpenoids) isolated from a Brazilian propolis [25]. Additionally, because of its antiviral, antibacterial, and antifungal activities, propolis has been used in human healthcare to treat colds, wounds, ulcers, and rheumatism [27,28]. Propolis was found to exhibit antagonistic effects against a number of Gram-positive cocci and rods [29]. A recent study by Shimizu et al. 2011 [30], showed that the ethanol extract of propolis from Brazil had antiviral activities. When administered orally or cutaneously to herpes simplex virus type 1 (HSV-1)-infected mice, the ethanol extract of propolis significantly reduced the herpetic skin lesions and enhanced delayed-type hypersensitivity [30]. Silici et al. 2006 [31] reported that propolis had antifungal activities against 15 strains belonging to four species of yeasts isolated from patients with superficial mycoses.
Traditionally, studies on the medicinal benefits of propolis have attributed its effects to its complex composition and to the synergistic effects among its complex chemical constituents [32,33]. Another emerging theme is that the chemical composition of propolis is highly dependent on the geographical location, botanical origin [34], and bee species [35]. In different ecosystems, there are different plant species, and these plants can vary in their secretion and exudates, and therefore provide diverse food sources to bees [16]. Thus, the variability in chemical composition among propolis from different sources can be large. For example, propolis produced in the Pacific region contains geranyl flavanones which are also a typical component for the African propolis [17]. The green propolis of Brazil has prenylated phenylpropanoids (e.g., artepillin C) and diterpenes as major components [17], while the propolis of temperate regions consists of flavonoids lacking the B-ring substituents, namely, chrysin, galangin, pinocembrin, pinobanksin, caffeic acid phenethyl ester [36]. These unique mixtures of constituents from different sources likely contribute to the observed multiple effects including not only the broad biological effects described above, but also the inhibition of nuclear factor κ-B, cell proliferation, cell arrest, and apoptosis [17,36].
In this study, we tested the effectiveness of the commercially available propolis purchased from Natural Factors in Coquitlam, British Columbia, Canada on the WNS agent Pd. We investigated the in vitro antagonistic properties of propolis against Pd. We also tested the inhibition of germule development when incubated with propolis. This study is the first investigation of the anti-Pd activities of propolis.
2. Results
2.1. Kirby–Bauer Diffusion Assay
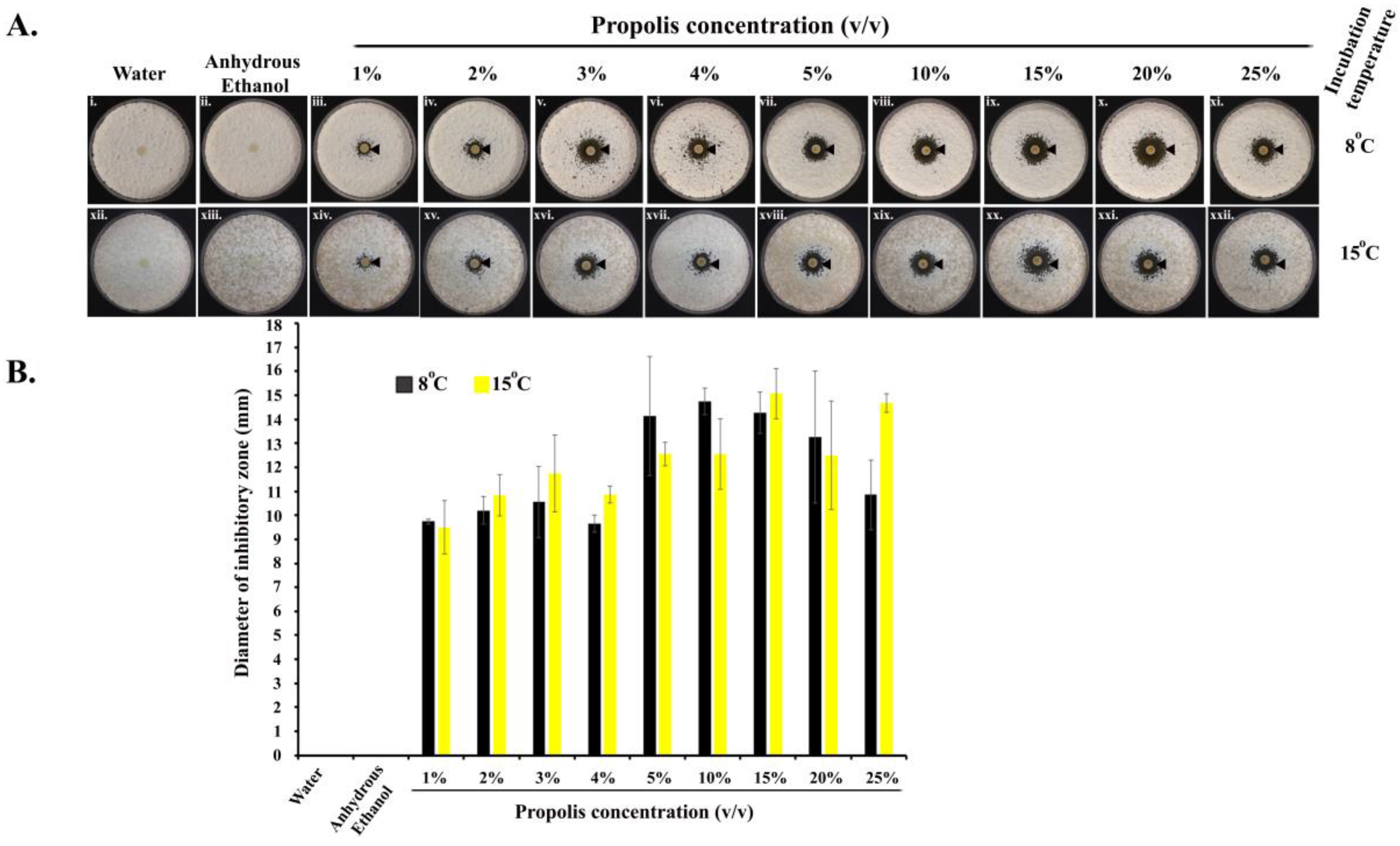
Propolis exhibited anti-Pd activities at all the concentrations (1–25%) we tested (in quadruplets), as revealed by a clear ‘zone of inhibition’ around each of the impregnated paper discs, in comparison to discs treated with water or anhydrous ethanol (Figure 1(Ai,ii,xii,xiii)). ANOVA tests showed that the different concentrations of propolis differed significantly in their inhibitory effects (Figure 1(Aiii–xi,xiv–xxii)). Propolis significantly inhibited the growth of Pd at 8 °C (one-way ANOVA; F = 8.309; df = 8; p = 0.100−4) as well as at 15 °C (one-way ANOVA; F = 8.704; df = 8; p = 0.839−5). Along the concentration gradient, at 8 °C, the diameter of the inhibitory zones initially increased with the increase in propolis concentration, reaching the highest value at the propolis concentration of 10%, and showed some decline at higher propolis concentrations. At 15 °C, though there were some variations, the inhibition also increased with increasing propolis concentrations, reaching the highest level at the concentration of 25% of propolis. The pairwise post-hoc T-test results are shown in Supplementary Tables S1 and S2.
2.2. Suppression of Germination of Pd Spores
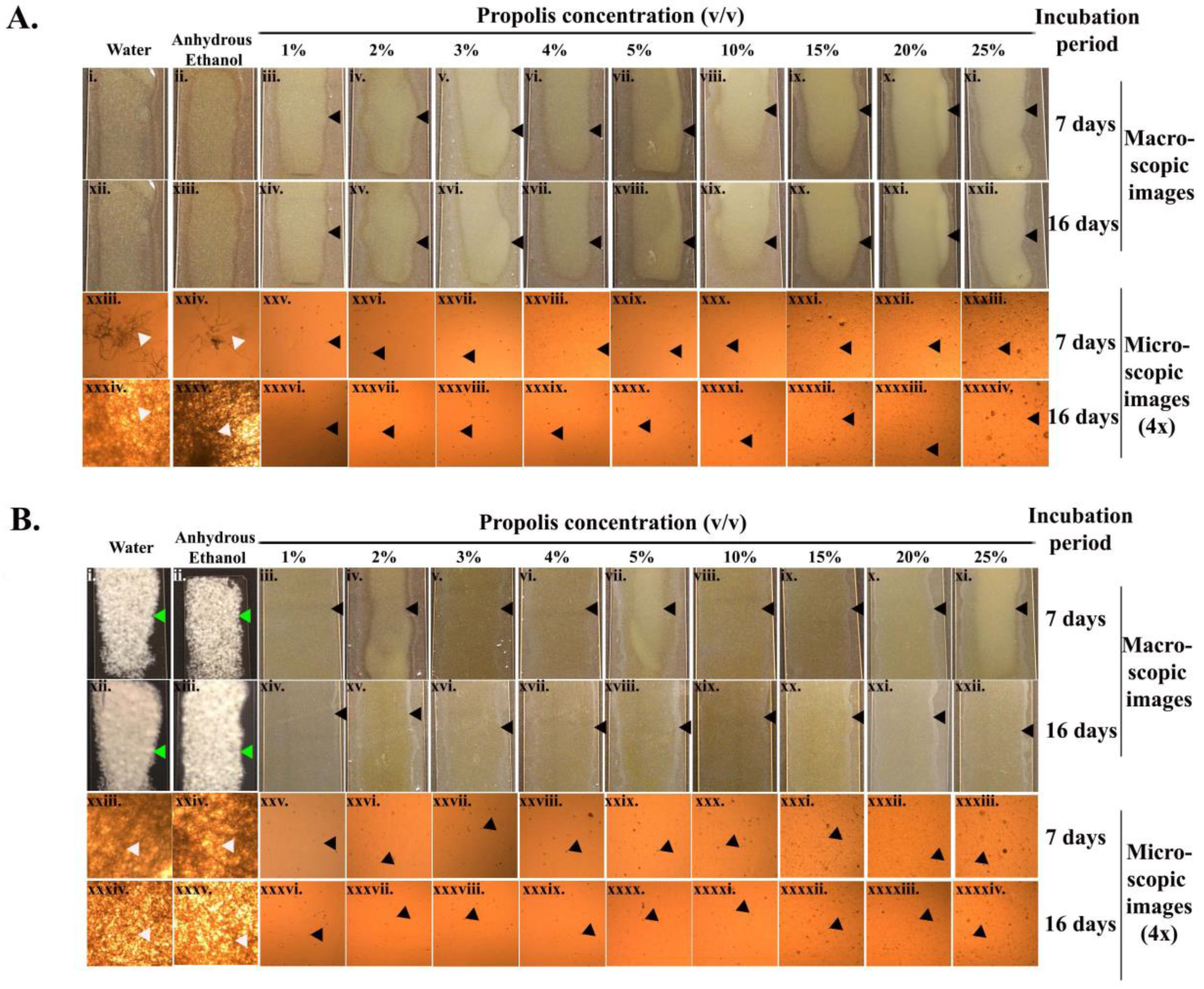
As revealed by the macroscopic and microscopic assay images (Figure 2A,B), there was a complete inhibition of Pd sporulation with all propolis concentrations throughout the entire incubation period (16 days) at both incubation temperatures tested. On the seventh day of incubation at 8 °C, more mycelial extensions (4× microscopic images, Figure 2(Axxiii,xxiv)) were observed in the samples treated with water or anhydrous ethanol. The mycelial growth became more confluent amongst the Pd spores treated with water and ethanol by the 16th day of incubation (Figure 2(Axxxiv,xxxv)). At 15 °C, the Pd spores treated with water and ethanol exhibited a confluent growth from the seventh day of incubation (Figure 2(Bi,ii,xxiii,xxiv)).
2.3. Microscopic Examination of the Treated Pd Spores
2.4. Chemical Composition of Propolis
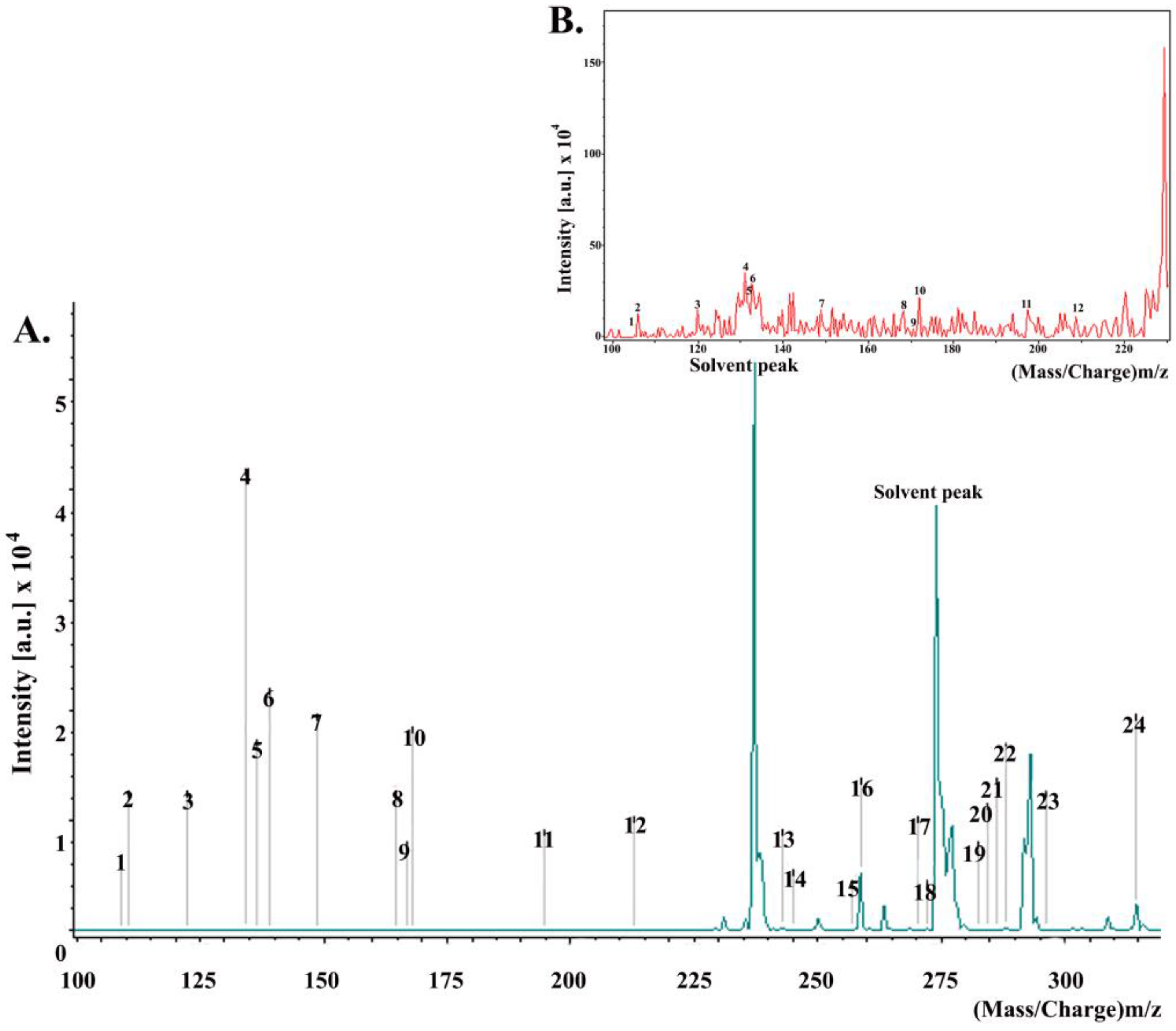
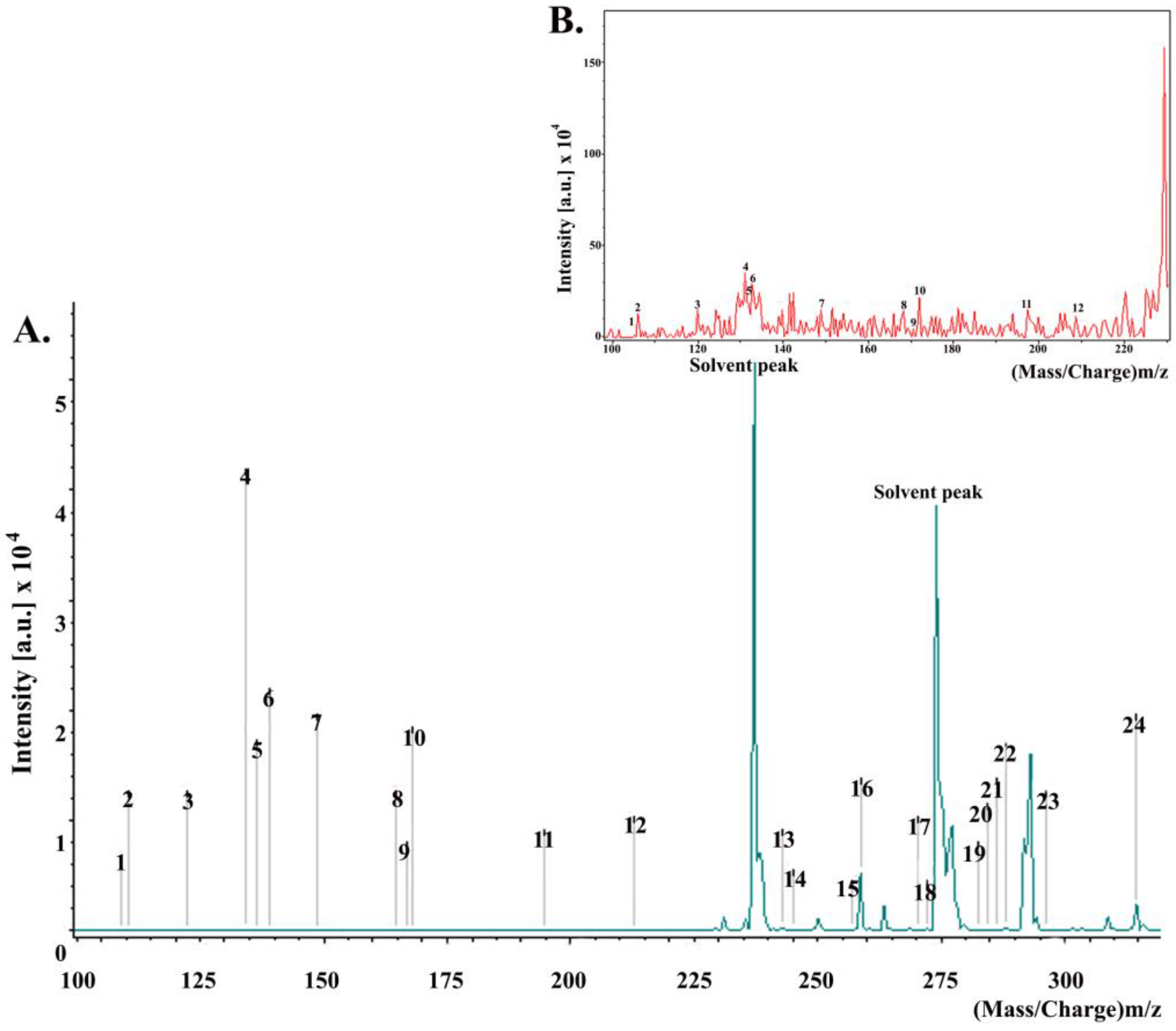
MALDI spectra (Figure 4) revealed that the major constituents of the propolis used in this study were aromatic acids, i.e., cinnamic acid and p-coumaric acid; dihydrochalcones, i.e., 2,4,6-trihydroxydihydrochalcone; fatty acids, i.e., stearic acid, palmitic acid; esters, i.e., benzyl methoxybenzoate.
3. Discussion
The populations of hibernating bats in North America are declining at unprecedented rates because of WNS [1,37]. Because insectivorous bats usually eat insects, they play important roles in the ecosystem and provide valuable pest control services to the agricultural and forestry sectors of the North American economy [11]. Our research has identified a new potential tool for combatting WNS that is threatening many bats across the continent. We have discovered that bee propolis may be used as an effective antifungal agent against Pd, the causative agent of WNS.
Our study has revealed that even low concentration (1%) of commercially available propolis (65% tincture) can completely inhibit Pd spore germination at both 8 °C and 15 °C. Unfortunately, Pd grows optimally between 8–16 °C [3], and the hibernating environment creates an ideal condition for Pd growth on hibernating bats. The incubation temperatures used in our testing were thus representative of the effective hibernation and fungal infection range that are also found in cave environments [38]. Additionally, the bioassay plates were kept until the 60th day after the 22nd day of observation, and we never found any encroachment of growth of Pd spores in the observed ZOI. This may indicate that propolis can completely inhibit Pd spore germination for up to 60 days when tested in the lab setting.
Propolis has previously been shown to exhibit antifungal, antibacterial, and antiviral properties and therefore has been widely used in human healthcare for treating ulcers, wounds, and rheumatisms [27,28]. We employed the Kirby–Bauer diffusion assay to identify fungicidal activities of propolis, while the germule suppression assay was performed in order to understand the possible inhibitory mechanism of the propolis against Pd spores germination. Similar studies by Cornelison el al. 2014 [13,39] have shown that bacteria-derived volatile compounds that include decanal, 2-ethyl-hexanol, nonanal, benzothiole, and N, N-dimethyloctylamine, completely inhibited the growth of conidia and radial mycelial extensions. Moreover, our findings show a deformation of the Pd spores exposed to propolis, a result consistent with a possible fungicidal mechanism of action. The deformation of the fungal conidia by propolis has never been reported before.
Though the propolis used in this study was purchased from a western Canadian company, Natural Factors, the company obtained the raw propolis materials from a variety of sources and geographic regions. Further examinations of our purchased propolis traced its origin to Mongolia. Unfortunately, no further information could be obtained, including the specific region within Mongolia, or the plants that bees were feeding on (personal communication). However, consistent with previous findings [40,41,42], our MALDI-TOF analysis of the tested propolis identified a range of constituents. These included benzyl benzoate, benzyl methoxybenzoate, benzyl dihydroxybenzoate, hydroxyacetophenone, 2,4,6-trihydroxydihydrochalcone, pinostrobin chalcone, 2,6-dihydroxy-4-methoxydihydrochalcone, 2,4,6-trihydroxy-4-methoxydihydrochalcone, cinnamic acid, and p-coumaric acid as major constituents. A previous study of Canadian propolis from two regions showed that the propolis from Victoria contained mainly p-hydroxyacetophenone, benzyl hydroxybenzoate, cinnamic acid, and dihydrochalcones, while that from Richmond had large amounts of cinnamic acid and p-coumaric acid [40]. However, both propolis samples showed significant antioxidant properties with a high level of radical scavenging activity. The other remaining compounds identified in our propolis also showed some geographic specificity in previous studies. For example, 3,3-dimethylallyl caffeate was reported from European poplar-type propolis [41,42], while we identified cinnamyl caffeate in our propolis. Other compounds from diverse locations include hydroquinone (Burdock et al. 1998 [42]) and benzyl alcohol [43]. Benzyl alcohol has anti-inflammatory, antibacterial, antitumour, hepatoprotective, and antioxidant activities [41]. Benzoic acid and 4-hydroxybenoic acid found in our samples were previously found in an Iranian propolis, and have shown antibacterial properties [44]. Other constituents revealed in our propolis included ferulic acid, oleic acid, stearic acid, palmitic acid, and pinobanksin 3-O-acetate. An earlier analysis of Anatolian propolis also identified the above mentioned compounds and showed that they exhibited antibacterial activities against Gram-positive bacteria such as Staphylococcus aureus (6538-P), Streptococcus sobrinus, Staphylococcus epidermidis, Streptococcus mutans, Enterococcus faecalis, and Micrococcus luteus [45]. Certain Gram-negative bacteria, such as Escherichia coli, Salmonella typhimurium, Pseudomonas aeruginosa, and Enterobacter aerogenes and yeast such as Candida albicans, C. tropicalis, and C. krusei were reported to be susceptible to the Anatolian propolis [45]. Lastly, Sakuranetin, one of the flavonoids identified in our propolis, was reported to exhibit antimicrobial activities against oral pathogens [46].
We have confirmed the complete inhibition of Pd spore germination even at a low concentration of propolis (1%). Propolis, also called “bee glue” [21,47], is soluble in anhydrous ethanol, which eliminates the resinous and sticky properties of this substance making it suitable for application on roost substrates. Our study contributes to a growing portfolio of biological and chemical measures for controlling the growth of Pd [10,12,13,42]. Future applications on bats, and tests in wild hibernacula are required to test the effectiveness of propolis outside of a laboratory setting.
The US Fish and Wildlife Service (USFWS) has also recommended a number of decontaminants. The only appropriate methods for laboratory and field decontamination of equipment and clothing include ethanol (≥60%), isopropanol (≥60%), isopropyl alcohol wipes (70%), hydrogen peroxide wipes (3%), Accel®, Clorox® bleach, Clorox® wipes, Clorox® Clean-Up cleaner + bleach, Hibiclens®, and Lysol® IC quaternary disinfectant cleaner. Whether propolis would be useful as a decontamination substance is yet to be seen, in any case the substances listed in the current US national WNS decontamination protocol are more readily available. The toxicity of many of these listed decontamination substances is of minor concern for an equipment decontamination protocol, but it is of utmost concern when these substances are employed for the mitigation of a disease. Naturally sourced anti-Pd substances like propolis could provide treatment options that are generally considered safe for mammals. Previous studies reported that Greek and Roman physicians prescribed propolis as a mouth disinfectant and for the topical therapy of cutaneous and mucosal wounds in humans [16]. More recently, a study showed that propolis paste applied on dogs’ cutaneous wounds resulted in better wound re-epithelization, contraction, and total wound healing than a placebo [48]. However, no study has examined the effects of propolis on bats.
Overall, our study has demonstrated the complete inhibition of Pd spore germination by propolis. However, significant research is still required, for example an investigation on whether a longer period of testing time in the laboratory would yield any additional results further indicating a real potential of propolis as one of the treatment options for WNS. The potential activities of the individual constituents of propolis and their combinations, as well as their synergistic interactions against Pd spore germination and mycelial growth also need to be identified. At present, the medicinal benefits of propolis have been attributed to its complex composition and to the potential synergistic effects of its chemical constituents. In addition, the chemical composition of propolis is highly dependent on its geographical origin, on bee species, and on the botanical food sources of the bees. Further investigations are needed in order to determine whether propolis, or its individual ingredients or combinations thereof may be an option for the treatment of Pd-infected cave or bats.
4. Materials and Methods
4.1. Cultivation of Pd Spores
The P. destructans M3906-2 strain was used in this study. This Pd strain was previously described by Khankhet et al. (2014) [49]. The cultivation and isolation of the Pd spores were performed as previously described [37]. Pd cultures were maintained on Sabouraud dextrose agar (SDA) plates at 15 °C. Pd spores were isolated from cultures by submerging the conidial lawn in Conidia Harvesting Solution (CHS) (0.05% Tween 80, 0.9% NaCl) for 5 min followed by mechanical scrapping and filtration through glass wool as previously described [13,14]. The concentration of the Pd spores was quantified by a haemocytometer and the spores were stored in phosphate buffer saline (PBS) at 4 °C until further use.
4.2. Kirby–Bauer Diffusion Assay
One hundred microliter of isolated Pd spores (1 × 107 spores/mL) were mixed with 250 mL of Sabouraud dextrose agar (SDA) media supplemented with chloramphenicol (34 mg/L) at 50 °C (Fisher Scientific, Fairlawn, NJ, USA) to avoid bacterial contamination. Approximately 20–22 mL of the mixture was poured into each of the 85 mm petri plates. Plates were air-dried in a laminar airflow hood. Eight millimeter (diameter) paper discs (Toyo Roshi Kaisha Ltd., Tokyo, Japan) were soaked in different concentrations (1%, 2%, 3%, 4%, 5%, 10%, 15%, 20%, and 25%) of commercially available propolis (65% extract) (Natural Factors, Coquitlam, BC, Canada), air-dried, and placed in the center of each seeded SDA medium plate along with sterile water and anhydrous ethanol (<0.005% water) (Sigma Aldrich, St. Louis, MO, USA); the latter two conditions were used as controls. Anhydrous ethanol was tested since it was used as a solvent to dissolve propolis in all dilutions (as per the manufacturer’s instructions). All plates with different propolis concentrations were incubated at 8 °C or 15 °C, including control plates, in duplicate. Anti-Pd activities were identified as zones of inhibition around the impregnated paper discs, and the diameters were measured in millimeters with an electronic Vernier caliper (Guilin, Guangxi, China). Notably, the measurement of the diameters for the zones of inhibition were recorded on the 22nd and 15th day of incubation at 8 °C and 15 °C, respectively, since Pd spores germinated slower at 8 °C than at 15 °C. Each treatment was repeated four times. The bioassay plates were kept until the 60th day after the observation performed at the 22nd day.
To determine whether the concentrations of propolis differed in their inhibitory effects, we used a single-factor ANOVA to analyze the quantitative-zone-of-inhibition data. If an overall difference was found, all pairwise comparisons were made using the two-tailed t-test. Since there were 36 pairwise comparisons [9 concentrations × (9 − 1)/2 = 36] at each of the two temperatures, a Bonferroni correction was applied to the typical p value of < 0.05 considered statistically significant. The corrected p value was 0.05/36 = 0.0013888.
4.3. Germule Suppression Assay
Sterile microscopic slides (24.5 × 76.2 mm) were layered with 600 µL of molten SDA (10%) at a temperature of 50 °C and premixed with 5 µL of Pd spores (1 × 107 spores/mL) and 5 µL of each of the indicated concentrations of propolis. Quadruplet slides were prepared for each of the propolis concentrations and were incubated in duplicate at 8 °C and 15 °C. The germination of the Pd spores was measured macroscopically by visualizing the white growth confluence of the Pd lawn, and microscopically by the Pd hyphal extension. Both macroscopic and microscopic images were acquired from slides on the 7th and 16th day of incubation. The microscopic images were taken using a DCM 130E digital camera for microscope (1.3 M pixels, CMOS chip) (AmScope, Irvine, CA, USA). The microscopic images were imported with the Scope Photo Software (AmScope, Irvine, CA, USA).
4.4. Analysis of Chemical Constituents
The chemical composition of propolis was determined using matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF-MS). The propolis sample consisted of 50% diluted propolis in methanol covered with 0.1020 M of α-cyano-4-hydroxy cinnamic acid (HCCA) in 1:4 (v/v) H2O/acetonitrile. A 1.0 µL diluted aliquot of the sample was first spotted on the plate and covered with 2.0 µL of matrix. The mass spectra were obtained using a bench-top Microflex MALDI-TOF MS from Bruker Daltonics® (Bremen, Germany) equipped with a pulsed nitrogen laser at 355 nm wavelength. The spectra were recorded from 40 to 2000 Da (positive mode), and from 100 to 400 Da (positive mode) using FlexControl 3.3 software (ion source 1: 19 kV; ion source 2: 15.5 kV; lens voltage: 9.45 kV; laser frequency: 60 Hz; pulsed ion extraction (PIE) delay: 120 ns). Other parameters are shown in Table 2. Mass gates of 400 m/z (positive mode) were set for all experiments. Individual mass spectra from each spot were acquired by averaging 350 laser shots. Data acquisition was set to automate, and the “random walk” movement was activated at 10 shots per raster during the sequence. The peak lists and intensities were calculated using the peak-picking centroid algorithm in FlexAnalysis 3.3 software.
Supplementary Materials
The following are available online at www.mdpi.com/2079-6382/7/1/2/s1, Table S1: Analysis of Variance (ANOVA) One-way (8 °C incubation temperature), Table S2: Analysis of Variance (ANOVA) One way (15 °C incubation temperature).
Acknowledgments
We are thankful to the United States Fish and Wildlife Service (Cheeptham and Lausen’s Grant #F15AS00188) for funding. Our sincere appreciation also goes to Cori Lausen of the Wildlife Conservation Society Canada and Mark Rakobowchuk of TRU for their time and assistance in editing this manuscript.
Author Contributions
N.C. conceived the idea to use propolis. S.G. and R.M.A. designed all the experiments under the close guidance and supervision of N.C. S.G. drafted the manuscript. R.M.A. conducted the experiments. N.C. secured funding. K.D. designed the experiments used to determine the chemical composition of propolis. Z.C.G. and R.M.K. conducted the experiments under the supervision of K.D. K.D. drafted the chemistry section of the manuscript. J.P.X. provided the P. destructans M3906-2 strain and contributed to the statistical analyses. N.C., S.G., J.P.X. and K.D. read and edited the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- United States Fish and Wildlife Service (USFWS). North American bat death toll exceeds 5.5 million from white-nose syndrome. In USFWS News Release; United States Fish and Wildlife Service (USFWS): Washington, DC, USA, 2012; pp. 1–2. [Google Scholar]
- United States Fish and Wildlife Service (USFWS). White Nose Sydromes: Where Is It Now? In USFWS News Release; United States Fish and Wildlife Service (USFWS): Washington, DC, USA, 2017.
- Verant, M.L.; Boyles, J.G.; Waldrep, W., Jr.; Wibbelt, G.; Blehert, D.S. Temperature-dependent growth of Geomyces destructans, the fungus that causes bat white-nose syndrome. PLoS ONE 2012, 7, e46280. [Google Scholar] [CrossRef] [PubMed]
- O’Donoghue, A.J.; Knudsen, G.M.; Beekman, C.; Perry, J.A.; Johnson, A.D.; DeRisi, J.L.; Craik, C.S.; Bennett, R.J. Destructin-1 is a collagen-degrading endopeptidase secreted by pseudogymnoascus destructans, the causative agent of white-nose syndrome. Proc. Natl. Acad. Sci. USA 2015, 112, 7478–7483. [Google Scholar] [CrossRef] [PubMed]
- Webb, P.I.; Speakman, J.R.; Racey, P.A. How hot is a hibernaculum? A review of the temperatures at which bats hibernate. Can. J. Zool. 1996, 74, 761–765. [Google Scholar]
- Davies, W.H. Hibernation: Ecology and physiological ecology. In Biology of Bats; Wimsatt, W.A., Ed.; Acedemic Press: New York, NY, USA; London, UK, 1970; Volume 1, pp. 265–300. [Google Scholar]
- Humphries, M.M.; Thomas, D.W.; Speakman, J.R. Climate-mediated energetic constraints on the distribution of hibernating mammals. Nature 2002, 418, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Speakman, J.R.; Thomas, D.W. Physiological ecology and enegetics of bats. In Bat Ecology; Kunz, T.H., Fenton, M.B., Eds.; The University of Chicago Press: Chicago, IL, USA; London, UK, 2003; pp. 431–434. [Google Scholar]
- Warnecke, L.; Turner, J.M.; Bollinger, T.K.; Misra, V.; Cryan, P.M.; Blehert, D.S.; Wibbelt, G.; Willis, C.K. Pathophysiology of white-nose syndrome in bats: A mechanistic model linking wing damage to mortality. Biol. Lett. 2013, 9, 20130177. [Google Scholar] [CrossRef] [PubMed]
- Cryan, P.M.; Meteyer, C.U.; Boyles, J.G.; Blehert, D.S. Wing pathology of white-nose syndrome in bats suggests life-threatening disruption of physiology. BMC Biol. 2010, 8, 135. [Google Scholar] [CrossRef] [PubMed]
- Boyles, J.G.; Cryan, P.M.; McCracken, G.F.; Kunz, T.H. Conservation. Economic importance of bats in agriculture. Science 2011, 332, 41–42. [Google Scholar] [CrossRef] [PubMed]
- Kunz, T.H.; Braun de Torrez, E.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Cornelison, C.T.; Keel, M.K.; Gabriel, K.T.; Barlament, C.K.; Tucker, T.A.; Pierce, G.E.; Crow, S.A. A preliminary report on the contact-independent antagonism of Pseudogymnoascus destructans by Rhodococcus rhodochrous strain DAP96253. BMC Microbiol. 2014, 14, 246. [Google Scholar] [CrossRef] [PubMed]
- Hoyt, J.R.; Cheng, T.L.; Langwig, K.E.; Hee, M.M.; Frick, W.F.; Kilpatrick, A.M. Bacteria isolated from bats inhibit the growth of Pseudogymnoascus destructans, the causative agent of white-nose syndrome. PLoS ONE 2015, 10, e0121329. [Google Scholar] [CrossRef] [PubMed]
- Boire, N.; Zhang, S.; Khuvis, J.; Lee, R.; Rivers, J.; Crandall, P.; Keel, M.K.; Parrish, N. Potent inhibition of Pseudogymnoascus destructans, the causative agent of white-nose syndrome in bats, by cold-pressed, terpeneless, valencia orange oil. PLoS ONE 2016, 11, e0148473. [Google Scholar] [CrossRef] [PubMed]
- Bankova, V.S.; de Castro, S.L.; Marcucci, M.C. Propolis: Recent advances in chemistry and plant origin. Apidologie 2000, 31, 3–15. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, C.P.; Wang, K.; Li, G.Q.; Hu, F.L. Recent advances in the chemical composition of propolis. Molecules 2014, 19, 19610–19632. [Google Scholar] [CrossRef] [PubMed]
- Choudhari, M.K.; Punekar, S.A.; Ranade, R.V.; Paknikar, K.M. Antimicrobial activity of stingless bee (Trigona sp.) propolis used in the folk medicine of western Maharashtra, India. J. Ethnopharmacol. 2012, 141, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Khacha-ananda, S.; Tragoolpua, K.; Chantawannakul, P.; Tragoolpua, Y. Propolis extracts from the northern region of Thailand suppress cancer cell growth through induction of apoptosis pathways. Investig. New Drugs 2016, 34, 707–722. [Google Scholar] [CrossRef] [PubMed]
- Wagh, V.D. Propolis: A wonder bees product and its pharmacological potentials. Adv. Pharmacol. Sci. 2013, 2013, 308249. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Hepburn, H.R.; Li, Y.; Chen, M.; Radloff, S.E.; Daya, S. Effects of ethanol and water extracts of propolis (bee glue) on acute inflammatory animal models. J. Ethnopharmacol. 2005, 100, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Ping, S.; Huang, S.; Hu, L.; Xuan, H.Z.; Zhang, C.P.; Hu, F.L. Molecular mechanisms underlying the in vitro anti-inflammatory effects of a Ffavonoid-rich ethanol extract from Chinese propolis (poplar type). Evid. Based Complement. Altern. Med. 2013, 2013, 127672. [Google Scholar]
- Xuan, H.; Zhao, J.; Miao, J.; Li, Y.; Chu, Y.; Hu, F. Effect of Brazilian propolis on human umbilical vein endothelial cell apoptosis. Food Chem. Toxicol. 2011, 49, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Xuan, H.; Zhu, R.; Li, Y.; Hu, F. Inhibitory effect of chinese propolis on phosphatidylcholine-specific phospholipase C activity in vascular endothelial cells. Evid. Based Complement. Altern. Med. 2011, 2011, 985278. [Google Scholar] [CrossRef] [PubMed]
- Ito, J.; Chang, F.R.; Wang, H.K.; Park, Y.K.; Ikegaki, M.; Kilgore, N.; Lee, K.H. Anti-AIDS agents. 48.(1) ANTI-HIV activity of moronic acid derivatives and the new melliferone-related triterpenoid isolated from Brazilian propolis. J. Nat. Prod. 2001, 64, 1278–1281. [Google Scholar] [CrossRef] [PubMed]
- Teerasripreecha, D.; Phuwapraisirisan, P.; Puthong, S.; Kimura, K.; Okuyama, M.; Mori, H.; Kimura, A.; Chanchao, C. In vitro antiproliferative/cytotoxic activity on cancer cell lines of a cardanol and a cardol enriched from Thai Apis mellifera propolis. BMC Complement. Altern. Med. 2012, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Silici, S.; Kutluca, S. Chemical composition and antibacterial activity of propolis collected by three different races of honeybees in the same region. J. Ethnopharmacol. 2005, 99, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Kujumgiev, A.; Tsvetkova, I.; Serkedjieva, Y.; Bankova, V.; Christov, R.; Popov, S. Antibacterial, antifungal and antiviral activity of propolis of different geographic origin. J. Ethnopharmacol. 1999, 64, 235–240. [Google Scholar] [CrossRef]
- Grange, J.M.; Davey, R.W. Antibacterial properties of propolis (bee glue). J. R. Soc. Med. 1990, 83, 159–160. [Google Scholar] [PubMed]
- Shimizu, T.; Takeshita, Y.; Takamori, Y.; Kai, H.; Sawamura, R.; Yoshida, H.; Watanabe, W.; Tsutsumi, A.; Park, Y.K.; Yasukawa, K.; et al. Efficacy of Brazilian propolis against herpes simplex virus type 1 infection in mice and their modes of antiherpetic efficacies. Evid. Based Complement. Altern. Med. 2011, 2011, 976196. [Google Scholar] [CrossRef] [PubMed]
- Silici, S.; Koc, A.N. Comparative study of in vitro methods to analyse the antifungal activity of propolis against yeasts isolated from patients with superficial mycoses. Lett. Appl. Microbiol. 2006, 43, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Amoros, M.; Simoes, C.M.; Girre, L.; Sauvager, F.; Cormier, M. Synergistic effect of flavones and flavonols against herpes simplex virus type 1 in cell culture. Comparison with the antiviral activity of propolis. J. Nat. Prod. 1992, 55, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Bueno-Silva, B.; Alencar, S.M.; Koo, H.; Ikegaki, M.; Silva, G.V.; Napimoga, M.H.; Rosalen, P.L. Anti-inflammatory and antimicrobial evaluation of neovestitol and vestitol isolated from Brazilian red propolis. J. Agric. Food Chem. 2013, 61, 4546–4550. [Google Scholar] [CrossRef] [PubMed]
- Salatino, A.; Fernandes-Silva, C.C.; Righi, A.A.; Salatino, M.L. Propolis research and the chemistry of plant products. Nat. Prod. Rep. 2011, 28, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Buckley, T.R.; Frati, F.; Stewart, J.B.; Beckenbach, A.T. Incorporating molecular evolution into phylogenetic analysis, and a new compilation of conserved polymerase chain reaction primers for animal mitochondrial DNA. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 545–579. [Google Scholar] [CrossRef]
- Fernandes-Silva, C.C.; Freitas, J.C.; Salatino, A.; Salatino, M.L. Cytotoxic activity of six samples of Brazilian propolis on sea urchin (Lytechinus variegatus) eggs. Evid. Based Complement. Altern. Med. 2013, 2013, 619361. [Google Scholar] [CrossRef] [PubMed]
- McArthur, R.L.; Ghosh, S.; Cheeptham, N. Improvement of protocols for the screening of biological control agents against white-nose syndrome. JEMI+ 2017, 2, 1–7. [Google Scholar]
- Langwig, K.E.; Frick, W.F.; Hoyt, J.R.; Parise, K.L.; Drees, K.P.; Kunz, T.H.; Foster, J.T.; Kilpatrick, A.M. Drivers of variation in species impacts for a multi-host fungal disease of bats. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef] [PubMed]
- Cornelison, C.T.; Gabriel, K.T.; Barlament, C.; Crow, S.A., Jr. Inhibition of Pseudogymnoascus destructans growth from conidia and mycelial extension by bacterially produced volatile organic compounds. Mycopathologia 2014, 177, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Christov, R.; Trusheva, B.; Popova, M.; Bankova, V.; Bertrand, M. Chemical composition of propolis from Canada, its antiradical activity and plant origin. Nat. Prod. Res. 2006, 20, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Bankova, V. Recent trends and important developments in propolis research. Evid. Based Complement. Altern. Med. 2005, 2, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Burdock, G.A. Review of the biological properties and toxicity of bee propolis (propolis). Food Chem. Toxicol. 1998, 36, 347–363. [Google Scholar] [CrossRef]
- Bankova, V.; Popova, M.; Trusheva, B. Propolis volatile compounds: Chemical diversity and biological activity: A review. Chem. Cent. J. 2014, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Trusheva, B.; Todorov, I.; Ninova, M.; Najdenski, H.; Daneshmand, A.; Bankova, V. Antibacterial mono- and sesquiterpene esters of benzoic acids from iranian propolis. Chem. Cent. J. 2010, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Uzel, A.; Sorkun, K.; Oncag, O.; Cogulu, D.; Gencay, O.; Salih, B. Chemical compositions and antimicrobial activities of four different Anatolian propolis samples. Microbiol. Res. 2005, 160, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Gomes, B.P.; Rosalen, P.L.; Ambrosano, G.M.; Park, Y.K.; Cury, J.A. In vitro antimicrobial activity of propolis and Arnica montana against oral pathogens. Arch. Oral Biol. 2000, 45, 141–148. [Google Scholar] [CrossRef]
- Santos, F.A.; Bastos, E.M.; Rodrigues, P.H.; de Uzeda, M.; de Carvalho, M.A.; Farias Lde, M.; Moreira, E.S. Susceptibility of Prevotella intermedia/Prevotella nigrescens (and Porphyromonas gingivalis) to propolis (bee glue) and other antimicrobial agents. Anaerobe 2002, 8, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Abu-Seida, A.M. Effect of propolis on experimental cutaneous wound healing in dogs. Vet. Med. Int. 2015, 2015, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Khankhet, J.; Vanderwolf, K.J.; McAlpine, D.F.; McBurney, S.; Overy, D.P.; Slavic, D.; Xu, J. Clonal expansion of the Pseudogymnoascus destructans genotype in North America is accompanied by significant variation in phenotypic expression. PLoS ONE 2014, 9, e104684. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Anti-Pd activity of propolis. (A) Images (i–xi) and (xii–xxii) indicate the activity of propolis at 8 °C and 15 °C, respectively. The black arrowheads indicate the zone of inhibition of Pd when treated with different concentrations of propolis in comparison to water and anhydrous ethanol treatments; (B) diameter of the zones of inhibition at 8 °C and 15 °C. The error bars are standard deviations of the diameters.
Figure 1.
Anti-Pd activity of propolis. (A) Images (i–xi) and (xii–xxii) indicate the activity of propolis at 8 °C and 15 °C, respectively. The black arrowheads indicate the zone of inhibition of Pd when treated with different concentrations of propolis in comparison to water and anhydrous ethanol treatments; (B) diameter of the zones of inhibition at 8 °C and 15 °C. The error bars are standard deviations of the diameters.
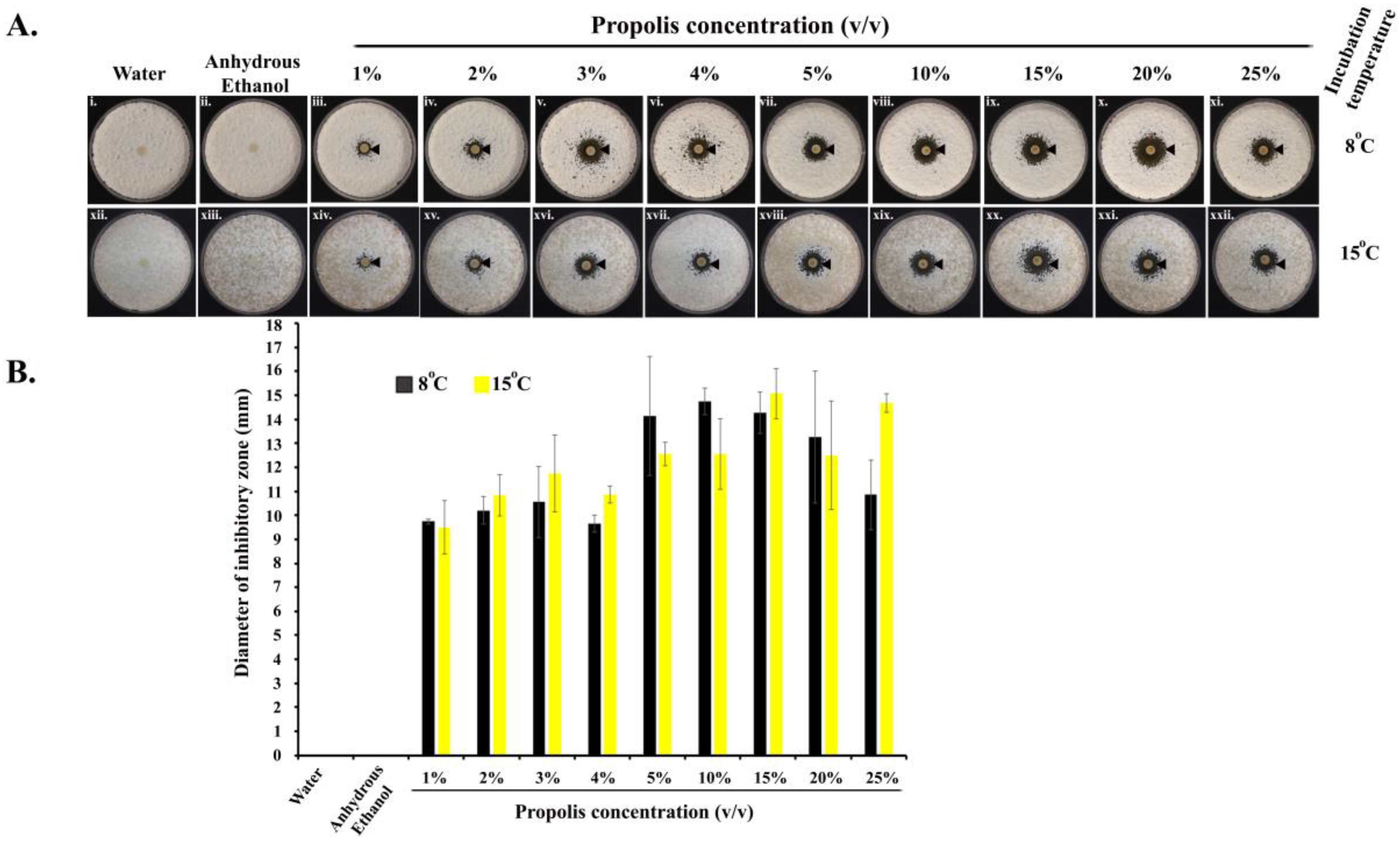
Figure 2.
Germule suppression assay. (A,B) represent the Pd germination assay for treatments with water, anhydrous ethanol, and various concentrations of propolis at 8 °C and 15 °C, respectively. The white arrowheads indicate the mycelial extension of the Pd spores at two different incubation temperatures. The black arrowheads indicate the inhibition of the Pd spores on exposure to propolis at different concentrations. The green arrowheads indicate the formation of white Pd lawns resulting from the treatment of spores with water or anhydrous ethanol.
Figure 2.
Germule suppression assay. (A,B) represent the Pd germination assay for treatments with water, anhydrous ethanol, and various concentrations of propolis at 8 °C and 15 °C, respectively. The white arrowheads indicate the mycelial extension of the Pd spores at two different incubation temperatures. The black arrowheads indicate the inhibition of the Pd spores on exposure to propolis at different concentrations. The green arrowheads indicate the formation of white Pd lawns resulting from the treatment of spores with water or anhydrous ethanol.
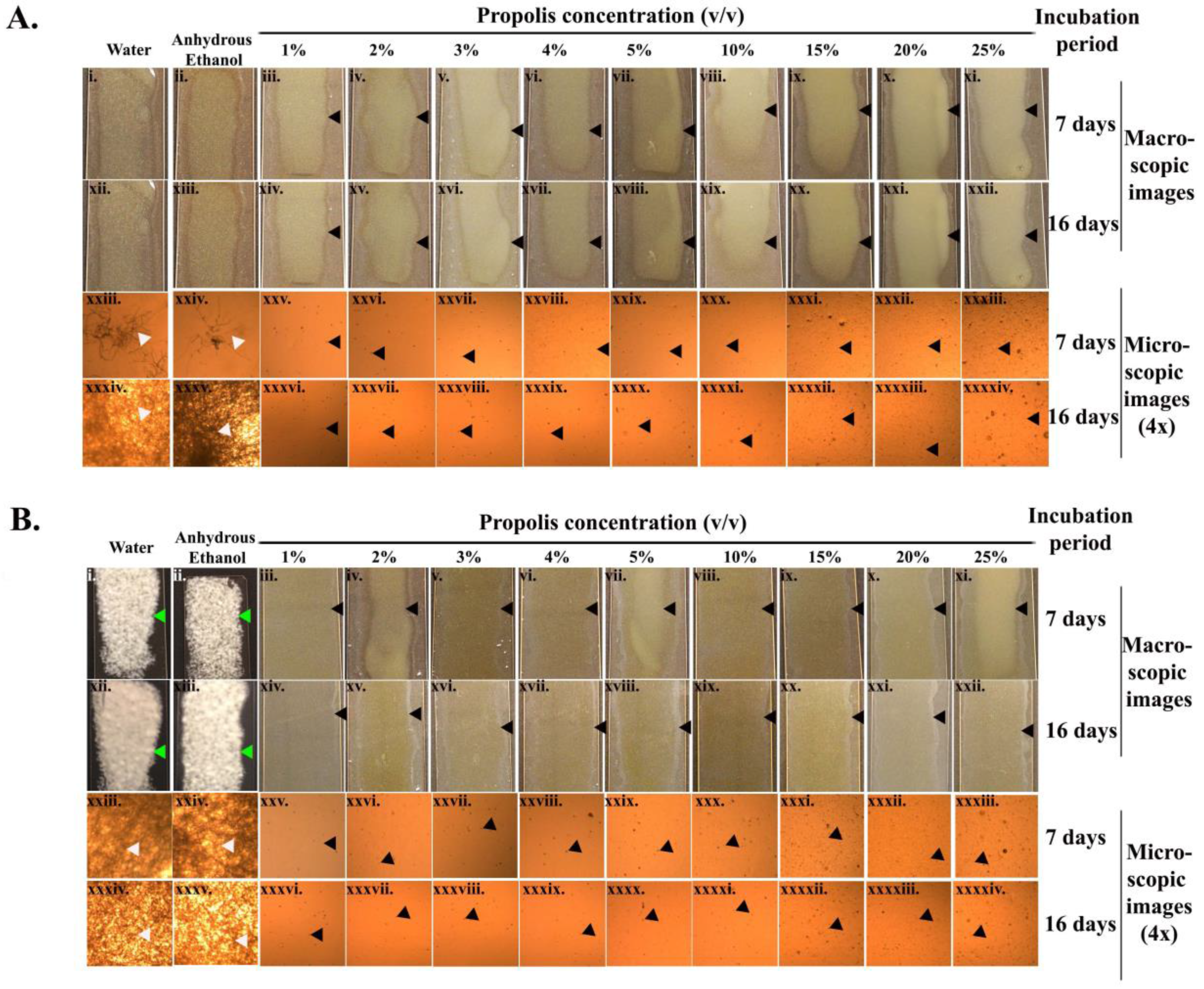
Figure 3.
Micrographs of Pd spores displayed at 10× and 40× magnification: (i–ii) elliptical shape of untreated Pd spores; (iii–iv) deformed Pd spores treated with 1% propolis.
Figure 3.
Micrographs of Pd spores displayed at 10× and 40× magnification: (i–ii) elliptical shape of untreated Pd spores; (iii–iv) deformed Pd spores treated with 1% propolis.

Figure 4.
(A) MALDI-TOF-MS of a propolis sample at a mass range of 100–400 Da. Each of the peaks on the mass spectrum represents a distinctive compound in our propolis sample. The numbers above the peaks correspond to the compounds listed in Table 1; (B) magnified version of the mass spectrum at a mass range of the 100–225 Da.
Figure 4.
(A) MALDI-TOF-MS of a propolis sample at a mass range of 100–400 Da. Each of the peaks on the mass spectrum represents a distinctive compound in our propolis sample. The numbers above the peaks correspond to the compounds listed in Table 1; (B) magnified version of the mass spectrum at a mass range of the 100–225 Da.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Composition of propolis as determined by MALDI-TOF-MS (the peaks corresponding to these values can be seen in the mass spectrum in Figure 4).
Table 1.
Composition of propolis as determined by MALDI-TOF-MS (the peaks corresponding to these values can be seen in the mass spectrum in Figure 4).
| Peak | Constituents Identified | Mass/Charge (m/z) | Intensity |
|---|---|---|---|
| 1 | Benzyl alcohol | 108.14 | 0.77 |
| 2 | Hydroquinone | 110.11 | 5.00 |
| 3 | Benzoic acid | 122.12 | 5.93 |
| 4 | Cinnamyl alcohol | 134.17 | 23.81 |
| 5 | Hydroxyacetophenone | 136.15 | 7.82 |
| 6 | 4-Hydroxybenzoic acid | 138.12 | 10.98 |
| 7 | Cinnamic acid | 148.16 | 14.81 |
| 8 | p-coumaric acid | 164.16 | 4.91 |
| 9 | 3-Phenyl-3-hydroxypropanoic acid | 166.18 | 5.91 |
| 10 | Sesquiterpenes | 168.31 | 13.90 |
| 11 | Ferulic acid | 194.18 | 3.91 |
| 12 | Benzyl benzoate | 212.25 | 6.85 |
| 13 | Benzyl methoxybenzoate | 242.27 | 211.88 |
| 14 | Benzyl dihydroxybenzoate | 244.24 | 105.16 |
| 15 | Palmitic acid | 256.43 | 64.16 |
| 16 | 2,4,6-Trihydroxydihydrochalcone | 258.27 | 5184.70 |
| 17 | Pinostrobin chalcone | 270.28 | 47.98 |
| 18 | 2,6-Dihydroxy-4-methoxydihydrochalcone | 272.25 | 132.69 |
| 19 | Oleic acid | 282.47 | 28.64 |
| 20 | Stearic acid | 284.31 | 50.69 |
| 21 | Sakuranetin | 286.27 | 44.71 |
| 22 | 2,4,6-Trihydroxy-4-methoxydihydrochalcone | 288.30 | 247.59 |
| 23 | Cinnamyl caffeate | 296.32 | 44.87 |
| 24 | Pinobanksin 3-O-acetate | 314.29 | 2331.83 |
Table 2.
Parameters of MALDI-TOF-MS.
| Parameters | Values |
|---|---|
| Laser | Pulsed nitrogen |
| Laser power | 20–80% |
| Peak selection (mass range) | 40–2000 Da |
| Sample rate | 0.05 GS/s |
| Mass range | Low range |
| Electronic gain | Enhanced 100 mV |
| Realtime smooth | Off |
| Spectrum size | 2069 pts |
| Spectrum delay | 307 pts |
| Laser frequency | 60.0 Hz |
| Laser attenuator offset | 17% |
| Laser attenuator range | 30% |
| Target | MSP 96 target polished steel |
| Matrix | α-cyano-4-hydroxy-cinnamic acid, HCCA |
| Sample | 50% diluted propolis in MeOH covered with 0.1020 M of HCCA in 1:4 (v/v) H2O/acetonitrile |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ghosh, S.; McArthur, R.; Guo, Z.C.; McKerchar, R.; Donkor, K.; Xu, J.; Cheeptham, N. Evidence for Anti-Pseudogymnoascus destructans (Pd) Activity of Propolis. Antibiotics 2018, 7, 2. https://doi.org/10.3390/antibiotics7010002
AMA Style
Ghosh S, McArthur R, Guo ZC, McKerchar R, Donkor K, Xu J, Cheeptham N. Evidence for Anti-Pseudogymnoascus destructans (Pd) Activity of Propolis. Antibiotics. 2018; 7(1):2. https://doi.org/10.3390/antibiotics7010002
Chicago/Turabian StyleGhosh, Soumya, Robyn McArthur, Zhi Chao Guo, Rory McKerchar, Kingsley Donkor, Jianping Xu, and Naowarat Cheeptham. 2018. "Evidence for Anti-Pseudogymnoascus destructans (Pd) Activity of Propolis" Antibiotics 7, no. 1: 2. https://doi.org/10.3390/antibiotics7010002
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.