An Early History of Neuroglial Research: Personalities



1
Scimed Biotechnologies, s.r.o., 25241 Zlatníky-Hodkovice, Czech Republic
2
Institute of Pharmacology and Toxicology, Faculty of Medicine in Plzeň, Charles University, 30605 Plzeň, Czech Republic
3
Faculty of Biology, Medicine and Health, The University of Manchester, M13 9PT Manchester, UK
4
Achucarro Center for Neuroscience, IKERBASQUE, Basque Foundation for Science, 48011 Bilbao, Spain
5
Center for Basic and Translational Neuroscience, Faculty of Health and Medical Sciences, University of Copenhagen, 2200 Copenhagen, Denmark
*
Authors to whom correspondence should be addressed.
Neuroglia 2018, 1(1), 245-257; https://doi.org/10.3390/neuroglia1010016
Submission received: 16 July 2018
/
Revised: 31 July 2018
/
Accepted: 6 August 2018
/
Published: 16 August 2018
Abstract
:Neuroscience, like most other divisions of natural philosophy, emerged in the Hellenistic world following the first experimental discoveries of the nerves connecting the brain with the body. The first fundamental doctrine on brain function highlighted the role for a specific substance, pneuma, which appeared as a substrate for brain function and, being transported through the hollow nerves, operated the peripheral organs. A paradigm shift occurred in 17th century when brain function was relocated to the grey matter. Beginning from the end of the 18th century, the existence of active and passive portions of the nervous tissue were postulated. The passive part of the nervous tissue has been further conceptualised by Rudolf Virchow, who introduced the notion of neuroglia as a connective tissue of the brain and the spinal cord. During the second half of the 19th century, the cellular architecture of the brain was been extensively studied, which led to an in-depth morphological characterisation of multiple cell types, including a detailed description of the neuroglia. Here, we present the views and discoveries of the main personalities of early neuroglial research.
1. Brief History of Neuroscience: From Ventricular-Pneumatic Doctrine to Cellular Structure
In the most ancient times, natural philosophy associated human intelligence with the heart and the cardiovascular system. This cardiocentric doctrine was predominant in ancient Egypt, in Judea, and in Persia; it was popular in the Hellenic world, and it remained a mainstream concept in Oriental medicine (most notably in China and in Japan) until the middle of 19th century [1]. In ancient Greece, Aristotle (384–322 B.C.) was the prominent proponent of the cardiovascular doctrine; Aristotle introduced the concept of pneuma, an air-like substance that was produced in the heart and that was distributed through the body. This pneuma, according to Aristotle, was a substrate of intelligence and cognition, and when released from the blood it triggered peripheral voluntary reactions. Within this paradigm, the brain was a bloodless organ that served for cooling pneuma and for moderate passions [2,3].
The first natural philosopher, anatomist, and physiologist who linked the brain with higher cognitive functions was Alcmaeon of Croton (6th–5th centuries B.C.). Alcmaeon defined intelligence as the main factor distinguishing between humans and other animals: “Man differs from other animals, because he alone comprehends, while the other animals perceive but do not understand, because understanding and perception are different things”, and realised that “all the senses are connected somehow with the brain” [4]. The brain as an organ of intelligence was recognised by Plato, by Socrates, and in the writings of the Hippocratic school. The Hippocratic corpus (~60 medical texts written by the members of Hippocrates’ school in the 5th–4th centuries B.C.) contains a chapter entitled “On the sacred disease”, which identifies the brain as an organ of cognition.
“It ought to be generally known that the source of our pleasure, merriment, laughter, and amusement, as of our grief, pain, anxiety, and tears, is none other than the brain. It is specially the organ which enables us to think, see, and hear, and to distinguish the ugly and the beautiful, the bad and the good, pleasant and unpleasant...” [5].
Systematic neuroanatomy was introduced by Herophilus of Chalcedon (335–280 B.C.) and Erasistratus of Cos (304–250 B.C.) who both worked in the Museum of Alexandria (Figure 1A,B). Herophilus identified nerves (including cranial nerves) and found that nerves connect the body with the brain. He further discovered the existence of motor and sensory nerves, and realised that motor nerves originate either in the brain or in the spinal cord. Herophilus was also the first to describe brain ventricles. Erasistratus continued the neuroanatomical studies of his predecessor, and was the first to found the direct relations between complexity of the brain and human intelligence [1,6,7,8]. Erasistratus was also the first to outline the principles of the ventricular-pneumatic theory of brain function. According to this theory, there are several classes of pneuma; the first type, the vital pneuma, is produced in the lungs from the inhaled air; this vital pneuma is subsequently distributed throughout the body by blood vessels, the vital pneuma diffuses to the tissues and sustains their life. The brain parenchyma converts the vital pneuma into the psychic pneuma, which is concentrated in the ventricles. The psychic pneuma represents the substrate of nervous function, including cognition. From the ventricles, the psychic pneuma is sent to the periphery through the hollow nerves; when released from nerve terminal, the psychic pneuma instigates peripheral responses [1,6,7,8]. Of note, Democritus (460–370 B.C.) suggested that the “psyche”, that is, the substrate of nerve function, is formed by the lightest atoms [3].
The ventricular-pneumatic theory of the brain function was further developed by Claudius Galen of Pergamon (129–200 A.D., Figure 1C). Galen regarded the pneuma as a special fluid that was produced by the brain parenchyma from the “vital spirit” produced by the lungs from the air; the final conversion of vital spirit to pneuma occurred in the choroid plexus, through which pneuma entered the ventricular system. The ventricles worked as a pump, which maintained the movement of pneuma through the motor nerves, and the aspiration of pneuma from the sensory nerves. The pneuma, which completely filled the nerves, made them rigid, which allowed the rapid conduction of signals similar to the pulse wave propagation through blood vessels. Signalling at the sensory nerve endings and motor nerve terminals occurred through microscopic pores that allowed free exchange of pneuma between the nerves and peripheral tissues. At the brain level, pneuma carrying sensory signals is delivered to the anterior ventricles, whereas the afferent signals to the muscles originate from the posterior ventricle [9]. Based on experiments on live animals which involved selective compression or lesioning of different parts of the brain and nerve ligation, Galen contemplated distinct localisation of functions within different anatomical structures. First, he demonstrated that damage to the brain parenchyma per se did not cause major deficits unless the ventricle was opened (apparently causing the escape of the pneuma). He also found that compression of anterior ventricle led to blindness, whereas compression of the posterior ventricle resulted in general paralysis. The damage to the ventricles resulted either in serious sensory deficits (anterior ventricle), or in collapse and death (middle and posterior ventricles).
The ventricular-pneumatic doctrine remained the main theory of neuroscience throughout the Middle Ages and the Renaissance. Anatomists and medics from both western world (Albertus Magnus, Tomas Aquinas, Roger Bacon, Petrus de Montagnana, Lodovico Dolce, Ghiradelli of Bologna, and Theodor Gull of Antwerp) and from Arabic countries (Ibn Sīnā and Ibn Rushd, known in Europe as Avicenna and Averroes), added further developments by assigning different functions to different parts of ventricular system (Figure 2). The anterior ventricle was usually associated with sensory input and sensory processing, the middle ventricle was linked to creativity and imagination, cognition and intellect, whereas the posterior ventricle was a seat for memory [9]. Leonardo da Vinci, who was an adept of the ventricular-pneumatic doctrine, made the first three-dimensional cast of the ventricles by filling them with melted wax (Figure 2C); when the wax was set, the brain tissues were removed, thus revealing the ventricular system [10]. According to Leonardo’s reckoning the anterior ventricles were responsible for bridging the sensory inputs with senso commune, while memory resided in the posterior ventricle.
Rene Descartes (1596–1650), who introduced the concept of reflexes, had his own view on the functioning of the nervous system. He assigned the nervous function to small corpuscles or “a very fine flame” which was released from nerve endings [14]. According to Descartes, peripheral stimulation triggered mechanical displacement of nerves that almost immediately caused the central end of the nerve to twitch. This in turn triggered the release of “animal spirit” or “a very fine flame” that in the form of minute particles, flew to the ventricles. The same particles carried signals through the nerves to the periphery. In the nerve endings, these signalling particles were released through miniature valves and diffused to the muscles, where they were taken up by congruent valves; after entering the muscles, these said particles initiated and regulated contraction. Descartes also suggested that volatile movements originate from the pineal gland surrounded by the cerebrospinal fluid. Small movements of the pineal gland are accordingly able to regulate the flow of the spiritus animales through a complex system of pipes and valves (Figure 3A).
The first departures form the ventricular-pneumatic views emerged in the 17th century when Marchello Malpighi and, to a larger extend Thomas Willis, noted the roles for the grey matter and brain parenchyma in nervous function [15,16]. Localisation of functions in the brain was initially performed by numerous phrenologists, including Georg Prochaska, Franz Joseph Gall, Johann Gaspard Spurzheim, and George Combe [17,18,19,20]. The first true functional brain centre was discovered by Paul Broca, who identified the area in the posterior-inferior part of the frontal cortex of the dominant hemisphere as a centre of speech [21]. Further advances in the localisation of brain functions were made by Gustav Theodor Fritsch and Edward Hitzig [22], and by systematic studies of Sir David Ferrier, who produced the first map of functional speciality of various brain regions in monkeys, including the motor and sensory (vision, heating and taste) areas [23,24,25]. The electrophysiological mapping of the motor cortex was finalised by Wilder G. Penfield who invented the concept of the motor and sensory “homunculus” [26].
These advances in neuroanatomy coincided with the emergence of neurophysiology. Probably the first true neurophysiologist was Albrecht von Haller, who performed physiological experiments and developed the theory of irritability and sensitivity [31,32]. By systematic injury to all organs and parts of organs by spraying with cold, hot or corrosive substances, by stinging, squeezing, and also by means of electricity, Haller, in observing the stimulus response of the organ part concerned, classified the two categories of irritable (irritabilis) and sensible (sensibilis) tissues [33]. Haller’s stimulation experiments, based on the assignment of the criteria irritable and sensitive, led for the first time to a precise, functional differentiation of the smallest components (fibres) of animal organisms (Figure 3B). Haller also noticed that in addition to the tissues that were directly responsible for irritation and sensitivity, there was another type of tissue which belonged to neither group. Haller called the third type of tissue the “cell tissue fibre” (Zellgewebsfaser), constituting “cellular tissue” (Zellgewebe). According to him, this was an inert tissue forming a filling or a basic substance, that encompassed all those components of the organism that Haller was unable to identify as either sensitive or irritable [34].
The ideas of cellular nature of the nervous tissue were developed in parallel. Starting from the mid-17th century, the notion of elementary units of life were contemplated by several natural philosophers, most notable by Pierre Gassendi and Robert Boyle. This coincided with the emergence of first microscopes that allowed visualisation of life units. The term “cell” was introduced in 1665 by Robert Hooke [35], who noted the semblance between regular microstructures that he observed in cork, with the monk’s cells in the monastery dormitories. The first animal cells were visualised and described by Antonie van Leeuwenhoek (1632–1723), who, by using self-made microscopes, found bacteria (he called them animalcules or little animals); he also observed erythrocytes, single muscle fibres, and followed the movements of live spermatozoids [36]. The cellular theory of life was formalised by Theodor Schwann and Matthias Jakob Schleiden [37,38].
In particular, Theodor Schwann (1810–1882), in his famous book devoted to the cell theory [38], summarized the properties of tissues as follows:
Upon these more or less important modifications of the cell-life the following classification of the tissues is based: 1st. Isolated, independent cells, which either exist in fluids, or merely lie unconnected and moveable, beside each other. 2d. Independent cells applied firmly together, so as to form a coherent tissue. 3d. Tissues, in which the cell-walls (but not the cell-cavities) have coalesced together, or with the intercellular substance. Lastly, tissues in which both the walls and cavities of many cells blend together. In addition to these, however, there is yet another very natural section of the tissues, namely, the fibre-cells, in which independent cells are extended out on one or more sides into bundles of fibres. The naturalness of this group will form my excuse for sacrificing logical classification to it, and inserting it as the fourth class (4th), consequently, that last mentioned, consisting of tissues, in which the cell-walls and cell-cavities coalesce, becomes the fifth (5th).(Cited from English translation; [39] p. 65)
Thus, according to Schwann, all tissues, including nervous and connective tissues, were composed from cells or their constituents; accordingly, all fibres were parts of the cells and they were cellular processes.
Probably the first microscopic observations of the cortex were made by Marcello Malpighi, who noted that nervous tissue is composed from numerous little formations, which he called “globules” or “little glands” ([40] and Figure 3C); similar structures were mentioned by other contemporary microscopists including Antonie van Leeuwenhoek (Figure 3D and [41]). Whether these structures indeed represented neural cells remains unknown. The very first true description of neural cells belongs to Emanuel Swedenborg (1688–1772) who described (in the 1740s) that the “globules” or “cerebellulas” (small brains) were independent functional entities that were interconnected with fibres; these globules and their projected fibres were the substrates for brain function: “From each cortical gland proceeds a single nerve fibre; this is carried down into the body, in order that it may take hold of some part of a sensation or produce some action” (quoted from [41]). Swedenborg’s writings unfortunately were not published until 1882 [42] and hence they did not influence the developments of cellular neuroscience. The globular structures of different sizes (which sizes vary between regions of the nervous system) have been also described by Giovanni Maria Della Torre (Figure 4A and [43]).
Peripheral nerves were first observed under microscope by van Leeuwenhoek, who described (in 1717, see [44]) small regular circular structures (that represented single axons) in the sagittal slice of the nerves isolated from cows or sheep (Figure 3D). Felice Gaspar Ferdinand Fontana produced, in 1780, the very first anatomical description of nerve and neural fibres or axons, noting that the nerve is composed “of a large number of transparent, uniform, and simple cylinders. These cylinders seem to be fashioned like a very thin, uniform wall or tunic which is filled, as far as one can see, with transparent, gelatinous fluid insoluble in water” [44].
One of the earliest description of cellular elements of the central nervous system have been made by Henri Dutrochet (1776–1847), who observed clusters of globular particles (corpuscules globuleux agglomeres) in the frog brain, which he suggested were the structural elements of the nervous tissue [46,47]. Dutrochet further concluded that these globular structures (nerve bodies) were “manufacturers of nervous energy”, while nerve fibres provided “nervous motion”. He further studied cerebral ganglia of molluscs Helix pomatia and Arion rufus, and reported clusters of spherical cells that were associated with smaller spherical or ovoid particles (Figure 4B). A century later, František Karel Studnička speculated that Dutrochet described large ganglion neurones surrounded by glial cells, which can be further identified as microglia, and thus Dutrochet was the first discoverer of neuroglia [48]. This inspiring idea however, remains a mere surmise Dutrochet by himself was oblivious to the existence of supportive cells of the nervous system.
The first documented microscopic images of leech neurones (Figure 4C) were obtained by Christian Gottfried Ehrenberg [49], while Jan Evangelista Purkyňe identified and imaged nerve cells of the cerebellum [50], and his pupil Gustav Gabriel Valentin observed neurones with nuclei (Figure 5A,B). In those days, the nerve cells were generally called globules or kugeln (spheres in German). Robert Bently Todd was the first to name these structures cells; he wrote: “The essential elements of the grey nervous matter are “vesicles” or cells, containing nuclei and nucleoli”. They have also been called nerve or ganglion “globules” [51]. In 1873, Camillo Golgi introduced the silver-chromate staining technique [52], which allowed much more detailed microscopic visualisation of neural cells.
2. The Concept of Neuroglia. The Birth of the Concept
Albrecht von Haller, in his treatise on irritability and sensitivity (see above) also noted that besides tissues involved in irritation and sensitivity, another type of tissue exists which belong to neither group and which Haller called as “cell tissue fibre” (Zellgewebsfaser), constituting “cellular tissue” (Zellgewebe). According to him, this was an inert tissue forming a filling or basic substance that encompassed all those components of the organism that Haller was unable to identify as either sensitive or irritable [33]. Haller, most likely for the first time, defined fibre tissue as an inert tissue forming a filling or basic substance in the brain tissue.
Almost 60 years after Haller’s work, German anatomist Theodor Keuffel (no personal data are available), who was a student of famous German physician, physiologist, anatomist, and psychiatrist Johann Christian Reil (1759–1813), published a book, based on his PhD thesis, which was dedicated to the histological examination of the central nervous system [45,54]. In the spinal cord slices, he described fine-needled fabric of the finest fibres; the whole fibrous substance was of reticular shape, as it would correspond to a fibrous network consisting of an innumerable amount of finest fibrils, which formed anastomoses between themselves in the most varied ways, and to connect with each other by countless branches (Figure 4D). The fibrous matter in the white, as well as in the grey matter, expressed a completely uniformly net-like structure without any predominant direction. The fibrils of the white and grey matter, merged directly with each other, so that it seemed to Keuffel that the fibrillary mesh ran through the entire cross-section of the spinal cord. The nature of the fibrous substance, according to him, was a compressed animal substance. What he observed, was probably the vascular network of the spinal cord, since by using his methods, all neuroglial cells disappeared and only vessels and the actual connective tissue remained (see also [55]). Thus, the main contribution of Keuffel was the concept of a histological microstructure that existed in the nervous tissue in addition to the nervous elements, which led to the conviction that the brain and spinal cord tissue also contained some sort of fibrillary tissue which was observed in the other organs of the body.
Gabriel Valentin (1810–1883) was a German physiologist; after finishing in the University of Breslau (now Wroclaw in Poland), he worked as an assistant to Jan Evangelista Purkyně. Valentin published results of his investigation of the structure if the nervous tissue, which was done most likely under the influence of Purkyně, in an extensive treatise [56,57]. According to Valentin:
The whole nervous system consists of two primary masses, namely the isolated spheres of the occupying masses and the isolated, continuous primitive fibres. The former are probably representatives of the creative, active, higher principle, the latter of the receiving and guiding, passive, lower principle. Each of these is enveloped by a cell-tissue sheath, the strength of which determines the intensity of the action of both heterogeneous parts on each other. These are the pure and peculiar formations of the nervous system.([56]; p. 157)
In Valentin’s model, the nervous tissue comprised spheres (Kugeln der Belegungsmassen), which represent cell bodies, primitive fibres (Primitivfasern), which represent nerve fibres, and intermediate substance (Zellgewebescheide) containing fibres or threads, which was most likely identical to what some authors later called as glial sheaths (Gliascheiden). In contrast to Keuffel, who considered this substance to be fibrous only, Valentin clearly identified it as cellular tissue (Figure 5A). He also described that the sheaths were always of a cellular nature where the cellular elements were interconnected by threadlike cellular tissue extensions. Valentin proposed that this whole formation evidently had the purpose of inhibiting the mutual direct action of the primitive fibres and the spheres, since he was convinced, on the basis of physiological experiments, that the sheaths had the properties of an insulator. The main contribution of Valentin was therefore the establishment of the concept of the nerve tissue structure, which moved towards the later neuroglial concept in two ways: first, the intermediate filling substance, other than nerve bodies and fibres, had a cellular structure with thread-like extensions, and second, this intermediate filling substance had insulating properties.
Purkyně’ was the first to provide topographic description of the ganglionic bodies, which from today’s perspective, corresponds to the agglomerations of various classes of nerve cells in different areas of the brain, spinal cord, and ganglia, and in which he described nuclei and nucleoli. Among them were also Purkinje cells, large neurons with many branching dendrites found in the cerebellum (Figure 5B). However, lesser known in fact, was that Purkyně has also a priority in the description of the processes of cells in the brain and spinal cord that were in close relationship with nearby blood vessels. As he pointed out in the extended and less known version of his abstract from the Prague meeting in 1837:
The processes of the ganglionic cells in the brain and in the spinal cord sometimes appear to be related to the large number of surrounding blood vessels, but it has never been proven with certainty, even less could be determined here about the reduction of very fine brain fibres [58].
What Purkyně most likely observed were the processes of perivascular astrocytes, which are closely associated with the brain capillaries.
The general view on the structure of the nervous system was significantly, and for a long period of time, affected by the concept suggested by the German professor emeritus of anatomy and physiology at Heidelberg, Friedrich Arnold (1803–1890). In his handbook of human anatomy [59] he stated, that the base substance (Grundmasse), the germinal matter, or the primary substance of the cellular tissue (Zellgewebe), is recognized, with the aid of good instruments and with moderate moderation of light, as a dull, finely granulated, molecular substance (moleculare Substanz) between the threads and bundles of the connective tissue. Concerning the nerve substance, he was convinced that it was composed of a (i) granular base substance (körnige Grundmasse), (ii) spherical, sphere-like and disk-shaped bodies, and (iii) primitive fibres and strips. The granular base substance he characterized as following:
The granular base substance occurs both in the white and grey matter; but in the latter, it seems, in greater quantity. The fact that it is also found in the marrow mass and here in no small quantity is proved by the examination of the intact upper marrow of small animals. The grains or granules are extremely small, measuring 0.0007″′–0.0005″′ [1.1–1.5 μm] and above, have either a bright and light or a greyish and dark appearance. They lie randomly between and around the closer and the next form components. In this fine-grained mass (feinkörnigen Masse), especially in the brain cortex and in the eye retina, one perceives light-yellowish, larger and smaller bodies of very dull appearance, rounded shape, and somewhat dark contours; they appear in different numbers, sometimes more scattered, now in denser masses, and appear in the former case as gaps in the granular base substance. According to their behaviour, they consist of protein and fat. I could not prove a compound of a nucleus and a bark or a skin on them; they are not vesicles, but simple molecular bodies (molekulare Körper).([59] p. 260)
Terms, such as moleculare Substanz, körnige Grundmasse and feinkörnigen Masse, were found throughout numerous works of various authors who described the structure of the nervous tissue in the middle of the 19th century. Most probably, such structures were an artefact resulted from the improper tissue processing methods used by researchers of that time.
Karel Rokytanský (in German Carl Freiherr von Rokitansky, 1804–1878) was a famous Czech physician; he studied medicine and worked until the end of his life in Vienna. Nowadays almost forgotten, Rokytanský extensively studied the pathological changes of the brain connective tissue [60,61]. In the publication, where he compared the development of tumour stroma with the development of other tissues, he described the similarity of the structure of brain tumours with brain connective tissue (Bindegewebs-Substanzen), and also argued for networked cellular formations in brain melanoma metastases [62]. In a publication dealing with general properties of connective tissue [63] Rokytanský also suggested that inflammation may significantly affect nerve connective tissue, which according to him was a storeroom (Lager) and scaffold (Gerüste) or binding substance (Bindemasse) for neural elements. Describing the scar formation in the brain, he stated:
If, in particular, these conditions are also formed in the brains with the inclusion of the higher sensory nerves and in the spinal cord, this requires the demonstration of a connective tissue substance as the basis and starting point of the new formation. A connective-tissue substance is here present, indeed, as a delicate, soft substance, which in the tissue appears as shagreened, and accumulates more abundantly in the ganglionic substance also containing abundant nuclei (Kernen), in which the elements of the brain-tissue are all embedded. It occurs in the ventricles as the lining of the same, ependyma. But not only here, rather also on the surface of the brain, it appears as a delicate clothing of the cerebral cortex, an outer ependyma formation (äussere Ependyma-Formation) that, similarly as in ventricles, is characterized by the appearance of simple and layered amyloid bodies (amyloider Körperchen). Accordingly, there is a storage and binding mass in the nervous centre, which forms a sheaths and a lining on the outer surface and in the inner spaces.([63]; p. 136)
Subsequent work of Rokytanský was dedicated exclusively to pathophysiology of nerve connective tissue [64]. He described different pathological states, when the primary change was an increase in the number of nuclei and the volume of the connective tissue, which separated the nerve cells, their fibres and vessels. In physiological conditions, to the contrary, connective tissue formed an almost homogeneous matrix. In degenerative disorders, he described an increase of the connective tissue volume and its fibrillation (most likely fibromatosis), which compressed nerve cells and their processes, thus instigating their subsequent atrophy and disintegration into fat particles (Fettkörnchen-Agglomerat) and colloid or amyloid bodies (corpora amylacea). Rokytanský found these degenerative changes in the pons, cerebellar peduncles, cerebral cortex, and even in the cranial nerves. Besides the extensive investigation of the brain connective substance pathology, Rokytanský clearly formulated the concept of the connective tissue as a binding substance, where neural elements are embedded, and a place of the starting point of their new formation. In this sense he might also influenced another famous pathologist of the time, Rudolph Virchow.
Rudolf Virchow (1821–1902) was a person of Renaissance proportion, being a medic, a cell biologist, an anatomist and histologist, an anthropologist, a hygienist, and a statesman. He was born on 13 September 1821 in Schivelbein, which is a small town in Pomerania. Virchow introduced the concept of cellular pathology epitomised in his widely known aphorism “Omnis cellula e cellula” (“every cell stems from another cell”; the erroneous belief that this aphorism belongs to Francois-Vincent Raspail [53,65] could not be verified and we could not find this phrase in Raspail’s writings). Pathological changes in cells or in cellular groups were, according to this concept, the seat of disease. In addition, Virchow was well known hygienist and advocate of social health. He was a member of the German Reichstag, a member of the municipal council of Berlin, and he organised the construction of modern water and sewage systems in the city of Berlin. Politically, he was at odds with Otto Bismarck, and their arguments led to a legendary duel in which Virchow, being challenged by Bismarck (and the argument was about hygienic values of the cities), chose as their weapons two pork sausages: a cooked sausage for himself, and a cooked sausage tainted with Trichinellala larva for Bismarck, who refused to fight under those conditions, considering them too dangerous.
When only 25-year-old, Virchow published a short paper dedicated to the structure of the connective tissue beneath the ependyma of brain ventricles [66]. He confirmed the presence of five basic elements of this tissue, previously described by Rokytanský [67]. In addition, Virchow also described small, round bodies, similar to glass beads or to dew drops, which were relatively densely packed without being interconnected, on the inner surface of the ventricles. If the tissue was well cleaned of blood, it was possible to observe how these units, being star-shaped, were interconnected by fine fibres, so that the entire area was covered by a fine net. Ten years later, Virchow published the same paper again, this time with commentaries [68]; in one of these comments he stated that:
Thus, according to my investigations, the ependyma consists not merely of an epithelium but essentially of a layer of connective tissue covered with epithelium, and although this can be easily removed from the surface, it forms no isolated membrane in the narrower sense of the word, but only the surface of the interstitial connective of the brain substance protruding above the surface... This connective substance forms in the brain, the spinal cord and the higher sensory nerves a type of putty (neuroglia), in which the nervous elements are immersed and which is the main deposition site for Corpora amylacea…([68]; p. 890)
However, the term “neuroglia” was not accepted instantly; for a while, many researchers used the term “nerve connective tissue”. It is still matter of debate as to whether Virchow also observed cells in what he named neuroglia. In the fresh connective tissue, Virchow have seen fine-grained material with elongated nuclei, which according to him, was mistaken for a special type of nerve substance. These elements, however, are far from the meaning of what we today call neuroglial or glial cells. The same concept of neuroglia, including images (Figure 6), was further elaborated by Virchow in his Opera Magna, Die Cellularpathologie in ihrer Begründung auf physiologische und pathologische Gewebelehre, which outlined the foundations of cellular pathophysiology. This book (which was in fact a stenograph of Virchow’s 20 lectures for students of Berlin Medical University Charite) was published first in German [69] and shortly afterwards in English [70]. Virchow’s concept of connective tissue was very similar to that which was published two years earlier by Rokytanský, who first suggested that the nerve connective tissue is a storeroom, scaffold, and binding material for elements of neural tissue [63]. However, it is evident from giving a special term for it (neuroglia), that Virchow was convinced that it significantly differed from other connective tissues in the body. Virchow’s concept of glia become universally acknowledged, and, albeit in a modified form, it has survived up to the present day.
Another important turning point to the cellular concept of the nervous connective tissue was represented by the research of Baltic-German physiologist and anatomist Georg Bidder (1810–1894) from the University of Dorpat (now Tartu, Estonia) and his students. Bidder, together with his former student Karl Kupffer (1829–1902), published an extensive work devoted to the investigation on the texture of the spinal cord and the development of its form elements [71]. Besides the general historical overviews, they also described their own results, which were obtained mainly in cold-blooded animals, that is, fishes and frogs. Bidder and Kupffer were aware of the importance of the connective tissue in the spinal cord; however, they did not consider elements of the connective tissue to be of nervous origin. Their work also included two large chapters devoted to connective tissue. As with other numerous researchers in the field, they were convinced that what they described as a connective tissue in the brain and in spinal cord shared similar properties with the other connective tissues in the body; therefore, one extensive chapter contained the detailed information on connective tissue in general, and in brain and spinal cord in particular. The second extensive chapter contained information on the methods, and how to distinguish the elements of the connective tissue from neural elements, neural bodies and nerve fibres. The publication is also important due to the fact that it gives most likely the first description of the connective tissue (neuroglial) cell in the grey matter of the spinal cord:
…there is also a third type of fibre associated with cells, which are distributed in large numbers throughout the grey matter. It has always been thought that these cells are nerve cells, and they undoubtedly refer to the statement that “the smallest” nerve cells are found in this part of the spinal cord. These are partly round, partly oblong, some without processes, some with two or more processes, and thus angular or star-shaped bodies of 0.003–0.004″′ [6.4–8.5 μm] in diameter, with deep dark contours, and a generally sharply delimited nucleus in the middle. They are fairly uniformly distributed throughout the grey matter, and no particular law can be demonstrated in their arrangement. The processes are often so delicate that they do not present two lateral boundary lines, but appear as a simple wavy line. The processes of neighbouring cells often overlap, giving rise to an appearance, as shown by the anastomosing processes of the bony bodies in a thin bone section. Of particular note, however, is the fact that the processes of these cells are sometimes associated with those fibres which emanate from the pointed and grey mass end of the spinal cord epithelial cells. If this connection alone would be sufficient to refute the nerve-cell nature of these elements, then the deficiency of the yellow colour, which in the chromic-acid preparation belongs to the unquestionable nerve-cells, and the missing connection with definite nerve-fibres, speak against it. Thus, these cells are connective tissue corpuscles, and the fibres associated with them are the admixtures to the connective tissue known as spiral or elastic fibres.([71], p. 45)
Unfortunately, this description had no accompanying figure. However, Kupffer’s earlier dissertation [72] contained results indicating that the connective tissue is an integral part of the spinal cord grey matter. Investigating the cellular elements in the frog spinal cord grey matter, Kupffler observed cells which he at first determined to be supposed nerve cells, but afterwards, he also observed that they had definite connections with undoubted connective tissue fibres. He afterwards had to classify the considerable part of these cells not as essential nerve elements, but as connective tissue corpuscles (Figure 5C).
3. Discovery of Glial Cells as Cellular Entities
3.1. Retinal Radial Glia: Samuel Pappenheim, Heinrich Müller, and Max Schultze
Probably the very first description of the histological structure of the retina was published by Samuel Moritz Pappenheim (1811–1882), who was Purkyně’s assistant in Wroclaw. They published several papers together on histology and physiology. Pappenheim also published several extensive monographs, one of which was dedicated to the structure of the eye [73]. All findings, especially the description of cellular elements, were based on a long period of research, and on the extensive amount of processed tissues, including bones, eye ball, and eye accessory organs of different species of animals and of humans. This book also contained the very first detailed description of the histological structure of the retina, which included the description of radially oriented cells (most likely radial glial cells, Figure 7A), of dark red granules (probably rhodopsin) and layers of ganglion cells.
Almost 10 years after Pappenheim, Heinrich Müller (1820–1864), who held a chair at the University of Würzburg, published a preliminary communication on the structure of the retina. In this paper, he described in detail the layers of the retina by using chromic acid [74]. He provided a description of the cell layers, of the radial cells extending from the inner to the outer layer, and also of cellular extensions in the granular layer, with these extensions containing the nuclei. Five years later, Müller published further descriptions of the retina [76], which contained figures of individual cellular elements in different species of animals, including radial glial cells, which were subsequently were named after him (Müller glia, Figure 7B).
Three years after Müller’s descriptions of the retina, German microscopic anatomist Max Johann Sigismund Schultze (1825–1874), published an advanced treatise on the structure of the retina [75] on the occasion of his appointment as a professor of anatomy and director of the Institute of Anatomy in Bonn. Besides the description of other structures, Schultze devoted the whole chapter to radial glial cells (he named them fibras Muellerianas to emphasize that they were described for the first time in detail by Heinrich Müller). The treatise, written in Latin, also contained images of retinal structures, and particularly of radial glial cells, which were of exceptional quality with the minutest details (Figure 7C).
3.2. Carl Bergmann
German anatomist, physiologist, and biologist Carl Georg Lucas Christian Bergmann (1814–1865), who was professor of anatomy and physiology at the University of Rostock, published a short paper about some structural relationships in the cerebellum and spinal cord [77]. In the first part of this paper, Bergmann confirmed previous descriptions of conical cells surrounding the central canal of the spinal cord of frog; processes of these cells entered into the grey matter of the spinal cord and were interconnected with each other (Figure 8A). However, he disagreed with the idea that cell processes were linked to elements of connective tissue. In the second part of the paper, Bergmann described the cellular structure of cerebellum of a newborn kitten, the preparation was stained with chromic acid. Between the grey matter and pia mater, he observed a great number of very fine, branched, parallel, and radially arranged fibres:
The isolated fibres showed short (probably broken) branches starting at acute angles. These have partially the direction toward the periphery, but also partly into the interior of the organ. The latter circumstance may contradict the assumption that the fibrils discussed belong to the branching of the ganglia bodies, whereas there are other reasons as well. (Especially in the case of the kitten, where these fibres are so clear, the processes are hardly to be found directly on the ganglion bodies.) On the other hand, it has become probable for these branches, which are so different in direction, that the fibres in question form a net in grey matter. On an isolated fibre, which protrudes by 0.004″′ [85 μm], is nicely visible ending embedded in the clear substance, the fibre here transients into some finer fibrils. The picture recalls (only noticeably gentler) the behaviour of the radial fibres of the retina, where they approach the membrana limitans [77].
Bergmann was the first to describe these fibres of specialized cerebellar astrocytes whose bodies are located in the layer of Purkyně neurons, with their processes running throughout the molecular layer, all the way up to the pia mater. These cells are known today as Bergmann glia.
3.3. Ludwig Mauthner
Ludwig Mauthner (1840–1894) who was born in Prague and was professor at Innsbruck University, and subsequently held a chair in ophthalmology in Vienna, described, in 1861, the cellular elements of the connective tissue of the brain and the spinal cord [78] in fishes (Figure 8B). Around the central canal of the spinal cord, he found epithelial cells with large processes (ependymoglial cells) which, without contact with other cell formations, extended to the periphery of the spinal cord and disappeared into the fibres of the spinal meninges. Mauthner observed that connective tissue cells were about 10 times smaller than the ganglion cells (as neurones ware called in those time). Connective tissue cells appeared in clusters in the ventral horns of the spinal cord, and formed a network consisting of their fibre-like processes. Similar cells were also observed in the cerebellar nuclei. Mauthner concluded that connective cells in the grey matter represented the basic elements of connective tissue; furthermore, he considered them as neural cells and not as the particles of connective tissue. He also assumed that the formations, which were previously considered to be connective tissue cells, were in fact larger nuclei of cells whose cytoplasm was so transparent it seemed that that an empty space surrounded the cell nucleus.
3.4. Lionel Beale
Physician and microscopist Lionel Smith Beale (1828–1906) was a professor at King’s College in London; he was also a President of the Royal Microscopical Society (Figure 8C). He studied in detail the structure of the nerve tissue, and wrote numerous publications in which he dealt with the development, internal structures, and interconnections of neurons. Beale had an opinion that connective tissue and connective tissue corpuscles are produced from the very same masses of germinal matter as those from which nerve cells and nerve fibres were developed, and that they are the remains of nerve cells, nerve fibres, and vessels which were in a state of functional activity at an earlier period of life. He was also convinced that true nerve fibres, which convey the nerve current do not lose themselves in the connective tissue or blend with it, nor are they connected with its corpuscles, but they form networks. In addition, a normal nerve fibre can always be distinguished from a fibre of connective tissue. However, Beale also considered the connective tissue of the brain in the following, rather philosophical (and prophetic!) manner:
It is possible that, for many years to come, some observers will persist in terming everything in which they fail to demonstrate distinct structure, connective tissue, and all nuclei which are not seen in their specimens to be in connexion with positive vessels, positive nerve-fibres, or other well-defined tissues besides fibrous tissues, connective-tissue corpuscles; but there is little doubt that when the changes occurring during the development of special tissues shall have been patiently worked out by the use of high powers and better means of preparation, opinions on the connective-tissue question will be completely changed. The idea of the necessity for a supporting tissue or framework will be given up, and many structures now included in “connective tissue” will be isolated, just as new chemical substances year after year are being discovered in the indefinite “extractive matters”.([79]; p. 25)
3.5. Carl Frommann
Carl Friedrich Wilhelm Frommann (1831–1892) was a German physician; in 1870 he habilitated at the University of Heidelberg and became a private lecturer for histology; in 1872 he moved to the University of Jena, where he was, in 1875, appointed as professor. In 1864 (being in Weimar as an ordinary doctor) he published a paper summarizing the results of staining of the spinal cord with silver nitrate [80]. In the longitudinal sections of the white matter of cow spinal cord, the connective tissue, which, as previously was proposed by Virchow, Frommann called “neuroglia”, appeared to be a very fine dark-brown network or mesh of interconnected fibres (Figure 8D). The elements of this web were quite variable, the larger were round, oval or square, the smaller one were usually round, and were either isolated or grouped; sometimes they formed long narrow rows. The fibre network was arranged in parallel with the nerve fibres. Cells appeared to be uniformly brown, or with a lighter core, even without sharp contours; sometimes it seemed that the nucleus was a bit darker than the protoplasm. The cells were, by means of their processes, connected to the fibre network, and were also interconnected. On the cross-section of the frozen spinal cord, which was thawed in the silver solution, the neuroglial web was not usually visible, and the shape of its elements resembled a very fine granular mass. Similar findings Frommann were also obtained in the human spinal cord.
Concerning the connective tissue fibres, Frommann believed “that all fibres originate from the cell processes and that they are hollow, and therefore, all white matter connective tissue consists of a continuous network of channels of varying size, which form cellular collectors and centres by numerous connections” ([80]; p. 46). In general, however, his papers and books clearly demonstrate that at his time, there was no clear definition of terms such as “glia”, “glia mass” (for which he used Leim-masse) and “connective tissue”, as evidenced by some of his statements. For instance, Frommann claimed that glial fibres (Glifasern) are found inside of the glial mass, or that “it is characteristic of connective tissue cells, unlike neurones, that they are interconnected with their processes to the fibre glial networks...” ([81]; p. 9).
3.6. Otto Friedrich Karl Deiters
German neuroanatomist Otto Friedrich Karl Deiters (1834–1863) spent most of his short professional career in Bonn. Deiters’s most famous work was his treatise on the structure of the nervous system, which substantially advanced studies of the cellular structure of the nerve tissue [82]. This book was published two years after Deiter’s untimely death (he perished at 29-year-old from typhoid), by his colleague Max Schultze, who, on the basis of several Deiter’s illustrations and notes, attempted to organize the results of his histological research as logically as possible. This book contains 13 chapters, and chapter 3 is dedicated to the brain connective tissue (Bindesubstanz). Similarly to his contemporaries, who were convinced of the existence of connective tissue in the brain and the spinal cord, Deiters sought to find its characteristic features in order to distinguish them from neurones. He postulated that the connective tissue in the central nervous system (CNS) differed from that in other organs:
One is accustomed to discuss only the distinction between connective tissue and nervous tissues in the central organs, or to call anything that is not nervous, briefly connective tissue. For the time being it would be more correct, as some wish, to separate nervous elementary parts from those which are not connected with the nervous system. The further distinction is certainly for the moment indifferent. That the non-nervous tissue of the central organs does not readily have the character of ordinary connective tissue is plausible, and here, too, certain conditions are certainly not justified in any way. If, therefore, many people oppose the designation of such parts as connective tissue, this is probably nothing more than that the concept of connective tissue is far from exhausted, and may perhaps reveal unexpected new sides.([82]; p. 35)
Deiters did not use the term “neuroglia”, although he was well acquainted with Virchow’s concepts. Deiters provided detailed descriptions of all elements of the nervous connective tissue. The basic structure of this tissue was spongy mass (schwammige Masse) or porous mass (poröse Grundmasse), containing several types of fibres. These were either connective tissue fibres (Bindegewebsfibrillen, elastischen Fasern) or intercellular support fibres (interzelluläre Stützfasern) which Deteirs suggest to put:
…aside the Mullerian fibres of the retina, and thus to establish another analogy between this tissue and those of the central nervous system… Thus, it may well be assumed that such intercellular support fibres may form an essential link in the whole tissue arrangement… It can generally be established as a law that such a streaky arrangement of the connective tissue mass goes hand in hand with a regular linear arrangement also of the nervous parts; such represents the retina, such in the cerebellum, in the Ammon’s horn (hippocampus), etc.([82]; p. 43)
Deiters also described and discussed the presence of the formations called “free nuclei” which could be in fact some transition forms of cells without detectable cytoplasm and processes. He also described in the connective tissue cellular elements which he named cells of the connective tissue (Bindesubstanzzelle):
Close to the glossy nucleus, which contains no nucleoli, we can see a mass of outgoing fibre tracts, which from the beginning have a firm though delicate appearance, a very sharp, smooth contour, a considerable shine, and which radiate to all sides. These are easily movable, twisting on isolated cells many times, and are not fragile. They divide very soon and then ramify in the most varied way under always forked splitting. I do not think that anyone who sees such an isolated element, will want to think of artifacts, accidental coagulation.([82]; p. 46)
Deiters observed these cells in both white and grey matter, in the substantia gelatinosa of the spinal cord, and in the medulla oblongata (Figure 9). Unfortunately, as stated almost 10 years later by Franz Boll (see further in text) “in general it is to be noted that the excellent disputes of Deiters, as well as his positive discovery of the true form of these connective-tissue cells, have been neglected by almost all his successors in an almost irresponsible manner.”
3.7. Leopold Besser
German physician Leopold Besser (1820–1906) worked as a provisional intern in the mental illness clinic in Siegburg, Germany. In 1866 he published a paper, which had a great impact on the research of neuroglia; this paper described the development of nerve tissue in the neonate brain with particular emphasis on the role of neuroglia [83]. He demonstrated the emergence of neuroglial cells not only in grey matter but also in the vicinity of capillaries (Figure 10A) and was convinced of its importance during nerve tissue development. Besser particularly noted:
This mass of connective tissue of central organs, which Rokytanský called “connective tissue of ependymal formation” (Bindegewebe der Ependym formation) and Virchow in 1846 suggested to name as “neuroglia”, Kölliker referred to as “connective substance forming networks”, or “the reticulum” (netzförmige Bindesubstanz, Reticulum), whose “spongy part” (Schwammigen Theil) were considered by Deiters to be “intercellular substance” (Intercellularsubstanz), and Hensen named its multi-nuclear elements “parenchymal cells” (Parenchymzellen), has in the brain of newborns so essential meaning and its formative character has a determined dimension and form that I have to name it as “neuroglia of newborns” (Neuroglia des Neugebornen) because it has properties that the adult reticulum does not have in its normal condition.([83]; p.309)
Although Rokytanský and Virchow suggested differences between the neuroglia found in nervous tissue vs connective tissue in the rest of the body (see [84]), the question of connective tissue containing fibres and loose nuclei on the one hand, and neuroglia on the other hand, still remained under debate. Besser adopted Virchow’s concept and used term “neuroglia” to emphasize its different properties from the connective tissue of other organs and systems. In particular, he wrote:
The name neuroglia—Nervenkitt—chosen by Virchow could be discussed with a bit of a criticism because this matter is far from being putty, clinging or sticking to cover the nerve elements. It is also not the most important supportive structure because much more closely arranged vessels are more important for the shape and support of nervous parts. But this matter has such specific features and is so different from all other types of connective tissue formations that it is quite justified to stick to a special name.([83]; p. 310)
According to Besser, glial mass contained corpuscles that could be easily observed around evolving blood capillaries. According to him, the glial matter itself consisted of two separate structures, which were, however in a mutual “genetic” relationship. Besser named them as “Glia-Kern” (glial nuclei) and “Glia-Reisernetz” (glial finely branched nets). Beser’s term “net” did not describe networks formed by the glial cell processes, as we know today, but the glial nets attached to “glial cores” and from them, growing fibrous felt. According to him, neuroglial origin, development, transformation, and interrelationship formed the embryology of the nervous basic parts. “Glial nuclei” (which today’s we recognise as glial cell bodies) were formed by dividing, and before reaching the size of red blood cells, they remained homogeneous. Shortly thereafter, however, with their growth, a fine granulation emerged on their surface from which developed protuberances and branched processes forming networks. The shape of the delicately branched processes seemed to Besser be round, like the branching of the small twigs of our trees. He was convinced that after death, these branches changed so much that we could not observe them anymore. These networks, however, in the course of further development, according to Besser, separated and gave rise to other structures.
Besser was of the opinion that all nerve elements of the human CNS developed from the part of neuroglia that had morphogenetic properties. According to him, the nuclei of the neurones were transformed glial nuclei, the neuronal bodies were formed from glial branched networks, and the neuronal processes were formed from glial processes. Nerve fibres developed from long branches of proliferating glial nets, and axons were formed by the conversion of fine fibres of branched nets. Besser also described the involvement of glial cells in the capillary development in the cerebellar cortex of newborns. As he noted, thousands of these glial structures touched each other in parallel layers that were stacked together on the capillaries and on the smallest blood vessels. The significance of Besser is that, besides the conviction that the neuroglia in the nervous tissue and the connective tissue were different, he also suggested their importance in the development of the CNS, and for the first time, displayed star-shaped neuroglial cells, probably astrocytes, attached on blood vessels.
3.8. Albert Kölliker
Albert Kölliker (1817–1905), a professor at University of Würzburg, in 1867 published the fifth edition of his extensive manual on the human microscopic anatomy [85]. This handbook, as well as its previous editions, contained a detailed summary of contemporary knowledge about the structure of nerve tissue. Besides the detailed description of the nervous elements of the brain and spinal cord tissue, that is, nerve fibres and nerve cells, Kölliker also dedicated a relatively large section of this handbook to the connective tissue (sometimes he referred to it as the supporting tissue—Stützsubstanz). In the spinal cord he described the connective substance as following:
…here, apart from the pia mater and its processes in the anterior cleft and the adventitia of larger vessels, there is no ordinary fibrillar connective tissue, but only a simple binding substance, consisting entirely of nets of star-shaped connective tissue cells (connective tissue bodies sap cells—Bindegewebskörperchen Saftzellen) or of scaffolds of coreless fibres and trabeculae resulting from the cell networks, as described in the general parts (§ 23) as part of the cytogenetic binding substance. These nets and scaffolds, which I denote by the name of the net-like binding substance (netzförmigen Bindesubstanz), where they occur singularly as the supporting substance of other tissue elements, are found in the spinal cord in both substances in such a development that they form a very significant part of the whole mass of the organ. In other words, they form a delicate skeleton running through the whole white and grey substance, which I shall call the reticulum of the central nervous system (Reticulum des zentralen Nervensystemes), which contains in its numerous gaps the cells and nerve tubes and carries the blood vessels as well.([85]; p. 266)
Kölliker believed that fine-grained matter containing numerous nuclei and observed by a number of authors in nerve connective tissue, was an artefact. Under much larger (up to 300 times) magnification, he observed that it consisted of connective tissue cells forming a network of structures. Perhaps under the influence of Deitres, who described and visualized connective tissue cells in some areas of the brain and spinal cord, Kölliker called one type of cells, which in earlier issues he named as “cells in the central grey cores of the human spinal cord”, connective tissue cells (Figure 10B). He also emphasized the role of the mentioned “reticulum” during pathological states, which, according to him, increased the proportion of interstitial matter that became fibrous.
3.9. Theodor Meynert
Austrian neuroanatomist and psychiatrist Theodor Hermann Meynert (1833–1892) worked together with Karel Rokytanský in Vienna. He studies the cortical structures of the brain cortex, as well as the interconnections of certain regions of the brain, through the neural pathways. Meynert also developed new cell markers and used thin serial slices stained with carmine or gold for quantitative measurements of neurones and investigation of the brain tissue structure. A comprehensive summary of his research was published in Stricker’s Manual of histology [86]. Concerning connective tissue, Meynert referred to it as to a basement-substance (Grundgewebe). Similarly to Besser, he pointed to the inconsistent terminology of his contemporaries, noting:
This basement-substance is called by Rokytanský ependymal formation (Ependymformation), by Virchow, neuroglia, by Kölliker, connective tissue (Bindesubstanz), by Deiters, spongy tissue (schwammige Substanz), by Henle and R. Wagner, fused ganglion-cell substance (zusammengeflossene Ganglienzellenmasse). Occurring in the olfactory lobe and ammon’s horn, it receives from Clarke the designation of gelatinous (gelatinöse), from Kupfer that of molecular substance (moleculare Substanz).(Cited from the English translation [87]; p. 660)
Unlike Deiters, who recognized only loose nuclei in the connective tissue of the cerebral cortex, Meynert insisted “upon the indisputable fact of the occurrence here of the star-shaped cells, with very little protoplasm, and a great number of the finest possible processes” (Figure 11A). Moreover, he also noticed, that “these cells swell up under certain pathological conditions and assume grotesque forms”. In addition, Meynert was the first to declare that the basic substance of the grey matter of the cerebral cortex contains spongioform structures made up from the processes of cells other than neurones.
3.10. Rudolf Arndt
German psychiatrist Rudolf Gottfried Arndt (1835–1900) studied the morphology of the brain tissue of large vertebrates and humans. He summarised his findings in a series of three papers [89,92,93]. In the first paper he described five to six layers in the cortex. In the first layer, there were intertwined parallel fibres, some of which were nervous, another belonging to the fibres of the connective tissue. The second layer, according to Arndt, consisted mainly of neuroglia, which was grainy-fibrous (körnig-faserigen) or spongy (schwammigen), and which was, according to him, differentiated protoplasm, neither nervous nor connective. In the neuroglia were nuclei and loose thin and thick fibres; some ended with the loops described by Gabriel Valentin. The third layer contained isolated nuclei and nerve cells with a nucleus, all immersed in neuroglia; their shape was either round, elliptical or triangular (Figure 11B). From the latter emerged horizontally and inwardly oriented processes, one thick process pointing to the surface of the cortex, which he considered to be an axon. Whether these processes served to interconnect cells, passed into nerve fibres, or dissolved in neuroglial tissue, Arndt was not certain.
The second paper mostly contained polemic about the works of Besser, Meynert, and others that had been published in the meantime. It also included consideration of the purpose of the corpuscles of the connective matter, which he believed to be a remnant of the development of nervous mass in the foetus. Arndt also accepted Besser’s terminology (Gliakern, Gliareiser) and an idea on the development of nerve tissue elements; he assumed that from these two glial components developed three brain elements: blood vessels, neurons, and nerve fibres. In the third, shorter paper, Arndt confirmed his findings from the previous publications and discussed the functional unity of ganglion bodies and their processes, together with the surrounding granular-fibrous substance:
Concerning the nerve cell of the central organ of an adult, I believe that according to the foregoing description the ganglion body together with its central processes and the part of granular-fibrous substance, which is genetically related to it, must be understood. Whether we will ever get to know them intact is questionable, indeed highly unlikely. It may not even exist on its own, although this is still quite conceivable, but it is probably so completely merged with adjacent parts, so intimately interwoven with them by their finest threads, that together they are only one, their creation form absolutely indivisible mass. But if this is the case, then we must logically consider the interganglionary granular-fibrous substance, the terminal fibre-network, as it turns out in the adult, to be a coalesced protoplasm modified for definite purposes, as an irritable tissue, the actual carrier of all is central operations.([89]; p. 329)
Arndt was thus one of the first researchers who hypothesized that the nerve cells and the surrounding granular-fibrous substance are developmentally related and form a single functional unit.
3.11. Ludwig Stieda
The Baltic German Ludwig Stieda (1837–1918) was professor of anatomy in Dorpat and Königsberg. He studied the structure of the neural tissue, predominantly in animals. Stieda published many works and he summarised his findings in two extensive publications [90,94]. According to him, the CNS consisted of nerve cell bodies, nerve fibres, connective tissue, blood vessels, and epithelium. There was no general agreement on the composition and terminology of brain connective tissue or neuroglia, and Stieda absolutely correctly pointed out that there was no agreement on the terminology on the nerve cells as well. As he noted, these formations were named by various names, such as ganglion bodies (Ganglienkörper), nerve bodies (Nervenkörper), ganglion balls (Ganglienkugeln), occupancy balls (Belegungskugeln), spinal bodies (Spinalkörpen), ganglion cells (Ganglienzellen), or nerve cells (Nervenzellen). Concerning the connective tissue, Stieda did not distinguished the true fibrous connective tissue in meninges from what was called connective tissue in the grey matter, where it, according to him, had a fine-grained appearance and contained nuclei of the basement substance (Figure 11C). He also did not consider the basement substance to be nervous, but to be a special category of supportive or connective tissue of the nervous system. Moreover, Stieda could not confirm the presence of the grained basement substance as a network of cells interconnected with anastomoses, as was described by other researchers.
3.12. Jakob Henle and Friedrich Merkel
Four years after the publication of Deiters’ monograph on the structure of nerve tissue, Jakob Henle (1809–1885), who was professor of anatomy in Gottingen, and his son-in-law Friedrich Merkel (1845–1919), published a paper dealing with the connective tissue of the CNS [91]. Although this work contained a detailed historical overview of connective tissue research, the authors did not even mention Deiters’ findings. The goal of the Henle and Merkel study was to determine whether fine-grained or reticular tissue had the properties of nervous substance, or was only a supportive, connective, essentially cementing substance. For the study of tissues from various regions of the brain and spinal cord of several mammals and humans, they used a variety of histological methods, from carmine staining followed by fixation with chromic acid, to boiling, using differently diluted acids and bases, or alcohol hardening. Henle and Merkel were convinced that the connective tissue consisted of a so-called molecular mass and the elements of the connective tissue (Figure 11D), that is, fibres, cells, and grains or nuclei:
These connective tissue cells could, according to the analogy of the nerve cells, be called bipolar, as opposed to the multipolar connective tissue cells, from which three or more fibres emanate. From multipolar cells merging with their processes, the above-mentioned reticular connective tissue develops; similar cells, growing in several threads, but usually preferentially in two opposite directions, owe their origin to the confused fibres of the innermost layer of the pia mater. The fibres can be traced over long distances and, although they branch out, do not appear to anastomose regularly; they are more of a felt than a net. The cells are observed in this layer only with difficulty, isolated, hardly without the help of Carmin absorption. More clearly, because more scattered and less obscured by fibres, they are found on the border of the pia mater and the cortical layer, and in the latter itself.([91]; p. 57)
Although Henle and Merkel referred to the term “neuroglia” only in relation to Virchow’s results and did not use it at all in the description of their own results, they suggested that the fibrous networks described by them could be considered as so-called neuroglia or Kölliker’s reticulum.
3.13. Camillo Golgi: Early Studies on Neuroglia
In the early 1870s, Camillo Golgi (1843–1926), while a 27-year-old employee of the University Clinic in Pavia, published his first brief communication on the connective tissue of the brain [95], which a year later, was translated to German [96]. Golgi studied the connective cells of the cortical regions of the human brain, using staining primarily with osmic acid. He observed circular, oval, or star-shaped cells with a large nucleus, from which several very long processes emerged in different directions. He even described anastomoses between the processes of neighbouring cells.
During the same year, Golgi began to publish an expanded three-part version of the previous communication with a detailed description of his findings [97,98,99]. In the three chapters, it contained a detailed description of the results obtained in the brain, cerebellum, and spinal cord. Golgi did not use the terms “neuroglia” or “glial cells” in these publications. The introductory historical overviews of the findings at the beginning of each chapter are very instructive. Golgi observed the cells described in the previous publication, in various regions of the brain and spinal cord (Figure 12A), and he summarized the results as following:
In summary, with the preparations in osmic acid I have demonstrated:
- That a very thin superficial layer of the brain cortex is exclusively constituted, except of some bundles of nerve fibres coming from the marrow, from the connective cells provided with a large number of thin, long and for the most part rigid unbranched extensions.
- That identical cells are scattered in considerable numbers in all the layers of the cortex, forming a continuous support tissue.
- That many extensions of the connective cells are inserted in the contour of the vessels, and that there is neither the space nor the perivascular lymphatic network… ([97]; p. 344).
In addition, Golgi employed several modifications of osmic acid staining, and also described the pericellular lymphatic space. By using different staining methods on very thin slices of the cortex, he was able to observe a large number of “elegant” connective cells with 10–30 very thin and long processes, very rarely branched. These branches separated at a short distance from the cell boundary, and he never observed more than two or three secondary branches, which were also very long and unbranched. Golgi also commented on the contents of the neuropil, which, according to today’s findings, is part of the grey matter that fills the space between the perikarya and the vessels, where the nerve fibres, the synapses, and the glial cell processes are located, and which appear to be amorphous under the light microscope. According to Golgi, the so-called fine-grained, possibly mesh-like, spongy or dotted molecular, amorphous or gelatinous matter originated either as a result of changes in the dead body, due to reagents or tissue processing. These effects led to the breakdown of the fibrillary substance, which, in his opinion, was likely to occur not only in the processes of connective tissue cells but also in the finest protoplasmic processes of neurons. It should be noted, however, that the above mentioned findings by Golgi were made before his discovery of the biochemical method of colouring the nerve tissue preparation with potassium dichromate and silver nitrate in 1873 [101], now known as the Golgi method or reazione nera.
3.14. Moritz Jastrowitz
German neurologist and psychiatrist Moritz Jastrowitz (1839–1912), working as an assistant at the clinic for mentally and neurotic patients in Berlin, published in 1870–1871 a two-part paper on the structure of nerve tissue during encephalitis and myelitis in children in the early postnatal period. In the first part, he described in detail the anamnesis of patients from whom he took brain tissue, and described the macroscopic tissue changes [102]. In the second part, he examined in detail the individual elements of nerve tissue and neuroglia, which, according to him, also played an important role during encephalitis [100].
Jastrowitz in his paper, clearly distinguished neuroglia and connective tissue, which consisted of spongious structures, fibres, granular (molecular) substances, and also lymphoid cells. The molecular substance, according to Jastrowitz, was associated with the myelination of nerve fibres, since during the development of nerve tissue, it was gradually lost. Therefore, he considered it not only as a supportive matter, but also as an embryonic tissue that was responsible for axonal isolation. Jastrowitz also believed that molecular substance played a more important role than connective matter, and therefore recommended not to include it among the other elements of connective tissue under the common name of “neuroglia”.
The most prominent non-nerve cells, which, according to Jastrowitz, no one had mentioned before, he found in the ependyma (Figure 12B). As he noticed, “If Virchow’s view, initially expressed by the ependymal layer of the fourth ventricle, that in it the neuroglia comes to light in its purity, also applies to this location, then these would be true prototypes of glial cells”. Jastrowitz described these cells as following:
They generally surpass those of the third layer in volume, not inconsiderably, especially with respect to the diameter of the base, and differ in shape and size from one another. Most are spindle-shaped, but also rounded and angular as well as cylindrical shapes are encountered; they are often narrowed at one end and may be quite pointed at this, sometimes at both ends. Their length is usually double and even three times their width, on average for the smaller ones 0.007–0.01: 0.004–5 [length 15–21 μm, width 8.7–10.6 μm] and for the large ones 0.012–0.017: 0.005–7 [length 25–36 μm, width 10.6–14.8 μm]. The protoplasm is a little more granulated and less transparent, usually it contains very large oval nucleus with several granules larger than nucleoli is often located at one end of the cell, and particularly is intensively coloured, therefore one easily overlooks its contents in the tissue. An immense amount of delicate, bright processes extend from them widely in all directions, often across each other, and finally disappear in the molecular mass, whose particles usually adhere to them in isolation. Despite the long distances they travel, divisions are exceptional, but often kinks and bends. They are chiefly those who give the whole structure a very characteristic appearance, which resembles that of a little spider.([100]; p. 169)
Jastrowitz, therefore, named these cells as “spider glial cells” (spinnenähnliche Gliazellen) or “spider cells” (Spinnezellen). Their number increased toward the surface of the brain ventricle so that they finally formed the epithelium of the ependyma. Jastrowitz disagreed with Besser, who believed that the glial elements were made up of separated glial nuclei and glial networks. The glial nuclei observed by Besser, according to Jastrowitz, corresponded in part to embryonic cells, in part to the nuclei of connective tissue cells and neurons, and partially to free nuclei.
3.15. Victor Butzke
Russian psychiatrist Victor Butzke (1845–1904), while collaborating with the German pathologist and histologist Eduard Rindfleisch (see below) at the Pathological-Anatomical Institute in Bonn, published in 1872 a paper dedicated to the fine microscopic structure of nerve cells and glia and their interrelationships [103].
Butzke has extensively described other types of cells than nerve cells. As he stated, “not everything must be considered as a connective substance, or even as a connective tissue, that is not recognized as a ganglion cell or nerve fibre in the central organs of the nervous system” ([103]; p. 590). He questioned the existence of the so-called “free” nuclei, believing that even these were surrounded by protoplasm that often passed into the processes, and he also distinguished glial elements with thin and thicker processes, partially oriented radially, which were found on the surface of the large brain (Figure 13A). Other glial elements were cells with a relatively large nucleus and radially emerging thick processes, which, according to Butzke, were scattered among the well-known real nerve elements of the grey matter of the cerebral cortex, and furthermore, he detached them into a special category, “real connective tissue corpuscles with their thicker and thinner processes” ([103]; p. 591; Figure 13A). In addition, he also expressed a hypothesis on the physiological role of the glial network in the nervous tissue, namely, its role in the formation of new connections between nerve cells:
The fine-grained fibrous mass is therefore not a simple putty substance (Kittsubstanz) for us, which is only intended to adhere the parts together; it is no longer the simple intermediate mass (Zwischenmasse) through which the coarser structures pass, but it also contains one enclosed within it highly subtle tissue in the form of a network, which is formed by the end fibrils of the ganglion processes and supported by the processes of the glial corpuscles (Gliakörperchen). I now put forward the hypothesis, which has never been formulated sufficiently clearly and sufficiently supported by reasons that the connection between the end branches of the ganglion bodies takes place through this fine irregular network. I have not seen clearly that connection, so I call my view a hypothesis. But if you look right where the finest branches are, and you see a network that starts directly from the final ramifications, what other conclusion could you make? … Nor are we aware of any facts that suggest that not every physiological connection between the elementary centres could be fixed, strengthened, and newly formed: in practice and in learning. But what about the notions of learning, forgetting, practicing, getting used to, etc., if all the connections of the functioning elements were given once and for all in rigid anastomoses, or in the entry and exit of nerve fibres into a ganglion body? … But that is probably no longer so necessary and, in order to come back specifically to the one organ which I subjected to the more detailed examination, I will ask, if not every impression which arrives at the brain through the senses and adheres to it is proof enough that there are suddenly created a hundred new connections that may exist for life? And is not every new thought that comes to us proof enough that out of the old connections that already existed, new ones were formed automatically (that is, without influence from outside), others were strengthened, others disappeared or become weaker? It is, after all, to quote Master Goethe in the end: “the thought factory is a weaver masterpiece, where—one stroke strikes a thousand connections.”([103]; p. 595)
Like Golgi, Butzke also drew attention to the particular funnel-like shape of some of the glial processes that were adjacent to the vessels or the meninges.
3.16. Louis-Antoine Ranvier
French physician and histologist Louis-Antoine Ranvier (1835–1922), who worked at the Collège de France in Paris, is universally known as a discoverer of the nodes of Ranvier, which are regularly spaced discontinuities of the myelin sheath, occurring at varying intervals along the length of a nerve fibre. It is less known, however, that Ranvier was also interested in the structure of connective tissue. In 1873, he published a brief note on the spinal cord connective elements [105]. Through a golden cannula, he perfused white and grey matter of the spinal cord with osmic acid, and thus in stained longitudinal slices, he visualised homogenous black structures in white matter and marbled black structures in the grey matter. In white matter, he described neural tubes (axons) with myelin sheaths, among which he observed fibrils or bundles of connective tissue fibrils. The small bundles of connective tissue in spinal cord white matter, according to Ranvier, did not produce anastomoses, but at some points four to eight or even more intersected one another. At this crossing, a round or oval flattened nucleus with small nucleoles surrounded by granular mass was often observed. This whole unit Ranvier named as a flattened cell of connective tissue (cellule plate de tissu conjonctif). The same structures he also described in detail in grey matter. Consequently, Ranvier noted that the connective tissue of the spinal cord was constituted by bundles of connective fibrils or fibres and flat cells.
Ten years later, Ranvier published an extended paper containing figures, describing his new achievements in neuroglial research [104]. Concerning the term “neuroglia” and the meaning of it, he wrote:
The word neuroglia is detestable and if I use it, it is only because it has passed into the language of histologists. All know that it designates the connective tissue of the nervous centres; but if they agree to adopt it, they do not agree on the organisation of the neuroglia itself, because some believe with Henle and Merkel, that it is formed of elements of the connective tissue ordinary; others, with Gerlach, that it is composed of elastic fibres or analogous to certain elastic fibres, that it consists entirely of branched cells of a special nature of Deiters cells (Boll, Golgi, etc.). Some, finally, think, with Schwalbe, that the cells which are between the nervous elements of the brain, the marrow, and the optic nerve are nothing but the lymphatic cells in migration. I myself have maintained that the marrow neuroglia is composed of fibres of any length, intersecting at certain points, at the levels of which are generally flattened cells.([104]; p. 177)
Using the method of hardening the nervous tissue of his own, Ranvier was convinced that the neuroglial cells do not have the rudimentary forms attributed to them by Deiters. According to him, their nucleus was well formed, their well-developed cell bodies were membranous, irregularly star-shaped, and showed shape-related accidents that depended to a large extent on the pressures exerted by neighbouring elements (Figure 13B). They had numerous processes which, as Ranvier believed, were nothing but fibres of any length, so distinct in the preparations made by means of the interstitial injections of osmic acid, and which, in these preparations, appeared to be independent of the cells. These fibres which passed through the neuroglial cells did not always follow a rectilinear path; a large number of them described curves or were folded in the shape of a loop.
Ranvier also believed that nerve cells and cells of the neuroglia had a common origin; they both proceeded from the primitive neuroepithelium, and although they may have had absolutely different physiological significances, they retained in their structures certain points that betrayed their common origin. He was also convinced that the granular substance, which various researchers had attached to the neuroglia, did not exist. Ranvier summarized the composition of the nervous tissue as follows:
The grey matter of the brain and that of the spinal cord are thus reduced to the same type. Both are composed of ganglion cells, myelinated nerve fibres, myelin-free nerve fibres, and neuroglia cells, leaving, of course, the blood vessels and their perivascular sheath, which belong to the ordinary connective tissue.([104]; p. 185)
Concerning the general view on the nerve tissue structure, Ranvier’s observations were correct. However, his description of the neuroglial cells as cells through which passed neuroglial fibres, was stepped aside and it significantly influenced neuroglial research in the next decade.
3.17. Eduard Rindfleisch
German pathologist and histologist Eduard Rindfleisch (1836–1908), who attained professorship in Bonn and in Würzburg, recognised the role of nerve connective tissue, which he called “neuroglia Virchovi”, especially during pathological processes. In his textbook of pathological histology, the third German edition of which was published in 1873, he dedicated the entire section to neuroglial cells (Figure 14A) [106]. Rindfleisch joined those researchers who discussed the question what known elements of the nervous tissue should be considered as the part of the neuroglia. He was convinced, that
As for the neuroglia of the medullary [white] substance, we may safely affirm that it comprises whatever elements are met with in the white matter, apart from nerve-fibres and blood-vessels; hence all the granules and nuclei in the white substance are neuroglia-cells and nuclei, so the soft fibrous-spongy material into which the nerve fibres are embedded is also neuroglia. Concerning the relation of these parts of neuroglia to each other, a maceration of fresh cerebral substance in 1/10 per cent of super-osmic acid for several days teaches that some of the cells are star-shaped and have numerous, finely branched processes. These cells are arranged at regular intervals and represent a network of connective tissue corpuscles in Virchow’s sense. Their finest processes form by numerous anastomoses the mentioned fibrous-spongy material, the actual nervous putty (Nervenkitt), so that it can be reasonably not mentioned as a special basic substance of this connective tissue.([106]; p. 570)
Rindfleisch, according to his preface to the first edition, “offered his new textbook to the student-youth of Germany”; therefore, his view on the neuroglia as a connective tissue containing star-shaped cells forming a network, had a big impact on a new generation of researchers dealing with the structure of the nervous tissue.
3.18. Franz Boll
German physiologist and histologist Franz Christian Boll (1849–1879) who held professorships in Genoa and Rome, is universally known as the discoverer of rhodopsin. In 1873, however, he published an extensive work describing in detail the structure and development of the brain and spinal cord [107]. The publication also contained results of his extensive examination of the nervous connective tissue.
Boll was convinced that the connective tissue, besides connective tissue fibres, also contained cells (Figure 14B) which were, as he pointed out, for the first time isolated, described, and illustrated in “their true form” by Deiters. However, he added, the Deiters name and the description of these cells did not appear in the writings of Henle and Merkel, Stieda, Gerlach, Kolliker, and Frommann, but the cells were described and illustrated by Jastrowitz, Golgi, and, to a certain extent, by Rindfleisch. For this reason, Boll was likely to continue to name these cells as the “Deiters’ cells” (Deiters’schen Zellen); this term was used for some time by other researchers simultaneously with the name “glial cells”. Today, however, by the term “Deiters’ cells”, or phalangeal cells, are named supporting cells of the Corti organ in the inner ear, which are not related to glial cells at all. Describing the glial cells in the ox spinal cord Boll noted that:
These cells are so peculiar that I cannot give them any analogue from the series of other known histological elements. Characteristic of these is the lack of a body or cell body. At first sight, some seem to imagine only a confused convolution of fine fibres, in the centre of which there is a nucleus-like structure. If, however, in most of these cells the actual cell-body is an absolutely vanishing minimum, the processes of the latter are all the more pronounced. Each of these cells emits a very large number of very long hair-thin processes, comparable to the fineness of connective tissue fibrils, of straight or only slightly tortuous course. The length of these fibrous processes is often very considerable; not infrequently, individuals are drawn through the entire field of view of the microscope and even further. Whether these fine processes ramify, I certainly do not want to decide; I have never received unambiguous pictures of such a place. On the other hand, on the basis of my experience, I do not necessarily dare deny ramification: in any case, such is one of the rarer occurrences. The direction of the processes is subject to countless differences: there are cells from which these fibres emanate multipolarly in all directions; besides, there are other cells where two dense fibrous bundles are detached on two opposing poles, and finally a third main form is not uncommon, where the whole mass of the fibrous processes is directed to one side, while from the opposite pole of the cell depart only very insignificant fibres, so that these cells offer a surprising similarity with a fine hair brush (brush cells—Pinselzellen).([107]; p. 7)
Thus Boll, in addition to the detailed description of glial cell as cells with the long hair-thin processes, also drew attention to the variety of shapes of the glial cells, and attempted to classify them. Boll further analysed the structure of the perivascular space. The presence of terminals of glial processes on the capillary walls before Boll, had already been indicated by Besser, Golgi, and Butzke (see above). Boll, however, described that capillaries in the brain and spinal cord of humans, cows, and sheep were surrounded by rows of “brush cells” of various sizes and shapes spreading numerous processes to the capillary walls. In addition, on the basis of the detailed study of the nerve tissue development, Boll was convinced that both glial and nerve cells were histologically distinct elementary parts of the CNS.
3.19. Hans Gierke
German anatomist Hans Paul Bernhard Gierke (1847–1886) attained professorship at the Imperial University of Tokyo and in Breslau (Wrocław). Already in 1882, he published his paper on the elements of the CNS, where he described nerve cells and neuroglia studied predominantly by the carmine method, which he, based on numerous experiments, considered to be the most suitable for tissue staining [108]. According to Gierke, “The skeleton for the nervous elements of the central organ, the network in which they are embedded, is composed of two different substances, the fibrillar connective tissue and Virchow’s so-called neuroglia, which is considered to be a subspecies of the reticular connective tissue.” [108]. He also included in his paper a detailed description and pictures of several forms of neuroglial cells (Figure 15), and discussed their relationship to nerve cells and fibres. In addition, Gierke was convinced that: “The neuroglia develops from elements of the ectoderm like the nerve cells and like the cells of the epidermis. Even this histogenetic fact speaks for a peculiar position of the neuroglia, even more so its appearance, its behaviour against reagents and the arrangement of the elements.” [108].
A year later, Gierke published an updated paper devoted to the supportive substance of the nervous system [110]. He described different types of neuroglial cells in the various regions of the brain and spinal cord, and at the same time, he moved forward in questioning what should be considered to be neuroglia:
The supporting substance consists of unformed and shaped elements, both of which must be grouped under the term Neuroglia. The unformed part, the basic substance (Grundsubstanz), has not yet received the due attention it deserves. It is the basis of the grey matter in which everything else is stored and quantitatively occupies about the third part of it…. The shaped elements of the supportive substance are cells and their processes.([110]; p.362)
In two years Gierke published results of the investigation of supportive substance of the CNS in two extensive publications [109,111]. He described in detail many modifications of his carmine staining method, which he still considered to be the best. Gierke made an attempt to clearly define the connective tissue and neuroglia. First of all, he was convinced that free grains observed in the connective tissue by Jastrowitz and Boll did not exist. Concerning the connective tissue, Gierke wrote:
The nervous elements of the central organs are not directly next to each other, but are separated from each other by a different mass, which can be named with a very indifferent name as a supporting substance (Stutzsubstanz) of the central nervous system. It is often called connective tissue (Bindegewebe) by the authors, and indeed regarded as “reticular connective tissue” (retikulären Bindesubstanz) in terms of its histological significance, and is presented as a subspecies of it in addition to the cytogenous connective substance of the mucous membranes and of many glands and the jelly-tissue… In any case, it is desirable not to name it that way any longer, but I shall name it in the following to have the shortest possible expression, with the name “neuroglia” (or nerve putty), or glia, introduced by Virchow. The supporting substance of the central organs consists of two components, the unformed and the shaped substance; both must be grouped together under the name of neuroglia, although of course the term “nerve putty” is less suitable for the cellular elements than for the matrix substance. When we shorten, as is customary, that word, we have in the terms “glial substance” and “glial cells” very convenient names, which I do not find at all like Ranvier in the above-quoted little work “detestable”.([109]; p. 452)
Gierke also tried to find out the general characteristics of the glial cells:
First, what is common to all glial cells? In most cases characteristic of them are the processes. Just as in the central nervous system, no nerve cells without processes occur, so are cells of the support substance without such unthinkable. In order to be able to take part in the formation of the framework for the embedded nervous elements, the glial cells must connect by means of processes with other similar elements. I can also say with the utmost certainty that there are absolutely no glial cells in the central nervous system, which are without processes, and which therefore lie isolated, without connection to the general scaffold. With regard to the number of processes, certain details are not to be specified. Most of the cells have very many, but even with very few occur. Cells with a single process, if they exist at all, are very rare; they might perhaps appear on the surface of the cerebral cortex as an exception. Bipolar cells occur in certain places, especially where long sutures are needed. The cell-body can then grow so much in the formation of the growing processes, that it only makes a small swelling in the middle of them, indeed one finds horn-threads in those places, which have no trace of a cell-body, but detectable in of the imaginary way. … Glial cells with three or very few processes proceeding to different sides occur in all parts of the central organ; yet they are much rarer than those with many processes.([109]; p. 466)
In general, due to the staining methods, which reached its borders, Gierke’s investigations did not bring extraordinary new information about the glial cells, if compared to the works of his predecessors. It became evident that the carmine staining has reached its limits, and in the following years, it withdrew its place in favour of the “black reaction” staining invented by Golgi.
4. Recapitulation
The notions of the passive elements of the CNS have emerged in the second half of 18th century, and at the beginning of 19th century. The idea of the brain connective tissue (Bindegewebs-Substanzen) was first introduced by Karel Rokytanský, and then it was developed by Rudolf Wirchow into the concept of neuroglia. In the course of the second half of the 19th century, many neuroanatomists dedicated their efforts to study neuroglia; as a result, numerous morphologically distinct cell types have been described. The turn of research toward neuroglia has been precipitated by the development of staining techniques which allowed much more precise morphological analysis of neural cells. This allowed several major types of neuroglia to be distinguished; in addition, the theories of glial roles in the physiology and pathophysiology of the nervous system began to evolve. Detailed descriptions of these studies will be covered in our papers that are to follow.
Author Contributions
All authors participated equally in writing this commentary.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Crivellato, E.; Ribatti, D. Soul, mind, brain: Greek philosophy and the birth of neuroscience. Brain Res. Bull. 2007, 71, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Clarke, E. Aristotelian concepts of the form and function of the brain. Bull. Hist. Med. 1963, 37, 1–14. [Google Scholar] [PubMed]
- Gross, C.G. Aristotle on the brain. Neuroscientist 1995, 1, 245–250. [Google Scholar] [CrossRef]
- Codellas, P.S. Alcmaeon of Croton: His life, work, and fragments. Proc. R. Soc. Med. 1932, 25, 1041–1046. [Google Scholar] [PubMed]
- Hippocrates. On the sacred disease. In The Medical Works of Hippocrates; Chadwick, J., Mann, W.N.T., Eds.; Blackwells: Oxford, UK, 1950; pp. 179–189. [Google Scholar]
- Longrigg, J. Greek Rational Medicine: Philosophy and Medicine from Alcmaeon to the Alexandrians; Routledge: New York, NY, USA, 1993. [Google Scholar]
- Von Staden, H. Herophilus: The Art of Medicine in Early Alexandria; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Wills, A. Herophilus, Erasistratus, and the birth of neuroscience. Lancet 1999, 354, 1719–1720. [Google Scholar] [CrossRef]
- Manzoni, T. The cerebral ventricles, the animal spirits and the dawn of brain localization of function. Arch. Ital. Biol. 1998, 136, 103–152. [Google Scholar] [PubMed]
- Pevsner, J. Leonardo da Vinci’s contributions to neuroscience. Trends Neurosci. 2002, 25, 217–220. [Google Scholar] [CrossRef]
- Reisch, G. Margarita Philosophica; Johann Schott: Freiburg, Germany, 1503. [Google Scholar]
- Fludd, R. Utriusque Cosmi Majoris Scilicet et Minoris Metaphysica, Physica Atque Technica Historia in duo Volumina Secundum Cosmi Differentiam Divisa. Tomus Primus de Macrososmi Historia in duo Tractatus Divisa; Oppenhemii, Aere Johan-Theodori de Bry, Typis Hieronymi Galleri: Frankfurt, Germany, 1617–1621. [Google Scholar]
- Da Vinci, L. Corpus of the Anatomical Studies in the Collection of Her Majesty, the Queen, at Windsor Castle; Harcourt Brace Jovanovich: Windsor, UK, 1978–1980. [Google Scholar]
- Descartes, R.; Schuyl, F. De Homine; Petrum Leffen, Franciscum Moyardum: Ludguni Batavorum (Leiden), The Netherlands, 1662. [Google Scholar]
- Malpighi, M. Opera Omnia; Royal SocietyTomis Duobus: London, UK, 1687. [Google Scholar]
- Willis, T. De Anima Brutorum Quae Hominis Vitalis ac Sentitiva est: Exercitationes Duae; Typis E.F. Impensis Ric. Davis: Oxon, UK, 1672. [Google Scholar]
- Combe, G. The Constitution of Man Considered in Relation to External Objects, 8th ed.; MacLahlan Steward & Co.: Edinburgh, UK, 1847. [Google Scholar]
- Gall, F.J. On the Functions of the Brain and Each of Its Parts: With Observations on the Possibility of Determining the Instincts, Propensities and Talents, or the Moral and Intellectual Dispositions of Men and Animals, by the Configuration of the Head, Trans, Winslow Lewis, Jr.; Marsh, Capen & Lyon: Boston, UK, 1835. [Google Scholar]
- Gall, F.J.; Spurzheim, J.G. Anatomie et Physiologie du Système Nerveux en Général et du Cerveau en Particulier Avec des Observations sur la Possibilité de Reconnaître Plusieurs Dispositions Intellectuelles et Morales de L’homme et des Animaux par la Configuration de Leurs Têtes, 4 Vols., with an Atlas of 100 Engraved Plates; Schoell: Paris, France, 1810–1819. [Google Scholar]
- Prochaska, G. Functions of the Nervous System (English Translation by T. LAYCOCK, 1851); Sydenham’s Society Series: London, UK, 1784. [Google Scholar]
- Broca, P. Perte de la parole, ramolissement chronique et destruction partielle du lobe anterieur gauche du cerveau (sur la siege de la faculte du langage). Bulletins et Mémoires de la Société D’anthropologie de Paris 1861, 2, 235–238. [Google Scholar]
- Fritsch, G.T.; Hitzig, E. Über die electrische erregbarkeit des grosshirns. Arch. Anat. Physiol. Wiss. Med. 1870, 37, 300–322. [Google Scholar]
- Ferrier, D. Experiments in the brain of monkeys. Proc. R. Soc. Lond. Ser. B. 1875, 23, 409–430. [Google Scholar] [CrossRef]
- Ferrier, D. The Functions of the Brain; Smith, Elder: London, UK, 1876. [Google Scholar]
- Ferrier, D. The Croonian Lectures on Cerebral Localisation; Smith, Elder & Co.: London, UK, 1890. [Google Scholar]
- Penfield, W.; Bouldrey, E. Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation. Brain 1937, 60, 389–443. [Google Scholar] [CrossRef]
- Descartes, R. L’homme, et un Traité de la Formation du Foetus du Mesme Autheur. Avec les Remarques de Louys de la Forge, 1st ed.; Nicolas Le Gras: Paris, France, 1664. [Google Scholar]
- Bidloo, G.; de Lairesse, G. Anatomia Humani Corporis; Joannis à Someren, Joannis à Dyk, Henrici & Theodori Boom: Amstelodami, The Netherlands, 1685. [Google Scholar]
- Leeuwenhoek, A.V. Alle de Briefen van Antoni van Leeuwenhoek/The Collected Letters of Antoni van Leeuwenhoek 1673–1696 (Eleven Volumes), Edited, Illustrated, and Annotated by Committees of Dutch Scientists; Swets & Zeitlinger: Amsterdam, The Netherlands, 1939–1983. [Google Scholar]
- Van Leeuwenhoek, A. Send-Brieven, Zoo aan de Hoog Edele Heeren van de Koninklyke Societeit te Londen; Adriaan Beman: Te Delft, The Netherlands, 1718. [Google Scholar]
- Von Haller, A. Elementa Physiologiae Corporis Humani, in 8 Volumes; Marci-Michael Bosquet & Sociorum: Lausanne, Switzerland, 1757–1778. [Google Scholar]
- Chvatal, A. Jiri Prochaska (1749–1820): Part 2: “De structura nervorum”—Studies on a structure of the nervous system. J. Hist. Neurosci. 2015, 24, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Von Haller, A. De Partibus Corporis Humani Sensilibus et Irritabilibus. Commentarii Societatis Regiae Scientiarum Gottingensis; Vadenhoek: Gottingen, Germany, 1753. [Google Scholar]
- Dierig, S. Neuronen-Doktrin und Neuroglia. PhD Thesis, Inaugural-Dissertation, University of Konstanz, Konstanz Germany, 1994. [Google Scholar]
- Hooke, R. Micrographia; Folio Society: London, UK, 2017. [Google Scholar]
- Shapiro, S. Antony van Leeuwenhoek; a review of his life and work. J. Biol. Photogr. Assoc. 1955, 23, 49–57. [Google Scholar] [PubMed]
- Schleiden, M.J. Beiträge zur phytogenesis. Arc. Anat. Physiol. Wiss. Med. 1838, 137–176. [Google Scholar]
- Schwann, T. Mikroskopische Untersuchungen über die Übereinstimmung in der Struktur und dem Wachstum der Tiere und Pflanzen; Sanderschen Buchhandlung: Berlin, Germany, 1839. [Google Scholar]
- Schwann, T. Microscopical Researches into the Accordance in the Structure and Growth of Animals and Plants; Sydenham Society: London, UK, 1847. [Google Scholar]
- Malpighi, M. De Cerebri Cortice. In De Viscerum Structura Exercitatio Anatomica; Giacomo Monti: Bologna, Italy, 1666. [Google Scholar]
- Gross, C.G. Emanuel Swedenborg: A neuroscientist before his time. Neuroscientist 1997, 3, 142–147. [Google Scholar] [CrossRef]
- Swedenborg, E. The Brain, Considered Anatomically, Physiologically and Phylosopically. (Translated and Edited by R.L. Tafel), in 4 Volumes; James Speirs: London, UK, 1882. [Google Scholar]
- Torre, G.M.D. Nuove Osservazioni Microscopiche; C. R. Somasco: Napoli, Italy, 1776. [Google Scholar]
- Bentivoglio, M. 1896–1996: The centennial of the axon. Brain Res. Bull. 1996, 41, 319–325. [Google Scholar] [CrossRef]
- Keuffel, G.G.T. Ueber das ruckenmark. Arch. Physiol. 1811, 10, 123–202. [Google Scholar]
- Dutrochet, M.H. Recherches Anatomiques et Physiologiques sur la Structure Intime des Animaux et des Végétaux, et sur leur Motilité; J.B.Bailliere: Paris, France, 1824. [Google Scholar]
- Dutrochet, M.H. Mémoires Pour Servir a L’historie Anatomique et Physiologique des Végétaux et des Animaux. II; J.-B. Bailliere: Paris, France, 1837; p. 688. [Google Scholar]
- Studnička, F.K. Aus der Vorgeschichte der Zellentheorie. H. Milne edwards, H. Dutrochet, F. Raspail, j.E. Purkinje. Anatomischer Anzeiger 1932, 73, 390–416. [Google Scholar]
- Ehrenberg, C.G. Beobachtungeiner Auffallenden Bisher Unerkannten Strukfurdes Seelenorgans bei Menschen und Thieren; Königlichen Akademie der Wissenschchaft: Berlin, Germany, 1836. [Google Scholar]
- Purkinje, J.E. Neueste Beobachtungen über die Struktur des Gehirns. In Opera omnia; Purkynova Spolecnost: Prague, Chec Republic, 1837; Volume 2, p. 3. [Google Scholar]
- Todd, R.B. The Descriptive and Physiological Anatomy of the Brain, Spinal Cord, Ganglions and Their Coverings; Sherwood, Gilbert and Piper: London, UK, 1845. [Google Scholar]
- Golgi, C. Opera Omnia; Hoepli: Milano, Italy, 1903. [Google Scholar]
- Tan, S.Y.; Brown, J. Rudolph virchow (1821–1902): “Pope of pathology”. Singap. Med. J. 2006, 47, 567–568. [Google Scholar]
- Keuffel, G.G.T. De Medulla Spinali. Doctoral Dissertation, Halae, Magdeburg, Germay, 1810. [Google Scholar]
- Weigert, C. Kenntnis der Normalen Menschlichen Neuroglia; Moritz Diesterweg: Frankfurt, Germany, 1895. [Google Scholar]
- Valentin, G. Über den verlauf und die letzten enden der nerven. Nova Acta Academiae Caesareae Leopoldino-Carolinae Germanicae Naturae Curiosorum. Verhandlungen der Kaiserlich Leopoldinisch-Carolinischen Deutschen Akademie der Naturforscher 1836, 18, 51–240. [Google Scholar]
- Chvatal, A. Discovering the structure of nerve tissue: Part 2: Gabriel Valentin, Robert Remak, and Jan Evangelista Purkyně. J. Hist. Neurosci. 2015, 24, 326–351. [Google Scholar] [CrossRef] [PubMed]
- Purkyně, J.E. Untersuchungen aus der Nerven-und Hirnanatomie. Isis 1838, 581–584. [Google Scholar]
- Arnold, F. Handbuch der Anatomie des Menschen; Verlag Herder: Freiburg, Germany, 1844. [Google Scholar]
- Rokitansky, C. Ueber die dendritischen Vegetationen auf Synovialhäuten. Zeitschrift der Kaiserlichen Königlichen Gesellschaft der Aerzte zu Wien 1851, 7, 1–8. [Google Scholar]
- Rokitansky, K. Über die Entwickelung der Krebsgerüste mit Hinblick auf das Wesen und die Entwickelung anderer Maschenwerke. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften. Mathematisch-Naturwissenschaftliche Classe. Wien 1852, 8, 391–405. [Google Scholar]
- Rokitansky, K. Über den Zottenkrebs. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften. Mathematisch-Naturwissenschaftliche Classe. Wien 1852, 8, 513–535. [Google Scholar]
- Rokitansky, K. Über das Auswachsen der Bindegewebs-Substanzen und die Beziehung desselben zur Entzündung. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften. Mathematisch-Naturwissenschaftliche Classe. Wien 1854, 13, 122–140. [Google Scholar]
- Rokitansky, K. Über Bindegewebs-Wucherung im Nervensysteme. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften. Mathematisch-Naturwissenschaftliche Classe. Wien 1857, 24, 517–536. [Google Scholar]
- Wright, N.A.; Poulsom, R. Omnis cellula e cellula revisited: Cell biology as the foundation of pathology. J. Pathol. 2012, 226, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Virchow, R. Ueber das granulirte Ansehen der Wandungen der Gehirnventrikel. Allgemeine Zeitschrift für Psychiatrie und Psychisch-Gerichtliche Medicin 1846, 3, 242–250. [Google Scholar]
- Rokitansky, C. Handbuch der Pathologischen Anatomie. II. Band; Braumüller & Seidel: Wien, Austria, 1844. [Google Scholar]
- Virchow, R. Ueber das granulirte Ansehen der Wandungen der Gehirnventrikel. In Gesammelte Abhandlungen zur Wissenschaftlichen Medicin.; Virchow, R., Ed.; Meidinger Sohn & Comp.: Frankfurt, Germany, 1856; pp. 885–891. [Google Scholar]
- Virchow, R. Die Cellularpathologie in Ihrer Begründung auf Physiologische und Pathologische Gewebelehre 20 Vorlesungen, Gehalten Während d. Monate Febr., März u. April 1858 im Patholog. Inst. Zu Berlin; August Hirschwald: Berlin, Germany, 1858. [Google Scholar]
- Virchow, R.L.K. Cellular pathology; John Churchill: London, UK, 1860; p. 511. [Google Scholar]
- Bidder, F.H.; Kupffer, C. Untersuchungen über die Textur des Rückenmarks und die Entwicklung Seiner Formelemente; Breikopf und Härtel: Leipzig, Germany, 1857; p. 151. [Google Scholar]
- Kupffer, C. De Medullae Spinalis Textura in Ranis Ratione Imprimis Habita Indolis Substantiae Cinereae; Typis Viduae, J.C. Schünmanni et C. Mattiesseni: Dorpati Livonorum, 1854. [Google Scholar]
- Pappenheim, S. Die Specielle Gewebelehre des Auges mit Rücksicht auf Entwicklungsgeschichte und Augenpraxis; Georg Philipp Aderholz: Breslau, Poland, 1842. [Google Scholar]
- Müller, H. Zur Histologie der Netzhaut. Zeitschrift für Wissenschaftliche Zoologie 1851, 3, 234–237. [Google Scholar]
- Schulze, M. Observationes de Retinae Structura Penitiori; Published Lecture at the Univeesity of Bonn: Bonn, Germany, 1859. [Google Scholar]
- Müller, H. Anatomisch-Physiologische Untersuchungen über die Retina des Menschen und der Wirbelthiere; Wilhelm Engelmann: Leipzig, Germany, 1856. [Google Scholar]
- Bergmann, C. Notiz über einige Structurverhältnisse des Cerebellum und Rückenmarks. Zeitschrift für Rationelle Medicin 1857, 8, 360–363. [Google Scholar]
- Mauthner, L. Über die Sogenannten Bindegewehskörperchen des Centralen Nervensystems. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften. Mathematisch-Naturwissenschaftliche Classe. Wien 1861, 43, 45–54. [Google Scholar]
- Beale, L. New Observations upon the Structure and Formation of Certain Nervous Centres, Tending to Prove That the Cells and Fibres of Every Nervous Apparatus form an Uninterrupted Circuit; John Churchill and Sons: London, UK, 1864. [Google Scholar]
- Frommann, C. Ueber die färbung der Binde- und Nervensubstanz des Rückenmarkes durch Argentum nitricum und über die Struktur der Nervenzellen. Archiv für Pathologische Anatomie und Physiologie und für Klinische Medicin 1864, 31, 129–153. [Google Scholar] [CrossRef]
- Frommann, C. Untersuchungen über die Normale und Pathologische Anatomie des Rückenmarks. Zweiter Theil; Friedrich Frommann: Jena, Germany, 1867. [Google Scholar]
- Deiters, O. Untersuchungen über Gehirn und Rückenmark des Menschen und der Säugethiere; Vieweg: Braunschweig, Germany, 1865. [Google Scholar]
- Besser, L. Zur Histogenese der nervösen Elementartheile in den Centralorganen des neugebornen Menschen. Archiv für Pathologische Anatomie und Physiologie und für Klinische Medicin 1866, 36, 305–334. [Google Scholar] [CrossRef]
- Chvatal, A. Discovering the structure of nerve tissue: Part 3: From Jan Evangelista Purkyne to Ludwig Mauthner. J. Hist. Neurosci. 2017, 26, 15–49. [Google Scholar] [CrossRef] [PubMed]
- Kölliker, A. Handbuch der Gewebelehre des Menschen für ärtze und Studirende, 5th ed.; Engelmann: Leipzig, Germany, 1867. [Google Scholar]
- Meynert, T. Vom Gehirne der Säugethiere. In Handbuch der Lehre von den Geweben des Menschen und der Thiere. Zweiter Band; Stricker, S., Ed.; Wilhelm Engelmann: Leipzig, Germany, 1872; pp. 694–809. [Google Scholar]
- Meynert, T. The brain of mammals. In A Manual of Histology; Stricker, S., Ed.; William Wood & Company: New York, NY, USA, 1872; pp. 650–766. [Google Scholar]
- Meynert, T. Der bau der Gross-Hirnrinde und seine örtlichen Verschiedenheiten, nebst einem pathologisch-anatomischen Corollarium. I. Vierteljahrsschrift für Psychiatrie in ihren Beziehungen zur Morphologie und Pathologie des Central-Nervensystems der physiologischen Psychologie. Statistik und Gerichtlichen Medicin 1867, 1, 77–124. [Google Scholar]
- Arndt, R. Studien über die Architektonik der Grosshirnrinde des Menschen. III. Archiv für Mikroskopische Anatomie 1869, 5, 317–331. [Google Scholar] [CrossRef]
- Stieda, L. Studien uer das Centrale Nervensystem der Wirbelthiere. Zeitschrift für Wissenschaftliche Zoologie 1869, 20, 273–456. [Google Scholar]
- Henle, J.; Merkel, F. Uber die sogenannte Bindesubstanz der Centralorgane des Nervensystems. Z. Med. 1869, 34, 49–82. [Google Scholar]
- Arndt, R. Studien über die Architektonik der Grosshirnrinde des Menschen. Archiv für Mikroskopische Anatomie 1867, 3, 441–476. [Google Scholar] [CrossRef]
- Arndt, R. Studien über die Architektonik der Grosshirnrinde des Menschen. II. Archiv für Mikroskopische Anatomie 1868, 4, 407–526. [Google Scholar] [CrossRef]
- Stieda, L. Studien über das Centrale Nervensystem der Wirbelthiere; Wilhelm Engelmann: Leipzig, Germany, 1870. [Google Scholar]
- Golgi, C. Sulla sostanza connettiva del cervello. Gazzetta Medica Italiana, Lombardia 1870, 19, 145–146. [Google Scholar]
- Golgi, C. Ueber die Bindesubstanz im Gehirne. Schmidt’s Jahrbücher der in-und Ausländischen Gesammten Medicin 1871, 150, 3–4. [Google Scholar]
- Golgi, C. Contribuzione alla fina anatomia degli organi centrali del sistema nervoso. I. Rivista Clinica di Bologna 1871, 1, 338–350. [Google Scholar]
- Golgi, C. Contribuzione alla fina anatomia degli organi centrali del sistema nervoso. II. Rivista Clinica di Bologna 1871, 1, 371–380. [Google Scholar]
- Golgi, C. Contribuzione alla fina anatomia degli organi centrali del sistema nervoso. III. Rivista Clinica di Bologna 1872, 2, 38–46. [Google Scholar]
- Jastrowitz, M. Studien über die Encephalitis und Myelitis des ersten Kindesalters. Archiv für Psychiatrie und Nervenkrankheiten 1871, 3, 162–213. [Google Scholar] [CrossRef]
- Golgi, C. Sulla struttura della sostanza grigia del cervello. Gazzetta Medica Italiana, Lombardia 1873, 6, 244–246. [Google Scholar]
- Jastrowitz, M. Encephalitis und Myelitis des ersten Kindersalters. Archiv für Psychiatrie und Nervenkrankheiten 1870, 2, 389–414. [Google Scholar] [CrossRef]
- Butzke, V. Studien uber den feineren Bau der Grosshirnrinde. Archiv für Psychiatrie und Nervenkrankheiten 1872, 3, 575–600. [Google Scholar] [CrossRef]
- Ranvier, L.A. De la névroglie. Archives de Physiologie Normale et Pathologique 1883, 3 S1, 177–185. [Google Scholar]
- Ranvier, L.A. Sur les éléments conjonctifs de la moelle épinière. Comptes Rendus Hebdomadaires des Séances de l’Académie des Sciences 1873, 77, 1299–1302. [Google Scholar]
- Rindfleisch, E. Lehrbuch der Pathologischen Gewebelehre zur Eeinführung in das Studium der Pathologischen Anatomie; Wilhelm Engelmann: Leipzig, Germany, 1873. [Google Scholar]
- Boll, F. Die Histiologie und Histiogenese der nervösen Centralorgane. Archiv für Psychiatrie und Nervenkrankheiten 1873, 4, 1–138. [Google Scholar] [CrossRef]
- Gierke, H. Beiträge zur Kenntniss der Elemente des centralen Nervensystems. Breslauer Aerztliche Zeitschrift 1882, 4, 157–160, 172–177. [Google Scholar]
- Gierke, H. Die Stützsubstanz des Centralnervensystems. I. Theil. Archiv für Mikroskopische Anatomie 1885, 25, 441–554. [Google Scholar] [CrossRef]
- Gierke, H. Die Stützsubstanz des Centralenervensystems. Neurologisches Centralblatt 1883, 2, 361–369, 385–392. [Google Scholar]
- Gierke, H. Die Stützsubstanz des Centralnervensystems. II. Theil. Archiv für Mikroskopische Anatomie 1886, 26, 129–228. [Google Scholar] [CrossRef]
Figure 1.
Founders of neuroanatomy and neurophysiology: (A) Herophilius, (B) Earsistratus, and (C) Galen, and the frontispiece of the Renaissance edition of Galen’s works (Galeno. Omnia quae extant opera. Venezia, eredi di Luca Antonio Giunta, 1556).
Figure 1.
Founders of neuroanatomy and neurophysiology: (A) Herophilius, (B) Earsistratus, and (C) Galen, and the frontispiece of the Renaissance edition of Galen’s works (Galeno. Omnia quae extant opera. Venezia, eredi di Luca Antonio Giunta, 1556).

Figure 2.
Pneumo-ventricular doctrine of the brain function. (A) The conceptual scheme of ventricular localisation of brain functions proposed by Albertus Magnus, as depicted in the later version from the 16th century, made by Gregorius Reisch, who was the Prior of the House of Carthusians at Freiburg and confessor to the Emperor Maximilian, and who published a concise encyclopaedia of knowledge in 1503 [11]. (B) Ventricular divisions of brain functions as presented in ‘The Art of Memory’ by English Paracelcian physician Robert Fludd [12]. (C) The three-dimensional depiction of the ventricular system as drawn by Leonardo da Vinci. Leonardo made a mould of the brain ventricles by injecting them with melted wax, which after the removal of soft tissues produced their precise cast [13]. “Make two vent-holes in the horns of the greater ventricles, and insert melted wax with a syringe, making a hole in the ventricle of memory; and through such a hole, fill the three ventricles of the brain. Then when the wax has set, take apart the brain, and you will see the shape of the ventricles exactly.” (cited from [10]). (D) The concept of brain functions and cross-section of the human head and cranial nerves according to Leonardo da Vinci. Since Leonardo used his left hand for writing and wrote from right to left, his drawings with descriptions are presented mirrored in this figure. On the left is a cross-section of the head with a symbolic image of three ventricles; it is possible to recognize the names of the individual layers, such as the cerebro (brain), dura mater, and pia mater (meninges), the cranio (skull), and the pericranio (periosteum). On the right is the “cell” theory of brain functions and the cranial nerves. In the first ventricle, Leonardo placed imprensiva, a term proposed by himself (and never used by anybody else) for a structure connecting the senses and senso commune; in the second ventricle, he placed senso comune; and in the third, he placed memo (memory) [10].
Figure 2.
Pneumo-ventricular doctrine of the brain function. (A) The conceptual scheme of ventricular localisation of brain functions proposed by Albertus Magnus, as depicted in the later version from the 16th century, made by Gregorius Reisch, who was the Prior of the House of Carthusians at Freiburg and confessor to the Emperor Maximilian, and who published a concise encyclopaedia of knowledge in 1503 [11]. (B) Ventricular divisions of brain functions as presented in ‘The Art of Memory’ by English Paracelcian physician Robert Fludd [12]. (C) The three-dimensional depiction of the ventricular system as drawn by Leonardo da Vinci. Leonardo made a mould of the brain ventricles by injecting them with melted wax, which after the removal of soft tissues produced their precise cast [13]. “Make two vent-holes in the horns of the greater ventricles, and insert melted wax with a syringe, making a hole in the ventricle of memory; and through such a hole, fill the three ventricles of the brain. Then when the wax has set, take apart the brain, and you will see the shape of the ventricles exactly.” (cited from [10]). (D) The concept of brain functions and cross-section of the human head and cranial nerves according to Leonardo da Vinci. Since Leonardo used his left hand for writing and wrote from right to left, his drawings with descriptions are presented mirrored in this figure. On the left is a cross-section of the head with a symbolic image of three ventricles; it is possible to recognize the names of the individual layers, such as the cerebro (brain), dura mater, and pia mater (meninges), the cranio (skull), and the pericranio (periosteum). On the right is the “cell” theory of brain functions and the cranial nerves. In the first ventricle, Leonardo placed imprensiva, a term proposed by himself (and never used by anybody else) for a structure connecting the senses and senso commune; in the second ventricle, he placed senso comune; and in the third, he placed memo (memory) [10].
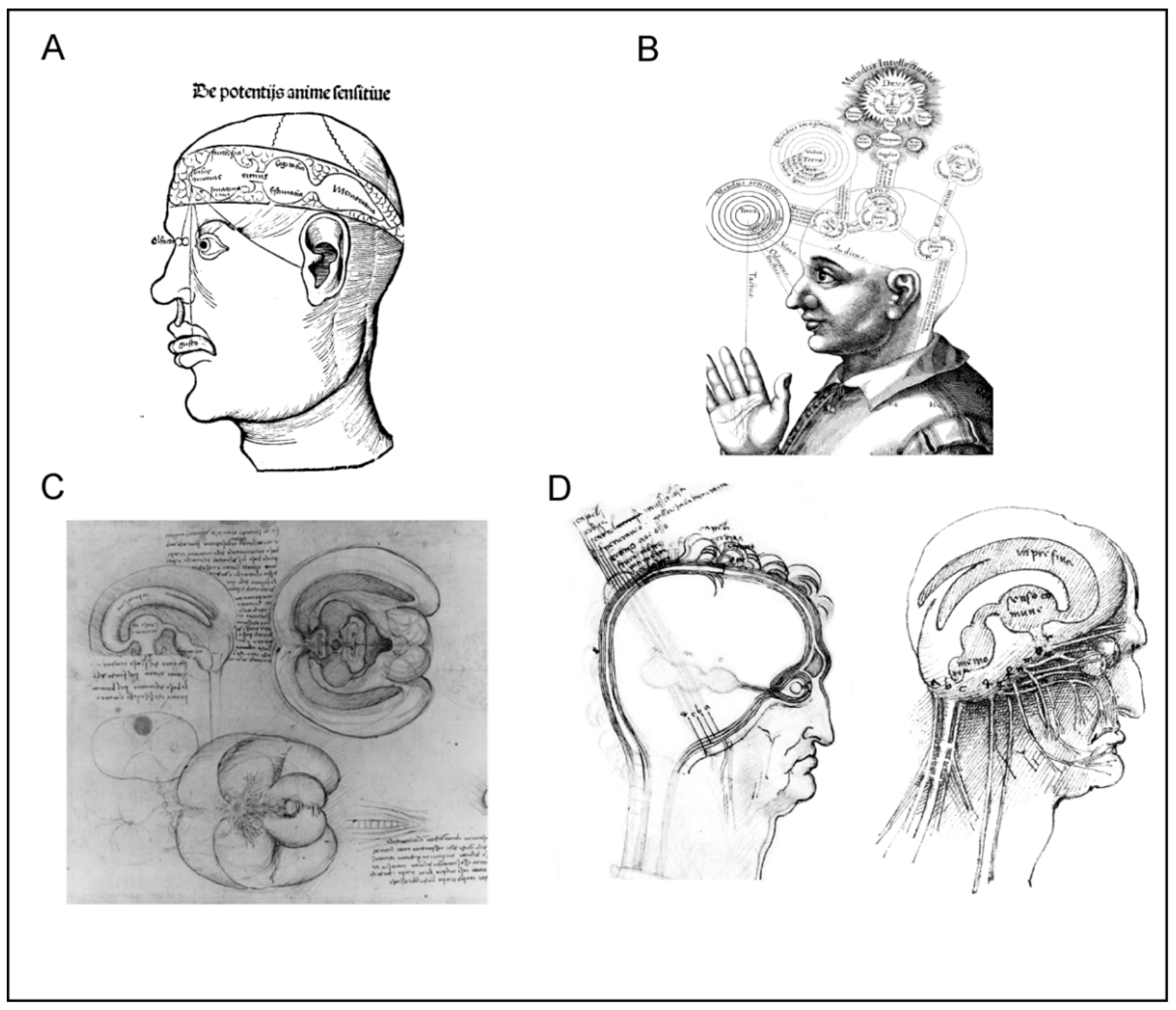
Figure 3.
Early morphological studies of the nervous system. (A) Nervous structure and the formation of voluntary movements as proposed by René Descartes. On the left is a diagram of the optic nerve comprising hollow tube-like structures [14]. On the right is the formation of voluntary movements regulated by the activity of the pineal gland [27]. (B) Albrecht von Haller, and the excerpt of his book De partibus corporis humani sensilibus et irritabilibus. Commentarii societatis regiae scientiarum gottingensis [26] in which he introduces the existence of irritable (irritabilis) and sensible (sensibilis) tissues. (C) Globular (gland-like) structures of the brain cortex according to Marcello Malpighi (reproduced from [28]). (D) Cross-section of a nerve (left, [29]) and the structure of the brain tissue after Antoni van Leeuwenhoek. On the right is a structure of dried pig brain tissue containing two vesicles [30].
Figure 3.
Early morphological studies of the nervous system. (A) Nervous structure and the formation of voluntary movements as proposed by René Descartes. On the left is a diagram of the optic nerve comprising hollow tube-like structures [14]. On the right is the formation of voluntary movements regulated by the activity of the pineal gland [27]. (B) Albrecht von Haller, and the excerpt of his book De partibus corporis humani sensilibus et irritabilibus. Commentarii societatis regiae scientiarum gottingensis [26] in which he introduces the existence of irritable (irritabilis) and sensible (sensibilis) tissues. (C) Globular (gland-like) structures of the brain cortex according to Marcello Malpighi (reproduced from [28]). (D) Cross-section of a nerve (left, [29]) and the structure of the brain tissue after Antoni van Leeuwenhoek. On the right is a structure of dried pig brain tissue containing two vesicles [30].
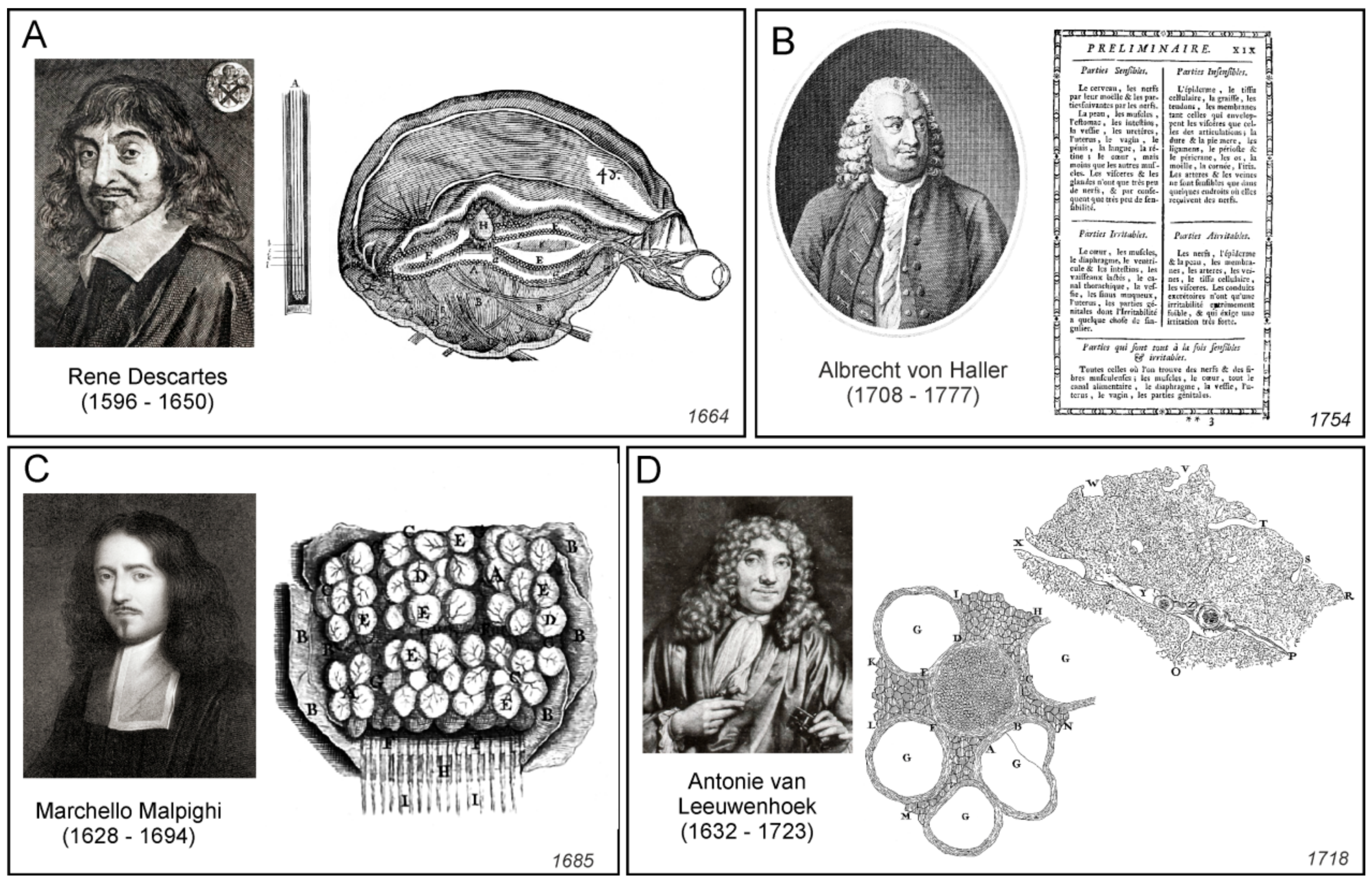
Figure 4.
The early studies of the elements of the nervous system. (A) Globular structure of the spinal cord tissue according to Giovanni Maria Della Torre, and the frontispiece of his book [36]. (B) Henri Dutrochet and his drawings of the globular corpuscules from mollusc cervical ganglia [38]. (C) Christian Gottfried Ehrenberg and his image of the nerve cell of the leech [41]. (D) Theodor Keuffel and the excerpt of his book on the structure of the spinal cord [45]. Right: the fibrous substance in the thin slice of ox spinal cord (top) and in the grey matter only (bottom) Top: a, b, c, d—the pia mater; f, g—two fibres extending from the pia mater and splitting into two centres (i); h—neurilem. Bottom: two centres of the grey matter; a—the end of the extension of the pia mater; b—the last fibrils into which this extension splits; c—the two centres of grey matter.
Figure 4.
The early studies of the elements of the nervous system. (A) Globular structure of the spinal cord tissue according to Giovanni Maria Della Torre, and the frontispiece of his book [36]. (B) Henri Dutrochet and his drawings of the globular corpuscules from mollusc cervical ganglia [38]. (C) Christian Gottfried Ehrenberg and his image of the nerve cell of the leech [41]. (D) Theodor Keuffel and the excerpt of his book on the structure of the spinal cord [45]. Right: the fibrous substance in the thin slice of ox spinal cord (top) and in the grey matter only (bottom) Top: a, b, c, d—the pia mater; f, g—two fibres extending from the pia mater and splitting into two centres (i); h—neurilem. Bottom: two centres of the grey matter; a—the end of the extension of the pia mater; b—the last fibrils into which this extension splits; c—the two centres of grey matter.

Figure 5.
First microscopic images of neural cells. (A) The first drawing of neurones with processes and nervous structures, made by Gustav Gabriel Valentin [48]. Middle: single globule of the human yellow matter, terminated by a tail-like process: (a) parenchyma, (b) tail-like process, (c) vesicle-like core similar to the germinal vesicle, and (d) bodies present on the surface. Right: fibres and globules in the sheep spinal cord. A thin section of the spinal cord in the cervical region, medulla oblongata: (a) the undulated longitudinal primitive fibres forming plexuses and (b) globules present in the interstitial space. (B) Jan Evangelista Purkyně and the first drawings of the Purkinje neurone made by him for the Congress of Physicians and Scientists Conference in Prague, in 1837. (C) Karl Kupffer [53] and the first drawing of the cellular elements in the frog spinal cord grey matter, which had “definite connections with undoubted connective tissue fibres”.
Figure 5.
First microscopic images of neural cells. (A) The first drawing of neurones with processes and nervous structures, made by Gustav Gabriel Valentin [48]. Middle: single globule of the human yellow matter, terminated by a tail-like process: (a) parenchyma, (b) tail-like process, (c) vesicle-like core similar to the germinal vesicle, and (d) bodies present on the surface. Right: fibres and globules in the sheep spinal cord. A thin section of the spinal cord in the cervical region, medulla oblongata: (a) the undulated longitudinal primitive fibres forming plexuses and (b) globules present in the interstitial space. (B) Jan Evangelista Purkyně and the first drawings of the Purkinje neurone made by him for the Congress of Physicians and Scientists Conference in Prague, in 1837. (C) Karl Kupffer [53] and the first drawing of the cellular elements in the frog spinal cord grey matter, which had “definite connections with undoubted connective tissue fibres”.
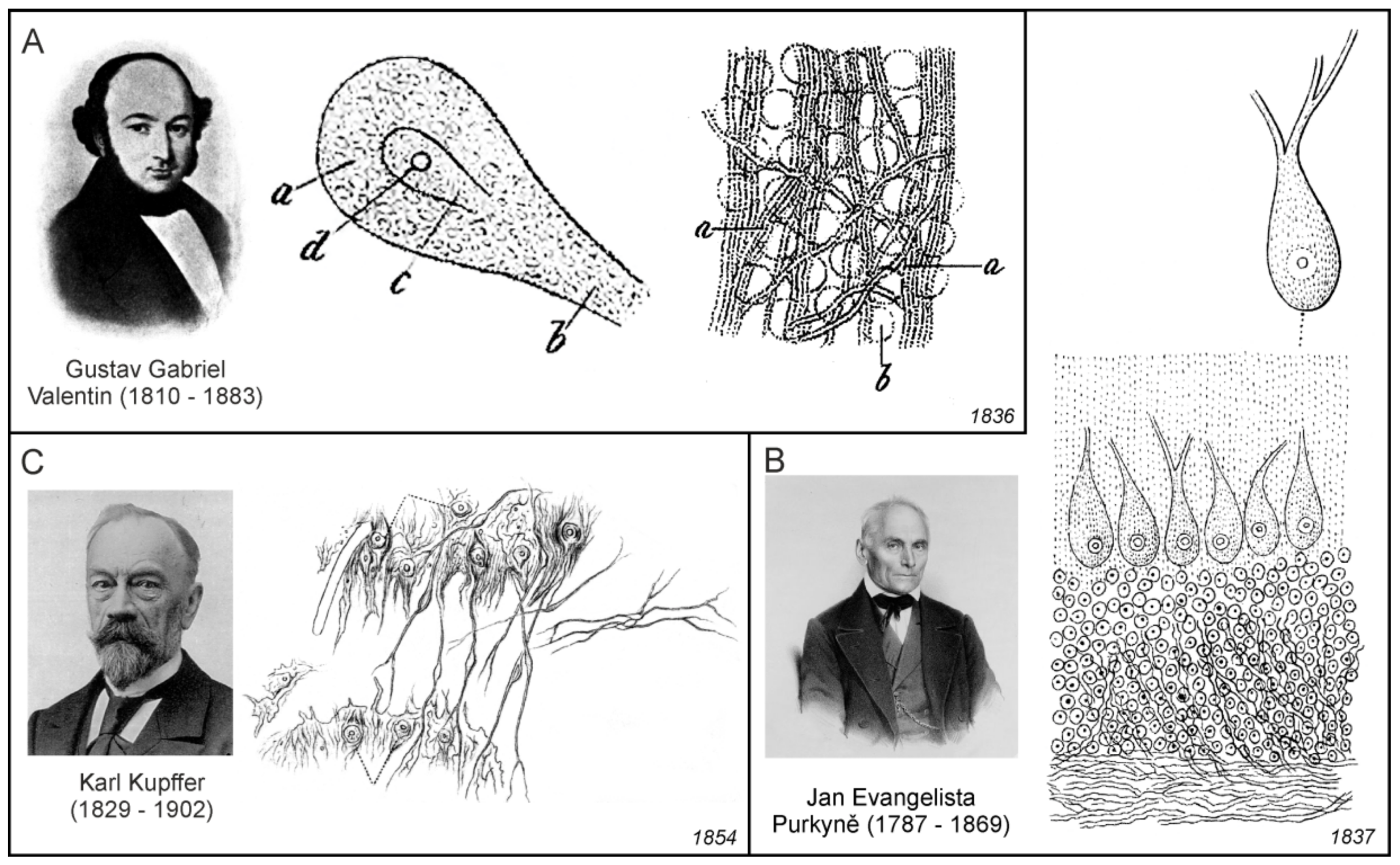
Figure 6.
Rudolf Virchow and the concept of neuroglia. (A) Rudolf Virchow, the frontispiece of his book Die cellularpathologie in ihrer begründung auf physiologische und pathologische [69,70] and the title of lecture 13, in which the concept of neuroglia was presented to the students of the Medical University of Berlin (Charite) on 3 April 1858. (B) Top: ependyma and neuroglia in the floor of the fourth ventricle. Between the ependyma and the nerve fibres is “the free portion of the neuroglia with numerous connective tissue corpuscles and nuclei”. Numerous corpora amylacea are also visible, shown enlarged below the main illustration (ca). E—ependymal epithelium; N—nerve fibres; v–w—blood vessels. Bottom: elements of neuroglia from the white matter of the human cerebral hemispheres. A—free nuclei with nucleoli; b—nuclei with partially destroyed cell bodies; c—complete cells. Reproduced from [69].
Figure 6.
Rudolf Virchow and the concept of neuroglia. (A) Rudolf Virchow, the frontispiece of his book Die cellularpathologie in ihrer begründung auf physiologische und pathologische [69,70] and the title of lecture 13, in which the concept of neuroglia was presented to the students of the Medical University of Berlin (Charite) on 3 April 1858. (B) Top: ependyma and neuroglia in the floor of the fourth ventricle. Between the ependyma and the nerve fibres is “the free portion of the neuroglia with numerous connective tissue corpuscles and nuclei”. Numerous corpora amylacea are also visible, shown enlarged below the main illustration (ca). E—ependymal epithelium; N—nerve fibres; v–w—blood vessels. Bottom: elements of neuroglia from the white matter of the human cerebral hemispheres. A—free nuclei with nucleoli; b—nuclei with partially destroyed cell bodies; c—complete cells. Reproduced from [69].

Figure 7.
(A). Probably the first image of radial astrocytes of the retina or Muller glial cells made by Samuel Pappenheim (a—the inner part, b—part of the retina, e—an outer part with a large spherical base changing into the fibre (c) and into the ending (d), [73], the panel also shows the frontispiece of his book Die specielle gewebelehre des auges mit rücksicht auf entwicklungsgeschichte und augenpraxis). (B): Radial astrocytes of the retina by Heinrich Müller [74]. (C): Radial astrocytes of the retina by Max Schultze [75]. The processes of Müller fibres passing into the outer nuclear layer (s), inner membrane (x) and the holes in membrana limitans interna (a), the holes in the network of fine membranes for optic nerves (b), so-called molecular network (c), the nuclei in Müller fibres (d), the space with the cell nuclei of the inner nuclear layer (e).
Figure 7.
(A). Probably the first image of radial astrocytes of the retina or Muller glial cells made by Samuel Pappenheim (a—the inner part, b—part of the retina, e—an outer part with a large spherical base changing into the fibre (c) and into the ending (d), [73], the panel also shows the frontispiece of his book Die specielle gewebelehre des auges mit rücksicht auf entwicklungsgeschichte und augenpraxis). (B): Radial astrocytes of the retina by Heinrich Müller [74]. (C): Radial astrocytes of the retina by Max Schultze [75]. The processes of Müller fibres passing into the outer nuclear layer (s), inner membrane (x) and the holes in membrana limitans interna (a), the holes in the network of fine membranes for optic nerves (b), so-called molecular network (c), the nuclei in Müller fibres (d), the space with the cell nuclei of the inner nuclear layer (e).

Figure 8.
First glioanatomists and their contributions. (A) Carl Bergmann and the excerpt of his paper [77] describing the anatomy of the cerebellum, including radial glia (Bergmann glial cells). (B) Ludwig Mauthner and his paper on the connective tissue of the brain [78]. (C) Lionel Beale and his book: New observations upon the structure and formation of certain nervous centres [79]. (D) The paper of Carl Frommann [80] and his images of stained neuroglia from the surface of the spinal cord longitudinal cut.
Figure 8.
First glioanatomists and their contributions. (A) Carl Bergmann and the excerpt of his paper [77] describing the anatomy of the cerebellum, including radial glia (Bergmann glial cells). (B) Ludwig Mauthner and his paper on the connective tissue of the brain [78]. (C) Lionel Beale and his book: New observations upon the structure and formation of certain nervous centres [79]. (D) The paper of Carl Frommann [80] and his images of stained neuroglia from the surface of the spinal cord longitudinal cut.

Figure 9.
Otto Deiters and his drawings of connective tissue cells (seemingly astrocyte, middle top—from the white matter, middle bottom—from the hypoglossal nucleus grey matter) and (at the right) neurone [82].
Figure 9.
Otto Deiters and his drawings of connective tissue cells (seemingly astrocyte, middle top—from the white matter, middle bottom—from the hypoglossal nucleus grey matter) and (at the right) neurone [82].
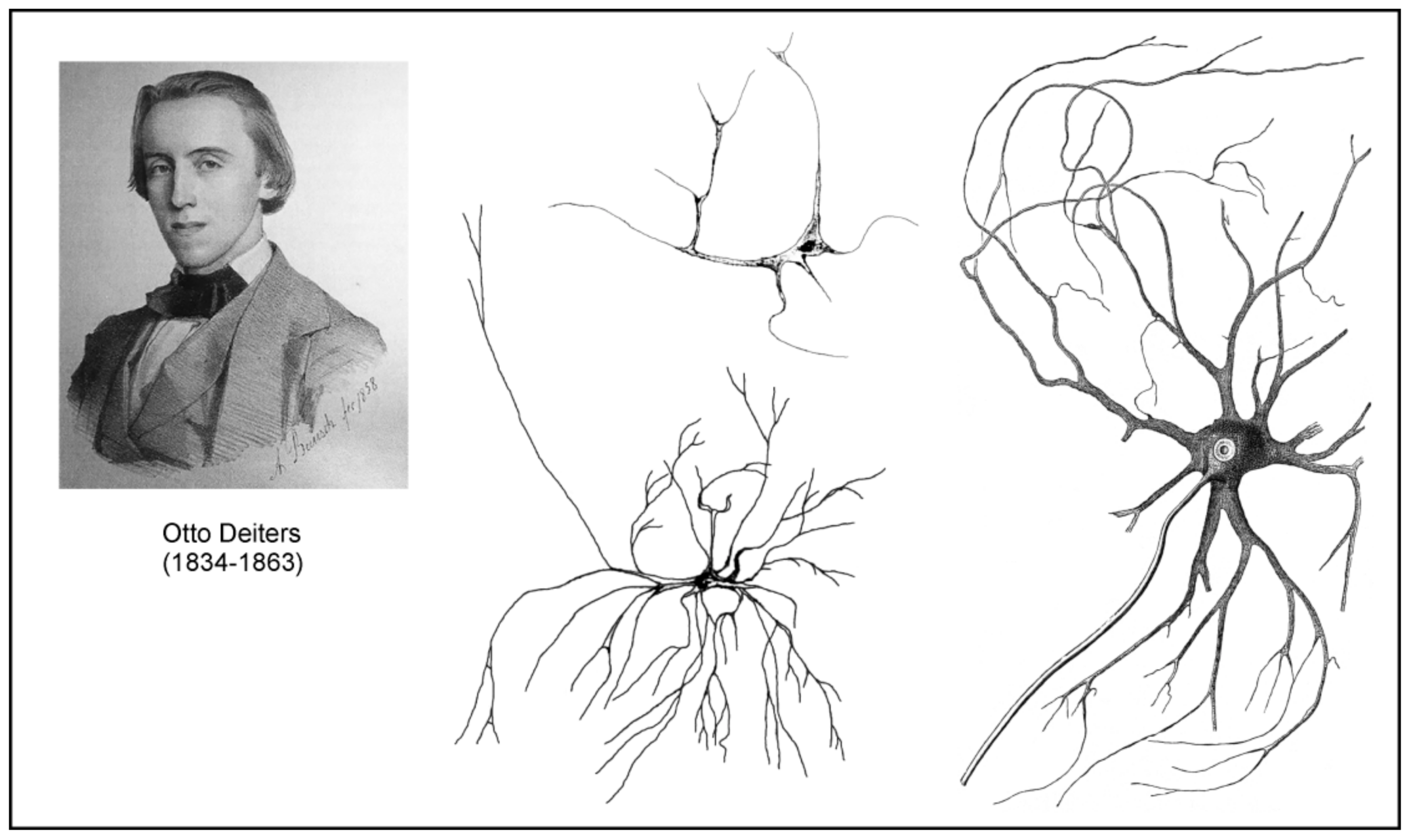
Figure 10.
(A): Images of connective tissue cells made by Leopold Besser, and the title page of his paper [83]. Top: vascular neoplasms in the brain of the newborn showing the glial formations located on the blood vessels. Bottom: formations on the cut of the neonatal cortex (left), including normal and damaged “glial nuclei” (1) during the cutting process; enlarged “glial nucleus” (right). (B): Images of connective tissue made by Albert Kölliker [85]. Middle: part of the connective tissue reticulum from the dorsal columns of the spinal cord, in two places the finest mesh forming reticulum is shown. Right: connective tissue cells in the grey matter of the human brain.
Figure 10.
(A): Images of connective tissue cells made by Leopold Besser, and the title page of his paper [83]. Top: vascular neoplasms in the brain of the newborn showing the glial formations located on the blood vessels. Bottom: formations on the cut of the neonatal cortex (left), including normal and damaged “glial nuclei” (1) during the cutting process; enlarged “glial nucleus” (right). (B): Images of connective tissue made by Albert Kölliker [85]. Middle: part of the connective tissue reticulum from the dorsal columns of the spinal cord, in two places the finest mesh forming reticulum is shown. Right: connective tissue cells in the grey matter of the human brain.

Figure 11.
(A) Images of connective tissue cells located between the nerve fibres, drawn by Theodor Meynert [88], (B) Brain cortex neurones embedded in the fine-grained connective tissue, by Rudolf Arndt [89]. (C) Brain cortex neurones embedded in the fine-grained connective tissue, by Ludwig Stieda [90]. (D) A surface layer containing fibres of white matter with visible axons and multipolar connective tissue cells, by Jakob Henle and Friedrich Merkel [91].
Figure 11.
(A) Images of connective tissue cells located between the nerve fibres, drawn by Theodor Meynert [88], (B) Brain cortex neurones embedded in the fine-grained connective tissue, by Rudolf Arndt [89]. (C) Brain cortex neurones embedded in the fine-grained connective tissue, by Ludwig Stieda [90]. (D) A surface layer containing fibres of white matter with visible axons and multipolar connective tissue cells, by Jakob Henle and Friedrich Merkel [91].
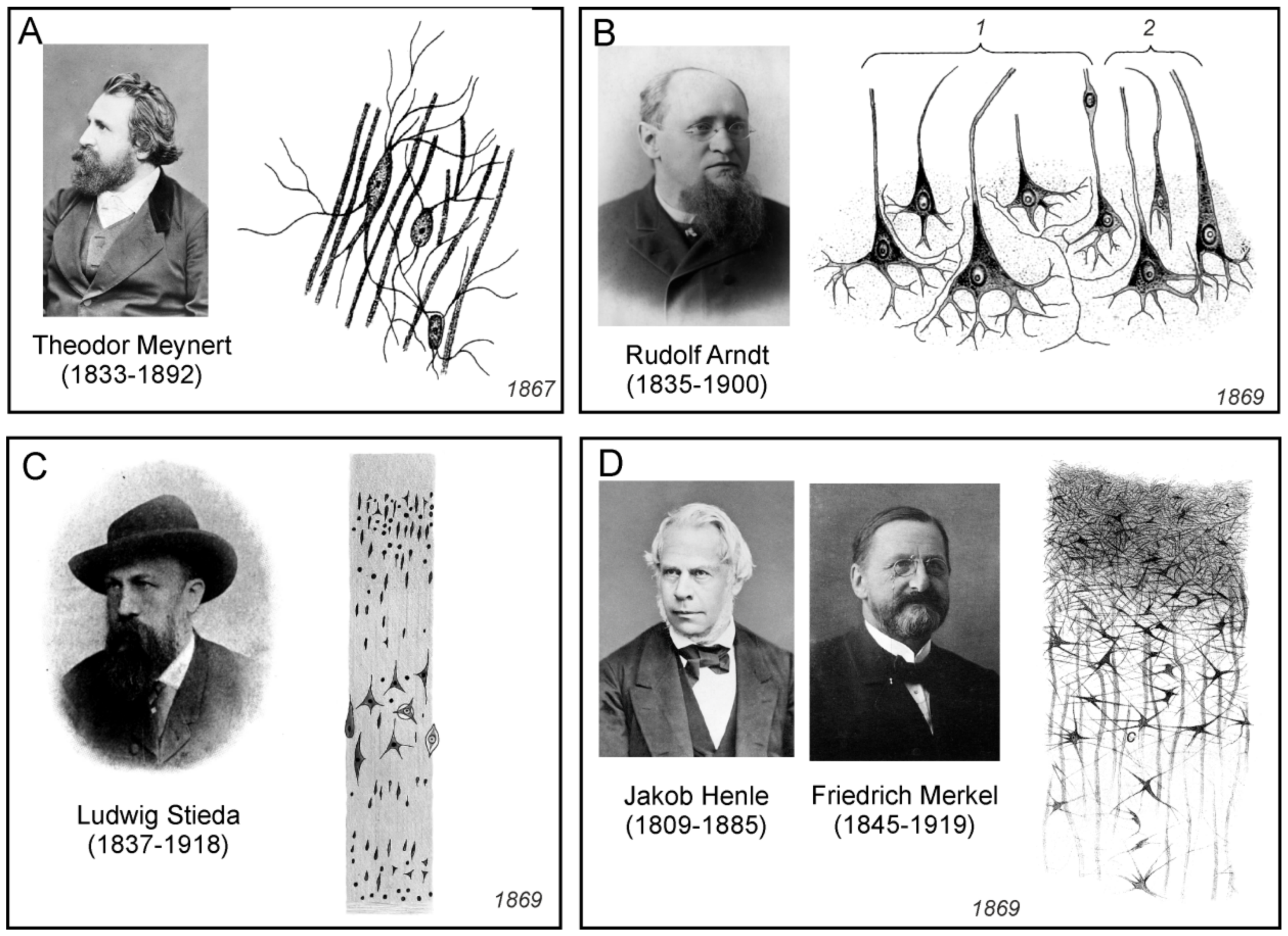
Figure 12.
(A) Images of connective tissue cell made by Camillo Golgi [96]. Middle: spinal cord cell with long longitudinal processes in deeper layers of white matter. Right: cross section of the capillary showing the connective cell processes closely adjacent to its contour. (B) Images of glial cells by Moritz Jastrowitz [100]. Isolated spider-like glial cells from the vicinity of the human brain ventricle ependym.
Figure 12.
(A) Images of connective tissue cell made by Camillo Golgi [96]. Middle: spinal cord cell with long longitudinal processes in deeper layers of white matter. Right: cross section of the capillary showing the connective cell processes closely adjacent to its contour. (B) Images of glial cells by Moritz Jastrowitz [100]. Isolated spider-like glial cells from the vicinity of the human brain ventricle ependym.

Figure 13.
(A) Images of glial cell with richly branched processes (middle) and connective tissue cell (right) drawn by Victor Butzke [103]. (B) Images of glial cells and neurons drawn by Louis-Antoine Ranvier [104]. Left: neuroglial cell of the lateral cords of the ox spinal cord. Middle: elements of the cortical layer of the cat’s brain; a, b, c, and d, four neuroglial cells; n, nerve cell; p its protoplasmic processes, v its axon. Right top: cells and fibres in the spinal cord. Right bottom: two spinal cord neurons (A,B) with axons (D) and protoplasmic processes (p), and glial cell (C) between them.
Figure 13.
(A) Images of glial cell with richly branched processes (middle) and connective tissue cell (right) drawn by Victor Butzke [103]. (B) Images of glial cells and neurons drawn by Louis-Antoine Ranvier [104]. Left: neuroglial cell of the lateral cords of the ox spinal cord. Middle: elements of the cortical layer of the cat’s brain; a, b, c, and d, four neuroglial cells; n, nerve cell; p its protoplasmic processes, v its axon. Right top: cells and fibres in the spinal cord. Right bottom: two spinal cord neurons (A,B) with axons (D) and protoplasmic processes (p), and glial cell (C) between them.


{kind=link}
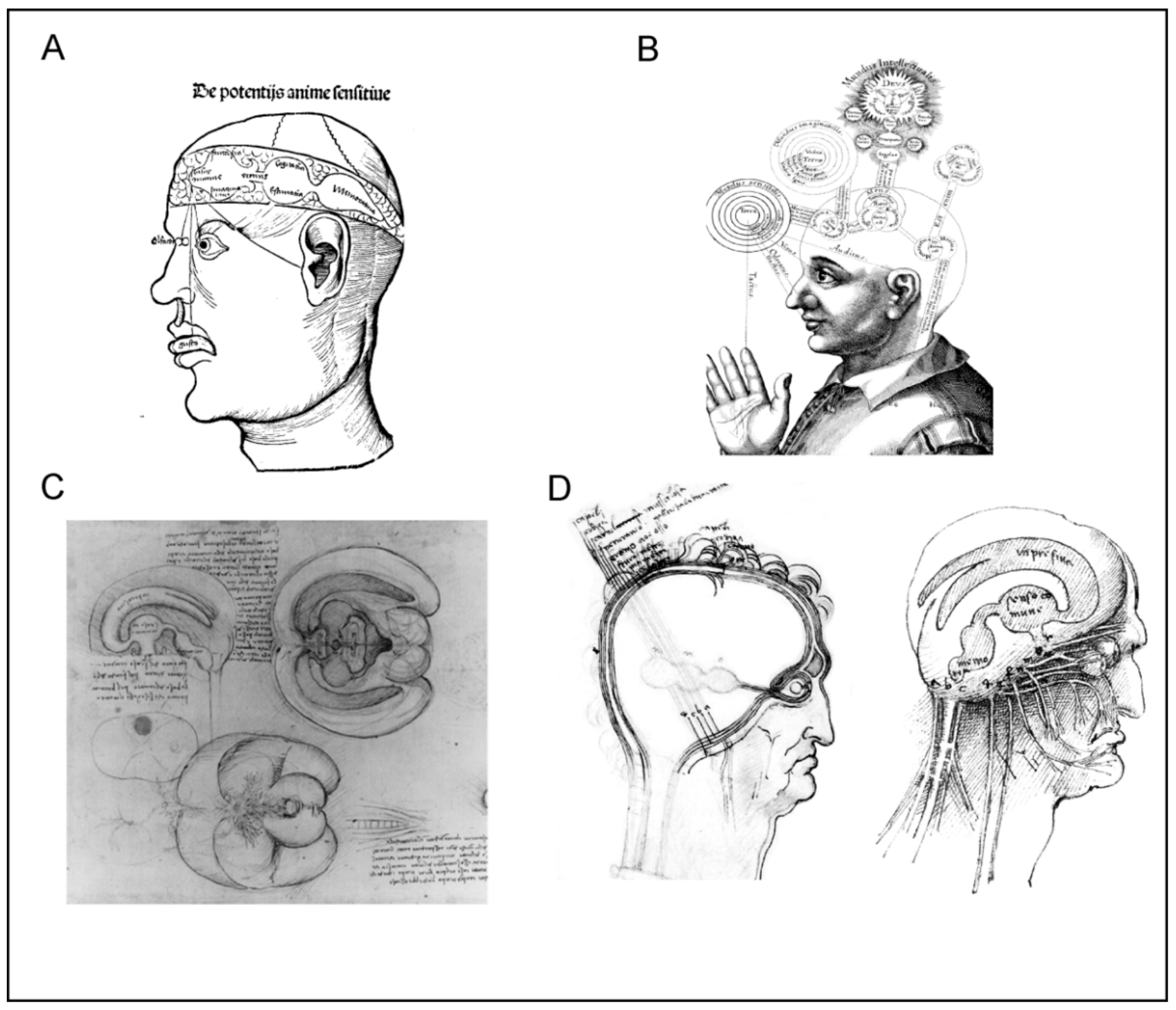{kind=link}
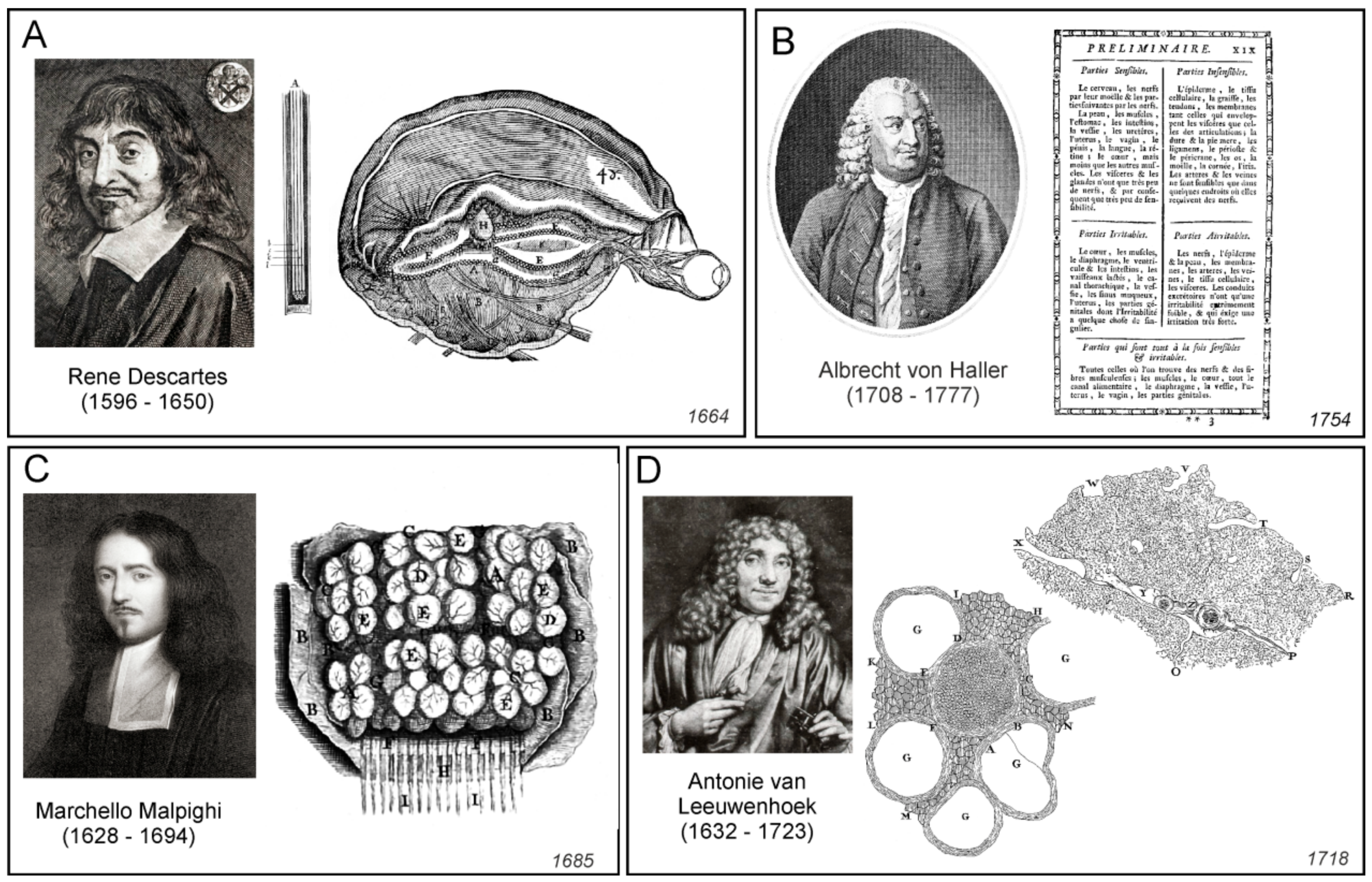{kind=link}
{kind=link}
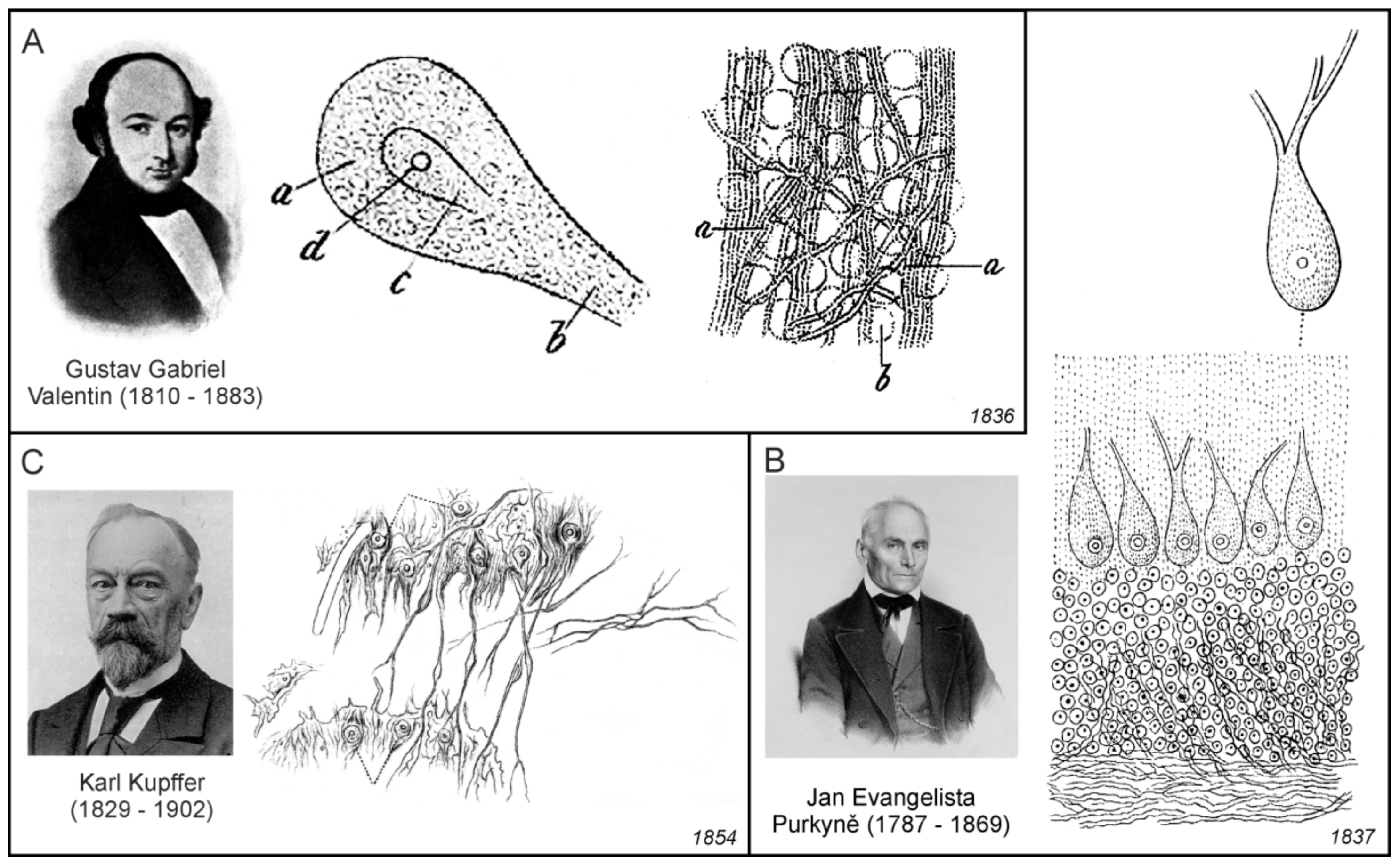{kind=link}
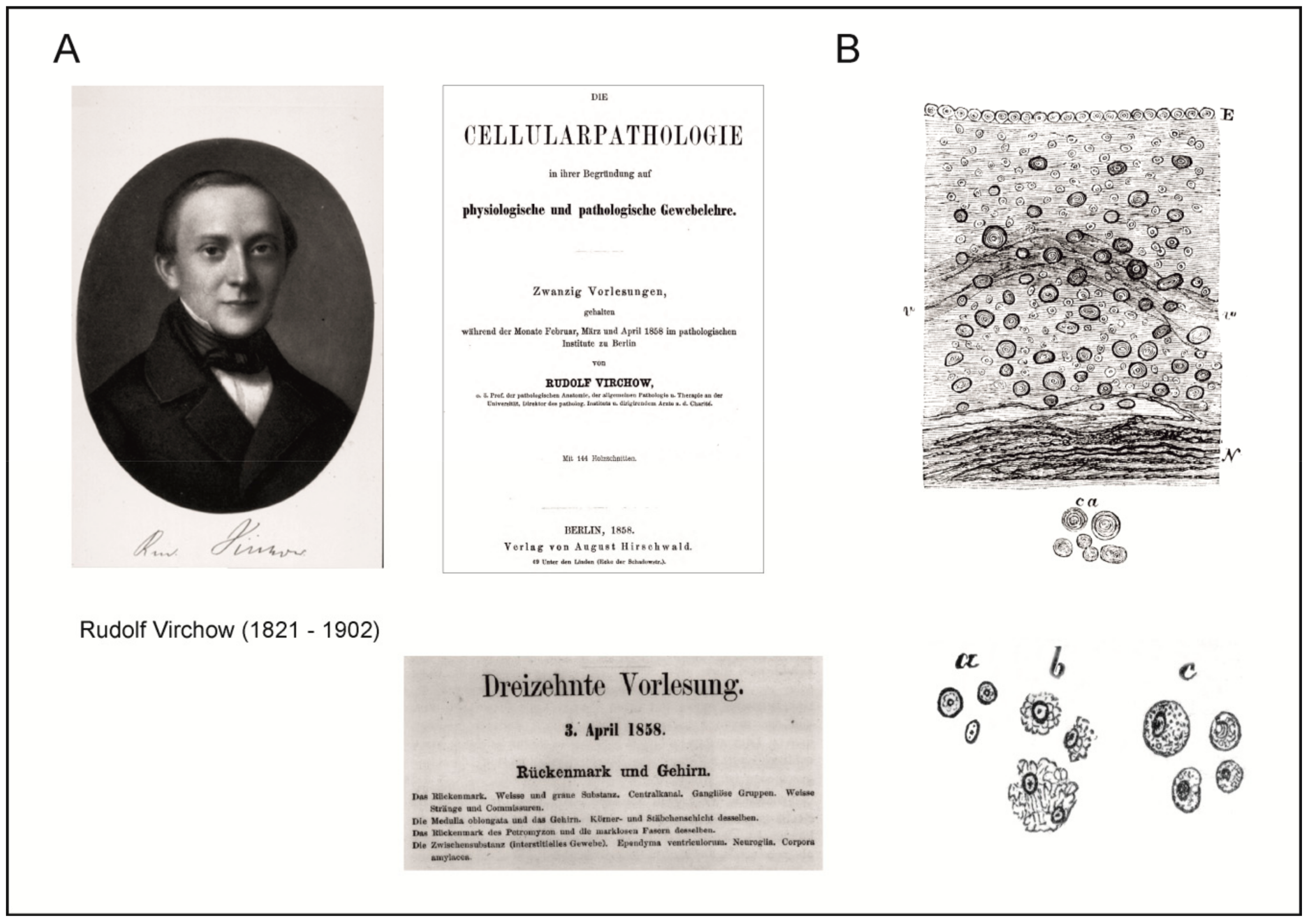{kind=link}
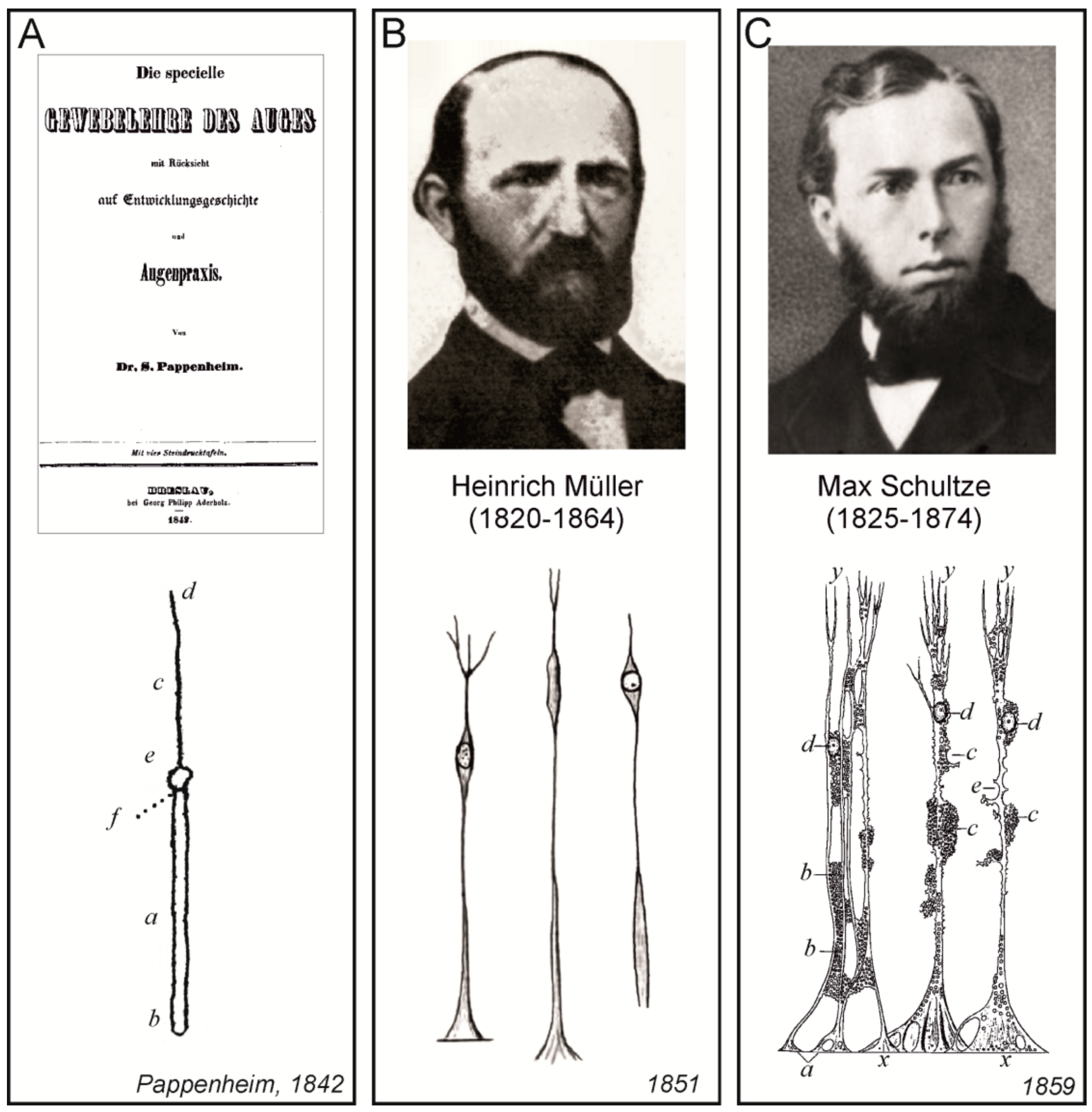{kind=link}
{kind=link}
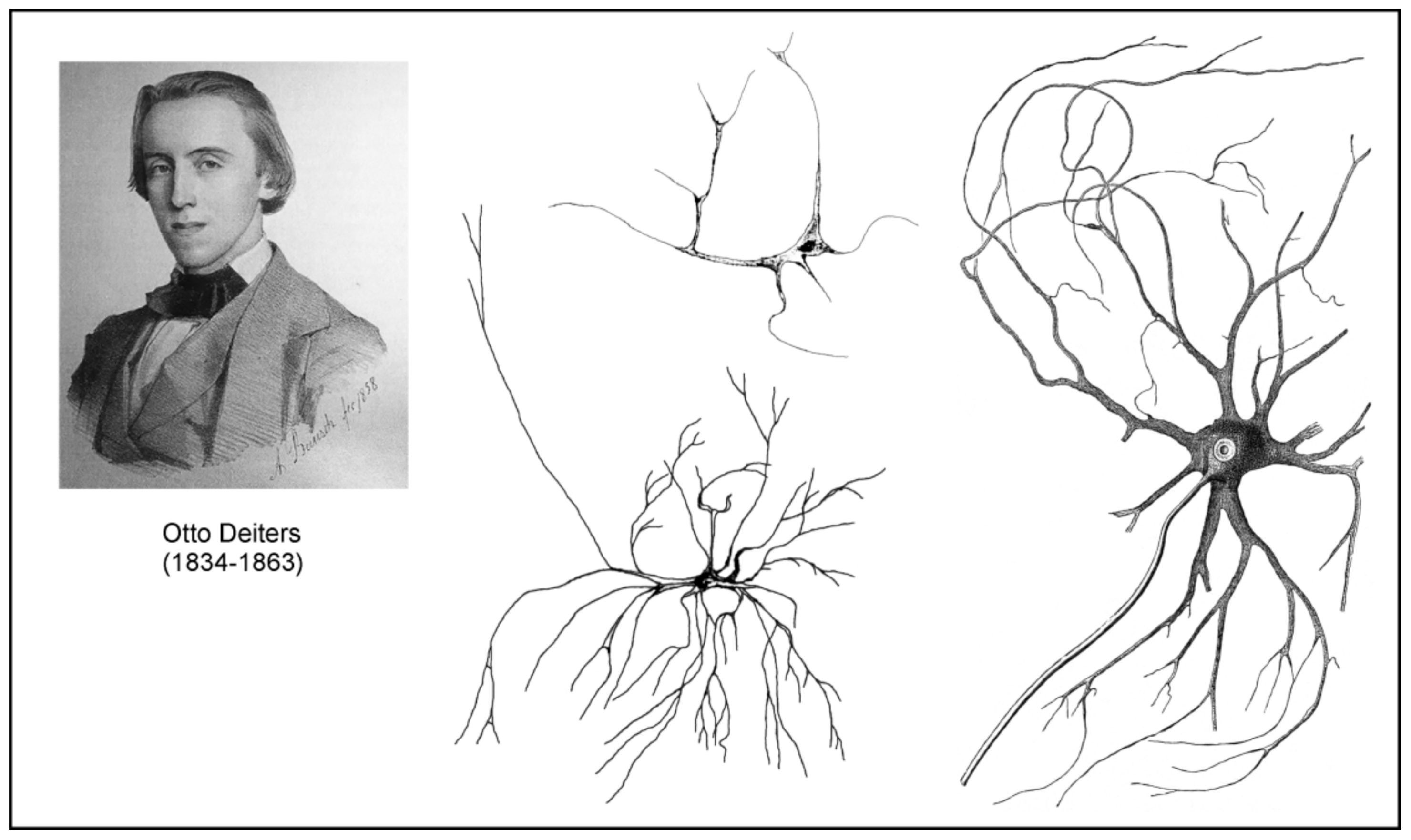{kind=link}
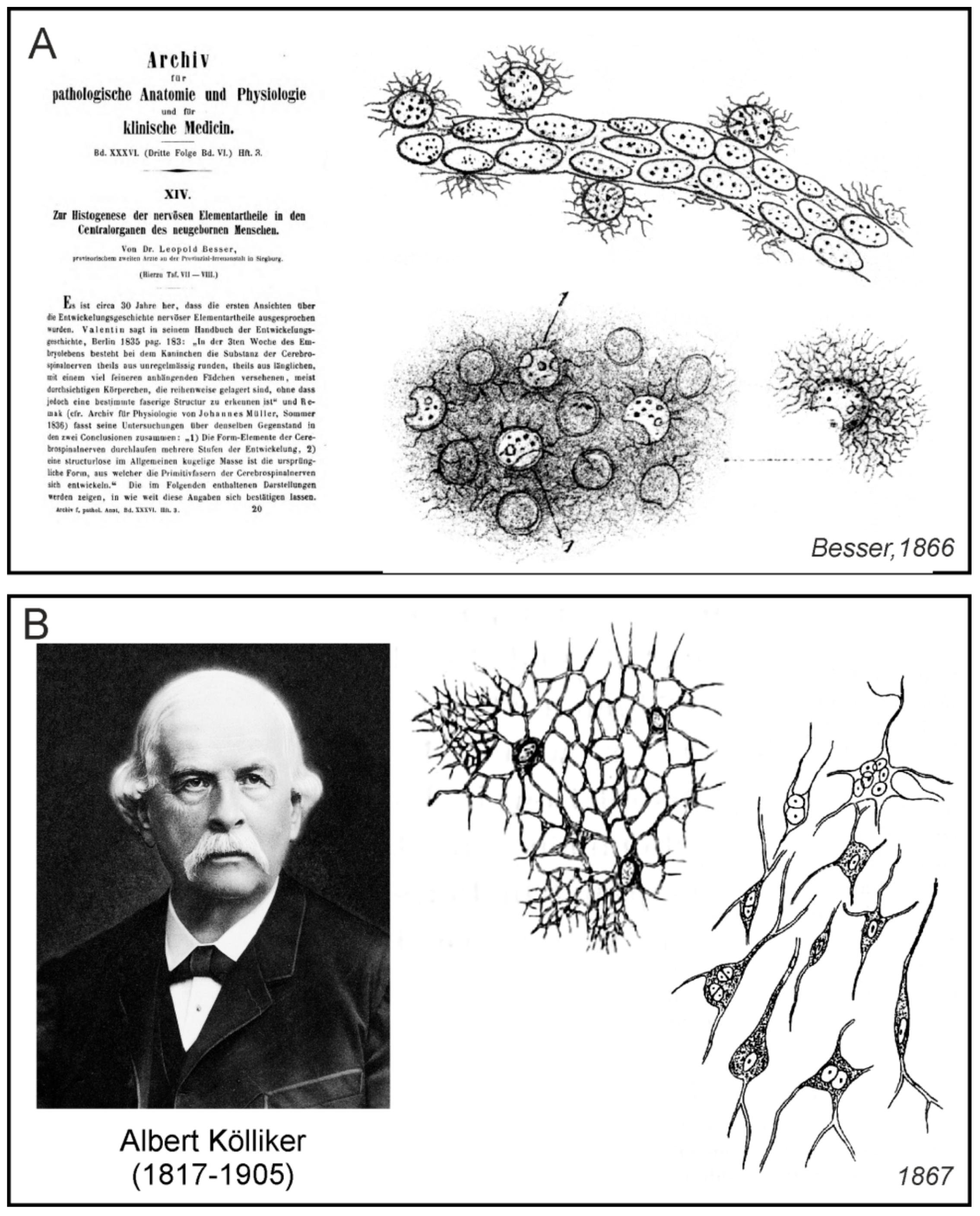{kind=link}
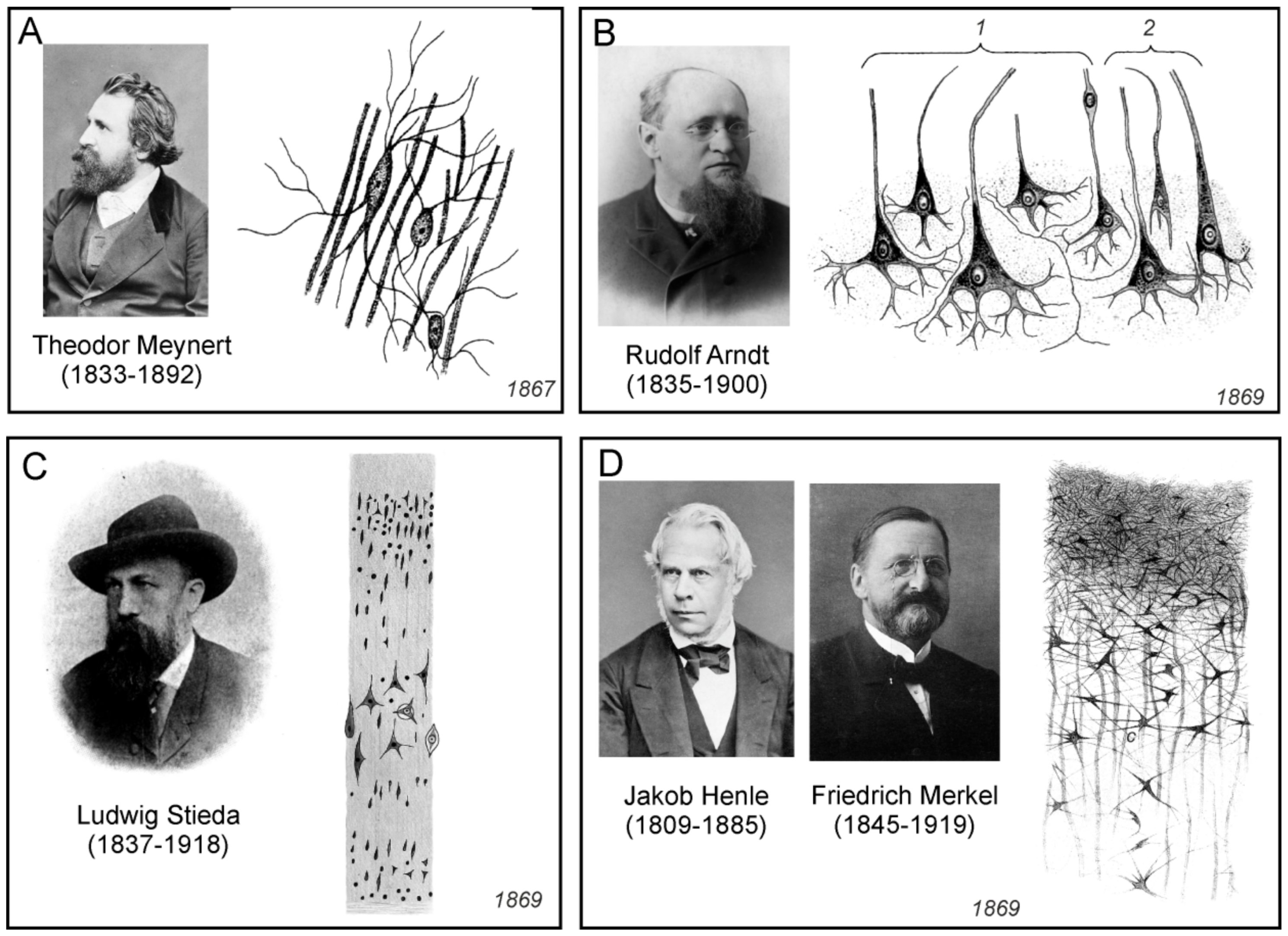{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 15.
Images of glia drawn by Hans Gierke [108,109]. Left: neuroglial cells from the white matter (1–3) and from the grey matter (4); proliferating neuroglial cell (5,6). Right top: large glial cell from the bottom of the fourth ventricle; bottom left: glial cell from the grey matter of the spinal cord; bottom right: glial cell from the white matter of the spinal cord.
Figure 15.
Images of glia drawn by Hans Gierke [108,109]. Left: neuroglial cells from the white matter (1–3) and from the grey matter (4); proliferating neuroglial cell (5,6). Right top: large glial cell from the bottom of the fourth ventricle; bottom left: glial cell from the grey matter of the spinal cord; bottom right: glial cell from the white matter of the spinal cord.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chvátal, A.; Verkhratsky, A. An Early History of Neuroglial Research: Personalities. Neuroglia 2018, 1, 245-257. https://doi.org/10.3390/neuroglia1010016
AMA Style
Chvátal A, Verkhratsky A. An Early History of Neuroglial Research: Personalities. Neuroglia. 2018; 1(1):245-257. https://doi.org/10.3390/neuroglia1010016
Chicago/Turabian StyleChvátal, Alexandr, and Alexei Verkhratsky. 2018. "An Early History of Neuroglial Research: Personalities" Neuroglia 1, no. 1: 245-257. https://doi.org/10.3390/neuroglia1010016