Genomics and Epigenetics of Malignant Mesothelioma

 , , and
, , and
Abstract
:1. Introduction
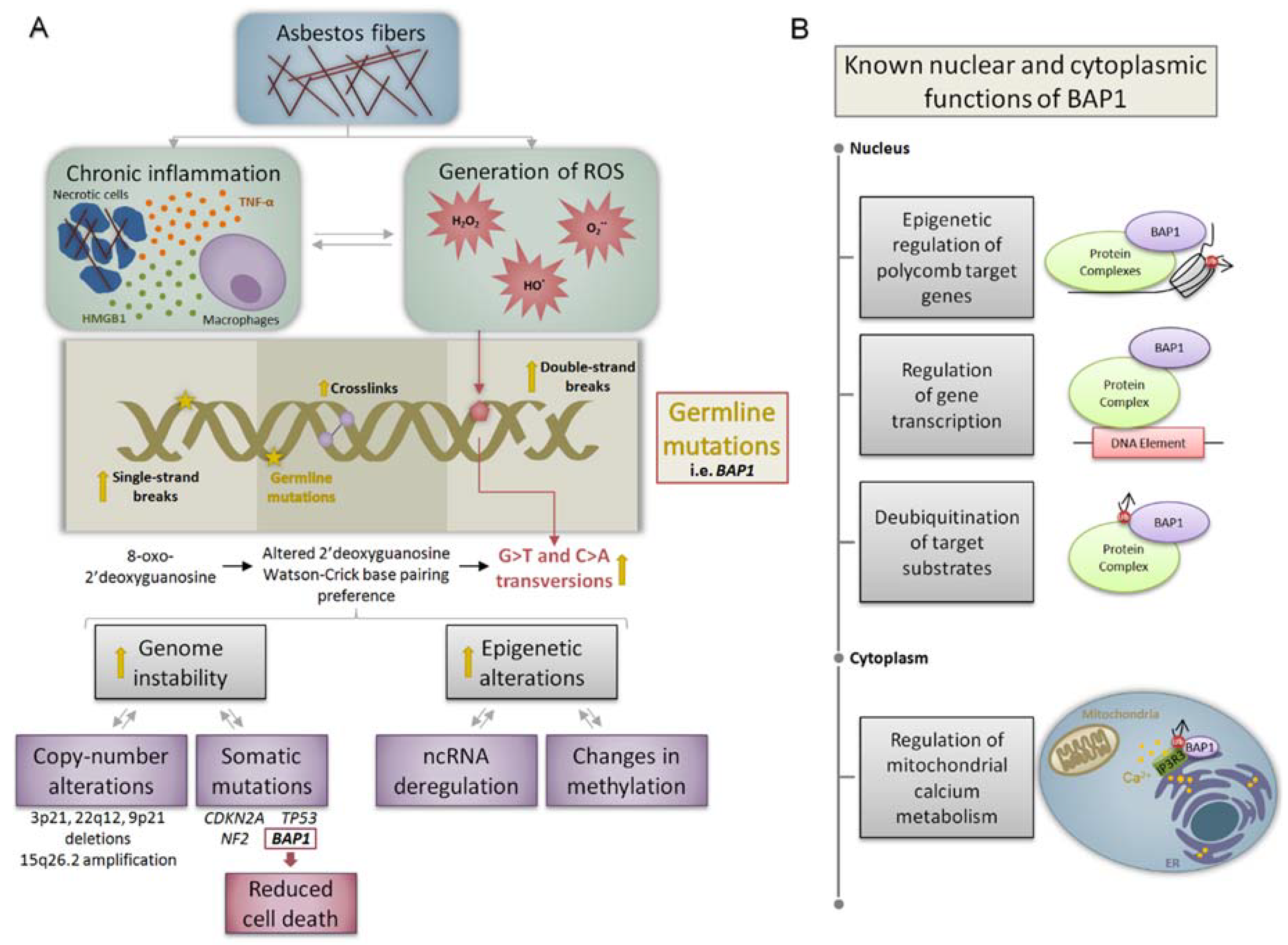
2. Molecular Mechanisms of Asbestos-Related Carcinogenesis
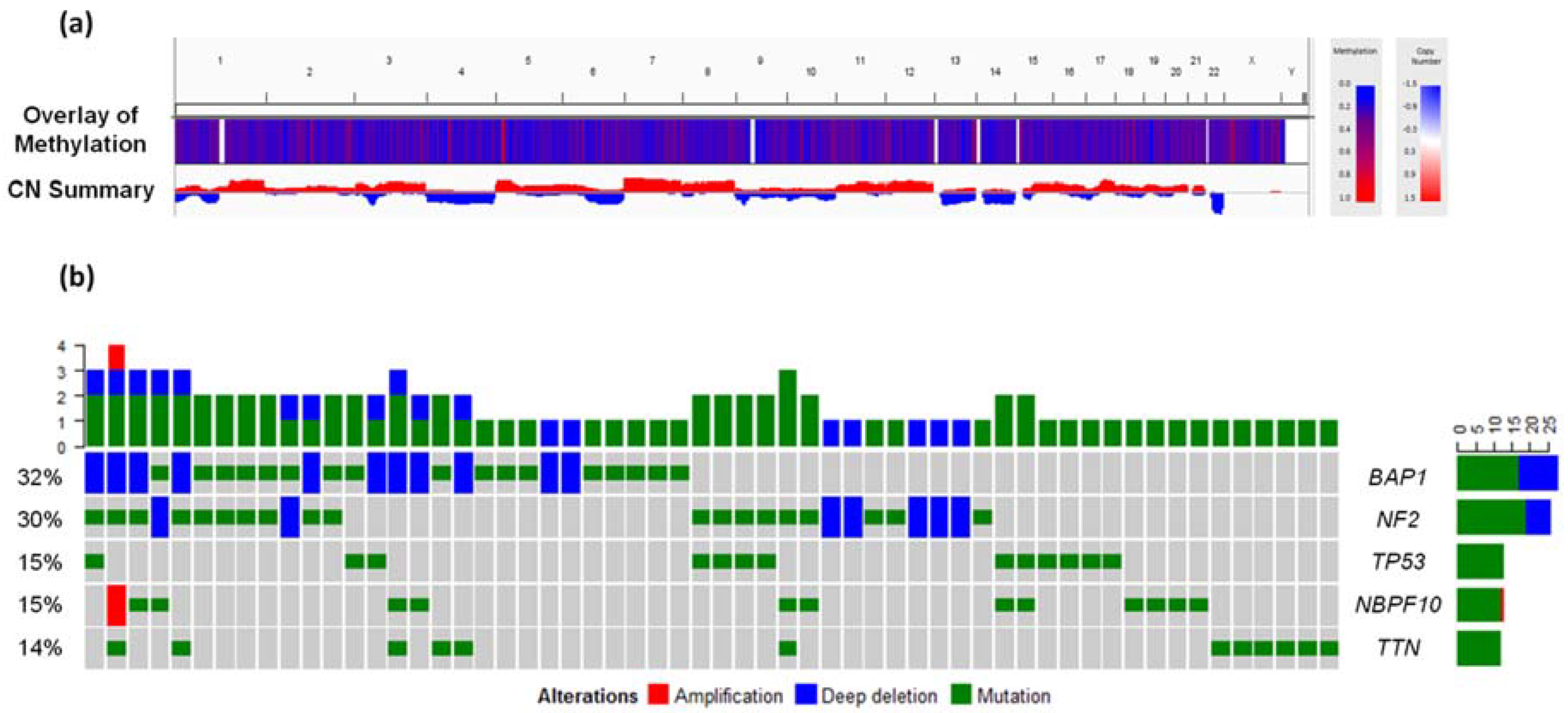
3. Genomic and Epigenetic Landscape of Malignant Mesothelioma
Genetic Variants Modifying the Risk of Malignant Mesothelioma
4. Clinically Relevant Genes Identified through High-Throughput Analyses
4.1. BRCA1-Associated Protein 1 (BAP1)
4.2. Deletion in 9p21
4.3. Additional Genomic Disruptions
5. Non-Coding Transcriptome as a Tissue-Specific Feature in Malignant Mesothelioma
5.1. MicroRNAs
5.2. Long Non-Coding RNAs
6. Conclusions and Future Challenges
Author Contributions
Funding
Conflicts of Interest
References
- Robinson, B.M. Malignant pleural mesothelioma: An epidemiological perspective. Ann. Cardiothorac. Surg. 2012, 1, 491–496. [Google Scholar] [PubMed]
- McDonald, J.C.; McDonald, A.D. The epidemiology of mesothelioma in historical context. Eur. Respir. J. 1996, 9, 1932–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, G. The latency period of mesothelioma among a cohort of British asbestos workers (1978–2005). Br. J. Cancer 2013, 109, 1965–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanphear, B.P.; Buncher, C.R. Latent period for malignant mesothelioma of occupational origin. J. Occup. Med. 1992, 34, 718–721. [Google Scholar] [PubMed]
- Rusch, V.W.; Giroux, D.; Kennedy, C.; Ruffini, E.; Cangir, A.K.; Rice, D.; Pass, H.; Asamura, H.; Waller, D.; Edwards, J.; et al. Initial analysis of the international association for the study of lung cancer mesothelioma database. J. Thorac. Oncol. 2012, 7, 1631–1639. [Google Scholar] [CrossRef] [PubMed]
- Global Burden of Disease Cancer Collaboration; Fitzmaurice, C.; Allen, C.; Barber, R.M.; Barregard, L.; Bhutta, Z.A.; Brenner, H.; Dicker, D.J.; Chimed-Orchir, O.; Dandona, R.; et al. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life-years for 32 Cancer Groups, 1990 to 2015: A Systematic Analysis for the Global Burden of Disease Study. JAMA Oncol. 2017, 3, 524–548. [Google Scholar] [PubMed]
- Carbone, M.; Ly, B.H.; Dodson, R.F.; Pagano, I.; Morris, P.T.; Dogan, U.A.; Gazdar, A.F.; Pass, H.I.; Yang, H. Malignant mesothelioma: Facts, myths, and hypotheses. J. Cell. Physiol. 2012, 227, 44–58. [Google Scholar] [CrossRef] [PubMed]
- The International Ban Asbestos Secretariat. Current Asbestos Bans. Available online: http://www.ibasecretariat.org/alpha_ban_list.php (accessed on 1 April 2018).
- Attanoos, R.L.; Churg, A.; Galateau-Salle, F.; Gibbs, A.R.; Roggli, V.L. Malignant Mesothelioma and Its Non-Asbestos Causes. Arch. Pathol. Lab. Med. 2018, 142, 753–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husain, A.N.; Colby, T.; Ordonez, N.; Krausz, T.; Attanoos, R.; Beasley, M.B.; Borczuk, A.C.; Butnor, K.; Cagle, P.T.; Chirieac, L.R.; et al. Guidelines for pathologic diagnosis of malignant mesothelioma: 2012 update of the consensus statement from the International Mesothelioma Interest Group. Arch. Pathol. Lab. Med. 2013, 137, 647–667. [Google Scholar] [CrossRef] [PubMed]
- Galateau-Salle, F.; Churg, A.; Roggli, V.; Travis, W.D.; World Health Organization Committee for Tumors of the Pleura. The 2015 World Health Organization Classification of Tumors of the Pleura: Advances since the 2004 Classification. J. Thorac. Oncol. 2016, 11, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Galateau-Sallé, F. Pathology of Malignant Mesothelioma; Springer: London, UK, 2010. [Google Scholar]
- Husain, A.N.; Colby, T.V.; Ordonez, N.G.; Allen, T.C.; Attanoos, R.L.; Beasley, M.B.; Butnor, K.J.; Chirieac, L.R.; Churg, A.M.; Dacic, S.; et al. Guidelines for Pathologic Diagnosis of Malignant Mesothelioma 2017 Update of the Consensus Statement From the International Mesothelioma Interest Group. Arch. Pathol. Lab. Med. 2018, 142, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Agalioti, T.; Giannou, A.D.; Stathopoulos, G.T. Pleural involvement in lung cancer. J. Thorac. Dis. 2015, 7, 1021–1030. [Google Scholar] [PubMed]
- Roberts, M.E.; Neville, E.; Berrisford, R.G.; Antunes, G.; Ali, N.J.; on behalf of the BTS Pleural Disease Guideline Group. Management of a malignant pleural effusion: British Thoracic Society Pleural Disease Guideline 2010. Thorax 2010, 65, ii32–ii40. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer (IARC). IARC monographs on the evaluation of the carcinogenic risk of chemicals to man: Asbestos. IARC Monogr. Eval. Carcinog. Risk Hum. 2009, 100C, 1–106. [Google Scholar]
- De Klerk, N.; Reid, A. Hazards of residential exposure to household asbestos. Lancet Public Health 2017, 2, e490–e491. [Google Scholar] [CrossRef]
- World Health Organization, Asbestos. Available online: http://www.who.int/ipcs/assessment/public_health/asbestos/en/ (accessed on 8 April 2018).
- Global Burden of Disease Cancer Collaboration; Fitzmaurice, C.; Dicker, D.; Pain, A.; Hamavid, H.; Moradi-Lakeh, M.; MacIntyre, M.F.; Allen, C.; Hansen, G.; Woodbrook, R.; et al. The Global Burden of Cancer 2013. JAMA Oncol. 2015, 1, 505–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, B.; Driscoll, T. Mesothelioma in Australia: Cresting the third wave. Public Health Res. Pract. 2016, 26. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.; Luce, D. The health impact of nonoccupational exposure to asbestos: What do we know? Eur. J. Cancer Prev. 2009, 18, 489–503. [Google Scholar] [CrossRef] [PubMed]
- CAREX Canada. Asbestos. Available online: https://www.carexcanada.ca/en/asbestos/ (accessed on 3 April 2018).
- Robledo, R.; Mossman, B. Cellular and molecular mechanisms of asbestos-induced fibrosis. J. Cell. Physiol. 1999, 180, 158–166. [Google Scholar] [CrossRef]
- Chew, S.H.; Toyokuni, S. Malignant mesothelioma as an oxidative stress-induced cancer: An update. Free Radic. Biol. Med. 2015, 86, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Kamp, D.W.; Weitzman, S.A. The molecular basis of asbestos induced lung injury. Thorax 1999, 54, 638–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Rivera, Z.; Jube, S.; Nasu, M.; Bertino, P.; Goparaju, C.; Franzoso, G.; Lotze, M.T.; Krausz, T.; Pass, H.I.; et al. Programmed necrosis induced by asbestos in human mesothelial cells causes high-mobility group box 1 protein release and resultant inflammation. Proc. Natl. Acad. Sci. USA 2010, 107, 12611–12616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Tubbs, A.; Nussenzweig, A. Endogenous DNA damage as a source of genomic instability in cancer. Cell 2017, 168, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Bononi, A.; Giorgi, C.; Patergnani, S.; Larson, D.; Verbruggen, K.; Tanji, M.; Pellegrini, L.; Signorato, V.; Olivetto, F.; Pastorino, S.; et al. BAP1 regulates IP3R3-mediated Ca2+ flux to mitochondria suppressing cell transformation. Nature 2017, 546, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network; Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, L.; Jensen, R.V.; De Rienzo, A.; Gordon, G.J.; Xu, Y.; Sugarbaker, D.J.; Bueno, R. Differentially expressed alternatively spliced genes in malignant pleural mesothelioma identified using massively parallel transcriptome sequencing. BMC Med. Genet. 2009, 10, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugarbaker, D.J.; Richards, W.G.; Gordon, G.J.; Dong, L.; De Rienzo, A.; Maulik, G.; Glickman, J.N.; Chirieac, L.R.; Hartman, M.L.; Taillon, B.E.; et al. Transcriptome sequencing of malignant pleural mesothelioma tumors. Proc. Natl. Acad. Sci. USA 2008, 105, 3521–3526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truini, A.; Coco, S.; Nadal, E.; Genova, C.; Mora, M.; Dal Bello, M.G.; Vanni, I.; Alama, A.; Rijavec, E.; Biello, F.; et al. Downregulation of miR-99a/let-7c/miR-125b miRNA cluster predicts clinical outcome in patients with unresected malignant pleural mesothelioma. Oncotarget 2017, 8, 68627–68640. [Google Scholar] [CrossRef] [PubMed]
- Gordon, G.J.; Rockwell, G.N.; Jensen, R.V.; Rheinwald, J.G.; Glickman, J.N.; Aronson, J.P.; Pottorf, B.J.; Nitz, M.D.; Richards, W.G.; Sugarbaker, D.J.; et al. Identification of novel candidate oncogenes and tumor suppressors in malignant pleural mesothelioma using large-scale transcriptional profiling. Am. J. Pathol. 2005, 166, 1827–1840. [Google Scholar] [CrossRef]
- Dragon, J.; Thompson, J.; MacPherson, M.; Shukla, A. Differential Susceptibility of Human Pleural and Peritoneal Mesothelial Cells to Asbestos Exposure. J. Cell. Biochem. 2015, 116, 1540–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, G.; Chmielecki, J.; Goparaju, C.; Heguy, A.; Dolgalev, I.; Carbone, M.; Seepo, S.; Meyerson, M.; Pass, H.I. Whole-exome sequencing reveals frequent genetic alterations in BAP1, NF2, CDKN2A, and CUL1 in malignant pleural mesothelioma. Cancer Res. 2015, 75, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Maki-Nevala, S.; Sarhadi, V.K.; Knuuttila, A.; Scheinin, I.; Ellonen, P.; Lagstrom, S.; Ronty, M.; Kettunen, E.; Husgafvel-Pursiainen, K.; Wolff, H.; et al. Driver Gene and Novel Mutations in Asbestos-Exposed Lung Adenocarcinoma and Malignant Mesothelioma Detected by Exome Sequencing. Lung 2016, 194, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.C.; Kim, H.K.; Lee, S.; Mendez, P.; Kim, J.W.; Woodard, G.; Yoon, J.H.; Jen, K.Y.; Fang, L.T.; Jones, K.; et al. Whole exome and targeted deep sequencing identify genome-wide allelic loss and frequent SETDB1 mutations in malignant pleural mesotheliomas. Oncotarget 2016, 7, 8321–8331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patch, A.M.; Nones, K.; Kazakoff, S.H.; Newell, F.; Wood, S.; Leonard, C.; Holmes, O.; Xu, Q.; Addala, V.; Creaney, J.; et al. Germline and somatic variant identification using BGISEQ-500 and HiSeq X Ten whole genome sequencing. PLoS ONE 2018, 13, e0190264. [Google Scholar] [CrossRef] [PubMed]
- Bueno, R.; De Rienzo, A.; Dong, L.; Gordon, G.J.; Hercus, C.F.; Richards, W.G.; Jensen, R.V.; Anwar, A.; Maulik, G.; Chirieac, L.R.; et al. Second generation sequencing of the mesothelioma tumor genome. PLoS ONE 2010, 5, e10612. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Tomson, B.N.; Buys, T.P.; Elkin, S.K.; Carter, J.L.; Kurzrock, R. Genomic Landscape of Malignant Mesotheliomas. Mol. Cancer Ther. 2016, 15, 2498–2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Iacono, M.; Monica, V.; Righi, L.; Grosso, F.; Libener, R.; Vatrano, S.; Bironzo, P.; Novello, S.; Musmeci, L.; Volante, M.; et al. Targeted next-generation sequencing of cancer genes in advanced stage malignant pleural mesothelioma: A retrospective study. J. Thorac. Oncol. 2015, 10, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Ladanyi, M.; Robinson, B.W.S.; Campbell, P.J. The TCGA malignant pleural mesothelioma (MPM) project: VISTA expression and delineation of a novel clinical-molecular subtype of MPM. J. Clin. Oncol. 2018, 36, 8516. [Google Scholar]
- National Mesothelioma Virtual Bank. Available online: www.data.mesotissue.org/mvb/home.seam?cid=840 (accessed on 10 June 2018).
- NCBI clinvar. Available online: www.ncbi.nlm.nih.gov/clinvar/ (accessed on 10 June 2018).
- Carbone, M.; Yang, H.; Pass, H.I.; Krausz, T.; Testa, J.R.; Gaudino, G. BAP1 and cancer. Nat. Rev. Cancer 2013, 13, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Bueno, R.; Stawiski, E.W.; Goldstein, L.D.; Durinck, S.; De Rienzo, A.; Modrusan, Z.; Gnad, F.; Nguyen, T.T.; Jaiswal, B.S.; Chirieac, L.R.; et al. Comprehensive genomic analysis of malignant pleural mesothelioma identifies recurrent mutations, gene fusions and splicing alterations. Nat. Genet. 2016, 48, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Knijnenburg, T.A.; Wang, L.; Zimmermann, M.T.; Chambwe, N.; Gao, G.F.; Cherniack, A.D.; Fan, H.; Shen, H.; Way, G.P.; Greene, C.S.; et al. Genomic and Molecular Landscape of DNA Damage Repair Deficiency across The Cancer Genome Atlas. Cell Rep. 2018, 23, 239–254.e236. [Google Scholar] [CrossRef] [PubMed]
- Hylebos, M.; Van Camp, G.; Vandeweyer, G.; Fransen, E.; Beyens, M.; Cornelissen, R.; Suls, A.; Pauwels, P.; van Meerbeeck, J.P.; Op de Beeck, K. Large-scale copy number analysis reveals variations in genes not previously associated with malignant pleural mesothelioma. Oncotarget 2017, 8, 113673–113686. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, Y.; Emi, M.; Hashimoto-Tamaoki, T.; Ohmuraya, M.; Sato, A.; Tsujimura, T.; Hasegawa, S.; Nakano, T.; Nasu, M.; Pastorino, S.; et al. High-density array-CGH with targeted NGS unmask multiple noncontiguous minute deletions on chromosome 3p21 in mesothelioma. Proc. Natl. Acad. Sci. USA 2016, 113, 13432–13437. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.T.; Thorvaldsdottir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorvaldsdottir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed]
- Chirac, P.; Maillet, D.; Lepretre, F.; Isaac, S.; Glehen, O.; Figeac, M.; Villeneuve, L.; Peron, J.; Gibson, F.; Galateau-Salle, F.; et al. Genomic copy number alterations in 33 malignant peritoneal mesothelioma analyzed by comparative genomic hybridization array. Hum. Pathol. 2016, 55, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.C.; Houseman, E.A.; Godleski, J.J.; Marsit, C.J.; Longacker, J.L.; Roelofs, C.R.; Karagas, M.R.; Wrensch, M.R.; Yeh, R.F.; Nelson, H.H.; et al. Epigenetic profiles distinguish pleural mesothelioma from normal pleura and predict lung asbestos burden and clinical outcome. Cancer Res. 2009, 69, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.C.; Houseman, E.A.; Poage, G.M.; Godleski, J.J.; Bueno, R.; Sugarbaker, D.J.; Wiencke, J.K.; Nelson, H.H.; Marsit, C.J.; Kelsey, K.T. Integrated profiling reveals a global correlation between epigenetic and genetic alterations in mesothelioma. Cancer Res. 2010, 70, 5686–5694. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, K.C.; Kaufman, A.S.; Schrump, D.S. Targeting the epigenome in malignant pleural mesothelioma. Transl. Lung Cancer Res. 2017, 6, 350–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillegass, J.M.; Miller, J.M.; MacPherson, M.B.; Westbom, C.M.; Sayan, M.; Thompson, J.K.; Macura, S.L.; Perkins, T.N.; Beuschel, S.L.; Alexeeva, V.; et al. Asbestos and erionite prime and activate the NLRP3 inflammasome that stimulates autocrine cytokine release in human mesothelial cells. Part. Fibre Toxicol. 2013, 10, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, M. Functional and cancer genomics of ASXL family members. Br. J. Cancer 2013, 109, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, H.; Prokop, J.; Karar, J.; Park, K.; Cao, L.; Harbour, J.W.; Bowcock, A.M.; Malkowicz, S.B.; Cheung, M.; Testa, J.R.; et al. Familial and Somatic BAP1 Mutations Inactivate ASXL1/2-Mediated Allosteric Regulation of BAP1 Deubiquitinase by Targeting Multiple Independent Domains. Cancer Res. 2018, 78, 1200–1213. [Google Scholar] [CrossRef] [PubMed]
- LaFave, L.M.; Beguelin, W.; Koche, R.; Teater, M.; Spitzer, B.; Chramiec, A.; Papalexi, E.; Keller, M.D.; Hricik, T.; Konstantinoff, K.; et al. Loss of BAP1 function leads to EZH2-dependent transformation. Nat. Med. 2015, 21, 1344–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.C.; Kim, N.Y.; Seo, Y.R.; Kim, Y. An Integrated Analysis of the Genome-Wide Profiles of DNA Methylation and mRNA Expression Defining the Side Population of a Human Malignant Mesothelioma Cell Line. J. Cancer 2016, 7, 1668–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casalone, E.; Allione, A.; Viberti, C.; Pardini, B.; Guarrera, S.; Betti, M.; Dianzani, I.; Aldieri, E.; Matullo, G. DNA methylation profiling of asbestos-treated MeT5A cell line reveals novel pathways implicated in asbestos response. Arch. Toxicol. 2018, 92, 1785–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Testa, J.R.; Cheung, M.; Pei, J.; Below, J.E.; Tan, Y.; Sementino, E.; Cox, N.J.; Dogan, A.U.; Pass, H.I.; Trusa, S.; et al. Germline BAP1 mutations predispose to malignant mesothelioma. Nat. Genet. 2011, 43, 1022–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matullo, G.; Guarrera, S.; Betti, M.; Fiorito, G.; Ferrante, D.; Voglino, F.; Cadby, G.; Di Gaetano, C.; Rosa, F.; Russo, A.; et al. Genetic variants associated with increased risk of malignant pleural mesothelioma: A genome-wide association study. PLoS ONE 2013, 8, e61253. [Google Scholar] [CrossRef] [PubMed]
- Cadby, G.; Mukherjee, S.; Musk, A.W.; Reid, A.; Garlepp, M.; Dick, I.; Robinson, C.; Hui, J.; Fiorito, G.; Guarrera, S.; et al. A genome-wide association study for malignant mesothelioma risk. Lung Cancer 2013, 82, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tunesi, S.; Ferrante, D.; Mirabelli, D.; Andorno, S.; Betti, M.; Fiorito, G.; Guarrera, S.; Casalone, E.; Neri, M.; Ugolini, D.; et al. Gene-asbestos interaction in malignant pleural mesothelioma susceptibility. Carcinogenesis 2015, 36, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.; Emri, S.; Dogan, A.U.; Steele, I.; Tuncer, M.; Pass, H.I.; Baris, Y.I. A mesothelioma epidemic in Cappadocia: Scientific developments and unexpected social outcomes. Nat. Rev. Cancer 2007, 7, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Roushdy-Hammady, I.; Siegel, J.; Emri, S.; Testa, J.R.; Carbone, M. Genetic-susceptibility factor and malignant mesothelioma in the Cappadocian region of Turkey. Lancet 2001, 357, 444–445. [Google Scholar] [CrossRef]
- McCambridge, A.J.; Napolitano, A.; Mansfield, A.S.; Fennell, D.A.; Sekido, Y.; Nowak, A.K.; Reungwetwattana, T.; Mao, W.; Pass, H.I.; Carbone, M.; et al. Progress in the Management of Malignant Pleural Mesothelioma in 2017. J. Thorac. Oncol. 2018, 13, 606–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bott, M.; Brevet, M.; Taylor, B.S.; Shimizu, S.; Ito, T.; Wang, L.; Creaney, J.; Lake, R.A.; Zakowski, M.F.; Reva, B.; et al. The nuclear deubiquitinase BAP1 is commonly inactivated by somatic mutations and 3p21.1 losses in malignant pleural mesothelioma. Nat. Genet. 2011, 43, 668–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Kadariya, Y.; Cheung, M.; Pei, J.; Talarchek, J.; Sementino, E.; Tan, Y.; Menges, C.W.; Cai, K.Q.; Litwin, S.; et al. Germline mutation of Bap1 accelerates development of asbestos-induced malignant mesothelioma. Cancer Res. 2014, 74, 4388–4397. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, T.; Tsuruga, T.; Kuroda, T.; Nishikawa, H.; Ohta, T. Functional Link between BRCA1 and BAP1 through Histone H2A, Heterochromatin and DNA Damage Response. Curr. Cancer Drug Targets 2016, 16, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Ismail, I.H.; Davidson, R.; Gagne, J.P.; Xu, Z.Z.; Poirier, G.G.; Hendzel, M.J. Germline mutations in BAP1 impair its function in DNA double-strand break repair. Cancer Res. 2014, 74, 4282–4294. [Google Scholar] [CrossRef] [PubMed]
- Krasinskas, A.M.; Bartlett, D.L.; Cieply, K.; Dacic, S. CDKN2A and MTAP deletions in peritoneal mesotheliomas are correlated with loss of p16 protein expression and poor survival. Mod. Pathol. 2010, 23, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Dacic, S.; Kothmaier, H.; Land, S.; Shuai, Y.; Halbwedl, I.; Morbini, P.; Murer, B.; Comin, C.; Galateau-Salle, F.; Demirag, F.; et al. Prognostic significance of p16/cdkn2a loss in pleural malignant mesotheliomas. Virchows Arch. 2008, 453, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Rios, F.; Chuai, S.; Flores, R.; Shimizu, S.; Ohno, T.; Wakahara, K.; Illei, P.B.; Hussain, S.; Krug, L.; Zakowski, M.F.; et al. Global gene expression profiling of pleural mesotheliomas: Overexpression of aurora kinases and P16/CDKN2A deletion as prognostic factors and critical evaluation of microarray-based prognostic prediction. Cancer Res 2006, 66, 2970–2979. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.C.; Sheffield, B.S.; Rodriguez, S.; Thompson, K.; Tse, C.H.; Gown, A.M.; Churg, A. Utility of BAP1 Immunohistochemistry and p16 (CDKN2A) FISH in the Diagnosis of Malignant Mesothelioma in Effusion Cytology Specimens. Am. J. Surg. Pathol. 2016, 40, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Husain, A.N.; Colby, T.V.; Ordonez, N.G.; Krausz, T.; Borczuk, A.; Cagle, P.T.; Chirieac, L.R.; Churg, A.; Galateau-Salle, F.; Gibbs, A.R.; et al. Guidelines for pathologic diagnosis of malignant mesothelioma: A consensus statement from the International Mesothelioma Interest Group. Arch. Pathol. Lab. Med. 2009, 133, 1317–1331. [Google Scholar] [PubMed]
- Miyanaga, A.; Masuda, M.; Tsuta, K.; Kawasaki, K.; Nakamura, Y.; Sakuma, T.; Asamura, H.; Gemma, A.; Yamada, T. Hippo pathway gene mutations in malignant mesothelioma: Revealed by RNA and targeted exon sequencing. J. Thorac. Oncol. 2015, 10, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, T.; Osada, H.; Murakami, H.; Tatematsu, Y.; Taniguchi, T.; Kondo, Y.; Yatabe, Y.; Hasegawa, Y.; Shimokata, K.; Horio, Y.; et al. YAP1 is involved in mesothelioma development and negatively regulated by Merlin through phosphorylation. Carcinogenesis 2008, 29, 2139–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, Y.; Hida, T.; Hamasaki, M.; Matsumoto, S.; Sato, A.; Tsujimura, T.; Kawahara, K.; Hiroshima, K.; Oda, Y.; Nabeshima, K. A combination of MTAP and BAP1 immunohistochemistry in pleural effusion cytology for the diagnosis of mesothelioma. Cancer Cytopathol. 2018, 126, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Thurneysen, C.; Opitz, I.; Kurtz, S.; Weder, W.; Stahel, R.A.; Felley-Bosco, E. Functional inactivation of NF2/merlin in human mesothelioma. Lung Cancer 2009, 64, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Deguen, B.; Goutebroze, L.; Giovannini, M.; Boisson, C.; van der Neut, R.; Jaurand, M.C.; Thomas, G. Heterogeneity of mesothelioma cell lines as defined by altered genomic structure and expression of the NF2 gene. Int. J. Cancer 1998, 77, 554–560. [Google Scholar] [CrossRef]
- Bianchi, A.B.; Mitsunaga, S.I.; Cheng, J.Q.; Klein, W.M.; Jhanwar, S.C.; Seizinger, B.; Kley, N.; Klein-Szanto, A.J.; Testa, J.R. High frequency of inactivating mutations in the neurofibromatosis type 2 gene (NF2) in primary malignant mesotheliomas. Proc. Natl. Acad. Sci. USA 1995, 92, 10854–10858. [Google Scholar] [CrossRef] [PubMed]
- Poulikakos, P.I.; Xiao, G.H.; Gallagher, R.; Jablonski, S.; Jhanwar, S.C.; Testa, J.R. Re-expression of the tumor suppressor NF2/merlin inhibits invasiveness in mesothelioma cells and negatively regulates FAK. Oncogene 2006, 25, 5960–5968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, I.M.; Kolev, V.N.; Vidal, C.M.; Kadariya, Y.; Ring, J.E.; Wright, Q.; Weaver, D.T.; Menges, C.; Padval, M.; McClatchey, A.I.; et al. Merlin deficiency predicts FAK inhibitor sensitivity: A synthetic lethal relationship. Sci. Transl. Med. 2014, 6, 237ra268. [Google Scholar] [CrossRef] [PubMed]
- Gordon, G.J.; Jensen, R.V.; Hsiao, L.L.; Gullans, S.R.; Blumenstock, J.E.; Richards, W.G.; Jaklitsch, M.T.; Sugarbaker, D.J.; Bueno, R. Using gene expression ratios to predict outcome among patients with mesothelioma. J. Natl. Cancer Inst. 2003, 95, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Pass, H.I. Biomarkers and prognostic factors for mesothelioma. Ann. Cardiothorac. Surg. 2012, 1, 449–456. [Google Scholar] [PubMed]
- De Reynies, A.; Jaurand, M.C.; Renier, A.; Couchy, G.; Hysi, I.; Elarouci, N.; Galateau-Salle, F.; Copin, M.C.; Hofman, P.; Cazes, A.; et al. Molecular classification of malignant pleural mesothelioma: Identification of a poor prognosis subgroup linked to the epithelial-to-mesenchymal transition. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2014, 20, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Huarte, M. The emerging role of lncRNAs in cancer. Nat. Med. 2015, 21, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, H.; Fabbri, M.; Calin, G.A. MicroRNAs and other non-coding RNAs as targets for anticancer drug development. Nat. Rev. Drug Discov. 2013, 12, 847–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hausser, J.; Zavolan, M. Identification and consequences of miRNA-target interactions—Beyond repression of gene expression. Nat. Rev. Genet. 2014, 15, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Guled, M.; Lahti, L.; Lindholm, P.M.; Salmenkivi, K.; Bagwan, I.; Nicholson, A.G.; Knuutila, S. CDKN2A, NF2, and JUN are dysregulated among other genes by miRNAs in malignant mesothelioma—A miRNA microarray analysis. Genes Chromosom. Cancer 2009, 48, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Borczuk, A.C.; Pei, J.; Taub, R.N.; Levy, B.; Nahum, O.; Chen, J.; Chen, K.; Testa, J.R. Genome-wide analysis of abdominal and pleural malignant mesothelioma with DNA arrays reveals both common and distinct regions of copy number alteration. Cancer Biol. Ther. 2016, 17, 328–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, T.; Karnan, S.; Fukui, T.; Yokoyama, T.; Tagawa, H.; Yokoi, K.; Ueda, Y.; Mitsudomi, T.; Horio, Y.; Hida, T.; et al. Genomic profiling of malignant pleural mesothelioma with array-based comparative genomic hybridization shows frequent non-random chromosomal alteration regions including JUN amplification on 1p32. Cancer Sci. 2007, 98, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Santarelli, L.; Strafella, E.; Staffolani, S.; Amati, M.; Emanuelli, M.; Sartini, D.; Pozzi, V.; Carbonari, D.; Bracci, M.; Pignotti, E.; et al. Association of MiR-126 with soluble mesothelin-related peptides, a marker for malignant mesothelioma. PLoS ONE 2011, 6, e18232. [Google Scholar] [CrossRef] [PubMed]
- Tomasetti, M.; Staffolani, S.; Nocchi, L.; Neuzil, J.; Strafella, E.; Manzella, N.; Mariotti, L.; Bracci, M.; Valentino, M.; Amati, M.; et al. Clinical significance of circulating miR-126 quantification in malignant mesothelioma patients. Clin. Biochem. 2012, 45, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, M.B.; Cheng, Y.Y.; Badrian, B.; Kao, S.C.; Creaney, J.; Edelman, J.J.; Armstrong, N.J.; Vallely, M.P.; Musk, A.W.; Robinson, B.W.; et al. Increased circulating miR-625-3p: A potential biomarker for patients with malignant pleural mesothelioma. J. Thorac. Oncol. 2012, 7, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- Mozzoni, P.; Ampollini, L.; Goldoni, M.; Alinovi, R.; Tiseo, M.; Gnetti, L.; Carbognani, P.; Rusca, M.; Mutti, A.; Percesepe, A.; et al. MicroRNA Expression in Malignant Pleural Mesothelioma and Asbestosis: A Pilot Study. Dis. Markers 2017, 2017, 9645940. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, H.; Lebanony, D.; Rosenwald, S.; Cohen, L.; Gibori, H.; Barabash, N.; Ashkenazi, K.; Goren, E.; Meiri, E.; Morgenstern, S.; et al. A diagnostic assay based on microRNA expression accurately identifies malignant pleural mesothelioma. J. Mol. Diagn. 2010, 12, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Gee, G.V.; Koestler, D.C.; Christensen, B.C.; Sugarbaker, D.J.; Ugolini, D.; Ivaldi, G.P.; Resnick, M.B.; Houseman, E.A.; Kelsey, K.T.; Marsit, C.J. Downregulated microRNAs in the differential diagnosis of malignant pleural mesothelioma. Int. J. Cancer 2010, 127, 2859–2869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, D.G.; Johnen, G.; Bryk, O.; Jockel, K.H.; Bruning, T. Identification of miRNA-103 in the cellular fraction of human peripheral blood as a potential biomarker for malignant mesothelioma—A pilot study. PLoS ONE 2012, 7, e30221. [Google Scholar] [CrossRef] [PubMed]
- Cavalleri, T.; Angelici, L.; Favero, C.; Dioni, L.; Mensi, C.; Bareggi, C.; Palleschi, A.; Rimessi, A.; Consonni, D.; Bordini, L.; et al. Plasmatic extracellular vesicle microRNAs in malignant pleural mesothelioma and asbestos-exposed subjects suggest a 2-miRNA signature as potential biomarker of disease. PLoS ONE 2017, 12, e0176680. [Google Scholar] [CrossRef] [PubMed]
- Muraoka, T.; Soh, J.; Toyooka, S.; Aoe, K.; Fujimoto, N.; Hashida, S.; Maki, Y.; Tanaka, N.; Shien, K.; Furukawa, M.; et al. The degree of microRNA-34b/c methylation in serum-circulating DNA is associated with malignant pleural mesothelioma. Lung Cancer 2013, 82, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, M.; Grauslund, M.; Ravn, J.; Sorensen, J.B.; Andersen, C.B.; Santoni-Rugiu, E. Diagnostic potential of miR-126, miR-143, miR-145, and miR-652 in malignant pleural mesothelioma. J. Mol. Diagn. 2014, 16, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.G.; Gawrych, K.; Casjens, S.; Brik, A.; Lehnert, M.; Taeger, D.; Pesch, B.; Kollmeier, J.; Bauer, T.T.; Johnen, G.; et al. Circulating miR-132-3p as a Candidate Diagnostic Biomarker for Malignant Mesothelioma. Dis. Markers 2017, 2017, 9280170. [Google Scholar] [CrossRef] [PubMed]
- Bononi, I.; Comar, M.; Puozzo, A.; Stendardo, M.; Boschetto, P.; Orecchia, S.; Libener, R.; Guaschino, R.; Pietrobon, S.; Ferracin, M.; et al. Circulating microRNAs found dysregulated in ex-exposed asbestos workers and pleural mesothelioma patients as potential new biomarkers. Oncotarget 2016, 7, 82700–82711. [Google Scholar] [CrossRef] [PubMed]
- Cappellesso, R.; Nicole, L.; Caroccia, B.; Guzzardo, V.; Ventura, L.; Fassan, M.; Fassina, A. Young investigator challenge: MicroRNA-21/MicroRNA-126 profiling as a novel tool for the diagnosis of malignant mesothelioma in pleural effusion cytology. Cancer Cytopathol. 2016, 124, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pass, H.I.; Goparaju, C.; Ivanov, S.; Donington, J.; Carbone, M.; Hoshen, M.; Cohen, D.; Chajut, A.; Rosenwald, S.; Dan, H.; et al. hsa-miR-29c* is linked to the prognosis of malignant pleural mesothelioma. Cancer Res. 2010, 70, 1916–1924. [Google Scholar] [CrossRef] [PubMed]
- Busacca, S.; Germano, S.; De Cecco, L.; Rinaldi, M.; Comoglio, F.; Favero, F.; Murer, B.; Mutti, L.; Pierotti, M.; Gaudino, G. MicroRNA signature of malignant mesothelioma with potential diagnostic and prognostic implications. Am. J. Respir. Cell. Mol. Biol. 2010, 42, 312–319. [Google Scholar] [CrossRef] [PubMed]
- De Santi, C.; Melaiu, O.; Bonotti, A.; Cascione, L.; Di Leva, G.; Foddis, R.; Cristaudo, A.; Lucchi, M.; Mora, M.; Truini, A.; et al. Deregulation of miRNAs in malignant pleural mesothelioma is associated with prognosis and suggests an alteration of cell metabolism. Sci. Rep. 2017, 7, 3140. [Google Scholar] [CrossRef] [PubMed]
- Kao, S.C.; Cheng, Y.Y.; Williams, M.; Kirschner, M.B.; Madore, J.; Lum, T.; Sarun, K.H.; Linton, A.; McCaughan, B.; Klebe, S.; et al. Tumor Suppressor microRNAs Contribute to the Regulation of PD-L1 Expression in Malignant Pleural Mesothelioma. J. Thorac. Oncol. 2017, 12, 1421–1433. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, M.B.; Cheng, Y.Y.; Armstrong, N.J.; Lin, R.C.; Kao, S.C.; Linton, A.; Klebe, S.; McCaughan, B.C.; van Zandwijk, N.; Reid, G. MiR-score: A novel 6-microRNA signature that predicts survival outcomes in patients with malignant pleural mesothelioma. Mol. Oncol. 2015, 9, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Nabeshima, K.; Hamasaki, M.; Shibuta, T.; Umemura, T. Upregulation of microRNA-31 associates with a poor prognosis of malignant pleural mesothelioma with sarcomatoid component. Med. Oncol. 2014, 31, 303. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, S.V.; Goparaju, C.M.; Lopez, P.; Zavadil, J.; Toren-Haritan, G.; Rosenwald, S.; Hoshen, M.; Chajut, A.; Cohen, D.; Pass, H.I. Pro-tumorigenic effects of miR-31 loss in mesothelioma. J. Biol. Chem. 2010, 285, 22809–22817. [Google Scholar] [CrossRef] [PubMed]
- Moody, H.L.; Lind, M.J.; Maher, S.G. MicroRNA-31 Regulates Chemosensitivity in Malignant Pleural Mesothelioma. Mol. Ther. Nucleic Acids 2017, 8, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.W.; Ferland-McCollough, D.; Jackson, T.J.; Bushell, M. microRNAs in cancer management. Lancet Oncol. 2012, 13, e249–258. [Google Scholar] [CrossRef]
- Enfield, K.S.; Pikor, L.A.; Martinez, V.D.; Lam, W.L. Mechanistic Roles of Noncoding RNAs in Lung Cancer Biology and Their Clinical Implications. Genet. Res. Int. 2012, 2012, 737416. [Google Scholar] [CrossRef] [PubMed]
- Minatel, B.C.; Martinez, V.D.; Ng, K.W.; Sage, A.P.; Tokar, T.; Marshall, E.A.; Anderson, C.; Enfield, K.S.S.; Stewart, G.L.; Reis, P.P.; et al. Large-scale discovery of previously undetected microRNAs specific to human liver. Hum. Genom. 2018, 12, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, E.A.; Sage, A.P.; Ng, K.W.; Martinez, V.D.; Firmino, N.S.; Bennewith, K.L.; Lam, W.L. Small non-coding RNA transcriptome of the NCI-60 cell line panel. Sci. Data 2017, 4, 170157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCall, M.N.; Kim, M.S.; Adil, M.; Patil, A.H.; Lu, Y.; Mitchell, C.J.; Leal-Rojas, P.; Xu, J.; Kumar, M.; Dawson, V.L.; et al. Toward the human cellular microRNAome. Genome Res. 2017, 27, 1769–1781. [Google Scholar] [CrossRef] [PubMed]
- Londin, E.; Loher, P.; Telonis, A.G.; Quann, K.; Clark, P.; Jing, Y.; Hatzimichael, E.; Kirino, Y.; Honda, S.; Lally, M.; et al. Analysis of 13 cell types reveals evidence for the expression of numerous novel primate- and tissue-specific microRNAs. Proc. Natl. Acad. Sci. USA 2015, 112, E1106–1115. [Google Scholar] [CrossRef] [PubMed]
- Quinn, L.; Finn, S.P.; Cuffe, S.; Gray, S.G. Non-coding RNA repertoires in malignant pleural mesothelioma. Lung Cancer 2015, 90, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.M.; Kirschner, M.B.; Cheng, Y.Y.; O’Byrne, K.J.; Gray, S.G.; Schelch, K.; Hoda, M.A.; Klebe, S.; McCaughan, B.; van Zandwijk, N.; et al. Long non-coding RNAs (lncRNAs) are dysregulated in Malignant Pleural Mesothelioma (MPM). PLoS ONE 2013, 8, e70940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felley-Bosco, E.; Rehrauer, H. Non-Coding Transcript Heterogeneity in Mesothelioma: Insights from Asbestos-Exposed Mice. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, E.; Suraokar, M.B.; Rodriguez, J.; Mino, B.; Lin, H.Y.; Rice, D.C.; Tsao, A.; Wistuba, I.I. Frequent coamplification and cooperation between C-MYC and PVT1 oncogenes promote malignant pleural mesothelioma. J. Thorac. Oncol. 2014, 9, 998–1007. [Google Scholar] [CrossRef] [PubMed]
- Renganathan, A.; Kresoja-Rakic, J.; Echeverry, N.; Ziltener, G.; Vrugt, B.; Opitz, I.; Stahel, R.A.; Felley-Bosco, E. GAS5 long non-coding RNA in malignant pleural mesothelioma. Mol. Cancer 2014, 13, 119. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.S.; Heery, R.; Gray, S.G. In Silico and In Vitro Analyses of LncRNAs as Potential Regulators in the Transition from the Epithelioid to Sarcomatoid Histotype of Malignant Pleural Mesothelioma (MPM). Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Parasramka, M.; Yan, I.K.; Wang, X.; Nguyen, P.; Matsuda, A.; Maji, S.; Foye, C.; Asmann, Y.; Patel, T. BAP1 dependent expression of long non-coding RNA NEAT-1 contributes to sensitivity to gemcitabine in cholangiocarcinoma. Mol. Cancer 2017, 16, 22. [Google Scholar] [CrossRef] [PubMed]
- Laury, A.R.; Hornick, J.L.; Perets, R.; Krane, J.F.; Corson, J.; Drapkin, R.; Hirsch, M.S. PAX8 reliably distinguishes ovarian serous tumors from malignant mesothelioma. Am. J. Surg. Pathol. 2010, 34, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhu, Z.; Watabe, K.; Zhang, X.; Bai, C.; Xu, M.; Wu, F.; Mo, Y.Y. Negative regulation of lncRNA GAS5 by miR-21. Cell Death Differ. 2013, 20, 1558–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Source | Number of Cases | Analysis | Platform | References |
|---|---|---|---|---|
| TCGA, Pan Cancer Atlas | 87 MPM tissue samples | RNASeq | Illumina HiSeq 2000 | [28] |
| EGAD00001001914 | 12 MPM cell lines | RNASeq | Illumina HiSeq 2000 | N/A |
| EGAD00001001915 | 211 MPM samples | RNASeq | Illumina HiSeq 2000 | |
| EGAD00001001916 | 207 MPM samples | Targeted Sequencing using SPET | Illumina HiSeq 2000 | |
| International Mesothelioma Program/Brigham and Woman’s Hospital/Harvard Medical School | 4 MPMs, 1 normal control, 1 lung adenocarcinoma (LAC) | Transcriptome Sequencing | Roche/454-pyrosequencing | [31,32] |
| Ospedale Policlinico San Martino (Genova, Italy) | 26 MPM tissue samples, and 3 non-malignant pleura samples | miRNA | Human miRNA Microarray Kit Release 19.0, 8 × 60 K | [33] |
| Brigham and Women’s Hospital/Harvard Medical School | 40 MPM samples, 5 normal pleura, 4 normal lung, 4 MPM cell lines, and 1 non-tumourigenic immortalized mesothelial cell line (SV40) | RNA | Affymetrix Human U133A | [34] |
| University of Vermont, College of Medicine | 4 mesothelial (pleural and peritoneal) cell lines (untreated and treated with asbestos) | RNA-Seq | Illumina HiSeq1000 | [35] |
| Source | Number of Cases | Analysis | Platform | Reference |
|---|---|---|---|---|
| TCGA Pan Cancer Atlas | 87 MPM samples | DNA-Seq, Copy Number | Illumina | [28] |
| NYU Cancer Research | 22 MPM and matched blood samples | Exome Sequencing, Copy Number | Illumina HiSeq | [36] |
| University of Helsinki | 21 malignant mesothelioma; 26 lung adenocarcinoma; 9 normal lung/blood samples of lung adenocarcinoma | Exome Sequencing | Illumina HiSeq | [37] |
| University of California, San Francisco | 1 MPM tissue sample and matched non-malignant tissue | Exome Sequencing | SOLiD 5500 | [38] |
| University of California, San Francisco | 78 MPM tissue samples from 69 MPM patients | Targeted Sequencing | Ion Torrent Personal Genome Machine | [38] |
| University of California, San Diego (Accession: PRJNA278669; ID: 278669) | 7 PeM samples, 7 whole blood samples | Exome Sequencing | Illumina HiSeq 2000 | N/A |
| EGAD00001001913 | 198 MPM Samples | Exome Sequencing | Illumina HiSeq 2500 | N/A |
| EGAD00001000360 | 232 mesothelioma samples | Genome Sequencing, Copy Number | Illumina HiSeq 2000 | N/A |
| EGAS00001002299/EGAS00001002298 | 3 pleural effusions and matched blood samples | Genome Sequencing | Illumina HiSeq X Ten/BGISEQ-500 | [39] |
| EGAD00001001917 | 1 cell line (NCI-H2495) | PacBio | PacBio RS II | N/A |
| The International Mesothelioma Program | 1 MPM sample and matched non-malignant tissue | Genome Sequencing | Illumina Genome Analyzer 2 and Roche/454-pyrosequencing | [40] |
| University of California, San Diego, Moores Cancer Centre | 42 mesothelioma samples (pleural: n = 23; peritoneal: n = 11; pericardial: n = 2; subtype unknown: n = 6) | Genome Sequencing | Illumina HiSeq 2000 | [41] |
| University of Turin | 123 MPM tissue samples | Targeted Sequencing | Ion Torrent Personal Genome Machine | [42] |
| Resource | Description |
|---|---|
| TCGA-MPM Project [43] | A recent analysis of 74 MPM cases with no previous treatment. Multiple high-throughput techniques were performed, including whole exome, mRNA, miRNA, ncRNA sequencing, as well as copy number analyses, DNA methylation, and reverse-phase protein array profiling. Data reveal novel extensive loss of heterozygosity in a subset of MPM cases, high expression of immune-checkpoint molecules, and a high prevalence of BAP1 alterations. |
| National Mesothelioma Virtual Bank [44] | Online databank of mesothelioma biosamples with associated statistics. Full access to the database allows viewing of individual patient clinical data. Tissue and blood samples can also be requested through this database. |
| NCBI ClinVar [45] | Database of human genetic variations that may be clinically relevant. The significance of each genetic variation to any type of disease is assessed, including malignant mesothelioma. Maintained by the National Institutes of Health (NIH), data are publicly available. |
| Classifier | Marker | Sample Type | Analysis | References |
|---|---|---|---|---|
| miR-126 | Early Diagnosis/Prognosis | Serum samples | Low levels of miR-126 could differentiate MPM from healthy individuals, as well as non-small cell lung cancer patients. Low-levels also indicates worse prognosis | [98,99] |
| miR-29c* miR-92a miR-196b | Early Diagnosis | Plasma samples | Higher levels detected in plasma of mesothelioma patients when compared to healthy controls | [100] |
| miR-625-3p | Early Diagnosis | Plasma/serum samples | Higher levels detected in plasma of mesothelioma patients when compared to healthy controls. Also found upregulated in tumor specimens | [100] |
| miR-16 miR-17 miR-486 | Early Diagnosis | Plasma and solid tissue samples | Downregulation in MPM and asbestos-exposed patients when compared to healthy controls | [101] |
| miR-141 miR-200a* miR-200b miR-200c miR-203 miR-205 miR-429 | Diagnosis | Solid tissue samples | Downregulation of the miR-200 family of miRs is able to differentiate MPM from lung adenocarcinomas | [102,103] |
| miR-200c miR-193a-3p miR-192 | Diagnosis | Solid tissue samples | Upregulation of miR-193a and downregulation of miR-200c and miR-192 are able to distinguish MPM from lung adenocarcinomas, adenocarcinomas from the gastrointestinal tract, renal cell carcinomas and other carcinomas | [102] |
| miR-103 | Diagnosis | Peripheral blood samples | Downregulation is able to differentiate mesothelioma patients from asbestos-exposed controls | [104] |
| miR-103a-3p miR-30e-3p | Diagnosis | Plasmatic extracellular vesicles | Expression pattern is able to distinguish MPM from past asbestos-exposed patients | [105] |
| miR-34-b/c | Diagnosis | Serum-circulating DNA | Increased promoter DNA methylation in MPM patients when compared to benign asbestos pleurisy cases and healthy volunteers | [106] |
| miR-126 miR-143 miR-145 miR-652 | Diagnosis | Solid tissue samples | Downregulation is capable of differentiating MPM from the corresponding non-malignant pleura | [107] |
| miR-132-3p | Diagnosis | Plasma samples | Downregulation of circulating miR-132 is able to differentiate mesothelioma patients from asbestos-exposed controls | [108] |
| miR-197-3p miR-1281 miR-32-3p | Diagnosis | Serum samples | Higher circulating levels detected in MPM patients when compared to healthy controls | [109] |
| miR-21 miR-126 | Diagnosis | Cell lines, solid tissue and cytologic specimens | Overexpression of miR-21 and downregulation of miR-126 are able to differentiate mesothelioma from non-neoplastic samples | [110] |
| miR-29c* | Prognosis | Solid tissue samples and cell lines | Increased expression of miR-29c* is associated with the epithelial subtype and able to predict a better prognosis | [111] |
| miR-17-5p miR-21 miR-29a miR-30c miR-30e-5p miR-106a miR-143 | Prognosis | Cell lines and solid tissue samples | Expression pattern is able to distinguish between different mesothelioma histopathological subtypes | [112] |
| miR-17-5p miR-30c | Prognosis | Cell lines and solid tissue samples | Downregulation is associated with better outcome in sarcomatoid mesothelioma patients | [112] |
| let-7c-5p miR-151a-5p | Prognosis | Solid tissue samples | Expression pattern correlate with overall survival and can be used to classify a risk group | [113] |
| miR-15b miR-16 miR-193a-3p miR-195 miR-200c | Prognosis | Solid tissue microarray | Downregulation is associated with increased expression of PD-L1 in MPM, which is a marker of poor prognosis | [114] |
| miR-17-5p miR-19b-3p miR-625-5p | Prognosis | Solid tissue samples | Downregulation is associated with a better prognosis in MPM patients | [115] |
| miR-31 | Prognosis | Solid tissue samples | Downregulation is able to distinguish MPM from reactive mesothelial proliferations. However, higher levels were found in sarcomatoid samples and associate with a worse prognosis. | [116] |
| miR-31 | Prognosis | Cell lines | Downregulation is associated with a worse prognosis and shorter time to tumor recurrence | [117] |
| miR-31 | Prognosis | Cell lines | Upregulation is associated with an intracellular accumulation of platinum, but with a decrease intranuclear concentration promoting chemoresistance | [118] |
| lncRNA | Analyses | Key Findings | References |
|---|---|---|---|
| NEAT1 | In silico analyses; Microarray; RT-qPCR |
| [125,126,130,131] |
| PAX8-AS1 | In silico analyses; Microarray; RT-qPCR |
| [126,132] |
| SNHG7 | In silico analyses; Microarray; RT-qPCR |
| [125,126] |
| PVT1 | In silico analyses; NGS; In vitro siRNA knockdown |
| [125,128,130] |
| GAS5 | In vitro and in silico analyses |
| [129,130,133] |
| EGFR-AS1 | In vitro and in silico analyses |
| [130] |
| PCAT6 | In silico analyses; Microarray; RT-qPCR |
| [130] |
| ZEB2-AS1 | In silico analyses |
| [130] |
| HOTAIR | In silico analyses |
| [130] |
| MORT | In silico analyses |
| [130] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sage, A.P.; Martinez, V.D.; Minatel, B.C.; Pewarchuk, M.E.; Marshall, E.A.; MacAulay, G.M.; Hubaux, R.; Pearson, D.D.; Goodarzi, A.A.; Dellaire, G.; et al. Genomics and Epigenetics of Malignant Mesothelioma. High-Throughput 2018, 7, 20. https://doi.org/10.3390/ht7030020
Sage AP, Martinez VD, Minatel BC, Pewarchuk ME, Marshall EA, MacAulay GM, Hubaux R, Pearson DD, Goodarzi AA, Dellaire G, et al. Genomics and Epigenetics of Malignant Mesothelioma. High-Throughput. 2018; 7(3):20. https://doi.org/10.3390/ht7030020
Chicago/Turabian StyleSage, Adam P., Victor D. Martinez, Brenda C. Minatel, Michelle E. Pewarchuk, Erin A. Marshall, Gavin M. MacAulay, Roland Hubaux, Dustin D. Pearson, Aaron A. Goodarzi, Graham Dellaire, and et al. 2018. "Genomics and Epigenetics of Malignant Mesothelioma" High-Throughput 7, no. 3: 20. https://doi.org/10.3390/ht7030020