A Review of Scrub Typhus (Orientia tsutsugamushi and Related Organisms): Then, Now, and Tomorrow

 ,
,
Abstract
:1. Introduction
2. History and Military Relevance of Scrub Typhus
3. Scrub Typhus Outbreaks
4. Antibiotic Resistance
5. Genotypes and Genome Sequencing
6. Serological Methods for Diagnosing Scrub Typhus
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization. WHO Recommended Surveillance Standards; World Health Organization: Geneva, Switzerland, 1999. [Google Scholar]
- Izzard, L.; Fuller, A.; Blacksell, S.D.; Paris, D.H.; Richards, A.L.; Aukkanit, N.; Nguyen, C.; Jiang, J.; Fenwick, S.; Day, N.P.; et al. Isolation of a novel Orientia species (O. chuto sp. nov.) from a patient infected in Dubai. J. Clin. Microbiol. 2010, 48, 4404–4409. [Google Scholar] [CrossRef] [PubMed]
- Balcells, M.E.; Rabagliati, R.; Garcia, P.; Poggi, H.; Oddo, D.; Concha, M.; Abarca, K.; Jiang, J.; Kelly, D.J.; Richards, A.L.; et al. Endemic scrub typhus-like illness, Chile. Emerg. Infect. Dis. 2011, 17, 1659–1663. [Google Scholar] [CrossRef] [PubMed]
- Dumler, J.S.; Barbet, A.F.; Bekker, C.; Dasch, G.A.; Palmer, G.H.; Ray, S.C.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: Unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and ‘HGE agent’ as subjective synonyms of Ehrlichia phagocytophila. Int. J. Syst. Evol. Microbiol. 2001, 51, 2145–2165. [Google Scholar] [PubMed]
- Odhiambo, A.M.; Maina, A.N.; Taylor, M.L.; Jiang, J.; Richards, A.L. Development and validation of a quantitative real-time polymerase chain reaction assay specific for the detection of Rickettsia felis and not Rickettsia felis-like organisms. Vector-Borne Zoonotic Dis. 2014, 14, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Andersson, J.O.; Andersson, S.G. A century of typhus, lice and rickettsia. Res. Microbiol. 2000, 151, 143–150. [Google Scholar] [CrossRef]
- Duh, D.; Punda-Polic, V.; Avsic-Zupanc, T.; Bouyer, D.; Walker, D.H.; Popov, V.L.; Jelovsek, M.; Gracner, M.; Trilar, T.; Bradaric, N. Rickettsia hoogstraalii sp. nov., isolated from hard-and soft-bodied ticks. Int. J. Syst. Evol. Microbiol. 2010, 60, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Huebner, R.J.; Jellison, W.L.; Armstrong, C. Rickettsialpox: A newly recognized rickettsial disease: V. Recovery of Rickettsia akari from a house mouse (Mus musculus). Public Health Rep. 1947, 62, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Milhano, N.; Palma, M.; Marcili, A.; Núncio, M.S.; de Carvalho, I.L.; de Sousa, R. Rickettsia lusitaniae sp. nov. isolated from the soft tick Ornithodoros erraticus (Acarina: Argasidae). Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Lipovsky, L. Studies of the food habits of postlarval stages of chiggers (Acarina, Trombicullidae). Univ. Kansas Sci. Bull. 1954, 36, 943–958. [Google Scholar]
- Mullen, G.R.; Durden, L.A. Medical and Veterinary Entomology; Academic Press: Cambridge, CA, USA, 2009. [Google Scholar]
- Shatrov, A.B. External morphology of the quiescent instars of trombiculid mites (Acariformes: Trombiculidae) with notes on their moulting processes. Acta Zool. 1999, 80, 85. [Google Scholar]
- Williams, R.W. A contribution to our knowledge of the bionomics of the common North American chigger, Eutrombicula alfreddugesi (Oudemans) with a description of a rapid collecting method. Am. J. Trop. Med. Hyg. 1946, 1, 243–250. [Google Scholar] [CrossRef]
- Krantz, G.W.; Walter, D.E. A Manual of Acarology, 3rd ed.; Texas Tech University Press: Lubbock, TX, USA, 2009; p. 816. [Google Scholar]
- Lerdthusnee, K.; Khlaimanee, N.; Monkanna, T.; Sangjun, N.; Mungviriya, S.; Linthicum, K.J.; Frances, S.P.; Kollars, T.M., Jr.; Coleman, R.E. Efficiency of Leptotrombidium chiggers (Acari: Trombiculidae) at transmitting Orientia tsutsugamushi to laboratory mice. J. Med. Entomol. 2002, 39, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.Z.; Guo, X.G.; Speakman, J.R.; Zuo, S.H.; Wu, D.; Wang, Q.H.; Yang, H. Abundances and host relationships of chigger mites in Yunnan Province, China. Med. Vet. Entomol. 2013, 27, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Domrow, R. Mite parasites of small mammals from scrub typhus foci in Australia. Aust. J. Zool. 1967, 15, 759–798. [Google Scholar] [CrossRef]
- Harrison, J.; Audy, J. Hosts of the mite vector of scrub typhus: I.—A check-list of the recorded hosts. Ann. Trop. Med. Parasitol. 1951, 45, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Rodkvamtook, W.; Gaywee, J.; Kanjanavanit, S.; Ruangareerate, T.; Richards, A.L.; Sangjun, N.; Jeamwattanalert, P.; Sirisopana, N. Scrub typhus outbreak, northern Thailand, 2006–2007. Emerg. Infect. Dis. 2013, 19, 774–777. [Google Scholar] [CrossRef] [PubMed]
- Audy, J.R. Red Mites and Typhus; Athlone Press: London, UK, 1968. [Google Scholar]
- Walker, J.S.; Gan, E.; Chye, C.T.; Muul, I. Involvement of small mammals in the transmission of scrub typhus in Malaysia: Isolation and serological evidence. Trans. R. Soc. Trop. Med. Hyg. 1973, 67, 838–845. [Google Scholar] [CrossRef]
- Traub, R.; Wisseman, C.L., Jr. Review article: The ecology of chigger-borne rickettsiosis (scrub typhus). J. Med. Entomol. 1974, 11, 237–303. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Walker, D.H.; Jupiter, D.; Melby, P.C.; Arcari, C.M. A review of the global epidemiology of scrub typhus. PLoS Negl. Trop. Dis. 2017, 11, e0006062. [Google Scholar] [CrossRef] [PubMed]
- Phasomkusolsil, S.; Tanskul, P.; Ratanatham, S.; Watcharapichat, P.; Phulsuksombati, D.; Frances, S.P.; Lerdthusnee, K.; Linthicum, K.J. Transstadial and transovarial transmission of Orientia tsutsugamushi in Leptotrombidium imphalum and Leptotrombidium chiangraiensis (Acari: Trombiculidae). J. Med. Entomol. 2009, 46, 1442–1445. [Google Scholar] [CrossRef] [PubMed]
- Frances, S.; Watcharapichat, P.; Phulsuksombati, D.; Tanskul, P. Transmission of Orientia tsutsugamushi, the aetiological agent for scrub typhus, to co-feeding mites. Parasitology 2000, 120, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Tilak, R.; Kunwar, R.; Wankhade, U.B.; Tilak, V. Emergence of Schoengastiella ligula as the vector of scrub typhus outbreak in Darjeeling: Has Leptotrombidium deliense been replaced? Indian J. Public Health 2011, 55, 92. [Google Scholar] [CrossRef] [PubMed]
- Brennan, J.M.; Goff, M.L. Keys to the genera of chiggers of the Western Hemisphere (Acarina: Trombiculidae). J. Parasitol. 1977, 63, 554–566. [Google Scholar] [CrossRef] [PubMed]
- Brennan, J.M.; Jones, E.K. Keys to the chiggers of North America with synonymic notes and descriptions of two new genera (Acarina: Trombiculidae). Ann. Entomol. Soc. Am. 1959, 52, 7–16. [Google Scholar] [CrossRef]
- Durden, L.A.; Wilson, N. Ectoparasitic and phoretic arthropods of Virginia opossums (Didelphis virginiana) in central Tennessee. J. Parasitol. 1990, 76, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Weitzel, T.; Dittrich, S.; López, J.; Phuklia, W.; Martinez-Valdebenito, C.; Velásquez, K.; Blacksell, S.D.; Paris, D.H.; Abarca, K. Endemic scrub typhus in South America. N. Engl. J. Med. 2016, 375, 954–961. [Google Scholar] [CrossRef] [PubMed]
- Kocher, C.; Jiang, J.; Morrison, A.C.; Castillo, R.; Leguia, M.; Loyola, S.; Ampuero, J.S.; Cespedes, M.; Halsey, E.S.; Bausch, D.G.; et al. Serologic evidence of scrub typhus in the Peruvian Amazon. Emerg. Infect. Dis. 2017, 23, 1389–1391. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, Y.; Fukatsu, T. Rickettsia infection in natural leech populations. Microb. Ecol. 2005, 49, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, Y.; Sameshima, S.; Kitade, O.; Kojima, J.; Fukatsu, T. Novel clade of Rickettsia spp. from leeches. Appl. Environ. Microbiol. 2002, 68, 999–1004. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.J.; Richards, A.L.; Temenak, J.; Strickman, D.; Dasch, G.A. The past and present threat of rickettsial diseases to military medicine and international public health. Clin. Infect. Dis. 2002, 34, S145–S169. [Google Scholar] [CrossRef] [PubMed]
- Cho, N.-H.; Seong, S.-Y.; Huh, M.-S.; Han, T.-H.; Koh, Y.-S.; Choi, M.-S.; Kim, I.-S. Expression of chemokine genes in murine macrophages infected with Orientia tsutsugamushi. Infect. Immun. 2000, 68, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Sudo, K.; Suzuki, K.; Aoyama, Y.; Nogami, S.; Tanaka, H.; Kawamura, A., Jr. Proliferating sites of Rickettsia tsutsugamushi in mice by different routes of inoculation evidenced with immunofluorescence. Jpn. J. Exp. Med. 1985, 55, 193. [Google Scholar] [PubMed]
- Rikihisa, Y.; Ito, S. Entry of Rickettsia tsutsugamushi into polymorphonuclear leukocytes. Infect. Immun. 1982, 38, 343–350. [Google Scholar] [PubMed]
- Paris, D.H.; Phetsouvanh, R.; Tanganuchitcharnchai, A.; Jones, M.; Jenjaroen, K.; Vongsouvath, M.; Ferguson, D.P.; Blacksell, S.D.; Newton, P.N.; Day, N.P. Orientia tsutsugamushi in human scrub typhus eschars shows tropism for dendritic cells and monocytes rather than endothelium. PLoS Negl. Trop. Dis. 2012, 6, e1466. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-H.; Cheong, T.-C.; Ha, N.-Y.; Ko, Y.; Cho, C.-H.; Jeon, J.-H.; So, I.; Kim, I.-K.; Choi, M.-S.; Kim, I.-S. Orientia tsutsugamushi subverts dendritic cell functions by escaping from autophagy and impairing their migration. PLoS Negl. Trop. Dis. 2013, 7, e1981. [Google Scholar] [CrossRef] [PubMed]
- Seong, S.-Y.; Choi, M.-S.; Kim, I.-S. Orientia tsutsugamushi infection: Overview and immune responses. Microbes Infect. 2001, 3, 11–21. [Google Scholar] [CrossRef]
- Dittrich, S.; Card, E.; Phuklia, W.; Rudgard, W.E.; Silousok, J.; Phoumin, P.; Bouthasavong, L.; Azarian, S.; Davong, V.; Dance, D.A.; et al. Survival and growth of Orientia tsutsugamushi in conventional hemocultures. Emerg. Infect. Dis. 2016, 22, 1460–1463. [Google Scholar] [CrossRef] [PubMed]
- Urakami, H.; Tsuruhara, T.; Tamura, A. Penetration of Rickettsia tsutsugamushi into cultured mouse fibroblasts (L cells): An electron microscopic observation. Microbiol. Immunol. 1983, 27, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Urakami, H.; Tsuruhara, T.; Tamura, A. Electron microscopic studies on intracellular multiplication of Rickettsia tsutsugamushi in L cells. Microbiol. Immunol. 1984, 28, 1191–1201. [Google Scholar] [CrossRef] [PubMed]
- Luksameetanasan, R.; Blacksell, S.D.; Kalambaheti, T.; Wuthiekanun, V.; Chierakul, W.; Chueasuwanchai, S.; Apiwattanaporn, A.; Stenos, J.; Graves, S.; Peacock, S.J. Patient and sample-related factors that effect the success of in vitro isolation of Orientia tsutsugamushi. Southeast Asian J. Trop. Med. Public Health 2007, 38, 91–96. [Google Scholar] [PubMed]
- Urakami, H.; Tsuruhara, T.; Tamura, A. Intranuclear Rickettsia tsutsugamushi in cultured mouse fibroblasts (L cells). Microbiol. Immunol. 1982, 26, 445–447. [Google Scholar] [CrossRef] [PubMed]
- Schaechter, M.; Bozeman, F.M.; Smadel, J.E. Study on the growth of rickettsiae. II. Morphologic observations of living rickettsiae in tissue culture cells. Virology 1957, 3, 160–172. [Google Scholar] [CrossRef]
- Gorvel, L.; Textoris, J.; Banchereau, R.; Amara, A.B.; Tantibhedhyangkul, W.; von Bargen, K.; Mignane, B.K.; Capo, C.; Ghigo, E.; Gorvel, J.-P. Intracellular bacteria interfere with dendritic cell functions: Role of the Type I interferon pathway. PLoS ONE 2014, 9, e99420. [Google Scholar] [CrossRef] [PubMed]
- Atwal, S.; Giengkam, S.; VanNieuwenhze, M.; Salje, J. Live imaging of the genetically intractable obligate intracellular bacteria Orientia tsutsugamushi using a panel of fluorescent dyes. J. Microbiol. Methods 2016, 130, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Siritantikorn, S.; Jintaworn, S.; Noisakran, S.; Suputtamongkol, Y.; Paris, D.H.; Blacksell, S.D. Application of ImageJ program to the enumeration of Orientia tsutsugamushi organisms cultured in vitro. Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 632–635. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.; Gan, E.; Chan, T.; Huxsoll, D. Clinical and immunologic responses of silvered leaf monkeys (Presbytis cristatus) to experimental reinfection with Rickettsia tsutsugamushi. J. Infect. Dis. 1981, 143, 558–561. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.S.; Delacruz, E.C.; Abalos, R.M.; Tan, E.V.; Jiang, J.; Richards, A.L.; Eamsila, C.; Rodkvantook, W.; Myint, K.S.A. Clinical and histological features of inoculation site skin lesions in cynomolgus monkeys experimentally infected with Orientia tsutsugamushi. Vector-Borne Zoonotic Dis. 2007, 7, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, N. Etiology of tsutsugamushi disease. J. Parasitol. 1920, 7, 53–68. [Google Scholar] [CrossRef]
- Fox, J.P. The long persistence of Rickettsia orientalis in the blood and tissues of infected animals. J. Immunol. 1948, 59, 109–114. [Google Scholar] [PubMed]
- Chan, T.; Jiang, J.; Temenak, J.; Richards, A. Development of a rapid method for determining the infectious dose (ID) 50 of Orientia tsutsugamushi in a scrub typhus mouse model for the evaluation of vaccine candidates. Vaccine 2003, 21, 4550–4554. [Google Scholar] [CrossRef]
- Groves, M.; Rosenstreich, D.; Taylor, B.; Osterman, J. Host defenses in experimental scrub typhus: Mapping the gene that controls natural resistance in mice. J. Immunol. 1980, 125, 1395–1399. [Google Scholar] [PubMed]
- Groves, M.G.; Kelly, D.J. Characterization of factors determining Rickettsia tsutsugamushi pathogenicity for mice. Infect. Immun. 1989, 57, 1476–1482. [Google Scholar] [PubMed]
- Groves, M.G.; Osterman, J.V. Host defenses in experimental scrub typhus: Genetics of natural resistance to infection. Infect. immun. 1978, 19, 583–588. [Google Scholar] [PubMed]
- Shirai, A.; Chan, T.C.; Gan, E.; Huxsoll, D.L. Persistence and reactivation of Rickettsia tsutsugamushi infections in laboratory mice. Jpn. J. Med. Sci. Biol. 1979, 32, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Oaks, S., Jr.; Ridgway, R.; Shirai, A.; Twartz, J. Scrub Typhus: Institute for Medical Research, Malaysia: Bulletin No. 21; Institute Penyelidikan Perubatan: Kuala Lampur, Malaysia, 1983. [Google Scholar]
- Ridgway, R.L.; Oaks, S.C., Jr.; LaBarre, D.D. Laboratory animal models for human scrub typhus. Lab. Anim. Sci. 1986, 36, 481–485. [Google Scholar] [PubMed]
- Soong, L.; Mendell, N.L.; Olano, J.P.; Rockx-Brouwer, D.; Xu, G.; Goez-Rivillas, Y.; Drom, C.; Shelite, T.R.; Valbuena, G.; Walker, D.H. An intradermal inoculation mouse model for immunological investigations of acute scrub typhus and persistent infection. PLoS Negl. Trop. Dis. 2016, 10, e0004884. [Google Scholar] [CrossRef] [PubMed]
- Sunyakumthorn, P.; Paris, D.H.; Chan, T.-C.; Jones, M.; Luce-Fedrow, A.; Chattopadhyay, S.; Jiang, J.; Anantatat, T.; Turner, G.D.; Day, N.P. An intradermal inoculation model of scrub typhus in Swiss CD-1 mice demonstrates more rapid dissemination of virulent strains of Orientia tsutsugamushi. PLoS ONE 2013, 8, e54570. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wen, B.; Wen, B.; Niu, D.; Chen, M.; Qiu, L. Induction of protective immunity against scrub typhus with a 56-kilodalton recombinant antigen fused with a 47-kilodalton antigen of Orientia tsutsugamushi Karp. Am. J. Trop. Med. Hyg. 2005, 72, 458–464. [Google Scholar] [PubMed]
- Shelite, T.R.; Saito, T.B.; Mendell, N.L.; Gong, B.; Xu, G.; Soong, L.; Valbuena, G.; Bouyer, D.H.; Walker, D.H. A hematogenously disseminated Orientia tsutsugamsushi-infected murine model of scrub typhus. PLoS Negl. Trop. Dis. 2014, 8, e2966. [Google Scholar] [CrossRef] [PubMed]
- Kundin, W.; Liu, C.; Harmon, P.; Rodina, P. Pathogenesis of scrub typhus infection (Rickettsia tsutsugamushi) as studied by immunofluorescence. J. Immunol. 1964, 93, 772–781. [Google Scholar] [PubMed]
- Xu, G.; Mendell, N.L.; Liang, Y.; Shelite, T.R.; Goez-Rivillas, Y.; Soong, L.; Bouyer, D.H.; Walker, D.H. CD8+ T cells provide immune protection against murine disseminated endotheliotropic Orientia tsutsugamushi infection. PLoS Negl. Trop. Dis. 2017, 11, e0005763. [Google Scholar]
- Keller, C.A.; Hauptmann, M.; Kolbaum, J.; Gharaibeh, M.; Neumann, M.; Glatzel, M.; Fleischer, B. Dissemination of Orientia tsutsugamushi and inflammatory responses in a murine model of scrub typhus. PLoS Negl. Trop. Dis. 2014, 8, e3064. [Google Scholar] [CrossRef] [PubMed]
- Lurchachaiwong, W.; Monkanna, T.; Leepitakrat, S.; Ponlawat, A.; Sattabongkot, J.; Schuster, A.L.; McCardle, P.W.; Richards, A.L. Variable clinical responses of a scrub typhus outbred mouse model to feeding by Orientia tsutsugamushi infected mites. Exp. Appl. Acarol. 2012, 58, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Peter, J.V.; Sudarsan, T.I.; Prakash, J.A.J.; Varghese, G.M. Severe scrub typhus infection: Clinical features, diagnostic challenges and management. World J. Crit. Care Med. 2015, 4, 244. [Google Scholar] [CrossRef] [PubMed]
- Biggs, H.M. Diagnosis and management of tickborne rickettsial diseases: Rocky Mountain spotted fever and other spotted fever group rickettsioses, ehrlichioses, and anaplasmosis—United States. MMWR Recomm. Rep. 2016, 65, 1–44. [Google Scholar] [CrossRef] [PubMed]
- Cascio, A.; Giordano, S.; Dones, P.; Venezia, S.; Iaria, C.; Ziino, O. Haemophagocytic syndrome and rickettsial diseases. J. Med. Microbiol. 2011, 60, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.J.; Yoo, I.H.; Lee, J.W.; Chung, N.G.; Cho, B.; Kim, H.K.; Kang, J.H. Life-threatening scrub typhus with hemophagocytosis and acute respiratory distress syndrome in an infant. J. Trop. Pediatr. 2013, 59, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Sankhyan, N.; Saptharishi, L.G.; Sasidaran, K.; Kanga, A.; Singhi, S.C. Clinical profile of scrub typhus in children and its association with hemophagocytic lymphohistiocytosis. Indian Pediatr. 2014, 51, 651–653. [Google Scholar] [CrossRef] [PubMed]
- Strickman, D.; Sheer, T.; Salata, K.; Hershey, J.; Dasch, G.; Kelly, D.; Kuschner, R. In vitro effectiveness of azithromycin against doxycycline-resistant and-susceptible strains of Rickettsia tsutsugamushi, etiologic agent of scrub typhus. Antimicrob. Agents Chemother. 1995, 39, 2406–2410. [Google Scholar] [CrossRef] [PubMed]
- Ives, T.J.; Manzewitsch, P.; Regnery, R.L.; Butts, J.D.; Kebede, M. In vitro susceptibilities of Bartonella henselae, B. quintana, B. elizabethae, Rickettsia rickettsii, R. conorii, R. akari, and R. prowazekii to macrolide antibiotics as determined by immunofluorescent-antibody analysis of infected Vero cell monolayers. Antimicrob. Agents Chemother. 1997, 41, 578–582. [Google Scholar] [PubMed]
- Kim, Y.-S.; Lee, H.J.; Chang, M.; Son, S.K.; Rhee, Y.E.; Shim, S.K. Scrub typhus during pregnancy and its treatment: A case series and review of the literature. Am. J. Trop. Med. Hyg. 2006, 75, 955–959. [Google Scholar] [PubMed]
- Phimda, K.; Hoontrakul, S.; Suttinont, C.; Chareonwat, S.; Losuwanaluk, K.; Chueasuwanchai, S.; Chierakul, W.; Suwancharoen, D.; Silpasakorn, S.; Saisongkorh, W. Doxycycline versus azithromycin for treatment of leptospirosis and scrub typhus. Antimicrob. Agents Chemother. 2007, 51, 3259–3263. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.J.; Fuerst, P.A.; Richards, A.L. The historical case for and the future study of antibiotic-resistant scrub typhus. Trop. Med. Infect. Dis. 2017, 2, 63. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Richards, A.L. Scrub typhus vaccines: Past history and recent developments. Hum. Vaccines 2007, 3, 73–80. [Google Scholar] [CrossRef]
- Kelly, D.J.; Fuerst, P.A.; Ching, W.-M.; Richards, A.L. Scrub typhus: The geographic distribution of phenotypic and genotypic variants of Orientia tsutsugamushi. Clin. Infect. Dis. 2009, 48, S203–S230. [Google Scholar] [CrossRef] [PubMed]
- Zinsser, H. Rats, Lice and History; Little, Brown, and Co., Inc.: Boston, MA, USA, 1934. [Google Scholar]
- Bayne-Jones, S. Epidemic Typhus in the Mediterranean Area during World War II with Special Reference to the Control of the Epidemic in Naples in the Winter of 1943–1944; Soule, M.H., Ed.; American Association for the Advancement of Science: Washington, DC, USA, 1948; pp. 1–15. [Google Scholar]
- Philip, C.B. Tsutsugamushi disease (scrub typhus) in World War II. J. Parasitol. 1948, 34, 169–191. [Google Scholar] [CrossRef] [PubMed]
- Maxcy, K.F. Scrub Typhus (Tsutsugamushi Disease) in the US Army during World War II; Soule, M.H., Ed.; American Association for the Advancement of Science: Washington, DC, USA, 1948; pp. 36–50. [Google Scholar]
- Kawamura, R. Studies on Tsutsugamushi Disease (Japanese Blood Fever); Spokesman Printing Company: Cincinnati, OH, USA, 1926; Volume 4. [Google Scholar]
- Mackie, T.T.; Davis, G.E.; Fuller, H.S.; Knapp, J.A.; Steinacker, M.L.; Stager, K.E.; Traub, R.; Jellison, W.L.; Millspaugh, D.D.; Austrian, R.C. Observations on tsutsugamusi disease (scrub typhus) in Assam and Burma: Preliminary report. Am. J. Epidemiol. 1946, 43, 195–218. [Google Scholar] [CrossRef]
- Philip, C.B. Scrub typhus and scrub itch. Commun. Dis. 1964, 2, 275–348. [Google Scholar]
- Woodward, T.E. Introduction to the history of the Armed Forces Medical Unit in Kuala Lampur, Malaya, and the Armed Forces Research Institute of Medical Sciences (AFRIMS) in Bangkok, Thailand; Lecture presented at the Walter Army Research Institute; Walter Army Research Institute: Washington, DC, USA, 1987. [Google Scholar]
- Berman, S.J.; Kundin, W.D. Scrub typhus in South Vietnam: A study of 87 cases. Ann. Intern. Med. 1973, 79, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Watt, G.; Chouriyagune, C.; Ruangweerayud, R.; Watcharapichat, P.; Phulsuksombati, D.; Jongsakul, K.; Teja-Isavadharm, P.; Bhodhidatta, D.; Corcoran, K.D.; Dasch, G.A. Scrub typhus infections poorly responsive to antibiotics in northern Thailand. Lancet 1996, 348, 86–89. [Google Scholar] [CrossRef]
- Jiang, J.; Chan, T.-C.; Temenak, J.J.; Dasch, G.A.; Ching, W.-M.; Richards, A.L. Development of a quantitative real-time polymerase chain reaction assay specific for Orientia tsutsugamushi. Am. J. Trop. Med. Hyg. 2004, 70, 351–356. [Google Scholar] [PubMed]
- Coleman, R.E.; Monkanna, T.; Linthicum, K.J.; Strickman, D.A.; Frances, S.P.; Tanskul, P.; Kollars, T.M., Jr.; Inlao, I.; Watcharapichat, P.; Khlaimanee, N.; et al. Occurrence of Orientia tsutsugamushi in small mammals from Thailand. Am. J. Trop. Med. Hyg. 2003, 69, 519–524. [Google Scholar] [PubMed]
- Maina, A.N.; Farris, C.M.; Odhiambo, A.; Jiang, J.; Laktabai, J.; Armstrong, J.; Holland, T.; Richards, A.L.; O’Meara, W.P. Q fever, scrub typhus, and rickettsial diseases in children, Kenya, 2011–2012. Emerg. Infect. Dis. 2016, 22, 883–886. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Richards, A.L. Scrub typhus: No longer restricted to the tsutsugamushi triangle. Trop. Med. Infect. Dis. 2017, 2. Accepted. [Google Scholar]
- Mahajan, S.K.; Rolain, J.M.; Sankhyan, N.; Kaushal, R.K.; Raoult, D. Pediatric scrub typhus in Indian Himalayas. Indian J. Pediatr. 2008, 75, 947–949. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.J.; Paris, D.H.; Newton, P.N. A systematic review of mortality from untreated scrub typhus (Orientia tsutsugamushi). PLoS Negl. Trop. Dis. 2015, 9, e0003971. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.D.; Yousuf, A.A.; Lerdthusnee, K.; Razee, A.; Chandranoi, K.; Jones, J.W. Scrub typhus reemergence in the Maldives. Emerg. Infect. Dis. 2003, 9, 1638–1641. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Ha, N.-Y.; Min, C.-K.; Kim, H.-I.; Yen, N.-T.; Lee, K.-H.; Oh, I.; Kang, J.-S.; Choi, M.-S.; Kim, I.-S.; et al. Diversification of Orientia tsutsugamushi genotypes by intragenic recombination and their potential expansion in endemic areas. PLoS Negl. Trop. Dis. 2017, 11, e0005408. [Google Scholar] [CrossRef] [PubMed]
- Usha, K.; Kumar, E.; Kalawat, U.; Kumar, B.S.; Chaudhury, A.; Gopal, D.V. Molecular characterization of Orientia tsutsugamushi serotypes causing scrub typhus outbreak in southern region of Andhra Pradesh, India. Indian J. Med. Res. 2016, 144, 597–603. [Google Scholar] [PubMed]
- Bonell, A.; Lubell, Y.; Newton, P.N.; Crump, J.A.; Paris, D.H. Estimating the burden of scrub typhus: A systematic review. PLoS Negl. Trop. Dis. 2017, 11, e0005838. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.K. Rickettsial diseases. J. Assoc. Physicians India 2012, 60, 37–44. [Google Scholar] [PubMed]
- Harris, P.N.A.; Oltvolgyi, C.; Islam, A.; Hussain-Yusuf, H.; Loewenthal, M.R.; Vincent, G.; Stenos, J.; Graves, S. An outbreak of scrub typhus in military personnel despite protocols for antibiotic prophylaxis: Doxycycline resistance excluded by a quantitative PCR-based susceptibility assay. Microbes Infect. 2016, 18, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Tan, Z.; Ren, D.; Zhang, X.; He, Y.; Bao, C.; Liu, D.; Yi, Q.; Qian, W.; Yin, J.; et al. Clinical characteristics and risk factors of an outbreak with scrub typhus in previously unrecognized areas, Jiangsu Province, China 2013. PLoS ONE 2015, 10, e0125999. [Google Scholar] [CrossRef] [PubMed]
- Tshokey, T.; Graves, S.; Tshering, D.; Phuntsho, K.; Tshering, K.; Stenos, J. Scrub typhus outbreak in a remote primary school, Bhutan, 2014. Emerg. Infect. Dis. 2017, 23, 1412–1414. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Luo, L.; Jing, Q.; Li, X.; Huang, Y.; Xiao, X.; Liu, L.; Wu, X.; Yang, Z. A city park as a potential epidemic site of scrub typhus: A case-control study of an outbreak in Guangzhou, China. Parasites Vectors 2014, 7, 513. [Google Scholar] [PubMed]
- Sharma, N.; Biswal, M.; Kumar, A.; Zaman, K.; Jain, S.; Bhalla, A. Scrub typhus in a tertiary care hospital in North India. Am. J. Trop. Med. Hyg. 2016, 95, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Vivekanandan, M.; Mani, A.; Priya, Y.S.; Singh, A.P.; Jayakumar, S.; Purty, S. Outbreak of scrub typhus in Pondicherry. J. Assoc. Physicians India 2010, 58, 24–28. [Google Scholar] [PubMed]
- Dass, R.; Deka, N.M.; Duwarah, S.G.; Barman, H.; Hoque, R.; Mili, D.; Barthakur, D. Characteristics of pediatric scrub typhus during an outbreak in the north eastern region of India: Peculiarities in clinical presentation, laboratory findings and complications. Indian J. Pediatr. 2011, 78, 1365–1370. [Google Scholar] [CrossRef] [PubMed]
- Sivarajan, S.; Shivalli, S. Clinical and paraclinical profile, and predictors of outcome in 90 cases of scrub typhus, Meghalaya, India. Infect. Dis. Poverty 2016, 5, 91. [Google Scholar] [CrossRef] [PubMed]
- Gurung, S.; Pradhan, J.; Bhutia, P.Y. Outbreak of scrub typhus in the north east Himalayan region-Sikkim: An emerging threat. Indian J. Med. Microbiol. 2013, 31, 72–74. [Google Scholar] [CrossRef] [PubMed]
- Stephen, S.; Sangeetha, B.; Ambroise, S.; Sarangapani, K.; Gunasekaran, D.; Hanifah, M.; Somasundaram, S. Outbreak of scrub typhus in Puducherry & Tamil Nadu during cooler months. Indian J. Med. Res. 2015, 142, 591–597. [Google Scholar] [PubMed]
- Sinha, P.; Gupta, S.; Dawra, R.; Rijhawan, P. Recent outbreak of scrub typhus in north western part of India. Indian J. Med. Microbiol. 2014, 32, 247–250. [Google Scholar] [PubMed]
- Krishna, M.R.; Vasuki, B.; Nagaraju, K. Scrub typhus: Audit of an outbreak. Indian J. Pediatr. 2015, 82, 537–540. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Singh, S.P.; Ahmad, N. A study of etiological pattern in an epidemic of acute febrile illness during monsoon in a tertiary health care institute of Uttarakhand, India. J. Clin. Diag. Res. JCDR 2014, 8, Mc01. [Google Scholar] [CrossRef] [PubMed]
- Takhar, R.P.; Bunkar, M.L.; Arya, S.; Mirdha, N.; Mohd, A. Scrub typhus: A prospective, observational study during an outbreak in Rajasthan, India. Natl. Med. J. India 2017, 30, 69–72. [Google Scholar] [PubMed]
- Subbalaxmi, M.V.; Madisetty, M.K.; Prasad, A.K.; Teja, V.D.; Swaroopa, K.; Chandra, N.; Upadhyaya, A.C.; Shetty, M.; Rao, M.N.; Raju, Y.S.; et al. Outbreak of scrub typhus in Andhra Pradesh—Experience at a tertiary care hospital. J. Assoc. Physicians India 2014, 62, 490–496. [Google Scholar] [PubMed]
- Singh, S.I.; Devi, K.P.; Tilotama, R.; Ningombam, S.; Gopalkrishna, Y.; Singh, T.B.; Murhekar, M.V. An outbreak of scrub typhus in Bishnupur District of Manipur, India, 2007. Trop. Dr. 2010, 40, 169–170. [Google Scholar] [CrossRef] [PubMed]
- 59 New Cases of Scrub Typhus in Chitwan. 2016. Available online: www.myrepublica.com/news/7173/ (accessed on 10 October 2016).
- Bastola, A.P.N. An outbreak of scrub typhus in Nepal following the 2015 Gorkha earthquake. Int. J. Infect. Dis. 2016, 53, 55. [Google Scholar] [CrossRef]
- Marks, M.; Joshua, C.; Longbottom, J.; Longbottom, K.; Sio, A.; Puiahi, E.; Jilini, G.; Stenos, J.; Dalipanda, T.; Musto, J. An outbreak investigation of scrub typhus in Western Province, Solomon Islands, 2014. West. Pac. Surveill. Response J. WPSAR 2016, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Krishna, V.P.; Manjunath, S.H.; Shrivastava, S.; Singh, V.; Dariya, S.S.; Soni, M.; Sharma, S. Analysis of two outbreaks of scrub typhus in Rajasthan: A clinico-epidemiological study. J. Assoc. Physicians India 2014, 62, 24–29. [Google Scholar] [PubMed]
- Likeman, R.K. Scrub typhus: A recent outbreak among military personnel in North Queensland. ADH Health 2006, 7, 10–13. [Google Scholar]
- McBride, W.J.; Taylor, C.T.; Pryor, J.A.; Simpson, J.D. Scrub typhus in North Queensland. Med. J. Aust. 1999, 170, 318–320. [Google Scholar] [PubMed]
- Chakraborty, S.; Sarma, N. Scrub typhus: An emerging threat. Indian J. Dermatol. 2017, 62, 478–485. [Google Scholar] [PubMed]
- St John, H.K.; Adams, M.L.; Masuoka, P.M.; Flyer-Adams, J.G.; Jiang, J.; Rozmajzl, P.J.; Stromdahl, E.Y.; Richards, A.L. Prevalence, distribution, and development of an ecological niche model of Dermacentor variabilis ticks positive for Rickettsia montanensis. Vector-Borne Zoonotic Dis. 2016, 16, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, J.A.; Dahm, N.M.; Ruiz, M.O.; Brown, W.M. Temporal and spatial distribution of tick-borne disease cases among humans and canines in Illinois (2000–2009). Environ. Health Insights 2014, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Ratmanov, P.; Mediannikov, O.; Raoult, D. Vectorborne diseases in West Africa: Geographic distribution and geospatial characteristics. Trans. R. Soc. Trop. Med. Hyg. 2013, 107, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, R.; Musella, V.; Descalzo, M.A.; Montoya, A.; Checa, R.; Marino, V.; Martín, O.; Cringoli, G.; Rinaldi, L.; Miró, G. Modelling the current distribution and predicted spread of the flea species Ctenocephalides felis infesting outdoor dogs in Spain. Parasites Vectors 2017, 10, 428. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.J.; Foley, D.H.; Richards, A.L. A spatiotemporal database to track human scrub typhus using the VectorMap application. PLoS Negl. Trop. Dis. 2015, 9, e0004161. [Google Scholar] [CrossRef] [PubMed]
- Cosson, J.F.; Galan, M.; Bard, E.; Razzauti, M.; Bernard, M.; Morand, S.; Brouat, C.; Dalecky, A.; Bâ, K.; Charbonnel, N. Detection of Orientia sp. DNA in rodents from Asia, West Africa and Europe. Parasites Vectors 2015, 8, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huxsoll, D.; Shirai, A.; Robinson, D.; Yap, L.; Lim, B. Presence of antibodies to scrub typhus and murine typhus in dogs from Selangor, peninsular, Malaysia. Southeast Asian J. Trop. Med. Public Health 1977, 8, 232–235. [Google Scholar] [PubMed]
- Smadel, J.E.; Woodward, T.E.; Ley, H.L., Jr.; Lewthwaite, R. Chloramphenicol (chloromycetin) in the treatment of tsutsugamushi disease (scrub typhus). J. Clin. Investig. 1949, 28, 1196. [Google Scholar] [CrossRef] [PubMed]
- Luce-Fedrow, A.; Mullins, K.; Kostik, A.P.; St John, H.K.; Jiang, J.; Richards, A.L. Strategies for detecting rickettsiae and diagnosing rickettsial diseases. Future Microbiol. 2015, 10, 537–564. [Google Scholar] [CrossRef] [PubMed]
- Smadel, J.E.; Woodward, T.E.; Ley, H.L., Jr.; Philip, C.B.; Traub, R.; Lewthwaite, R.; Savoor, S. Chloromycetin in the treatment of scrub typhus. Science 1948, 108, 160–161. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.; Saunders, J.; Singh, S.; Huxsoll, D.; Shirai, A. Single dose doxycycline therapy for scrub typhus. Trans. R. Soc. Trop. Med. Hyg. 1978, 72, 412–416. [Google Scholar] [CrossRef]
- Rajapakse, S.; Rodrigo, C.; Fernando, S.D. Drug treatment of scrub typhus. Trop. Dr. 2011, 41, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-H.; Lee, C.; Chang, W.H.; Choi, S.W.; Choi, J.E.; Kim, Y.S.; Cho, S.R.; Ryu, J.; Pai, C.H. Short-course doxycycline treatment versus conventional tetracycline therapy for scrub typhus: A multicenter randomized trial. Clin. Infect. Dis. 1995, 21, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Panpanich, R. Antibiotics for treating scrub typhus. Cochrane Libr. 2002. [Google Scholar] [CrossRef] [PubMed]
- Alanis, A.J. Resistance to antibiotics: Are we in the post-antibiotic era? Arch. Med. Res. 2005, 36, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Corwin, A.; Soderquist, R.; Suwanabun, N.; Sattabongkot, J.; Martin, L.; Kelly, D.; Beecham, J. Scrub typhus and military operations in Indochina. Clin. Infect. Dis. 1999, 29, 940–941. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Marienau, K.J.; May, L.A.; Beecham, H.J., III; Wilkinson, R.; Ching, W.-M.; Richards, A.L. Laboratory diagnosis of two scrub typhus outbreaks at Camp Fuji, Japan in 2000 and 2001 by enzyme-linked immunosorbent assay, rapid flow assay, and Western blot assay using outer membrane 56-kd recombinant proteins. Am. J. Trop. Med. Hyg. 2003, 69, 60–66. [Google Scholar] [PubMed]
- Rodkvamtook, W.; Ruang-areerate, T.; Gaywee, J.; Richards, A.L.; Jeamwattanalert, P.; Bodhidatta, D.; Sangjun, N.; Prasartvit, A.; Jatisatienr, A.; Jatisatienr, C. Isolation and characterization of Orientia tsutsugamushi from rodents captured following a scrub typhus outbreak at a military training base, Bothong district, Chonburi Province, Central Thailand. Am. J. Trop. Med. Hyg. 2011, 84, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Silpapojakul, K. Scrub typhus in the western pacific region. Ann. Acad. Med. 1997, 26, 794–800. [Google Scholar]
- Kim, M.S.; Baek, J.H.; Lee, J.-S.; Chung, M.-H.; Lee, S.M.; Kang, J.-S. High in vitro infectivity of a doxycycline-insensitive strain of Orientia tsutsugamushi. Infect. Chemother. 2013, 45, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Chung, E.; Kim, E.; Sea, J. A case of doxycycline-resistant tsutsugamushi meningoencephalitis. Neurol Asia 2014, 19, 205–206. [Google Scholar]
- Mathai, E.; Rolain, J.; Verghese, G.; Abraham, O.; Mathai, D.; Mathai, M.; Raoult, D. Outbreak of scrub typhus in southern India during the cooler months. Ann. N. Y. Acad. Sci. 2003, 990, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Tantibhedhyangkul, W.; Angelakis, E.; Tongyoo, N.; Newton, P.N.; Moore, C.E.; Phetsouvanh, R.; Raoult, D.; Rolain, J.-M. Intrinsic fluoroquinolone resistance in Orientia tsutsugamushi. Int. J. Antimicrob. Agents 2010, 35, 338–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thipmontree, W.; Tantibhedhyangkul, W.; Silpasakorn, S.; Wongsawat, E.; Waywa, D.; Suputtamongkol, Y. Scrub typhus in northeastern Thailand: Eschar distribution, abnormal electrocardiographic findings, and predictors of fatal outcome. Am. J. Trop. Med. Hyg. 2016, 95, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Watt, G.; Kantipong, P.; Jongsakul, K.; Watcharapichat, P.; Phulsuksombati, D.; Strickman, D. Doxycycline and rifampicin for mild scrub-typhus infections in northern Thailand: A randomised trial. Lancet 2000, 356, 1057–1061. [Google Scholar] [CrossRef]
- Smadel, J.E.; Ley, H.L., Jr.; Diercks, F.H.; Cameron, J. Persistence of Rickettsia tsutsugamushi in tissues of patients recovered from scrub typhus. Am. J. Hyg. 1952, 56, 294–302. [Google Scholar] [PubMed]
- Silpapojakul, K.; Ukkachoke, C.; Krisanapan, S.; Silpapojakul, K. Rickettsial meningitis and encephalitis. Arch. Intern. Med. 1991, 151, 1753–1757. [Google Scholar] [CrossRef] [PubMed]
- Rajapakse, S.; Rodrigo, C.; Fernando, D. Scrub typhus: Pathophysiology, clinical manifestations and prognosis. Asian Pac. J. Trop. Med. 2012, 5, 261–264. [Google Scholar] [CrossRef]
- Angelakis, E.; Biswas, S.; Taylor, C.; Raoult, D.; Rolain, J.-M. Heterogeneity of susceptibility to fluoroquinolones in Bartonella isolates from Australia reveals a natural mutation in gyrA. J. Antimicrob. Chemother. 2008, 61, 1252–1255. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Raoult, D.; Rolain, J.-M. A bioinformatic approach to understanding antibiotic resistance in intracellular bacteria through whole genome analysis. Int. J. Antimicrob. Agents 2008, 32, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Heisig, P.; Tschorny, R. Characterization of fluoroquinolone-resistant mutants of Escherichia coli selected in vitro. Antimicrob. Agents Chemother. 1994, 38, 1284–1291. [Google Scholar] [CrossRef] [PubMed]
- Minnick, M.F.; Wilson, Z.R.; Smitherman, L.S.; Samuels, D.S. gyrA mutations in ciprofloxacin-resistant Bartonella bacilliformis strains obtained in vitro. Antimicrob. Agents Chemother. 2003, 47, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Jee, H.-G.; Chung, M.-H.; Lee, S.-G.; Kim, I.-S.; Chang, V.-H. Transmission of scrub typhus by needlestick from a patient receiving pefloxacin. Scand. J. Infect. Dis. 1996, 28, 411–412. [Google Scholar] [CrossRef] [PubMed]
- Mathai, E.; Rolain, J.; Verghese, L.; Mathai, M.; Jasper, P.; Verghese, G.; Raoult, D. Scrub typhus during pregnancy in India. Trans. R. Soc. Trop. Med. Hyg. 2003, 97, 570–572. [Google Scholar] [CrossRef]
- Tsai, C.-C.; Lay, C.-J.; Wang, C.-L.; Ho, Y.-H.; Wang, L.-S.; Chen, L.-K. Levofloxacin versus tetracycline antibiotics for the treatment of scrub typhus. Int. J. Infect. Dis. 2010, 14, e62–e67. [Google Scholar] [CrossRef] [PubMed]
- Cracco, C.; Delafosse, C.; Baril, L.; Lefort, Y.; Morelot, C.; Derenne, J.-P.; Bricaire, F.; Similowski, T. Multiple organ failure complicating probable scrub typhus. Clin. Infect. Dis. 2000, 31, 191–192. [Google Scholar] [CrossRef] [PubMed]
- Eaton, M.; Cohen, M.T.; Shlim, D.R.; Innes, B. Ciprofloxacin treatment of typhus. JAMA 1989, 262, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-C.; Ko, W.-C.; Lee, H.-L.; Chen, H.-Y. Clinical manifestations and complications of rickettsiosis in southern Taiwan. J. Formos. Med. Assoc. 2002, 101, 385–392. [Google Scholar] [PubMed]
- Cho, N.H.; Kim, H.R.; Lee, J.H.; Kim, S.Y.; Kim, J.; Cha, S.; Darby, A.C.; Fuxelius, H.H.; Yin, J.; Kim, J.H.; et al. The Orientia tsutsugamushi genome reveals massive proliferation of conjugative type IV secretion system and host-cell interaction genes. Proc. Natl. Acad. Sci. USA 2007, 104, 7981–7986. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.-M.; Chao, C.-C.; Lei, H.; Li, B.; Tsai, S.; Hung, G.-C.; Ching, W.-M.; Lo, S.-C. Genomic sequencing of Orientia tsutsugamushi strain Karp, an assembly comparable to the genome size of the strain Ikeda. Genome Announc. 2016, 4, e00702–e00716. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.-M.; Chao, C.-C.; Lei, H.; Li, B.; Tsai, S.; Hung, G.-C.; Ching, W.-M.; Lo, S.-C. Intraspecies comparative genomics of three strains of Orientia tsutsugamushi with different antibiotic sensitivity. Genomics Data 2017, 12, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.; Yamashita, A.; Kurokawa, K.; Morimoto, T.; Ogawa, M.; Fukuhara, M.; Urakami, H.; Ohnishi, M.; Uchiyama, I.; Ogura, Y.; et al. The whole-genome sequencing of the obligate intracellular bacterium Orientia tsutsugamushi revealed massive gene amplification during reductive genome evolution. DNA Res. 2008, 15, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Shishido, A. Strain variation of Rickettsia orientalis in the complement fixation test. Jpn. J. Med. Sci. Biol. 1964, 17, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Shishido, A. Identification and serological classification of causative agent of scrub typhus in Japan. Jpn. J. Med. Sci. Biol. 1962, 15, 308. [Google Scholar]
- Dohany, A.L.; Shirai, A.; Robinson, D.M.; Ram, S.; Huxsoll, D.L. Identification and antigenic typing of Rickettsia tsutsugamushi in naturally infected chiggers (Acarina: Trombiculidae) by direct immunofluorescence. Am. J. Trop. Med. Hyg. 1978, 27, 1261–1264. [Google Scholar] [CrossRef] [PubMed]
- Shirai, A.; Robinson, D.M.; Brown, G.W.; Gan, E.; Huxsoll, D.L. Antigenic analysis by direct immunofluorescence of 114 isolates of Rickettsia tsutsugamushi recovered from febrile patients in rural Malaysia. Jpn. J. Med. Sci. Biol. 1979, 32, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Kawabata, N.; Tamura, A.; Urakami, H.; Ohashi, N.; Murata, M.; Yoshida, Y.; Kawamura, A. Immunological properties of Rickettsia tsutsugamushi Kawasaki strain, isolated from a patient in Kyushu. Microbiol. Immunol. 1986, 30, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Tamura, A.; Ohashi, N.; Urakami, H.; Takahashi, K.; Oyanagi, M. Analysis of polypeptide composition and antigenic components of Rickettsia tsutsugamushi by polyacrylamide gel electrophoresis and immunoblotting. Infect. Immun. 1985, 48, 671–675. [Google Scholar] [PubMed]
- Ohashi, N.; Tamura, A.; Sakurai, H.; Yamamoto, S. Characterization of a new antigenic type, Kuroki, of Rickettsia tsutsugamushi isolated from a patient in Japan. J. Clin. Microbiol. 1990, 28, 2111–2113. [Google Scholar] [PubMed]
- Ohashi, N.; Nashimoto, H.; Ikeda, H.; Tamura, A. Diversity of immunodominant 56-kda type-specific antigen (TSA) of Rickettsia tsutsugamushi. Sequence and comparative analyses of the genes encoding TSA homologues from four antigenic variants. J. Biol. Chem. 1992, 267, 12728–12735. [Google Scholar] [PubMed]
- Ohashi, N.; Nashimoto, H.; Ikeda, H.; Tamura, A. Cloning and sequencing of the gene (tsg56) encoding a type-specific antigen from Rickettsia tsutsugamushi. Gene 1990, 91, 119–122. [Google Scholar] [CrossRef]
- Stover, C.; Marana, D.; Carter, J.; Roe, B.; Mardis, E.; Oaks, E. The 56-kilodalton major protein antigen of Rickettsia tsutsugamushi: Molecular cloning and sequence analysis of the sta56 gene and precise identification of a strain-specific epitope. Infect. Immun. 1990, 58, 2076–2084. [Google Scholar] [PubMed]
- Ohashi, N.; Koyama, Y.; Urakami, H.; Fukuhara, M.; Tamura, A.; Kawamori, F.; Yamamoto, S.; Kasuya, S.; Yoshimura, K. Demonstration of antigenic and genotypic variation in Orientia tsutsugamushi which were isolated in Japan, and their classification into type and subtype. Microbiol. Immunol. 1996, 40, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, H.; Murai, K.; Okayama, A.; Nagatomo, Y.; Tachibana, N.; Tsubouchi, H. Genotypic identification of Rickettsia tsutsugamushi by restriction fragment length polymorphism analysis of DNA amplified by the polymerase chain reaction. Am. J. Trop. Med. Hyg. 1996, 54, 647–651. [Google Scholar] [CrossRef] [PubMed]
- Kawamori, F.; Akiyama, M.; Sugieda, M.; Kanda, T.; Akahane, S.; Yamamoto, S.; Ohashi, N.; Tamura, A. Two-step polymerase chain reaction for diagnosis of scrub typhus and identification of antigenic variants of Rickettsia tsutsugamushi. J. Vet. Med. Sci. 1993, 55, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Qiang, Y.; Tamura, A.; Urakami, H.; Makisaka, Y.; Koyama, S.; Fukuhara, M.; Kadosaka, T. Phylogenetic characterization of Orientia tsutsugamushi isolated in Taiwan according to the sequence homologies of 56-kDa type-specific antigen genes. Microbiol. Immunol. 2003, 47, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Blacksell, S.D.; Luksameetanasan, R.; Kalambaheti, T.; Aukkanit, N.; Paris, D.H.; McGready, R.; Nosten, F.; Peacock, S.J.; Day, N.P.J. Genetic typing of the 56-kDa type-specific antigen gene of contemporary Orientia tsutsugamushi isolates causing human scrub typhus at two sites in north-eastern and western Thailand. FEMS Immunol. Med. Microbiol. 2008, 52, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Enatsu, T.; Urakami, H.; Tamura, A. Phylogenetic analysis of Orientia tsutsugamushi strains based on the sequence homologies of 56-kDa type-specific antigen genes. FEMS Microbiol. Lett. 1999, 180, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.-Y.; Tsai, K.-H.; Yu, S.-K.; Cheng, C.-H.; Yang, J.-S.; Su, C.-L.; Hu, H.-C.; Wang, H.-C.; Huang, J.-H.; Shu, P.-Y. Phylogenetic analysis of 56-kDa type-specific antigen gene of Orientia tsutsugamushi isolates in Taiwan. Am. J. Trop. Med. Hyg. 2010, 83, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.E.; Siritantikorn, S.; Rolain, J.M.; Suputtamongkol, Y.; Hoontrakul, S.; Charoenwat, S.; Losuwanaluk, K.; Parola, P.; Raoult, D. Detection of new genotypes of Orientia tsutsugamushi infecting humans in Thailand. Clin. Microbiol. Infect. 2008, 14, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Tay, S.T.; Rohani, Y.M.; Ho, T.M.; Shamala, D. Sequence analysis of the hypervariable regions of the 56 kDa immunodominant protein genes of Orientia tsutsugamushi strains in Malaysia. Microbiol. Immunol. 2005, 49, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.W.; Choi, Y.K.; Baek, Y.H.; Seong, M.H. Phylogenetic analysis of the 56-kDa type-specific protein genes of Orientia tsutsugamushi in central Korea. J. Korean Med. Sci. 2012, 27, 1315–1319. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Kim, D.M.; Lee, S.H.; Jang, M.S.; Neupane, G.P. Phylogenetic analysis of the 56 kDa protein genes of Orientia tsutsugamushi in southwest area of Korea. Am. J. Trop. Med. Hyg. 2011, 84, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.-R.; Tsai, H.-P.; Tsui, P.-Y.; Weng, M.-H.; Kuo, M.-D.; Lin, H.-C.; Chen, K.-C.; Ji, D.-D.; Chu, D.-M.; Liu, W.-T. Genetic typing of the 56-kDa type-specific antigen gene of Orientia tsutsugamushi strains isolated from chiggers from wild-caught rodents in Taiwan. Appl. Environ. Microbiol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-W.; Lee, C.K.; Kwak, Y.G.; Moon, C.; Kim, B.-N.; Kim, E.S.; Kang, J.M.; Lee, C.-S. Antigenic drift of Orientia tsutsugamushi in South Korea as identified by the sequence analysis of a 56-kDa protein-encoding gene. Am. J. Trop. Med. Hyg. 2010, 83, 930–935. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Blacksell, S.D.; Phetsouvanh, R.; Phongmany, S.; Rolain, J.M.; Day, N.P.; Newton, P.N.; Raoult, D. Genotyping of Orientia tsutsugamushi from humans with scrub typhus, Laos. Emerg. Infect. Dis. 2008, 14, 1483–1485. [Google Scholar] [CrossRef] [PubMed]
- Varghese, G.M.; Janardhanan, J.; Mahajan, S.K.; Tariang, D.; Trowbridge, P.; Prakash, J.A.; David, T.; Sathendra, S.; Abraham, O. Molecular epidemiology and genetic diversity of Orientia tsutsugamushi from patients with scrub typhus in 3 regions of India. Emerg. Infect. Dis. 2015, 21, 64. [Google Scholar] [CrossRef] [PubMed]
- Ruang-areerate, T.; Jeamwattanalert, P.; Rodkvamtook, W.; Richards, A.L.; Sunyakumthorn, P.; Gaywee, J. Genotype diversity and distribution of Orientia tsutsugamushi causing scrub typhus in Thailand. J. Clin. Microbiol. 2011, 49, 2584–2589. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Cho, N.H.; Kim, S.Y.; Bang, S.Y.; Chu, H.; Choi, M.S.; Kim, I.S. Fibronectin facilitates the invasion of Orientia tsutsugamushi into host cells through interaction with a 56-kDa type-specific antigen. J. Infect. Dis. 2008, 198, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.; Kurokawa, K.; Fukuhara, M.; Urakami, H.; Yamamoto, S.; Yamazaki, K.; Ogura, Y.; Ooka, T.; Hayashi, T. Genome comparison and phylogenetic analysis of Orientia tsutsugamushi strains. DNA Res. 2010, 17, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Duong, V.; Blassdell, K.; May, T.T.; Sreyrath, L.; Gavotte, L.; Morand, S.; Frutos, R.; Buchy, P. Diversity of Orientia tsutsugamushi clinical isolates in Cambodia reveals active selection and recombination process. Infect. Genet. Evol. 2013, 15, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Sonthayanon, P.; Peacock, S.J.; Chierakul, W.; Wuthiekanun, V.; Blacksell, S.D.; Holden, M.T.; Bentley, S.D.; Feil, E.J.; Day, N.P. High rates of homologous recombination in the mite endosymbiont and opportunistic human pathogen Orientia tsutsugamushi. PLoS Negl Trop Dis 2010, 4, e752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phetsouvanh, R.; Sonthayanon, P.; Pukrittayakamee, S.; Paris, D.H.; Newton, P.N.; Feil, E.J.; Day, N.P. The diversity and geographical structure of Orientia tsutsugamushi strains from scrub typhus patients in Laos. PLoS Negl. Trop. Dis. 2015, 9, e0004024. [Google Scholar] [CrossRef] [PubMed]
- Wongprompitak, P.; Duong, V.; Anukool, W.; Sreyrath, L.; Mai, T.T.X.; Gavotte, L.; Moulia, C.; Cornillot, E.; Ekpo, P.; Suputtamongkol, Y. Orientia tsutsugamushi, agent of scrub typhus, displays a single metapopulation with maintenance of ancestral haplotypes throughout continental South East Asia. Infect. Genet. Evol. 2015, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bozeman, F.M.; Elisberg, B.L. Serological diagnosis of scrub typhus by indirect immunofluorescence. Proc. Soc. Exp. Biol. Med. 1963, 112, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Iida, T.; Kawashima, H.; Kawamura, A. Direct immunofluorescence for typing of tsutsugamushi disease rickettsia. J. Immunol. 1965, 95, 1129–1133. [Google Scholar] [PubMed]
- Blacksell, S.D.; Bryant, N.J.; Paris, D.H.; Doust, J.A.; Sakoda, Y.; Day, N.P. Scrub typhus serologic testing with the indirect immunofluorescence method as a diagnostic gold standard: A lack of consensus leads to a lot of confusion. Clin. Infect. Dis. 2007, 44, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Coleman, R.E.; Sangkasuwan, V.; Suwanabun, N.; Eamsila, C.; Mungviriya, S.; Devine, P.; Richards, A.L.; Rowland, D.; Ching, W.-M.; Sattabongkot, J.; et al. Comparative evaluation of selected diagnostic assays for the detection of IgG and IgM antibody to Orientia tsutsugamushi in Thailand. Am. J. Trop. Med. Hyg. 2002, 67, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Philip, R.; Casper, E.; Ormsbee, R.; Peacock, M.; Burgdorfer, W. Microimmunofluorescence test for the serological study of Rocky Mountain spotted fever and typhus. J. Clin. Microbiol. 1976, 3, 51–61. [Google Scholar] [PubMed]
- Gupta, N.; Chaudhry, R.; Thakur, C.K. Determination of cutoff of ELISA and immunofluorescence assay for scrub typhus. J. Glob. Infect. Dis. 2016, 8, 97. [Google Scholar] [PubMed]
- Horton, K.C.; Jiang, J.; Maina, A.; Dueger, E.; Zayed, A.; Ahmed, A.A.; Pimentel, G.; Richards, A.L. Evidence of rickettsia and orientia infections among abattoir workers in Djibouti. Am. J. Trop. Med. Hyg. 2016, 95, 462–465. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Minamishima, Y. Serodiagnosis of tsutsugamushi fever (scrub typhus) by the indirect immunoperoxidase technique. J. Clin. Microbiol. 1982, 15, 1128–1132. [Google Scholar] [PubMed]
- Suwanabun, N.; Chouriyagune, C.; Eamsila, C.; Watcharapichat, P.; Dasch, G.A.; Howard, R.S.; Kelly, D.J. Evaluation of an enzyme-linked immunosorbent assay in Thai scrub typhus patients. Am. J. Trop. Med. Hyg. 1997, 56, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Dasch, G.A.; Halle, S.; Bourgeois, A.L. Sensitive microplate enzyme-linked immunosorbent assay for detection of antibodies against the scrub typhus rickettsia, Rickettsia tsutsugamushi. J. Clin. Microbiol. 1979, 9, 38–48. [Google Scholar] [PubMed]
- Land, M.V.; Ching, W.M.; Dasch, G.A.; Zhang, Z.; Kelly, D.J.; Graves, S.R.; Devine, P.L. Evaluation of a commercially available recombinant-protein enzyme-linked immunosorbent assay for detection of antibodies produced in scrub typhus rickettsial infections. J. Clin. Microbiol. 2000, 38, 2701–2705. [Google Scholar] [PubMed]
- Chen, H.W.; Zhang, Z.; Huber, E.; Mutumanje, E.; Chao, C.C.; Ching, W.M. Kinetics and magnitude of antibody responses against the conserved 47-kilodalton antigen and the variable 56-kilodalton antigen in scrub typhus patients. Clin. Vaccine Immunol. 2011, 18, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Ha, N.Y.; Kim, Y.; Choi, J.H.; Choi, M.S.; Kim, I.S.; Kim, Y.S.; Cho, N.H. Detection of antibodies against Orientia tsutsugamushi Sca proteins in scrub typhus patients and genetic variation of SCA genes of different strains. Clin. Vaccine Immunol. 2012, 19, 1442–1451. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Yeo, S.J.; Park, S.J.; Woo, Y.J.; Kim, M.W.; Kim, S.H.; Chang, I.A.; Jeon, S.H.; Park, B.J.; Song, G.J.; et al. Improvement of the diagnostic sensitivity of scrub typhus using a mixture of recombinant antigens derived from Orientia tsutsugamushi serotypes. J. Korean Med. Sci. 2013, 28, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Vallee, J.; Thaojaikong, T.; Moore, C.E.; Phetsouvanh, R.; Richards, A.L.; Souris, M.; Fournet, F.; Salem, G.; Gonzalez, J.P.; Newton, P.N. Contrasting spatial distribution and risk factors for past infection with scrub typhus and murine typhus in Vientiane City, Lao PDR. PLoS Negl. Trop. Dis. 2010, 4, e909. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Myers, T.E.; Rozmajzl, P.J.; Graf, P.C.; Chretien, J.P.; Gaydos, J.C.; Richards, A.L. Seroconversions to rickettsiae in US military personnel in South Korea. Emerg. Infect. Dis. 2015, 21, 1073–1074. [Google Scholar] [CrossRef] [PubMed]
- Blacksell, S.D.; Jenjaroen, K.; Phetsouvanh, R.; Wuthiekanun, V.; Day, N.P.; Newton, P.N.; Ching, W.-M. Accuracy of AccessBio immunoglobulin M and total antibody rapid immunochromatographic assays for the diagnosis of acute scrub typhus infection. Clin. Vaccine Immunol. 2010, 17, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Anitharaj, V.; Stephen, S.; Pradeep, J.; Park, S.; Kim, S.H.; Kim, Y.J.; Kim, E.Y.; Kim, Y.W. Serological diagnosis of acute scrub typhus in southern India: Evaluation of InBios Scrub Typhus Detect IgM rapid test and comparison with other serological tests. J. Clin. Diagn. Res. 2016, 10, DC07–DC10. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.C.; Zhangm, Z.; Weissenberger, G.; Chen, H.W.; Ching, W.M. Lateral flow rapid test for accurate and early diagnosis of scrub typhus: A febrile illness of historically military importance in the Pacific Rim. Mil. Med. 2017, 182, 369–375. [Google Scholar] [CrossRef] [PubMed]
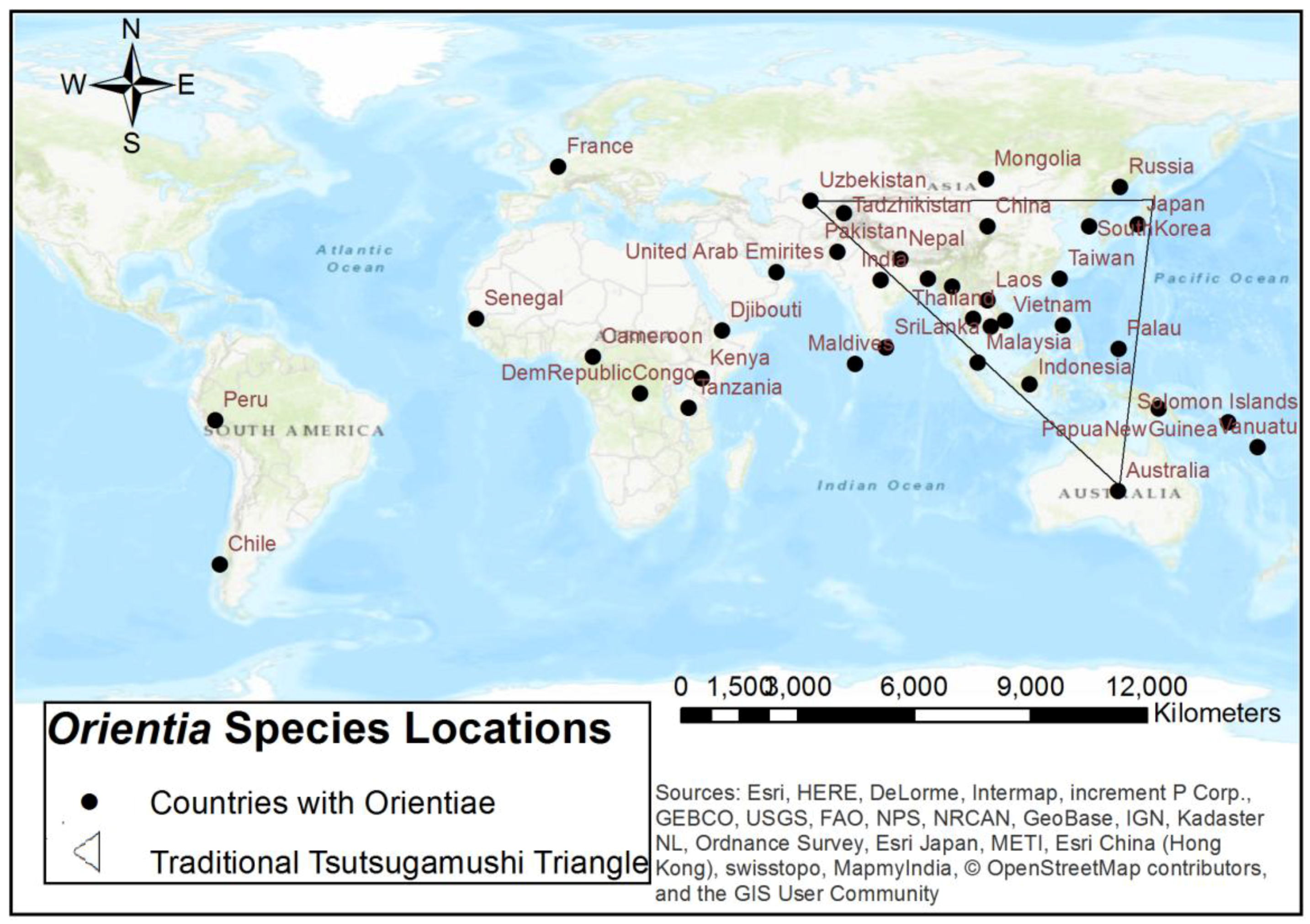

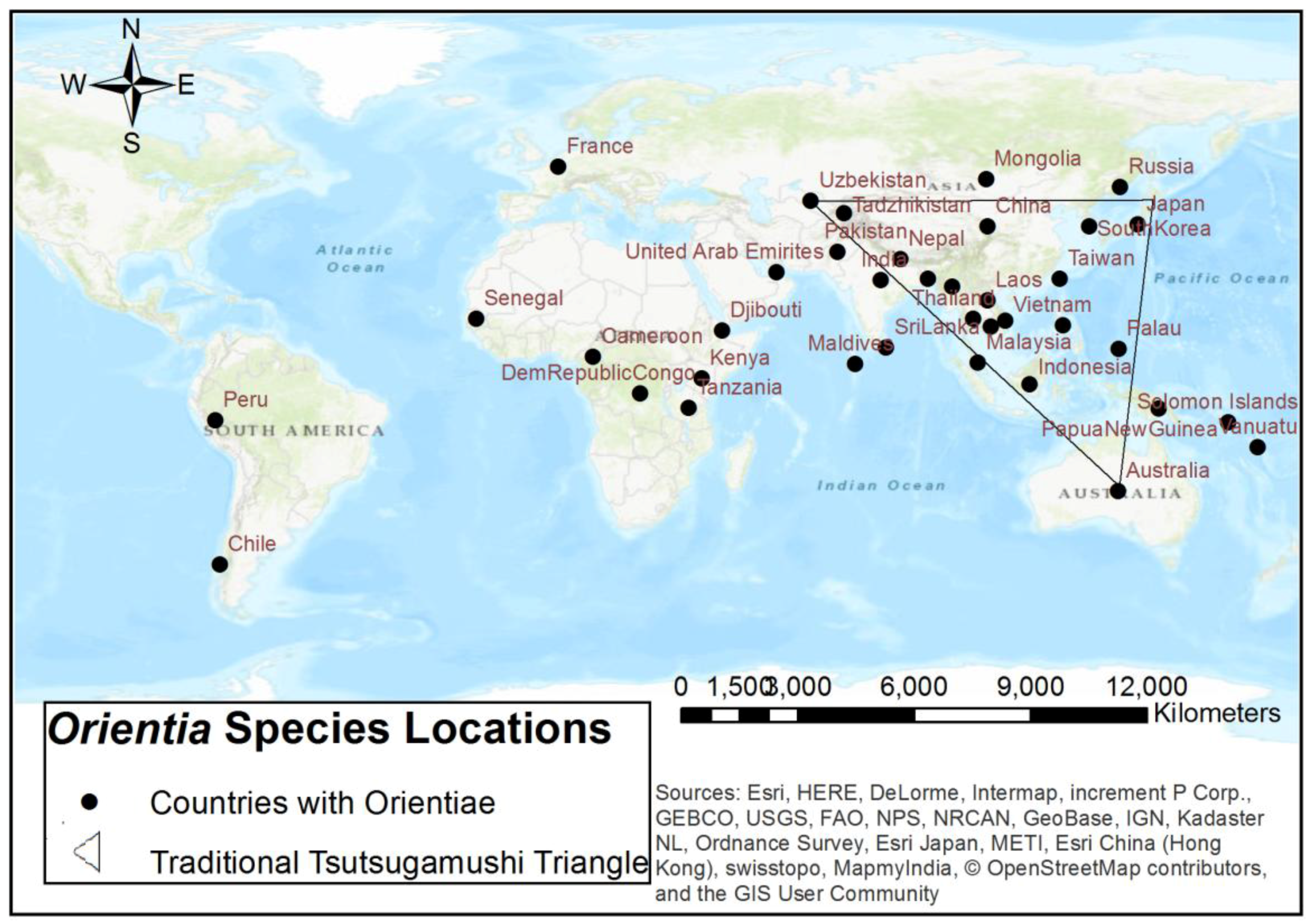{kind=link}
| Country | Province/State | Year | # Laboratory Confirmed or Suspected Cases | Fatalities | Outbreak Duration | Age in Years | Treatment | Laboratory Diagnosis | Other Differentials Ruled Out | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Australia | Northern Queensland | 2011 | 124 | 0 | 21 April–5 May 2011 | >18 | Doxycycline | IFA, PCR, isolation | Not done | Harris et al., 2016 |
| China | Jiangsu | 2013 | 271 | 0 | Hu et al., 2015 | |||||
| China | Guangdong | 2012 | 29 | 4 | 1 May–17 June 2012 | 24–86 | Not provided | WF, PCR, | Weil-Felix, PCR | Wei et al., 2014 # |
| Bhutan | Thimphu | 2014 | 12 | 2 | August–October 2014 | 6–15 | Not provided | RDT, IFA | Malaria, dengue, S. enteritica | Tshokey et al., 2017 |
| India | Northern India | 2013–2014 | 228 | 0 | July 2013–31 December 2014 | >12 | Doxycycline, azithromycin, ceftriaxone | ELISA IgM, PCR & sequencing | Malaria, Widal, leptospirosis | Sharma et al., 2016 |
| India | Himachal Pradesh (northern India) | 2008 | 5 | 1 | Not provided | 15–Mar | Doxycycline, azithromycin $ ceftriaxone ^ | WF, MIF | Rickettsial only | Mahajan et al., 2008 # |
| India | Pondicherry | 2008 | 50 | 1 | April 2006–April 2008 | 14–91 | Doxycycline | WF, eschar | Widal, dengue, malaria leptospirosis | Vivekanandan et al., 2010 |
| India | Meghalaya | 2010 | 24 | 0 | October 2009–January 2010 | Pediatric | Doxycycline, Chloramphenicol ^ Azithromycin $ | WF | Malaria | Dass et al., 2011 # |
| India | Meghalaya | 2012 | 90 | 5 | September 2011–August 2012 | Adults | Doxycycline, azithromycin $ | Rapid immunochromato-graphic test | Typhoid fever, malaria, leptospirosis, dengue | Sivarajan et al., 2016 |
| India | Puducherry | 2013 | 28 | 0 | September 2012–March 2013 | 1–89 | Doxycycline, azithromycin $ | Rapid immunochromato-graphic test, ELISA, WF paired sera | Typhoid, dengue, leptospirosis, malaria | Stephen et al., 2015 |
| India | Sikkim | 2011 | 63 | 0 | January–December 2011 | >2 | Doxycycline, azithromycin $ | Rapid immunochromato-graphic test, WF, ELISA, | Widal, Malaria, urine culture | Gurung et al., 2013 |
| India | Rajasthan | 2012 | 42 | 7 | October–December 2012 | 3–78 | Doxycycline | ELISA IgM | Widal, malaria, dengue | Sinha et al., 2014 |
| India | Chennai | 2011 | 52 | 0 | September 2010–June 2011 | 0–16 | - | ELISA, IgM, eschar | Not provided | Krishna et al., 2015 |
| India | Rajasthan | 2014 | 66 | 14 | July–October 2014 | Not provided | Doxycyline * azithromycin | ELISA IgM | Malaria, leptospirosis, dengue, typhoid, viral pharyngitis | Takhar et al., 2017 |
| India | Uttarakhand | 2014 | 69 | 1 | July–November 2013 | 12–80 | Not provided | IgM ELISA | Typhoid fever, malaria, dengue | Singh et al., 2014 |
| India | Andhra-pradesh | 2014 | 176 | 8 | August 2011–December 2012 | >12 | Doxycycline | Weil-Felix, rapid immunochromato-graphic test | Other causes excluded | Subbalaxmi et al., 2014 # |
| India | Andhra-pradesh | 2011–2013 | 258 | 0 | September–December 2011–2013 | 2–89 | Tetracyclines | IgM ELISA, PCR | Typhoid, chikungunya, dengue | Usha et al., 2016 |
| India | Bishnipur-Manipur | 2007 | 38 | 2 | May–September 2007 | 0–≥45 | Doxycyline, azithromycin | WF | Not provided | Singh et al., 2010 |
| Thailand | Chiang Mai | 2007 | 65 | 0 | June 2006–May 2007 | 0–13 | Not provided | IFA, PCR isolation | Not provided | Rodkvamtook et al., 2013 |
| Nepal | Chaitwan District | 2016 | 264 | 8 | May–October 2016 | Not provided | Not provided | Not provided | Not available | http://www.myrepublica.com/news/7173/ |
| Nepal | Kathmandu | 2015 | 23 | 0 | August–October 2015 | Not provided | Doxycycline, azithromycin, ceftriaxone | ELISA IgM | Not available | Bastola and pant, 2016 |
| Solomon Islands | Western Province | 2014 | 9 | 0 | 1–30 May 2014 | >11 | Doxycycline, chloramphenicol $ | IFA/MIF | Malaria | Marks et al., 2016 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luce-Fedrow, A.; Lehman, M.L.; Kelly, D.J.; Mullins, K.; Maina, A.N.; Stewart, R.L.; Ge, H.; John, H.S.; Jiang, J.; Richards, A.L. A Review of Scrub Typhus (Orientia tsutsugamushi and Related Organisms): Then, Now, and Tomorrow. Trop. Med. Infect. Dis. 2018, 3, 8. https://doi.org/10.3390/tropicalmed3010008
Luce-Fedrow A, Lehman ML, Kelly DJ, Mullins K, Maina AN, Stewart RL, Ge H, John HS, Jiang J, Richards AL. A Review of Scrub Typhus (Orientia tsutsugamushi and Related Organisms): Then, Now, and Tomorrow. Tropical Medicine and Infectious Disease. 2018; 3(1):8. https://doi.org/10.3390/tropicalmed3010008
Chicago/Turabian StyleLuce-Fedrow, Alison, Marcie L. Lehman, Daryl J. Kelly, Kristin Mullins, Alice N. Maina, Richard L. Stewart, Hong Ge, Heidi St. John, Ju Jiang, and Allen L. Richards. 2018. "A Review of Scrub Typhus (Orientia tsutsugamushi and Related Organisms): Then, Now, and Tomorrow" Tropical Medicine and Infectious Disease 3, no. 1: 8. https://doi.org/10.3390/tropicalmed3010008