
Effect of Feed Protein:Lipid Ratio on Growth Parameters of African Catfish Clarias gariepinus after Fish Meal Substitution in the Diet with Bambaranut (Voandzeia subterranea) Meal and Soybean (Glycine max) Meal


Abstract
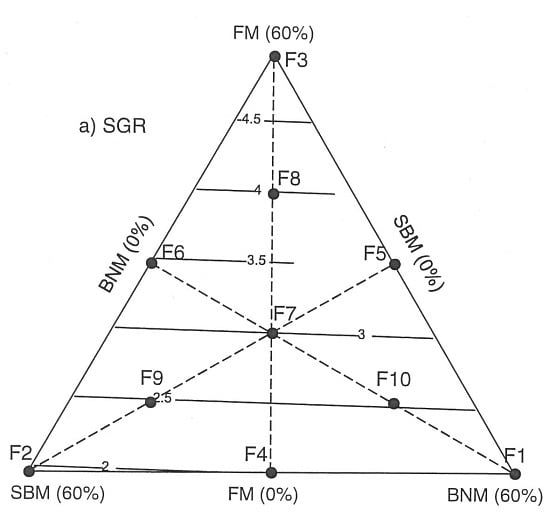
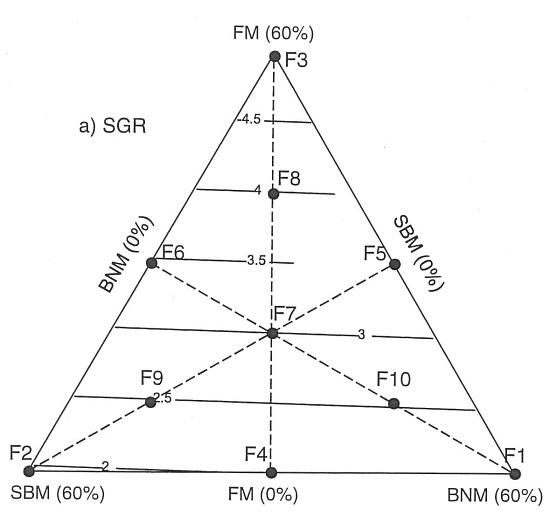
:
1. Introduction
2. Results
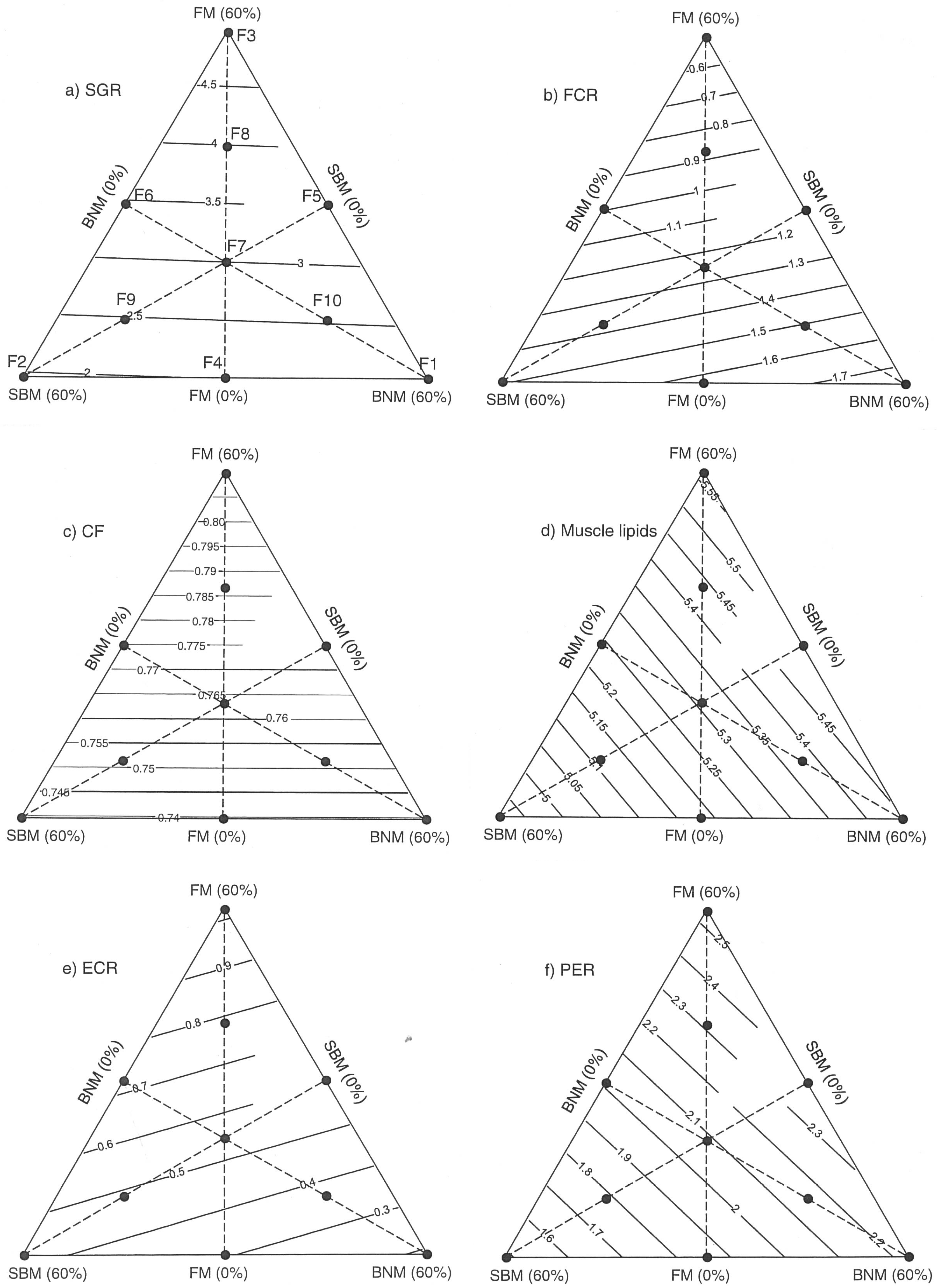
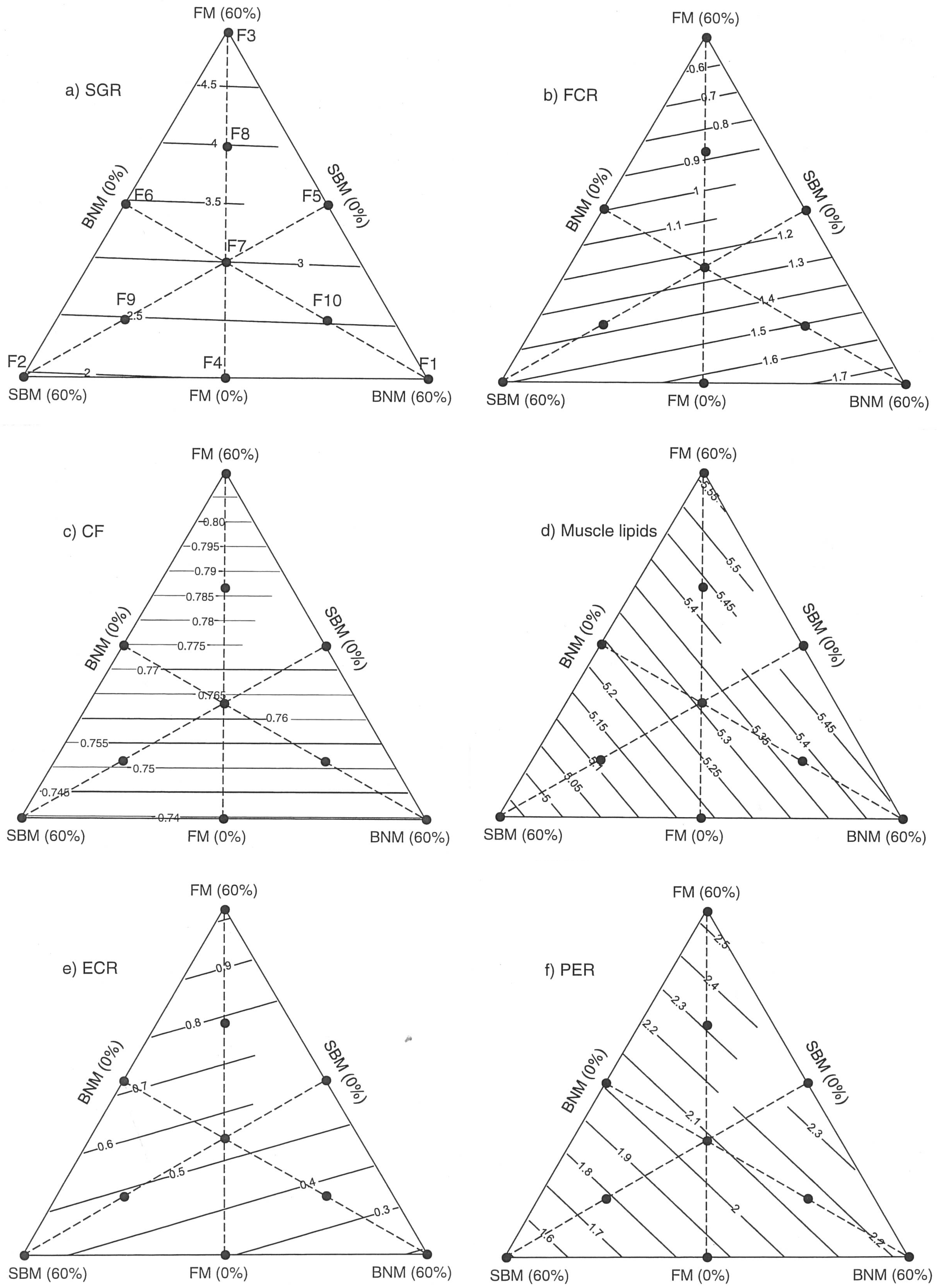
2.1. Effects of Protein:Lipid Ratios on Growth and Feed Utilization
2.2. Growth and Nutritional Performance Based on Response Surfaces
3. Discussion
4. Materials and Methods
4.1. Preparation of Feeds
4.2. Experimental Setup
4.3. Chemical Analyses
4.4. Calculations and Statistical Analyses
4.5. Mixture Design Principle
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Naylor, R.L.; Hardy, R.W.; Bureau, D.P.; Chiu, A.; Elliott, M.; Farrell, A.P.; Forster, I.; Gatlin, D.M.; Goldburg, R.J.; Hua, K.; et al. Feeding aquaculture in an era of finite resources. Proc. Natl. Acad. Sci. USA 2009, 106, 15103–15110. [Google Scholar] [CrossRef] [PubMed]
- Hardy, R.W. Utilization of plant protein in fish diets: Effects of global demand and supplies of fishmeal. Aquac. Res. 2010, 41, 770–776. [Google Scholar] [CrossRef]
- Drew, M.D.; Borgeson, T.L.; Thiessen, D.L. A review of processing of feed ingredients to enhance diet digestibility in finfish. Anim. Feed Sci. Technol. 2007, 138, 118–136. [Google Scholar] [CrossRef]
- Shipton, T.; Hecht, T. A Synthesis of the Formulated Animal and Aquafeed Industry in Sub-Saharan African. Available online: http://www.fao.org/docrep/008/a0042e/a0042e03.htm (accessed on 27 December 2016).
- Davies, O.A.; Ezenwa, N.C. Groundnut cake as alternative protein source in the diet of Clarias gariepinus fry. Int. J. Sci. Nat. 2010, 1, 73–76. [Google Scholar]
- Enyidi, U.D.; Mgbenka, B.O. Replacement of fish meal with bambaranut waste meal in the diets of larval African catfish Clarias gariepinus Burchell (1822). Br. J. Appl. Sci. Technol. 2015, 5, 526–537. [Google Scholar] [CrossRef]
- Imorou Toko, I.; Fiogbe, E.M.; Kestermont, P. Mineral status of African catfish (Clarias gariepinus) fed diets containing graded levels of soybean or cottonseed meals. Aquaculture 2008, 275, 298–305. [Google Scholar] [CrossRef]
- Azaza, M.S.; Wassim, K.; Mensi, F.; Abdelmouleh, A.; Brini, B.; Karaiem, M.M. Evaluation of faba beans (Vicia faba L. var. minuta) as a replacement for soybean meal in practical diets of juvenile Nile tilapia Oreochromis niloticus. Aquaculture 2009, 287, 174–179. [Google Scholar]
- Gatlin, D.M., III; Barrows, F.T.; Brown, P.; Dabrowski, K.; Gaylord, G.T.; Hardy, R.W.; Herman, E.; Hu, G.; Krogdahl, Å.; Nelson, R.; et al. Expanding the utilization of sustainable plant products in aquafeeds: A review. Aquac. Res. 2007, 38, 551–579. [Google Scholar] [CrossRef]
- Francis, G.; Makkar, P.S.H.; Becker, K. Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish. Aquaculture 2001, 199, 197–227. [Google Scholar] [CrossRef]
- Torstensen, B.E.; Espe, M.; Sanden, M.; Stubhaug, I.; Waagbo, R.; Hemre, G.-I.; Fontanillas, R.; Nordgarden, U.; Hevroy, E.M.; Olsvik, P.; et al. Novel production of Atlantic salmon (Salmo salar) protein based on combined replacement of fish meal and fish oil with plant meal and vegetable oil blends. Aquaculture 2008, 285, 193–200. [Google Scholar] [CrossRef]
- Karunaratne, A.; Azam-Ali, S.; Sasey, A.; Adu-dapaah, H.; Crout, N. Modelling the effect of temperature, soil moisture and photoperiod on growth and development of bambara groundnut (Vigna subterranean (L.) Verdc.): BAMGRO model. Global issues. Paddock action. In Proceedings of the 14th Australian Agronomy Conference, Adelaide, Australia, September 2008; Unkovich, M.J., Ed.; Australian Society of Agronomy: Adelaide, Australia, 2008; p. 4. [Google Scholar]
- Enyidi, U.D. Production of feeds for African catfish Clarias gariepinus using plant proteins. Jyvask. Stud. Biol. Sci. 2012, 251. [Google Scholar]
- Dakora, F.D.; Muofhe, L.M. Nitrogen fixation and nitrogen nutrition in symbiotic bambara groundnut (Vigna subterranea (L.) Verdc.) and Kersting’s bean (Macrotyloma geocarpum (Harms) Marech. et Baud.). In Proceedings of the Workshop on Conservation and Improvement of Bambara Groundnut (Vigna subterranea (L.) Verdc.), Harare, Zimbabwe, 14–16 November 1995; Hekler, J., Begemann, F., Mushonga, J., Eds.; Biodiversity International: Harare, Zimbabwe, 1995; pp. 72–78. [Google Scholar]
- Basu, S.; Roberts, J.A.; Azam-Ali, S.N.; Mayes, S. Bambara groundnut. In Pulses, Sugar and Tuber Crops. Genome Mapping and Molecular Breeding in Plants; Kole, C., Ed.; Springer: Berlin, Germany, 2007; pp. 159–171. [Google Scholar]
- Vielma, J.; Koskela, J.; Ruohonen, K.; Jokininen, I.; Kettunen, J. Optimal diet composition for European whitefish (Coregonous lavaretus): Carbohydrate stress and immune parameters responses. Aquaculture 2003, 225, 3–16. [Google Scholar] [CrossRef]
- Ruohonen, K.; Koskela, J.; Vielma, J.; Kettunen, J. Optimal diet composition for European whitefish (Coregonus lavaretus): Analysis of growth and nutrient utilization in mixture model trials. Aquaculture 2003, 225, 27–39. [Google Scholar] [CrossRef]
- Ruohonen, K.; Simpson, S.J.; Raubeheimer, D. A new approach to diet optimization: A re-analysis using European whitefish (Coregonus lavaretus). Aquaculture 2007, 267, 147–156. [Google Scholar] [CrossRef]
- Kim, L.O.; Lee, S. Effects of the dietary protein and lipids levels on growth and body composition of bagrid catfish, Pseudobagrus fulvidraco. Aquaculture 2005, 243, 323–329. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Fish and Shrimp; National Research Council of the National Academies: Washington, DC, USA, 2011; p. 363. [Google Scholar]
- Kim, K.-D.; Lim, S.G.; Kang, Y.J.; Kim, K.-W.; Son, M.H. Effects of dietary protein and lipid levels on growth and body composition of juvenile far eastern catfish Silurus asotus. Asian-Australas. J. Anim. Sci. 2012, 25, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Santinha, P.J.M.; Medale, F.; Corraze, G.; Gomes, E.F.S. Effects of the dietary protein:lipid ratio on growth and nutrient utilization in gilthead seabream (Sparus aurata L.). Aquac. Nutr. 1995, 5, 147–156. [Google Scholar] [CrossRef]
- Forster, I.P.; Dominy, W.G.; Lawrence, A.L.; Castille, F.L.; Patnaik, S. Optimization of a research diet for the Pacific white shrimp, Litopenaeus vannamei, using mixture model methodology. Aquaculture 2010, 298, 260–266. [Google Scholar] [CrossRef]
- Minjarez-Osorio, C.; Castillo-Alvarado, S.; Gatlin III, D.M.; González-Félix, M.L.; Perez-Velazquez, M.; Rossi, W., Jr. Plant protein sources in the diets of the sciaenids red drum (Sciaenops ocellatus) and shortfin corvina (Cynoscion parvipinnis): A comparative study. Aquaculture 2016, 453, 122–129. [Google Scholar] [CrossRef]
- Ruohonen, K.; Kettunen, J. Effective experimental designs for optimizing fish feeds. Aquac. Nutr. 2004, 10, 145–151. [Google Scholar] [CrossRef]
- Khan, M.; Abidi, S.F. Optimum histidine requirement of fry African catfish Clarias gariepinus (Burchell). Aquac. Res. 2010, 40, 1000–1010. [Google Scholar] [CrossRef]
- Fagbenro, O.A.; Davies, S.J. Use of soybean flour (dehulled, solvent-extracted soybean) as a fish meal substitute in practical diets for African catfish, Clarias gariepinus (Burchell 1822). J. Appl. Ichthyol. 2001, 17, 64–69. [Google Scholar] [CrossRef]
- Hu, M.; Wang, Y.; Wang, Q.; Zhao, M.; Xiong, B.; Qian, X.; Zhao, Y.; Luo, Z. Replacement of fish meal by rendered animal protein ingredients with lysine and methionine supplementation to practical diets for gibel carp, Carassius auratus gibelio. Aquaculture 2008, 275, 260–265. [Google Scholar] [CrossRef]
- Cheng, Z.J.; Hardy, W.R.; Usry, J.L. Effects of lysine supplementation in plant protein-based diets on the performance of rainbow trout (Oncorhynchus mykiss) and apparent digestibility coefficients of nutrients. Aquaculture 2003, 215, 255–265. [Google Scholar] [CrossRef]
- Yang, H.J.; Liu, Y.J.; Tian, X.L.; Liang, Y.G.; Hao, R.L. Effects of supplemental lysine and methionine on growth performance and body composition for grass carp (Ctenopharyngodon idella). Am. J. Agric. Biol. Sci. 2010, 5, 222–227. [Google Scholar] [CrossRef]
- Ng, W.-K.; Lim, P.-K.; Boey, P.-L. Dietary and palm oil source affects growth, fatty acid composition and muscle α-tocopherol concentration of African catfish, (Clarias gariepinus). Aquaculture 2003, 215, 229–243. [Google Scholar] [CrossRef]
- Manning, B.B.; Li, M.H.; Robinson, E.D.; Peterson, B.C. Enrichment of channel catfish (Ictalurus punctatus) fillets with conjugated linoleic acid and ω-3 fatty acids by dietary manipulation. Aquaculture 2006, 261, 337–342. [Google Scholar] [CrossRef]
- Storebakken, T.; Refstie, S.; Ruyter, B. Soy products as fat and protein sources in fish feeds for intensive aquaculture. In Soy in Animal Nutrition; Drackley, J.K., Ed.; Federation of Animal Science Societies: Champaign, IN, USA, 2000; pp. 127–170. [Google Scholar]
- Boyd, C.E.; Tucker, C.; Mc Nevin, A.; Bostick, K.; Clay, J. Indicators of resource use efficiency and environmental performance in fish and crustacean aquaculture. Rev. Fish. Sci. 2007, 15, 327–360. [Google Scholar] [CrossRef]
- Parrish, C.C. Determination of total lipid classes and fatty acids in aquatic samples. In Lipids in Freshwater Ecosystems; Wetzel, R.G., Art, M.T., Wainmann, B.C., Eds.; Springer: New York, NY, USA, 1999; pp. 4–20. [Google Scholar]
- Directive, C. Establishing community methods for the determination of amino acids, crude oils and fats, and olaquindox in feeding stuffs and amending directive 71/393/EEC, annex part a. Determination of amino acids. Off. J. Eur. Commun. L 1998, 257, 14–23. [Google Scholar]
- Onyimonyi, A.E.; Ugwu, S.O.C. Performance of laying hens fed varying dietary levels of bambara (Voandzeia subterrenea Thouars) offals. Int. J. Poult. Sci. 2007, 6, 223–226. [Google Scholar] [CrossRef]
- Cornell, J. Experiments with Mixtures: Designs, Models, and the Analysis of Mixture Data; John Wiley & Sons, Inc.: New York, NY, USA, 2002. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 20 January 2017).

{kind=link}
{kind=link}
| Diet | Initial wt. (g) | Final wt. (g) | SGR | FCR | PPV | FFE | PER |
|---|---|---|---|---|---|---|---|
| Feed 1 | 33.9 ± 0.00 | 49.2 ± 5.20 f | 1.83 ± 0.14 d | 1.65 ± 0.47 c | 16.8 ± 0.00 e | 0 | 1.64 ± 00 b |
| Feed 2 | 35.8 ± 1.30 | 55.2 ± 5.9 f | 1.94 ± 0.64 d | 1.49 ± 0.23 c | 15.3 ± 0.01 e | 0 | 1.16 ± 0.59 b |
| Feed 3 | 35.1 ± 3.50 | 122.2 ± 2.5 a | 4.92 ± 1.01 a | 0.67 ± 0.06 a | 32.9 ± 0.08 b | 1.99 ± 0.18 d | 2.46 ± 0.22 a |
| Feed 4 | 35.5 ± 1.40 | 63.6 ± 1.30 e | 2.08 ± 0.80 c | 1.69 ± 0.27 c | 19.2 ± 0.03 d | 0 | 2.72 ± 0.09 a |
| Feed 5 | 35.6 ± 2.83 | 111.2 ± 5.30 b | 3.77 ± 0.80 b | 0.98 ± 0.04 b | 37.8 ± 0.03 a | 1.30 ± 2.03 c | 2.34 ± 0.05 a |
| Feed 6 | 34.6 ± 1.90 | 109.3 ± 1.1 b | 3.74 ± 0.57 b | 0.84 ± 0.04 b | 31.1 ± 0.01 b | 1.12 ± 0.06 b | 2.30 ± 0.13 a |
| Feed 7 | 34.9 ± 6.40 | 85.8 ± 5.90 c | 3.22 ± 0.35 b | 1.11 ± 0.11 e | 28.7 ± 0.01 b | 1.00 ± 0.02 b | 2.07 ± 0.03 a,b |
| Feed 8 | 35.4 ± 4.24 | 106.9 ± 6.70 b | 3.94 ± 0.82 b | 0.82 ± 0.06 b | 30.0 ± 0.01 b | 1.46 ± 0.04 c | 2.37 ± 0.06 a |
| Feed 9 | 34.7 ± 3.50 | 58.1 ± 5.90 e,f | 1.84 ± 0.63 d | 1.62 ± 0.60 c | 15.1 ± 0.02 e | 0.72 ± 0.02 a | 1.41 ± 0.09 b |
| Feed 10 | 36.2 ± 1.20 | 76.4 ± 2.80 d | 2.67 ± 0.29 c | 1.31± 0.1 d | 25.7 ± 0.03 c | 0.58 ± 0.01 a | 2.06 ± 0.03 a,b |
| Amino Acid | F1 | F2 | F3 | F4 | F5 | F6 | F7 | F8 | F9 | F10 |
|---|---|---|---|---|---|---|---|---|---|---|
| Arginine | 11.49 | 23.33 | 28.55 | 17.41 | 20.02 | 25.94 | 21.13 | 24.84 | 22.23 | 16.31 |
| Histidine | 4.63 | 7.93 | 10.51 | 6.28 | 7.57 | 9.22 | 7.69 | 9.10 | 7.81 | 6.16 |
| Isoleucine | 7.47 | 13.88 | 19.82 | 10.28 | 13.26 | 16.86 | 13.47 | 16.65 | 13.68 | 10.08 |
| Leucine | 12.17 | 23.60 | 32.96 | 17.88 | 22.57 | 28.28 | 22.91 | 27.94 | 23.26 | 17.54 |
| Lysine | 9.07 | 19.21 | 34.15 | 14.14 | 21.62 | 26.69 | 20.82 | 27.49 | 20.02 | 14.95 |
| Methionine | 2.30 | 4.51 | 12.91 | 3.40 | 7.60 | 8.71 | 6.57 | 9.74 | 5.54 | 4.43 |
| Cysteine | 1.77 | 5.05 | 4.99 | 3.41 | 3.38 | 5.02 | 3.93 | 4.46 | 4.49 | 2.85 |
| Phenylalanine | 8.61 | 15.93 | 17.55 | 12.27 | 7.17 | 16.74 | 14.03 | 15.79 | 14.98 | 11.32 |
| Tyrosine | 7.17 | 11.44 | 14.08 | 9.30 | 10.63 | 12.76 | 10.90 | 12.49 | 11.17 | 9.03 |
| Threonine | 5.12 | 12.10 | 18.15 | 8.61 | 11.64 | 15.13 | 11.79 | 14.98 | 11.95 | 8.45 |
| Valine | 7.55 | 16.51 | 27.00 | 12.03 | 17.28 | 21.76 | 17.02 | 22.02 | 16.77 | 12.90 |
| Ingredient | F1 | F2 | F3 | F4 | F5 | F6 | F7 | F8 | F9 | F10 |
|---|---|---|---|---|---|---|---|---|---|---|
| BNM | 600 | 0 | 0 | 300 | 300 | 0 | 200 | 100 | 100 | 400 |
| SBM | 0 | 600 | 0 | 300 | 0 | 300 | 200 | 100 | 400 | 100 |
| FM | 0 | 0 | 600 | 0 | 300 | 300 | 200 | 400 | 100 | 100 |
| Bloodmeal | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Wheat | 205 | 205 | 205 | 205 | 205 | 205 | 205 | 205 | 205 | 205 |
| Premix a | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 |
| Fish oil | 70 | 70 | 70 | 70 | 70 | 70 | 70 | 70 | 70 | 70 |
| Total | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 |
| Proximate composition (%) | ||||||||||
| Protein | 27.3 | 47.0 | 55.4 | 39.7 | 43.9 | 52.4 | 43.9 | 52.1 | 46.8 | 37.5 |
| Fat | 18.4 | 13.8 | 21.9 | 13.7 | 18.1 | 17.2 | 17.8 | 18.2 | 15.4 | 13.3 |
| Moisture | 5.5 | 6.5 | 6.7 | 6.3 | 6.9 | 6.1 | 6.1 | 6.3 | 6.4 | 6.2 |
| Protein:Lipid | 1.5:1 | 3.4:1 | 2.5:1 | 2.9:1 | 2.4:1 | 3.0:1 | 2.4:1 | 2.8:1 | 2.9:1 | 2.8:1 |
| Ash | 0.2 | 0.7 | 5.9 | 3.0 | 4.2 | 2.2 | 3.7 | 5.4 | 1.5 | 0.7 |
| Amino Acid | FM | BNM | SBM |
|---|---|---|---|
| Arginine | 45.4 | 17.0 | 36.7 |
| Histidine | 16.5 | 6.7 | 12.2 |
| Isoleucine | 31.3 | 9.4 | 21.4 |
| Leucine | 51.9 | 17.2 | 36.3 |
| Lysine | 55.7 | 13.9 | 30.8 |
| Methionine | 20.8 | 3.1 | 6.8 |
| Cysteine | 7.4 | 2.0 | 7.5 |
| Phenylalanine | 27.1 | 12.2 | 24.4 |
| Tyrosine | 22.0 | 10.5 | 17.6 |
| Threonine | 29.0 | 7.3 | 18.9 |
| Valine | 43.0 | 10.6 | 25.5 |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enyidi, U.D.; Pirhonen, J.; Kettunen, J.; Vielma, J. Effect of Feed Protein:Lipid Ratio on Growth Parameters of African Catfish Clarias gariepinus after Fish Meal Substitution in the Diet with Bambaranut (Voandzeia subterranea) Meal and Soybean (Glycine max) Meal. Fishes 2017, 2, 1. https://doi.org/10.3390/fishes2010001
Enyidi UD, Pirhonen J, Kettunen J, Vielma J. Effect of Feed Protein:Lipid Ratio on Growth Parameters of African Catfish Clarias gariepinus after Fish Meal Substitution in the Diet with Bambaranut (Voandzeia subterranea) Meal and Soybean (Glycine max) Meal. Fishes. 2017; 2(1):1. https://doi.org/10.3390/fishes2010001
Chicago/Turabian StyleEnyidi, Uchechukwu D., Juhani Pirhonen, Juhani Kettunen, and Jouni Vielma. 2017. "Effect of Feed Protein:Lipid Ratio on Growth Parameters of African Catfish Clarias gariepinus after Fish Meal Substitution in the Diet with Bambaranut (Voandzeia subterranea) Meal and Soybean (Glycine max) Meal" Fishes 2, no. 1: 1. https://doi.org/10.3390/fishes2010001