Effects of Elevated Temperature and Potassium on Biomass and Quality of Dark Red ‘Lollo Rosso’ Lettuce



1
Department of Plant and Soil Sciences, Mississippi State University, North Mississippi Research and Extension Center, Verona, MS 38879, USA
2
Department of Plant Sciences, The University of Tennessee, Knoxville, TN 37996, USA
*
Author to whom correspondence should be addressed.
Horticulturae 2018, 4(2), 11; https://doi.org/10.3390/horticulturae4020011
Submission received: 2 May 2018
/
Revised: 12 June 2018
/
Accepted: 14 June 2018
/
Published: 16 June 2018
(This article belongs to the Special Issue Plant Production in Controlled Environment)
Abstract
:Lettuce is an economically important crop for small and medium-sized growers. When grown in adverse environmental conditions, lettuce is vulnerable to a deterioration of yield and quality. Research concerning the impact of elevated potassium (K) levels on leafy vegetables, such as lettuce, is lacking. Therefore, seeds of dark-red ‘Lollo’ lettuce were germinated under greenhouse conditions at 25/20 °C (day/night). Plants were transferred into 11-L containers and placed into growth chambers at 25 and 33 °C. Plants were grown with K treatments of 117.3 (control), 234.6 (2×), 469.2 (4×), and 4) 938.4 (8×) mg·L−1. Increasing K treatments resulted in a negative quadratic response on lettuce dry mass and generated 14% more leaf calcium at 234.6 mg·L−1. An increase in temperature from 25 to 33 °C increased leaf dry matter and biomass by 40% and 43%, respectively. Leaf water content increased by 3% as temperature increased. Plants grown at 33 °C had greater quercetin glycosides compared to plants grown at 25 °C. The results from this study suggest that temperature is a stronger regulatory factor than increasing K in the determination of lettuce yield and quality. Increasing K concentration to 234.6 mg·L−1 results in greater concentrations of leaf minerals without compromising plant yield.
1. Introduction
Lettuce is a nutritious plant that contains vitamins such as A, K, C, and folates [1]. Due to its comparatively large consumption in raw form, lettuce is also a notable source of minerals such as iron (Fe), calcium (Ca), and magnesium (Mg) as well as antioxidants. Red-leaf lettuce cultivars are especially high in phytonutrients such as phenolics and anthocyanins, which have been linked to the reduced incidence of chronic and degenerative diseases [2]. According to the 2012 United States census of agriculture, lettuce (head, leaf, and romaine combined) ranks first in total production value of fresh market vegetables. In the southern United States, open field and greenhouse lettuce production occur most commonly in the fall and winter seasons. This allows growers to take advantage of cooler temperatures and shorter days, and to maximize economic returns due to fewer overhead requirements and increased product quality. However, the progressing growth of the population and affluent communities coincides with public demand for year-round access to locally sourced, high-quality lettuce.
When grown in adverse environmental conditions, lettuce yield and quality can be severely diminished. For example, subjecting lettuce to high air and root-zone temperatures accelerates the inflorescence initiation (bolting) and results in reduced marketability [3,4]. This issue is especially exaggerated during the summer in the southern United States due to consistent exposure to high temperatures and long days, which can increase bolting and lettuce bitterness. Moreover, Jenni reported that crisphead lettuce exposed to heat stress for a 3 or 5-day period, two weeks after heading resulted in 46% of mature lettuce heads with rib discoloration [5]. Additionally, impacts on yield and quality are heavily associated with fertilization. For example, previous research has reported that increasing nitrogen (N) from 0 to 60 mg·L−1 resulted in greater shoot mass in lettuce grown at deep water culture, and lettuce supplied with 400 mg·L−1 N and supplemental light resulted in accelerated growth and shoot dry matter [6,7]. However, Fu et al. [8] found that leafy lettuce grown under high light (220 mol·m−2·s−1) and low N (7 mmol·L−1) resulted in increases in shoot biomass and vitamin C. Crisphead lettuce grown with a shortage of Ca resulted in chlorosis at the leaf margins and loss of apical dominance. Additionally, limiting boron (B) caused leaves to become wrinkled, stunted plant growth, and resulted in the loss of apical dominance [9]. Conversely, Barickman et al. [10] reported that romaine lettuce grown with increasing potassium (K) concentrations showed improvement in biomass, leaf sucrose, and leaf K content. However, increasing K resulted in decreases in some leaf minerals such as Ca, Mg, and sulfur (S) [10,11].
While K is not a known component of any functional plant molecules or structures, it is essential for growth and required in relatively large quantities. Potassium plays a vital role in protein synthesis, activation of enzymes, and photosynthesis [12,13]. Additionally, it is an essential factor in the translocation of photo-assimilates from source to sink tissues, stomatal conductance, and the maintenance of cell turgor pressure [10,13,14]. Potassium also plays a critical role in the survival of plants under stressful environmental conditions by protecting against light-induced cell damage, drought, and enhanced resistance to damage from low temperatures [15] These abiotic stresses commonly result in the increased production of reactive oxygen species (ROS); however, potassium lowers the production of ROS by maintaining the electron transport chain and reducing NAD(P)H oxidase activity [15].
Studies of the influence of K concentration on yield and quality concerning leaf lettuce are lacking, although research on crisphead lettuce and several other crops have examined its effect. For example, crisphead lettuce and Chinese cabbage grown in increasing levels of potassium nitrate (KNO3) (1, 5, or 10 mM) showed maximum yield at 5 mM, decreased leaf sodium (Na) and chloride (Cl) content, and increased leaf N and K content in both saline and non-saline conditions [16]. Moreover, Tzortzakis [17] reported that Ca and K enrichment alleviated salinity-induced stress in hydroponically grown endives. Promising supplemental K work has been conducted in muskmelon (Cucumis melo). For example, late-season foliar applications of two different K formulations improved fruit marketability attributes such as yield, firmness, and sugar content [12]. Additionally, increases in human-health related bioactive compounds, ascorbic acid and β-carotene, were observed [12]. However, research on adequate and elevated levels of K as it relates to lettuce yield and quality is inconclusive. For example, the fresh and dry shoot weight of ‘South Bay’ lettuce plantlets were unaffected by increasing K levels from 15 to 60 mg L−1 [6]. Bres and Weston [18] concluded that increasing K or pH levels did not consistently affect tipburn incidence in three lettuce cultivars. Conversely, Fallovo [19] found that seasonal changes in combination with increasing fertilizer concentrations resulted in plant growth and yield increases in hydroponic leafy lettuce. The current evidence on the effect of K treatments on lettuce yield and quality has produced some promising results. However, further research is needed to understand this dynamic relationship better. Thus, the purpose of this study was to determine the effect of temperature and adequate and elevated K levels on growth chamber produced red-leaf lettuce biomass accumulation, mineral nutrient uptake, and phenolic content.
2. Materials and Methods
2.1. Plant Culture and Harvest
Seeds of Dark Red ‘Lollo’ Rosso lettuce (Johnny’s Selected Seed, Waterville, ME, USA) were sown into 2.5 × 2.5-cm growing cubes (Grodan, Hedehuse, Denmark), and germinated in greenhouse conditions (Verona, MS; 34° N, 89° W) at 25/20 °C (day/night). After seedlings developed their first true leaf, three plantlets were transplanted into a 11-L Rubbermaid© (Rubbermaid, Atlanta, GA, USA) Roughneck plastic storage container. The dimensions of the container were 40.1 cm × 26.2 cm × 17.8 cm (L × W × H). Plants were transplanted into holes that were drilled into the lids of the containers. The tubs were filled with 10-L−1 of nutrient solution using a modified half-strength Hoagland formulation [20]. Elemental concentrations of half-strength nutrient solution consisted of (mg·L−1): N (105), P (91.5), Ca (80.2), Mg (24.6), S (32.0), Fe (1.0), B (0.25), Mo (0.005), Cu (0.01), Mn (0.25), and Zn (0.025). The nutrient solutions were aerated via tubing connected to an air blower (Model: 4-LPM, Uniclife Co., Guangzhou, China). Containers were then placed inside one of two growth chambers (Percival Scientific, Perry, IA), and plants were subjected to a 12/12 h (day/night) photoperiod and maintained at 65% relative humidity. Each chamber was divided into two shelves and equipped with 2-incandescent, white light bulbs and 14-fluorescent bulbs per shelf, generating a photosynthetic flux of 360 μmol·m–2·s–1 at plant canopy level. The experimental design for this study was a randomized complete block, with eight treatments and four replications in a 2 × 4 factorial arrangement. Treatments consisted of two temperature treatments of 25 and 33 °C and four K treatments of (1) 117.3 mg·L−1, (2) 234.6 mg·L−1, (3) 469.2 mg·L−1, and (4) 938.4 mg·L−1. Containers were monitored and supplemented with reverse osmosis (RO) water as needed. Plants were grown for 30 days before harvesting the three plants per treatment and then separated into roots and shoots before being weighed for total root and shoot biomass. A 10 to 15 g subsample was retrieved from each treatment and replication, freeze-dried (FreeZone® 2.5 L, Model Number: 7670520, Labconco Corp., Kansas City, MO, USA), and reweighed to determine plant biomass. Lettuce subsamples were then analyzed for nutritional quality.
2.2. Flavonoid Analysis
Flavonoid analysis was conducted according to Chen [21]. Briefly, lettuce leaf samples were ground using a mortar and pestle for a homogenous subsample. A 0.04 g subsample was extracted in a 2 mL microcentrifuge tube by adding 1.0 mL of extraction solvent (60:37:3) consisting of methanol, de-ionized water, and formic acid. The samples were then vortexed for 1 min and centrifuged at 20,000 gn for 15 min. After centrifugation, the samples were filtered through a 0.45 μm syringe filter and collected in a 2 mL HPLC vial for analysis. Separation parameters and flavonoid quantification were carried out with authentic standards using an Agilent 1260 series HPLC with a multiple wavelength detector (Agilent Technologies, Willington, DE, USA). Chromatographic separations were achieved using a 150 × 4.6 mm i.d., 2.6 μm analytical scale Kinetex F5 reverse-phase column (Phenomenex, Torrance, CA, USA), which allows for effective separation of chemically similar flavonoid compounds. The column was equipped with a Kinetex F5 12.5 × 4.6 mm i.d. guard cartridge and holder (Phenomenex), and it was maintained at 30 °C using a thermostat column compartment. All separations were achieved using a mobile gradient phase of reverse osmosis (RO) water adjusted to pH 2.5 with trifluoroacetic acid and acetonitrile. Anthocyanin analysis was similar to the flavonoid determination procedure with slight modification. Briefly, 0.04 g of red lettuce subsamples were extracted in a 2 mL microcentrifuge tube by adding 1.0 mL of extraction solvent (50:40:10) consisting of water, methanol, and acetic acid. The samples were then vortexed for 1 min and centrifuged at 20,000 gn rpm for 15 min. After centrifugation, the samples were filtered through a 0.45 μm syringe filter and collected in a 2 mL HPLC vial for analysis.
2.3. Mineral Composition
Nutrient analysis was conducted according to Barickman et al. [22] with slight modifications. Briefly, a 5.0-g subsample of fresh leaf tissue, which was combined with 10-mL of 70% HNO3, was digested in a microwave digestion unit (Model: Ethos, Milestone Inc., Shelton, CT, USA). Leaves were collected and dried for 48-h in a forced air oven (model large; Fisher Scientific, Atlanta, GA, USA) at 65 °C. Dried samples were ground to homogeneity using liquid nitrogen, and 0.5-g subsamples were weighed for analysis. Nutrient analysis was conducted using an inductively coupled plasma mass spectrometer (ICP-MS; Agilent Technologies, Inc., Wilmington, DE, USA). The ICP-MS system was equipped with an octopole collision/reaction cell, Agilent 7500 ICP-MS ChemStation software, a Micromist nebulizer, a water-cooled quartz spray chamber, and a CETAC (ASX-510, CETAC Inc., Omaha, NE, USA) auto-sampler. The instrument was optimized daily in terms of sensitivity (lithium: Li, yttrium: Y, thallium: Tl), level of oxide, and doubly charged ion using tuning solution containing 10 μg∙L-1 of Li, Y, Tl, cerium (Ce), and cobalt (Co) in a 2% HNO3/0.5% HCl (v/v) matrix. Tissue nutrient concentrations are expressed on a dry weight (DW) basis.
2.4. Statistical Analysis
Data were subjected to the GLIMMIXED procedure and mean separation using Tukey’s Honest Significant Difference test (P ≤ 0.05) with SAS statistical software (Version 9.4; SAS Institute, Cary, NC, USA).
3. Results
3.1. Temperature and Treatment Effect on Plant Growth
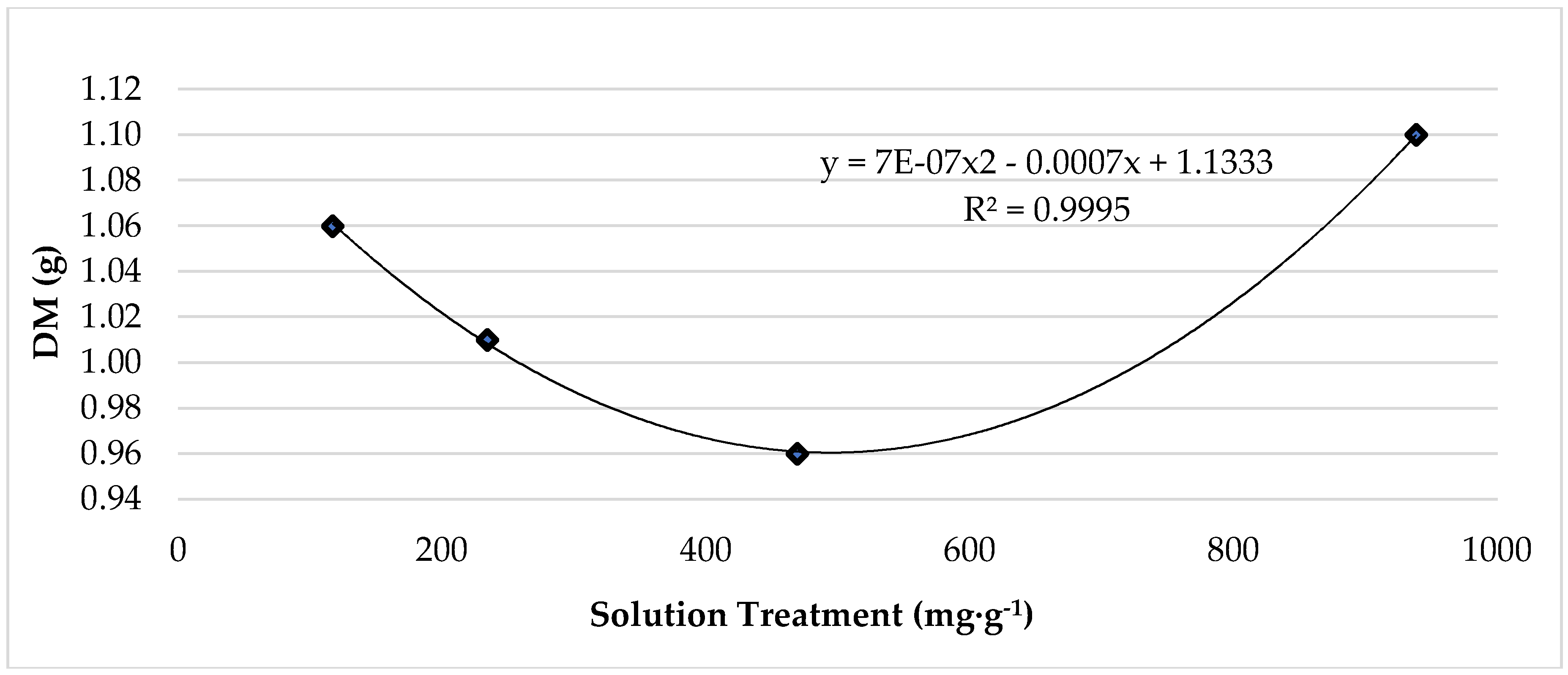
While leaf fresh mass (FM) was not impacted, lettuce leaf dry mass (DM), biomass (FM:DM), and water content were significantly affected (Table 1). Leaf DM increased by 40% from 25 °C to 33 °C, and leaf biomass increased from 0.07-g to 0.10-g (43%). Conversely, lettuce subjected to 25 °C produced the highest leaf water content, which was 3% larger compared to lettuce grown at 33 °C. Pertaining to lettuce roots, temperature treatments had a significant effect on root fresh mass and dry mass but did not significantly impact biomass or water content (Table 1). Fresh mass and dry mass were significantly higher among plants grown at 25 °C. Averaging 10.11-g, root fresh mass was 214% greater than the fresh mass of roots grown at 33 °C, which averaged 3.22-g. Similarly, root dry mass was 186% greater with respect to plants grown at 25 °C. Nutrient solution treatment did not significantly affect growth and production parameters. The highest level of K, 938.4 mg·L−1, resulted in the most dry mass (Figure 1).
3.2. The Effect of Temperature and Treatment on Leaf Phenolics and Mineral Content
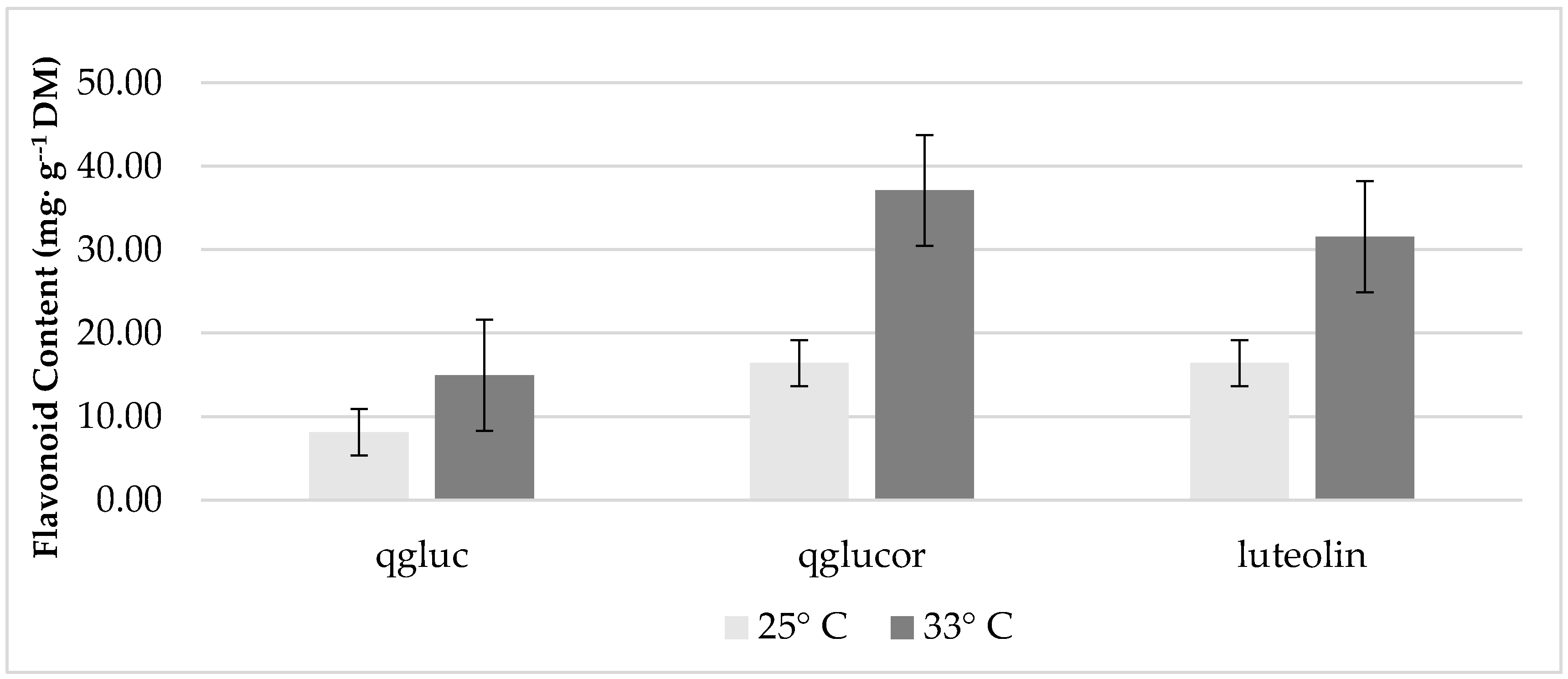
Increasing K concentrations did not significantly affect concentrations of lettuce phenolic compounds. However, plants grown at 33 °C exhibited a greater accumulation of quercetin glycosides and luteolin compared to plants grown at 25 °C (Figure 2).
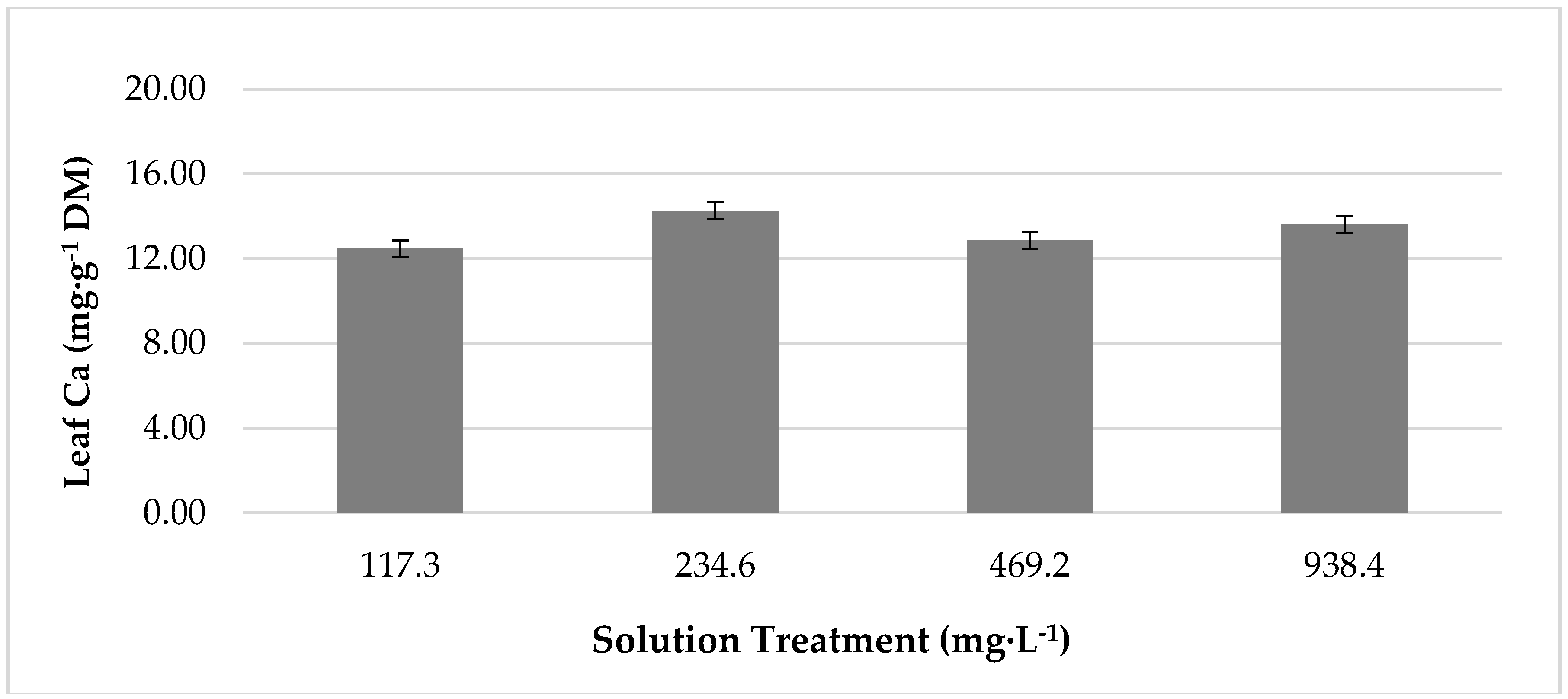
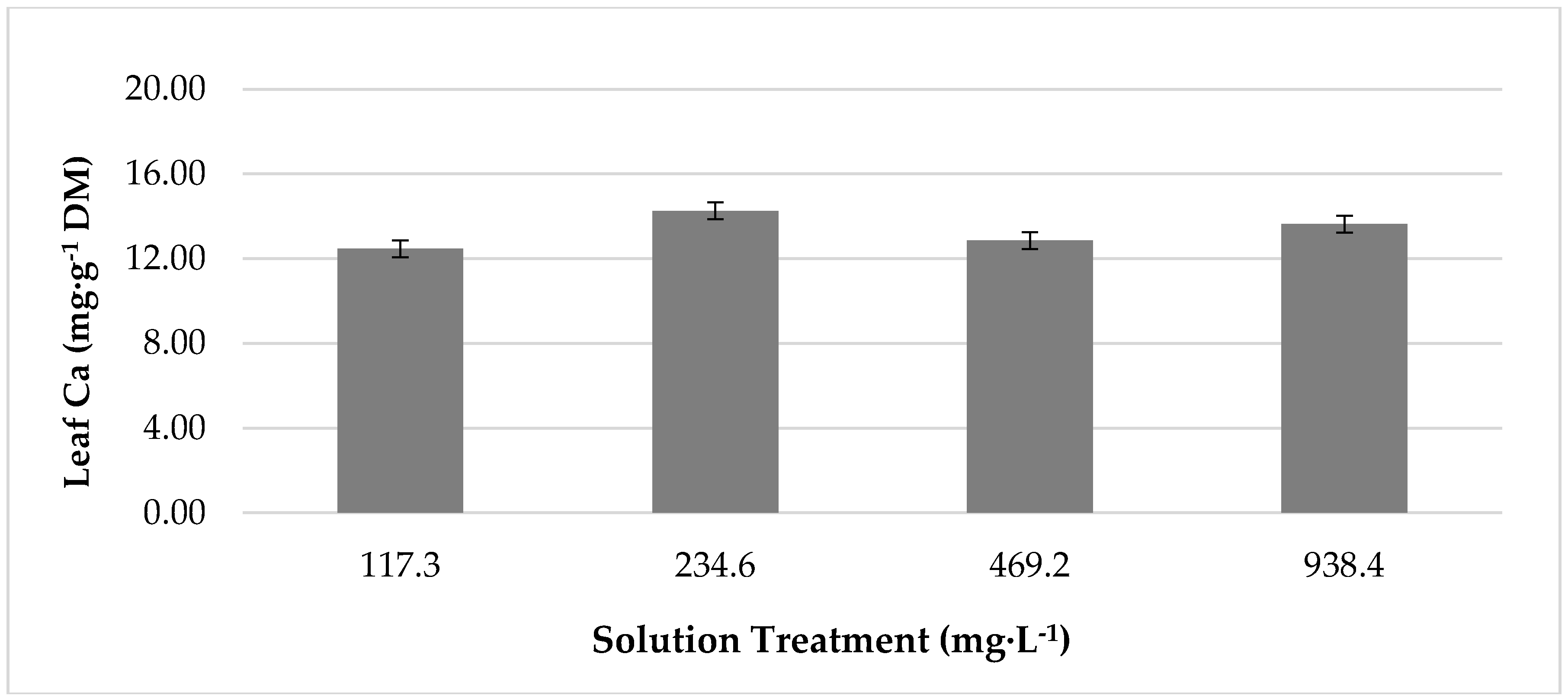
Growth chamber temperature had a significant effect on leaf concentrations of macronutrients Mg, K, and Ca (Table 2). Concentrations of all macronutrients were greatest in plants grown at 25 °C. At 33 °C, Mg concentration decreased by 19%, K decreased by 11%, and Ca declined by 13% from values at 25 °C. Additionally, the temperature had a significant impact on concentrations of micronutrients manganese (Mn) and molybdenum (Mo) (Table 2). Concentrations of both nutrients were significantly greater in plants grown at 25 °C. At 33 °C, Mn concentration was reduced by 42%, and Mo content declined by 35% from values at 25 °C. Nutrient solution treatment had no effect onleaf nutrient concentrations for any minerals except for Ca (Figure 3), which showed the greatest concentration in plants produced with 234.6 mg·L−1 of K. This was 14% greater than the concentration produced by lettuce under 117.3 mg·L−1 of K. However, lettuce produced with 938.4 mg·L−1 of K was statistically comparable. Moreover, lettuce grown with 938.4 mg·L−1 was comparable to lettuce grown with 117.3 mg·L−1 and 469.2 mg·L−1 of K.
3.3. The Effect of Temperature and Treatment Interactions on Leaf Mineral Content
Interactions between temperature and K solution treatment significantly affected nutrient concentrations of phosphorous (P), sulfur (S), and copper (Cu) only (Table 3). Plants grown at 25 °C in 117.3 mg·L−1 of K produced maximum levels of P (8.50 mg·g−1), followed respectively by 469.2 mg·L−1 (7.69 mg·g−1), 938.4 mg·L−1 (7.61 mg·g−1), and 234.6 mg·L−1 (7.40 mg·g−1), which were comparable to each other. Plants grown at 33 °C produced the lowest P concentrations, and treatments did not significantly differ. Plants grown at 25 °C accumulated the highest concentrations of sulfur, but treatments were not significantly different from each other. Plants grown at 33 °C in 117.3 mg·L−1 produced the greatest concentration of sulfur (9.99 mg·g−1), resulting in comparable amounts to plants grown at 25 °C; whereas, plants grown at 33 °C in 234.6, 469.2, and 938.4 mg·L−1 produced comparable concentrations to each other. Additionally, copper concentrations were greatest in plants grown with 469.2 mg·L−1 at 25 °C, which decreased 14% with plants grown with 938.4 mg·L−1 of K and an additional 20% and 2% in plants grown with 234.6 and 117.3 mg·L−1, respectively. Plants grown at 33 °C with 117.3 mg·L−1 accumulated the highest concentrations of copper (3.22 μg·g−1 DW) which declined 58%, 43%, and 53% with K treatments of 234.6, 469.2, and 938.4 mg·L−1, respectively.
4. Discussion
The current study examined how increasing K concentration in a deep-water culture production system affected the growth and quality of ‘Dark Red’ ‘Lollo’ Rosso lettuce under mild and extreme temperatures. Lettuce is a cool season crop with a preferred temperature range of 7–24 °C. Inflorescence initiation is encouraged by exposure to high temperatures and photoperiods of ≥13 h [23]. Producing lettuce above these thresholds affects plasma membrane fluidity, microtubules and microfilaments, and the photosynthetic apparatus [24]. This causes an intracellular signal cascade that ultimately reduces plant yield and quality. Previous studies have examined the effect of elevated temperatures on yield, quality, and growth rate for many vegetables. For example, plants grown at supra-optimal temperatures resulted in smaller tomato fruit, reduced flower and fruit set in bell peppers, and decreased net photosynthesis and chlorophyll content of Chinese cabbage [25,26,27]. Adverse temperatures also altered the form and color of cucumber and eggplant fruit and increased the relative growth rate of lettuce plants by 33% to 85% [28,29].
The results of the current study diverged from the common effects of heat stress on lettuce and other crops. Differences in production temperatures resulted in an inverse relationship between root and shoot development. Plants grown at 33 °C demonstrated greater production of shoot fresh mass, dry mass, and biomass, while plants grown at 25 °C had greater leaf water content, root fresh mass, and root dry mass. The degree to which lettuce can tolerate adverse temperatures is known to vary greatly among cultivars. In at least one variety trial comparing 21 cultivars of red-leaf lettuce, ‘Lollo Rosso’ was among the top four cultivars that exhibited the greatest resistance to heat stress [30]. Thus, plants produced at the highest temperature in the current study may have withstood deleterious effects on fresh mass, dry mass, and biomass due to heat tolerance inherent in the ‘Lollo Rosso’ cultivar. Moreover, the lack of exogenous stress on plants produced in milder temperatures may have allowed for the greater generation of root fresh and dry mass.
With respect to the production of secondary metabolites, increasing K concentration did not affect lettuce flavonoid and phenolic acid content. The highest concentrations of phenolics were notably present in lettuce plants produced at the highest temperature treatment, which is consistent with the literature demonstrating increased production of phenolics under warm temperatures in lettuce, tomato, and watermelon [31,32,33]. Instead, elevating the temperature to 33 °C resulted in significant differences of flavonoid content in lettuce plants. Unlike amino acids, sugars, and fatty acids, flavonoid and phenolic acid compounds are not required by plants for survival [33]. However, production of secondary metabolites is an adaptive mechanism by plants to aid in protection from environmental stressors. For example, high light intensity often results in the production of secondary metabolites called anthocyanins in flowering plants. Anthocyanins offer plants additional photoprotection due to the ability to absorb radiation in the photosynthetically active spectrum [34,35,36].
Previous research has examined the role of flavonoids and phenolic acids in mitigating abiotic stress such as extreme heat. For example, Oh et al. [37] reported that concentrations of chicoric and chlorogenic acid, quercetin-3-O-glucoside, and luteolin-7-O-glucoside increased significantly in five-week-old lettuce plants subjected to heat shock treatments. Additionally, flavonoid content increased in three pigmented baby lettuce cultivars as growing season progressed from February to May [38]. Conversely, Boo et al. [39] stated that total polyphenol concentrations decreased with increasing heat treatments. However, another study concluded that lettuce cultivar was a larger contributing factor to the accumulation of phenolic compounds among five tested varieties [40]. The results of the current study suggest that elevating temperatures were responsible for the increases in flavonoid and phenolic content, since light intensity and photoperiod were equal and held constant in both temperature treatments.
Increasing concentrations of K did not significantly affect the uptake of leaf mineral nutrients except for calcium, which increased among plants treated with 2× (234.6 mg·L−1) and 8× (938.4 mg·L−1) levels of K. Although not statistically significant, a visible trend was observed among plants exposed to 2× treatments of K, containing the greatest concentrations Mg, K, Fe, Mn, and Mo. Although K is not a constituent of any functional plant molecules or structures, it is vital to plant growth and development and is known to affect the uptake of mineral nutrients such as Ca and Mg [13]. Several studies have explored the impact of increasing nutrient solution concentrations on leaf mineral nutrient content. For example, Fallovo et al. [19] investigated the effects of increasing nutrient solution electrical conductivity (EC) on yield and quality of leafy lettuce. Results suggested that concentrations of N, P, K, and Mg increased concerning increasing nutrient solution EC. Similar results were confirmed by Chiloane [41], who found that N and P had a tendency to increase with increasing solution EC, and K increased with increasing solution EC before declining with the highest treatments.
Furthermore, Barickman et al. [10] demonstrated that elevating K in red-romaine lettuce grown in a soilless production medium increased leaf concentration of K by 43% but resulted in decreases in Ca, Mg, and S concentrations. Additionally, results from this study indicated that leaf mineral nutrient content increased before reaching a saturation point and declined with the greatest K treatments [10]. The results of the current study are consistent with the findings of previous studies. In comparison to control treatments, several nutrients increased with 2× (234.6 mg·L−1) treatments before content declined at 4× (469.2 mg·L−1) and 8× (938.4 mg·L−1). Additionally, concentrations of the macronutrients Mg, K, and Ca and micronutrients Mn and Mo were significantly greater at the lowest temperature treatment, suggesting that lack of exogenous temperature stress supported root productivity and facilitated the uptake and assimilation of minerals better than plants experiencing abiotic stress.
In conclusion, continuing research is needed to develop a better understanding of the dynamic relationship between abiotic stress, such as adverse temperatures and elevated EC, and the deterministic role these factors play in lettuce growth, development, and nutritional quality. While previous research has demonstrated the efficacy of increasing K on positive growth and qualitative characteristics, results vary among crop species and cultivars. In this situation, the application of increasing levels of K produced comparable levels of leaf DM at 117.3 mg·L−1 and 938.4 mg·L−1 and resulted in 14% more leaf calcium content at 234.6 mg·L−1, compared to the control treatment of 117.3 mg·L−1. However, previous research suggested that the beneficial effect of increasing K on some leaf mineral nutrients in lettuce may come with a reduction in others due to competition at the site of root uptake, generating inconsistency in nutritional quality. Ultimately, differences in growth chamber temperatures generated the most significant impact on lettuce biomass, phenolic content, and leaf mineral concentrations. Therefore, the results from this study suggest that temperature is a stronger regulatory factor than increasing K in the determination of lettuce yield and quality; however, increasing K concentration to 234.6 mg·L−1 resulted in higher concentrations of leaf calcium without compromising lettuce yield or quality.
Author Contributions
For this research article, T.C.B. conceived and designed the experiments; W.L.S. performed the experiments; T.C.B. and W.L.S. analyzed the data; T.C.B., W.L.S., and C.E.S. contributed sample analysis; W.L.S. wrote the manuscript; T.C.B. and C.E.S. edited the manuscript.
Funding
This publication is a contribution of the Mississippi Agriculture and Forestry Experiment Station and supported by the USDA NIFA Hatch S-294 Project MIS 146030.
Acknowledgments
The authors would like to thank the research team, Thomas Horgan, Cameron Tate, and Susan Worthy, for all their help with the project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Romani, A.; Pinelli, P.; Galardi, C.; Sani, G.; Cimato, A.; Heimler, D. Polyphenols in Greenhouse and Open-Air-Grown Lettuce. Food Chem. 2002, 79, 337–342. [Google Scholar] [CrossRef]
- Tsormpatsidis, E.; Henbest, R.G.C.; Davis, F.J.; Battey, N.H.; Hadley, P.; Wagstaffe, A. UV irradiance as a major influence on growth, development and secondary products of commercial importance in Lollo Rosso lettuce “Revolution” grown under polyethylene films. Environ. Exp. Bot. 2008, 63, 232–239. [Google Scholar] [CrossRef]
- Simonne, A.; Simonne, E.; Eitenmiller, R.; Coker, C.H. Bitterness and composition of lettuce varieties grown in the southeastern United States. Horttechnology 2002, 12, 721–726. [Google Scholar]
- Waycott, W. Photoperiodic Response of Genetically Diverse Lettuce Accessions. J. Am. Soc. Hortic. Sci. 1995, 120, 460–467. [Google Scholar]
- Jenni, S. Rib Discoloration: A Physiological Disorder Induced by Heat Stress in Crisphead Lettuce. HortScience 2005, 40, 2031–2035. [Google Scholar]
- Soundy, P.; Cantliffe, D.J.; Hochmuth, G.J.; Stoffella, P.J. Nutrient Requirements for Lettuce Transplants Using a Floatation Irrigation System II. Potassium. HortScience 2001, 36, 1071–1074. [Google Scholar]
- Masson, J.; Tremblay, N.; Gosselin, A. Nitrogen Fertilization and HPS Supplementary Lighting Influence Vegetable Transplant Production I. Transplant Growth. J. Am. Soc. Hortic. Sci. 1991, 116, 594–598. [Google Scholar]
- Fu, Y.; Li, H.; Yu, J.; Liu, H.; Cao, Z.; Manukovsky, N.S.; Liu, H. Interaction effects of light intensity and nitrogen concentration on growth, photosynthetic characteristics and quality of lettuce (Lactuca sativa L. Var. youmaicai). Sci. Hortic. (Amsterdam) 2017, 214, 51–57. [Google Scholar] [CrossRef]
- Petrazzini, L.L.; Souza, G.A.; Rodas, C.L.; Emrich, E.B.; Carvalho, J.G.; Souza, R.J. Nutritional deficiency in crisphead lettuce grown in hydroponics. Hortic. Bras. 2014, 32, 310–313. [Google Scholar] [CrossRef] [Green Version]
- Barickman, T.C.; Horgan, T.E.; Wheeler, J.R.; Sams, C.E. Elevated Levels of Potassium in Greenhouse-grown Red Romaine Lettuce Impacts Mineral Nutrient and Soluble Sugar Concentrations. HortScience 2016, 51, 504–509. [Google Scholar]
- Inthichack, P.; Nishimura, Y.; Fukumoto, Y. Effect of potassium sources and rates on plant growth, mineral absorption, and the incidence of tip burn in cabbage, celery, and lettuce. Hortic. Environ. Biotechnol. 2012, 53, 135–142. [Google Scholar] [CrossRef]
- Lester, G.E.; Jifon, J.L.; Makus, D.J. Impact of potassium nutrition on postharvest fruit quality: Melon (Cucumis melo L.) case study. Plant Soil 2010, 335, 117–131. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: San Diego, CA, USA, 2012. [Google Scholar]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Achilea, O. Alleviation of salinity—Induced stress in cash crops by multi-K (Potassium nitrate), five cases typifying the underlying pattern. Acta Hortic. 2002, 573, 43–48. [Google Scholar] [CrossRef]
- Tzortzakis, N.G. Potassium and calcium enrichment alleviate salinity-induced stress in hydroponically grown endives. Hortic. Sci. 2010, 37, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Bres, W.; Weston, L. A Nutrient Accumulation and Tipburn in {NFT-grown} Lettuce at Several Potassium and {pH} Levels. HortScience 1992, 27, 790–792. [Google Scholar]
- Fallovo, C.; Rouphael, Y.; Cardarelli, M.; Rea, E.; Battistelli, A.; Colla, G. Yield and quality of leafy lettuce in response to nutrient solution composition and growing season. J. Food Agric. Environ. 2009, 7, 456–462. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Ca. Agric. Exp. Stn. Circ. 1950, 347, 1–32. [Google Scholar]
- Chen, H.J.; Inbaraj, B.S.; Chen, B.H. Determination of phenolic acids and flavonoids in Taraxacum formosanum kitam by liquid chromatography-tandem mass spectrometry coupled with a post-column derivatization technique. Int. J. Mol. Sci. 2012, 13, 260–285. [Google Scholar] [CrossRef] [PubMed]
- Barickman, T.C.; Kopsell, D.A.; Sams, C.E. Effect of selenium and sulfur fertilization on glucosinolate and isothiocyanates in Arabidopsis thaliana and rapid-cycling Brassica oleracea. Acta Hortic. 2014, 1040, 31–38. [Google Scholar] [CrossRef]
- Jenni, S.; Truco, M.J.; Michelmore, R.W. Quantitative trait loci associated with tipburn, heat stress-induced physiological disorders, and maturity traits in crisphead lettuce. Theor. Appl. Genet. 2013, 126, 3065–3079. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, V.M.; Soengas, P.; Alonso-Villaverde, V.; Sotelo, T.; Cartea, M.E.; Velasco, P. Effect of temperature stress on the early vegetative development of Brassica oleracea L. BMC Plant Biol. 2015, 15, 145. [Google Scholar] [CrossRef] [PubMed]
- Adams, S.R. Effect of Temperature on the Growth and Development of Tomato Fruits. Ann. Bot. 2001, 88, 869–877. [Google Scholar] [CrossRef] [Green Version]
- Erickson, A.N.; Markhart, A.H. Flower Production, Fruit Set, and Physiology of Bell Pepper during Elevated Temperature and Vapor Pressure Deficit. J. Am. Soc. Hortic. Sci. 2001, 126, 697–702. [Google Scholar]
- Yuan, L.; Yuan, Y.; Liu, S.; Wang, J.; Zhu, S.; Chen, G.; Hou, J.; Wang, C. Influence of high temperature on photosynthesis, antioxidative capacity of chloroplast, and carbon assimilation among heat-tolerant and heat-susceptible genotypes of nonheading Chinese cabbage. HortScience 2017, 52, 1464–1470. [Google Scholar] [CrossRef]
- Gruda, N. Impact of environmental factors on product quality of greenhouse vegetables for fresh consumption. CRC Crit. Rev. Plant Sci. 2005, 24, 227–247. [Google Scholar] [CrossRef]
- Gent, M.P.N. Effect of temperature on composition of hydroponic lettuce. Acta Hortic. 2016, 1123, 95–100. [Google Scholar] [CrossRef]
- Lafta, A.; Turini, T.; Sandoya, G.V.; Mou, B. Field Evaluation of Green and Red Leaf Lettuce Genotypes in the Imperial, San Joaquin, and Salinas Valleys of California for Heat Tolerance and Extension of the Growing Seasons. HortScience 2017, 52, 40–48. [Google Scholar] [CrossRef]
- Leng, P.; Itamura, H.; Yamamura, H.; Deng, X.M. Anthocyanin accumulation in apple and peach shoots during cold acclimation. Sci. Hortic. (Amsterdam) 2000, 83, 43–50. [Google Scholar] [CrossRef]
- Christie, P.J.; Alfenito, M.R.; Walbot, V. Impact of low-temperature stress on general phenylpropanoid and anthocyanin pathways: Enhancement of transcript abundance and anthocyanin pigmentation in maize seedlings. Planta 1994, 194, 541–549. [Google Scholar] [CrossRef]
- Becker, C. Impact of Radiation, Temperature and Growth Stage on the Concentration of Flavonoid Glycosides and Caffeic Acid Derivatives in Red Leaf Lettuce. Ph.D. Thesis, Technische Universitat Berlin, Fakultat III-Prozesswissenschaften, Berlin, Germany, 2014. [Google Scholar] [CrossRef]
- Becker, C.; Klaering, H.P.; Kroh, L.W.; Krumbein, A. Cool-cultivated red leaf lettuce accumulates cyanidin-3-O-(6″-O-malonyl)-glucoside and caffeoylmalic acid. Food Chem. 2014, 146, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Becker, C. Flavonoids and phenolic acids in lettuce: How can we maximize their concentration? and why should we? Acta Hortic. 2016, 1142, 1–10. [Google Scholar] [CrossRef]
- Steyn, A.W.J.; Wand, S.J.E.; Holcroft, D.M.; Jacobs, G. Anthocyanins in Vegetative Tissues: A Proposed Unified Function in Photoprotection. New Phytol. 2016, 155, 349–361. [Google Scholar] [CrossRef]
- Oh, M.; Carey, E.E.; Rajashekar, C.B. Plant Physiology and Biochemistry Environmental stresses induce health-promoting phytochemicals in lettuce. Plant Physiol. Biochem. 2009, 47, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Marin, A.; Ferreres, F.; Barberá, G.G.; Gil, M.I. Weather variability influences color and phenolic content of pigmented baby leaf lettuces throughout the season. J. Agric. Food Chem. 2015, 63, 1673–1681. [Google Scholar] [CrossRef] [PubMed]
- Boo, H.-O.; Heo, B.-G.; Gorinstein, S.; Chon, S.-U. Positive effects of temperature and growth conditions on enzymatic and antioxidant status in lettuce plants. Plant Sci. 2011, 181, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Bunning, M.L.; Kendall, P.A.; Stone, M.B.; Stonaker, F.H.; Stushnoff, C. Effects of Seasonal Variation on Sensory Properties and Total Phenolic Content of 5 Lettuce Cultivars. J. Food Sci. 2010, 75, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Chiloane, T.S. Effect of Nutrient Concentration and Growing Seasons on Growth, Yield and Quality of Leafy Lettuce (Lactuca sativa L.) in a Hydroponic System. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2012; pp. 1–79. [Google Scholar]
Figure 1.
The effect of increasing potassium treatment on grow chamber, red ‘Lollo’ lettuce dry mass. The standard error of the mean was: Leaf DM ± 0.04.
Figure 1.
The effect of increasing potassium treatment on grow chamber, red ‘Lollo’ lettuce dry mass. The standard error of the mean was: Leaf DM ± 0.04.
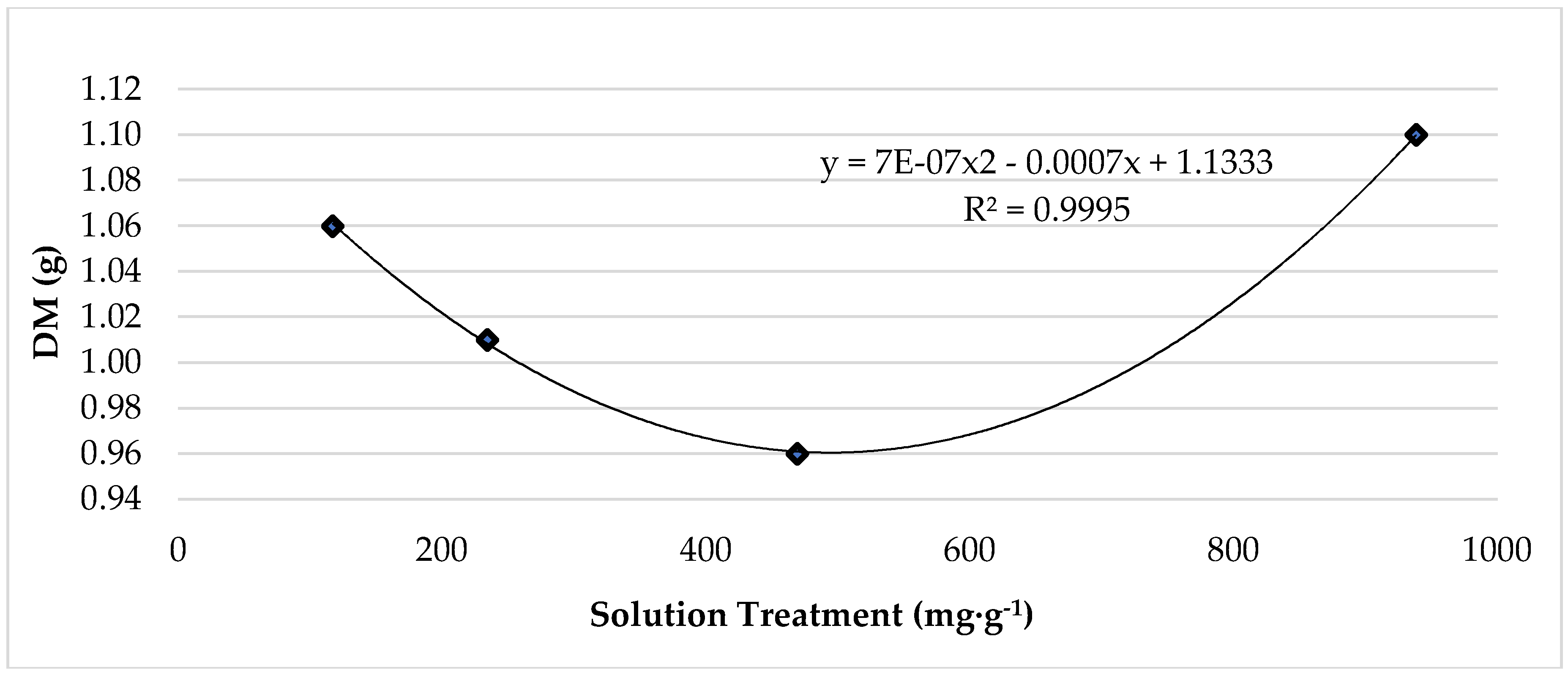
Figure 2.
The effect of temperature on concentrations of growth chamber, red ‘Lollo’ lettuce flavonoids. Abbreviations: qgluc- quercetin glucoside; qglucor- quercetin glucuronide. The standard error of the mean was: qgluc ± 1.77; qglucor ± 7.20; luteolin ± 5.36.
Figure 2.
The effect of temperature on concentrations of growth chamber, red ‘Lollo’ lettuce flavonoids. Abbreviations: qgluc- quercetin glucoside; qglucor- quercetin glucuronide. The standard error of the mean was: qgluc ± 1.77; qglucor ± 7.20; luteolin ± 5.36.
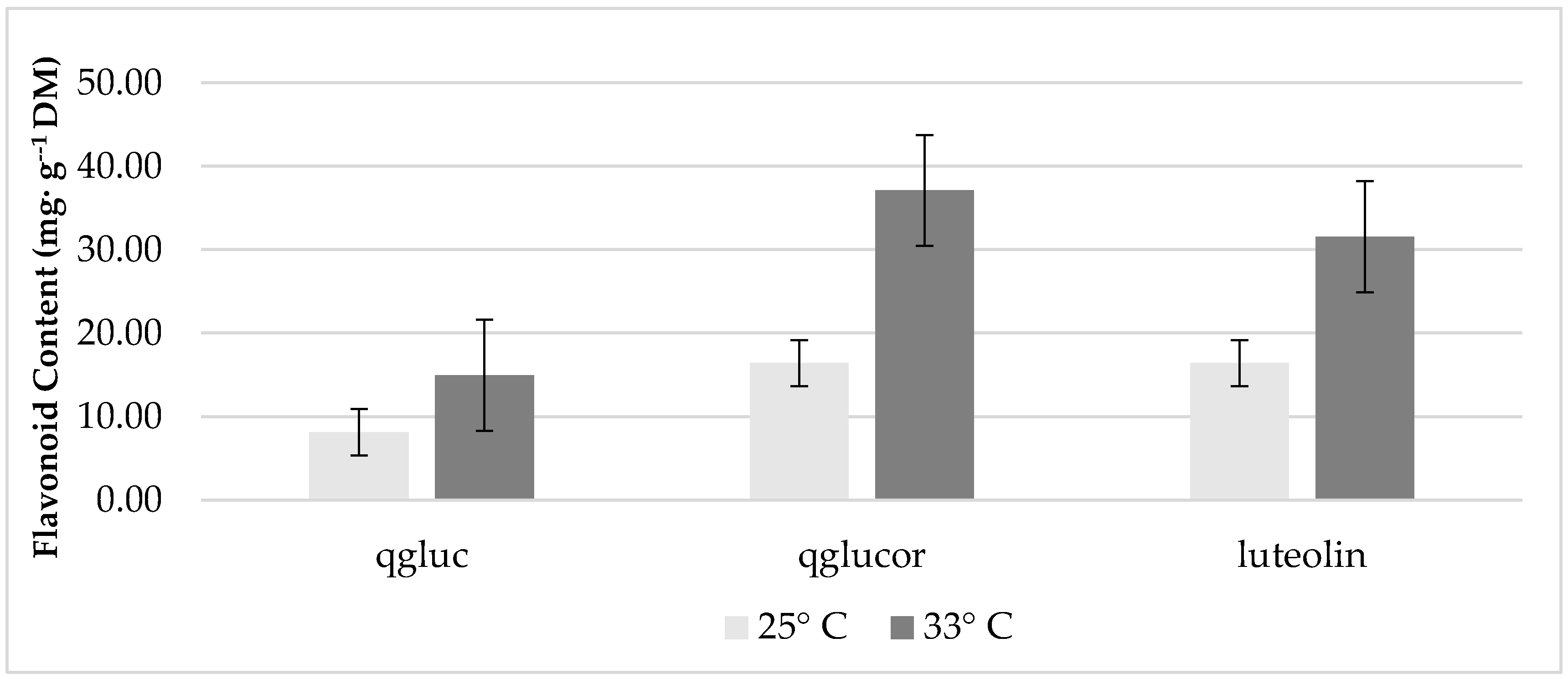
Figure 3.
The effect of increasing potassium concentration of growth chamber produced, red ‘Lollo’ lettuce calcium concentration. The standard error of the mean was: Calcium ± 0.66.
Figure 3.
The effect of increasing potassium concentration of growth chamber produced, red ‘Lollo’ lettuce calcium concentration. The standard error of the mean was: Calcium ± 0.66.

{kind=link}
{kind=link}
{kind=link}
Table 1.
The effect of grow chamber temperature on red, ‘Lollo’ lettuce shoot and root fresh mass, dry mass, DM:FM ratio, and water content.
Table 1.
The effect of grow chamber temperature on red, ‘Lollo’ lettuce shoot and root fresh mass, dry mass, DM:FM ratio, and water content.
| Temp (°C) | Leaf FM (g) | Leaf DM (g) | Leaf DM:FM a | Leaf Water (%) | Root FM (g) | Root DM (g) | Root DM:FM a | Root Water (%) |
|---|---|---|---|---|---|---|---|---|
| 25 | 12.12 a | 0.86 b | 0.07 b | 0.93 a | 10.11 a | 0.40 a | 0.04 a | 0.96 a |
| 33 | 12.47 a | 1.20 a | 0.10 a | 0.90 b | 3.22 b | 0.14 b | 0.05 a | 0.95 a |
| P-value b,c | ns | *** | *** | *** | *** | *** | ns | ns |
a DM:FM ratio is reported in grams of dry to grams of fresh mass; b The standard error of the mean was Leaf FM ± 0.30; Leaf DM ± 0.03; Leaf DM:FM ± 0.002; Leaf Water ± 0.002; Root FM ± 0.80; Root DM ± 0.03; Root DM:FM ± 0.003; Root Water ± 0.003; c ns, *, **, *** indicate non-significant or significant at P ≤ 0.05, 0.01, 0.001, respectively.
Table 2.
The effect of growth chamber temperature on elemental nutrient concentrations of red, ‘Lollo’ lettuce.
Table 2.
The effect of growth chamber temperature on elemental nutrient concentrations of red, ‘Lollo’ lettuce.
| Elemental Nutrient Concentrations a | ||||||||
|---|---|---|---|---|---|---|---|---|
| (mg·g−1 DM) | (μg·g−1 DM) | |||||||
| Temp | Mg | K | Ca | Fe | B | Mn | Zn | Mo |
| 25 | 4.11 a | 76.48 a | 14.25 a | 168.28 a | 32.44 a | 136.07 a | 25.49 a | 0.60 a |
| 33 | 3.32 b | 67.73 b | 12.34 b | 138.46 a | 31.44 a | 79.64 b | 28.17 a | 0.39 b |
| P-value b,c | *** | *** | ** | ns | ns | *** | ns | * |
a Abbreviations: Mg-Magnesium; K-Potassium; Ca-Calcium; Fe-Iron, B-Boron; Mn-Manganese; Zn-. Zinc; Mo-Molybdenum; b The standard error of the mean was Mg ± 0.18; K ± 2.67; Ca ± 0.57; Fe ± 11.68; B ± 0.84; Mn ± 9.22; Zn ± 4.85; Mo ± 0.05; c ns, *, **, *** indicate non-significant or significant at P ≤ 0.05, 0.01, 0.001, respectively.
Table 3.
The effect of the interaction between temperature and treatment on elemental nutrient concentrations of growth chamber red ‘Lollo’ lettuce.
Table 3.
The effect of the interaction between temperature and treatment on elemental nutrient concentrations of growth chamber red ‘Lollo’ lettuce.
| Elemental Nutrient Concentrations a | ||||
|---|---|---|---|---|
| (mg·g−1 DM) | (μg·g−1 DM) | |||
| Temperature (°C) | K Treatment (mg·L−1) | P | S | Cu |
| 25 | 117.3 | 8.50 a | 10.41 a | 2.59 a,b,c |
| 25 | 234.6 | 7.40 b | 10.61 a | 2.64 a,b,c |
| 25 | 469.2 | 7.69 b | 10.59 a | 3.82 a |
| 25 | 938.4 | 7.61 b | 10.02 a,b | 3.30 a,b |
| 33 | 117.3 | 6.20 c | 9.99 ab | 3.22 a,b |
| 33 | 234.6 | 6.54 c | 8.94 a,b | 1.36 b,c,d |
| 33 | 469.2 | 6.33 c | 8.82 a,b | 0.78 c,d |
| 33 | 938.4 | 6.33 c | 9.51 a,b | 0.37 d |
| P-value b,c | * | * | * | |
a Abbreviations: P-Phosphorous; S-Sulfur; Cu-Copper; b The standard error of the mean was p ± 0.22; S ± 0.28; Cu ± 0.70; c ns, *, **, *** indicate non-significant or significant at P ≤ 0.05, 0.01, 0.001, respectively.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sublett, W.L.; Barickman, T.C.; Sams, C.E. Effects of Elevated Temperature and Potassium on Biomass and Quality of Dark Red ‘Lollo Rosso’ Lettuce. Horticulturae 2018, 4, 11. https://doi.org/10.3390/horticulturae4020011
AMA Style
Sublett WL, Barickman TC, Sams CE. Effects of Elevated Temperature and Potassium on Biomass and Quality of Dark Red ‘Lollo Rosso’ Lettuce. Horticulturae. 2018; 4(2):11. https://doi.org/10.3390/horticulturae4020011
Chicago/Turabian StyleSublett, William L., T. Casey Barickman, and Carl E. Sams. 2018. "Effects of Elevated Temperature and Potassium on Biomass and Quality of Dark Red ‘Lollo Rosso’ Lettuce" Horticulturae 4, no. 2: 11. https://doi.org/10.3390/horticulturae4020011
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.