Endophytic Fungi: A Source of Potential Antifungal Compounds



1
TERI-Deakin Nano Biotechnology Centre, The Energy and Resources Institute (TERI), Darbari Seth Block, IHC Complex, Lodhi Road, New Delhi 110003, India
2
Department of Biotechnology, Motilal Nehru National Institute of Technology, Allahabad 211004, India
3
Department of Biotechnology, Thapar Institute of Engineering & Technology, Deemed to be a University, Patiala, Punjab 147004, India
*
Author to whom correspondence should be addressed.
J. Fungi 2018, 4(3), 77; https://doi.org/10.3390/jof4030077
Submission received: 20 May 2018
/
Revised: 13 June 2018
/
Accepted: 16 June 2018
/
Published: 25 June 2018
(This article belongs to the Special Issue Fungal Endophytes in Plants)
Abstract
:The emerging and reemerging forms of fungal infections encountered in the course of allogeneic bone marrow transplantations, cancer therapy, and organ transplants have necessitated the discovery of antifungal compounds with enhanced efficacy and better compatibility. A very limited number of antifungal compounds are in practice against the various forms of topical and systemic fungal infections. The trends of new antifungals being introduced into the market have remained insignificant while resistance towards the introduced drug has apparently increased, specifically in patients undergoing long-term treatment. Considering the immense potential of natural microbial products for the isolation and screening of novel antibiotics for different pharmaceutical applications as an alternative source has remained largely unexplored. Endophytes are one such microbial community that resides inside all plants without showing any symptoms with the promise of producing diverse bioactive molecules and novel metabolites which have application in medicine, agriculture, and industrial set ups. This review substantially covers the antifungal compounds, including volatile organic compounds, isolated from fungal endophytes of medicinal plants during 2013–2018. Some of the methods for the activation of silent biosynthetic genes are also covered. As such, the compounds described here possess diverse configurations which can be a step towards the development of new antifungal agents directly or precursor molecules after the required modification.
1. Introduction
The undisputed potential of fungi to produce bioactive secondary metabolites has long been established. To date, merely 5% of the entire number of species has been elaborated of an estimated 1.5 million. Out of these (69,000 species), merely 16% (11,500) have been cultured [1]. Amongst the fungi, endophytes represent a wide source of unexplored and uncharacterized microorganisms capable of producing novel metabolites. Endophytes generally exist asymptomatically, coexisting with their hosts and representing an underutilized group of microorganisms for the discovery of new compounds. Endophytes produce diverse metabolites and have the ability to synthesize compounds which are solely produced and isolated from higher plants [2,3]. Strobel and Daisy [4] commented that endophytes could be a goldmine of secondary metabolites. Pestalotiopsis sp. can be considered as “the E. coli of the rain forests” and P. microspora, a “microbial factory” of bioactive secondary metabolites. As per them, numerous chemical structures such as Ambuic acid, Cryptocandin, Taxol, Torreyanic acid, Subglutinol A and B, and many others have been identified. An array of metabolites of different chemical classes profiles have been deciphered, such as alkaloids, cytochalasines, flavonoids, furandiones, phenylpropanoids, lignans, peptides, phenol, phenolic acids, steroids, terpenoids, quinones, aliphatic acid, and chlorinated compounds. Secondary metabolites derived from endophytes comprise classes of compounds such as steroids, xanthones, phenols, isocoumarins, perylene derivatives, quinines, furandiones, terpenoids, depsipeptides, and cytochalasins, which are identified to possess biological activities with antibiotic, antiviral, volatile antibiotic, anticancer, antioxidant, insecticidal, antidiabetic, and immunosuppressive properties [5,6,7,8,9]. Endophytes play a major role in the physiological activities of host plants, influencing the enhancement of stress, insects, nematodes, and disease resistance [10,11,12,13].
This review covers the antifungal fungal metabolites reported from endophytic fungi from medicinal plants during 2013–2018 and their potential as antifungal agents. The antifungal activity of these compounds against the selected fungal pathogens are described briefly and some details such as producing organisms, plant sources, place of collections, and the antifungal properties of many of these compounds are shown in Table 1 and Table 2.
2. Medicinal Plants
2.1. Compounds Produced by Coelomycetes
Pestalotiopsis is an important genus of coelomycetes and different species of this genus have been identified for production of bioactive compounds for various biological properties which include antimicrobial, antifungal, antiviral, antineoplastic, and antioxidant activities [14]. Some of the antifungal reported from this genus includes a new monoterpene lactone, (3R,4R,6R,7S)-7-hydroxyl-3,7-dimethyl-oxabicyclo[3.3.1]nonan-2-one (1) (Figure 1), along with one related known compound, (3R,4R)-3-(7-methylcyclohexenyl)-propanoic acid (2) (Figure 1), were discovered from endophytic fungus Pestalotiopsis foedan obtained from the branch of Bruguiera sexangula in Hainan, China. Compounds 1–2 showed antifungal activity against Botrytis cinerea and Phytophthora nicotianae with MIC values of 3.1 and 6.3 µg/mL, respectively, while the known antifungal drug ketoconazole showed comparable activity (MIC 3.1 µg/mL each) Compound 2 also exhibited satisfactory activity against Candida albicans (MIC value of 50 µg/mL) while ketoconazole showed MIC of 6.3 µg/mL [15].
Plants of Dendrobium officinale were collected in Yandang Mountain, Zhejiang Province, China. Pestalotiopsis sp. DO14 was obtained from the shoots of D. officinale endophytic fungus which yielded two novel antifungal constituents, (4S,6S)-6-[(1S,2R)-1,2-dihydroxybutyl]-4-hydroxy-4-methoxytetrahydro-2H-pyran-2-one (3) and (6S,2E)-6-hydroxy-3-methoxy-5-oxodec -2-enoic acid (4), and two known compounds, LL-P880γ (5) and LL-P880α (6) (Figure 1) were isolated. Compounds 3–6 exhibited good anti-fungal activities (MIC ≤ 50 µg/mL) against C. albicans, Cryptococcus neoformans, Trichophyton rubrum, and Aspergillus fumigatus. Compounds 3 and 4 possess the strong activities with the MIC values ≤25 µg/mL against tested strains [16].
Endophytic fungus Pestalotiopsis fici obtained from the branches of Camellia sinensis collected from the suburb of Hangzhou, China was the source of a new a-pyrone derivative ficipyrone A (7) (Figure 1). Compound 7 showed antifungal activity against the plant pathogen Gibberella zeae with an IC50 value of 15.9 µM (the positive control ketoconazole showed an IC50 value of 6.02 µM) [17].
Endophytic fungus Pestalotiopsis mangiferae associated with Mangifera indica Linn collected from Maduravoyal, Tamil Nadu Province, India was the source of a new phenolic compound 4-(2,4,7-trioxa-bicyclo[4.1.0]heptan-3-yl) phenol (8) (Figure 1). Compound 8 exhibited strong antifungal activity against C. albicans with MIC value of 0.039 µg/mL, while nystatin showed MIC 10.0 µg/mL [18].
Phomopsis is an important genus that is a prolific producer of bioactive compounds including Cytochalasin H (9) (Figure 1), which was isolated from the endophytic fungus Phomopsis sp. of Senna spectabilis (Fabaceae) collected from São Paulo, Brazil. Compound 9 exhibited activity against Cladosporium cladosporioides and C. sphaerosphermum with MIC values of 10.0 and 25.0 µg, respectively, while the MIC of nystatin the reference compound was 1.0 µg [19].
Endophytic fungus Phomopsis sp. isolated from Aconitum carmichaeli collected in Huize County, Yunnan Province, China yielded (14β,22E)-9,14-dihydroxyergosta-4,7,22-triene-3,6-dione (10) and (5α,6β,15β,22E)-6-ethoxy-5,15 -dihydroxyergosta-7,22-dien-3-one (11), calvasterol A (12), and ganodermaside D (13) (Figure 1). All compounds were evaluated for their antifungal activities against Candida albicans, Aspergillus niger, Pyricularia oryzae, Fusarium avenaceum, Hormodendrum compactum, and Trichophyton gypseum. Compound 10 exhibited average antifungal activities against C. albicans, H. compactum, and A. niger, with MIC values of 64, 64, and 128 µg/mL, respectively. Compound 11 showed poor inhibitory activity against C. albicans and F. avenaceum with MIC values of 128 µg/mL. Compounds 12 and 13 showed average inhibitory activities against F. avenaceum (MIC 64 µg/mL for both compounds). Only compound 12 exhibited weak antifungal activities against P. oryzae and T. gypseum (MIC of 128 and 256 µg/mL, respectively) [20].
Diaporthe maritima an endophytic fungus obtained from needles of Picea sp. in the Acadian forest of Eastern Canada yielded three dihydropyrones, phomopsolides A (14), B (15), and C (16), and a stable alpha-pyrone (17) (Figure 1). Compound 14 demonstrated growth inhibition at 25 µM against Microbotryum violaceum and Saccharomyces cerevisiae whereas Compounds 15–17 were active at 250 µM [21].
Another coelomycete Phoma is known to produce diverse compounds [22]. From Phoma sp. an endophytic fungus of the plant Fucus serratus yielded phomalacton (18), (3R)-5-hydroxymellein (19) and emodin (20) (Figure 1). Phomalactone (18), (3R)-5-hydroxymellein (19) and emodin (20) Compounds 18–20 exhibited antifungal activity against Microbotryum violaceum with 5, 6 and 5 mm zone of inhibition (0.05 mg was pipetted onto 9 mm sterile filter disk) [23].
Viridicatol (21) (Figure 1), tenuazonic acid (22), alternariol (23), and alternariol monomethyl ether (24) (Figure 1) were isolated from endophytic fungi Phoma sp. WF4 of Eleusine coracana grown under semi-hydroponic conditions Arkell Field Station, Arkell, ON, Canada. Compounds 21–24 caused reasonable breakage of Fusarium graminearum hyphae in vitro [24].
Endophytic fungus Rhizopycnis vagum Nitaf 22 obtained from the healthy root of Nicotiana tabacum grown at China Agricultural University Beijing, China was the source of Rhizopycnin D (25) and TMC-264 (26) (Figure 2). Compounds 25 and 26 showed strong inhibition of the spore germination of Magnaporthe oryzae with IC50 values of 9.9 and 12.0 µg/mL, respectively [25].
A new polychlorinated triphenyl diether named microsphaerol (27) (Figure 2) has been isolated from the endophytic fungus Microsphaeropsis sp. (internal strain No. 8883), and phytochemical investigation of the endophytic fungus Seimatosporium sp. (internal strain No. 8883) associated with Salsola oppositifolia from Playa del Ingles (Gomera, Spain) led to the isolation of a new naphthalene derivative named seimatorone (28) (Figure 2). In antifungal assay, compounds 27 and 28 showed activity against Microbotryum violaceum with 9 and 5 mm zone of inhibition (0.05 mg was pipetted onto 9 mm sterile filter paper disk). In addition, there was some growth within the zone of inhibition [26].
Endophytic fungus Colletotrichum gloeosporioides associated with Michelia champaca isolated from São Paulo State University (UNESP), Araraquara, São Paulo, Brazil was found to be source of a new compound, 2-phenylethyl 1H-indol-3-yl-acetate (29) (Figure 2). Compound 29 displayed good activity against Cladosporium cladosporioides and C. sphaerospermum which was analogous to nystatin, the positive control [27].
Colletonoic acid (30) (Figure 2) was isolated from Colletotrichum sp. from Gomera (Spain). Colletonoic acid exhibit antifungal activity against Microbotryum violaceum with 7 mm zone of inhibition (0.05 mg was pipetted onto 9 mm a sterile filter paper disk) [28].
Coniothyrium sp., an endophytic fungus associated with Salsola oppostifolia from Gomera in the Canary Islands, was the source of known hydroxy anthraquinones 1,7-dihydroxy3-methyl-9,10-anthraquinone (31), 1,6-dihydroxy-3-methyl-9,10-anthraquinone (phomarin) (32), and 1-hydroxy-3-hydroxymethyl-9,10-anthraquinone (33) (Figure 2) along with four new derivatives having a tetralone moiety, namely coniothyrinones A–C (34–36) (Figure 6) and D (37) (Figure 2). The absolute configurations of coniothyrinones A (34), B (35), and D (37) were determined by TDDFT calculations of CD spectra, allowing the determination of the absolute configuration of coniothyrinone C (36) as well. Coniothyrinones A (34), B (35), and D (37) could be used as ECD reference compounds in the determination of absolute configuration for related tetralone derivates. Compounds 31–37 showed inhibitory effects against the fungus Microbotryum violaceum with 7, 10, 8, 7.5, 6, 8 and 7.5 mm zone of inhibition (0.05 mg/9-mm sterile filter paper disk). Compounds 32 and 34 exhibited strong antifungal activity against M. violaceum (10 and 9 mm zone of inhibition) and B. cinerea (7.5 and 12.5 mm zone of inhibition) when tested under similar conditions [29].
2.2. Compounds Produced by Ascomycetes
Xylaria is very important ascomycetous genus and a good sources of novel bioactive compounds, and some of the compounds reported to have drug-able properties relevant for drug discovery [30,31]. Nine oxygenated guaiane-type sesquiterpenes (38–46) and three isopimarane diterpenes (47–49), (Figure 2) were obtained from Xylaria sp. YM 311647, an endophytic fungus associated with Azadirachta indica collected from Yuanjiang County, Yunnan Province, China. All compounds were evaluated for their antifungal activities against Candida albicans, Aspergillus niger, Pyricularia oryzae, Fusarium avenaceum, and Hormodendrum compactum. Compounds 38–46 were moderately active against C. albicans and H. compactum (MIC values ranging from 32 to 256 µg/mL), while compound 47–49 were more active against all the tested strains (MIC values ranging from 16 to 256 µg/mL). Compound 49 exhibited the most promising activity against C. albicans and P. oryzae with MIC values of 16 µg/mL [32].
Endophytic fungus Xylaria sp. YM 311647 associated with Azadirachta indica from Yuanjiang County, China was also reported to produce five new guaiane sesquiterpenes, (50–54) (Figure 3). The antifungal activities of 50–54 were evaluated by means of the broth microdilution method against C. albicans, A. niger, P. oryzae, F. avenaceum and H. compactum Compounds 50–54 exhibited average or poor antifungal activities against P. oryzae and H. compactum (MIC values in the range of 32–256 µg/mL). Among them, 53 exhibited the most promising inhibitory activity against P. oryzae with a MIC value of 32 µg/mL. Compounds 52 and 53 showed average antifungal activities against H. compactum with MIC values of 64 µg/mL. In addition, 53 and 54 exhibited the most promising antifungal activities against C. albicans with MIC values of 32 µg/mL. Compound 52 showed average inhibitory activities against C. albicans, A. niger, and H. compactum with MIC values of 64 µg/mL. All compounds showed no notable inhibitory activities against Fusarium avenaceum [33]. Amazonian endophytic fungus X. feejeensis residing in Croton lechleri yielded nonenolide, xyolide (55) (Figure 3). Compound 55 exhibited antifungal activity against oomycetes Pythium ultimum with a MIC value of 425 µM [34].
Endophytic fungus Xylaria sp. XC-16 associated with Toona sinensis was isolated from Yangling, Shaanxi Province, China and was observed to produce a potent antifungal compound Cytochalasin Z28 (56) (Figure 3), displaying enhanced activity with an MIC of 12.5 µM as opposed to the antifungal activity possessed by hymeaxszol possessing an MIC value of 25 µM against the plant pathogen Gibberella saubinetti [35].
Various isolates of Xylaria produce griseofulvin (57) (Figure 3); Xylaria sp. PSU G12 associated with Garcinia hombroniana [36], and X. cubiensis residing in Asimina triloba [37], along with 13 strains of Xylaria sp. inhabiting Pinus strobus and six strains associated with Vaccinum augustifolium found in the Acadian forest of New Brunswick and Nova Scotia Canada [38] are known to produce griseofulvin and a few of them can also produce dechlorogriseofulvin (58) (Figure 3) [36,38].
Griseofulvin (57) is very potent against the phytopathogenic fungi, but not against oomycetes [38]. Griseofulvin has been validated to possess antifungal activity against Alternaria mali, B. cinerea, C. gloeosporioides, Corticium sasaki, Fusarium oxysporum and Magnaporthe grisea in vitro with IC50 values of 18.0, 5.0, 1.7, 11.0, 30.0, and 1.7 µg/mL, respectively. Dechlorogriseofulvin (58) demonstrated poor activity, with an IC50 value of 200 µg/mL for each fungus. Griseofulvin (57) also hinders the growth of M. grisea, C. sasaki, B. cinerea, Puccinia recondite and Blumeria graminis f. sp. hordei in vivo, with a percentage of fungal control of 95, 100, 60, 90 and 90, respectively, at 150 µg/mL. Griseofulvin (57) is used to cure dermatophytic infections caused by fungi such as Epidermophyton and Trichophyton species [38].
Chaetomium is another genus of ascomycete and prolific producer of bioactive compounds [39,40]. Chaetoglobosin A (59) and D (60) (Figure 3) were isolated Chaetomium globosum CDW7, an endophyte from Ginkgo biloba located in Taixing and Nanjing in Jiangsu Province and Chengdu in Sichuan Province, China. Compounds 59 and 60 showed antifungal activity against Sclerotinia sclerotiorum with IC50 values of 0.35 and 0.62 µg/mL, respectively, compared with carbendazim (0.17 µg/mL) [41].
Compounds Chaetomugilin A (61), Chaetomugilin D (62) Chaetoglobosin A (59), Chaetoglobosin B (63), Chaetoglobosin E (64), Chaetoglobosin F (65), and Penochalasin G (66) (Figure 3) were obtained from C. globosum endophyte obtained from seeds of Panax notoginseng collected at the Wenshan, Yunnan, China. Compounds 59 and 61–66 exhibited antifungal activity against Phoma herbarum (MIC in the range of 16–128 µg/mL) and Epicoccum nigrum (MIC in the range of <1–16 µg/mL). Both fungi are phytopathogenic fungi causing root rot of Panax notoginseng [42].
Ergosta-5,7,22-trien-3beta-ol (67) (Figure 3) was isolated from Chaetomium cupreum ZJWCF079 of Macleaya cordata. It exhibited antifungal activity against Sclerotinia sclerotiorum and B. cinereal, plant pathogenic fungi with EC50 values of 125 µg/mL and 190 µg/mL respectively, but had no effects on Pythium ultimum, Rhizoctonia solani and F. oxysporum [43].
Chaetoglobosin A (59) (Figure 3), D (60), E (64), C (68), (Figure 3) G (69), and R (70) (Figure 4) were isolated from Chaetomium globosum No.04 obtained from barks of Ginkgo biloba, growing in Linyi, Shandong Province, China. Compounds 59, 60, 64, and 68–70 showed good growth inhibitory activity at a concentration of 20 µg/disk, against Rhizopus stolonifer and Coniothyrium diplodiella [44].
A new tetranorlabdane diterpenoids botryosphaerin H (71) and a known tetranorlabdane diterpenes 13,14,15,16-tetranorlabd-7-en-19,6β:12,17-diolide (72) (Figure 4) were obtained from Botryosphaeria sp. P483, an endophyte of Huperzia serrata collected in Xichou County, Yunnan Province, China. When tested at 100 µg/disk, compound 71 showed zone of inhibition of 9, 7, 7, 8, and 8 mm, against Gaeumannomyces graminis, Fusarium solani, Pyricularia oryzae, Fusarium moniliforme, and F. oxysporum while compound 72 showed zone of inhibition of 12, 10, 10, 11, 13 mm against G. graminis, F. solani, P. oryzae, F. moniliforme, and F. oxysporum. The standard Carbendazim (50 µg/disk) exhibited activity against G. graminis, F. solani, P. oryzae, F. moniliforme, and F. oxysporum, with the zone of inhibition of 14, 18, 15, 17 and 15, mm respectively [45].
Endophytic fungus Botryosphaeria dothidea KJ-1 associated with the stems of Melia azedarach collected at Yangling, Shaanxi Province, China was the source of pycnophorin (73), stemphyperylenol (74), chaetoglobosin C (68), djalonensone (75) (Figure 4), alternariol (76), β-sitosterol glucoside (77), and 5-hydroxymethylfurfural (78) (Figure 4). Stemphyperylenol (74) exhibited good antifungal activity against Alternaria solani the plant pathogen with the MIC value of 1.57 µM comparable to commonly used fungicide, the carbendazim. Compounds 68, 73, and 75–78 showed good to average antifungal activities against A. solani (MICs of 6.25−25 µM) [46].
Two eicosanoic acids, 2-amino-3,4-dihydroxy-2-25-(hydroxymethyl)-14-oxo-6,12-eicosenoic acid (79) and myriocin (80) (Figure 4), were isolated from Mycosphaerella sp. an endophytic fungus of Eugenia bimarginata DC. (Myrtaceae) collected in Brazil (Savannah). These compounds displayed antifungal activities against several isolates of C. neoformans and C. gattii, with MIC values for compound 79 ranging from 1.3 to 2.50 µg/mL and for compound 80 was 0.5 µg/mL [47]. Both compounds exhibited antifungal activities against several isolates of C. neoformans and C. gattii, with MIC values ranging from 0.49 to 7.82 µM for compound 79 and 0.48–1.95 µM for compound 80 in another study. When checked by the checkerboard microtiter assay, both compounds exhibited synergistic activity against C. gattii with amphotericin B. Ultrastructural analysis divulges various signs of damage in C. gattii and C. neoformans cells treated with compounds 79 and 80, including deformities in cell shape, depressions on the surface, and withered cells. Compounds 79 and 80 showed less loss of cellular material in cells of C. gattii compared to those treated with amphotericin B.
The difference in cellular material loss increased in a test compound concentration-dependent manner. Compound 80 also induced the formation of several pseudohyphae, suggesting that it could reduce virulence in C. gattii cells [48].
Endophytic fungus Guignardia sp., associated with Euphorbia sieboldiana collected from Nanjing, Jiangsu, China was the source of guignardone N (81) and guignardic acid (82) (Figure 4). Both compounds were evaluated for their inhibitory effects alone and with fluconazole on the growth and biofilms of Candida albicans. At 6.3 µg/mL combined with 0.031 µg/mL of fluconazole, compounds 81 and 82 were found to have prominent inhibition on the growth of C. albicans with fractional inhibitory concentration (FIC) index values of 0.23 and 0.19, respectively. Combined with fluconazole, both (40 µg/mL for (81) and 20 µg/mL for (82) could also inhibit C. albicans biofilms and reverse the tolerance of C. albicans biofilms to fluconazole [49].
Antifungal hyalodendriol C (83), rhizopycnin D (84), palmariol B (85), TMC-264 (86), penicilliumolide B (87) and alternariol 9-methyl ether (88) (Figure 4) were obtained from the endophytic fungus Hyalodendriella sp. Ponipodef 12 associated with the healthy stems of the “Neva” hybrid of Populus deltoides Marsh × P. nigra L. were collected from Longhua in Hebei Province of China. Compound 83 displayed antifungal effects against the spore germination of M. oryzae with potent inhibition with the IC50 value of 11.6 µg/mL, which was comparable with the positive control, carbendazim (IC50 6.9 µg/mL) [50]. Previously, it found that rhizopycnin D (84), palmariol B (85), TMC-264 (86), penicilliumolide B (87), and alternariol 9-methyl ether (88) exhibits antifungal activity against the spore germination of M. oryzae [51,52].
Mellein (89) (Figure 4), was isolated from Pezicula sp. associated with the twigs of Forsythia viridissima, Zhejiang Province, Southeast China. Antifungal activity of this compound was tested against B. cinerea, Pythium ultimum, Fusarium oxysporium f. sp. cucumerinum, Colletotrichum orbiculare, Verticillium dahliae, Pyricularia oryzae, Pestalotia diospyri, Sclerotinia sclerotiorum and Fulvia fulva. Compound 89 displayed antifungal activity against 9 plant pathogenic fungi, esp. B. cinerea and F. fulva with EC50 values below 50 µg/mL [53].
Endophytic fungus Nodulisporium sp. A21 associated with the leaves of Ginkgo biloba collected from Nanjing in Jiangsu Province, China was a source of anti-phytopathogenic sporothriolide (90) (Figure 4). In mycelia growth inhibition method, sporothriolide (90) showed antifungal activity against Rhizoctonia solani with the EC50 value of 3.04 µg/mL (11.6 µM) while the EC50 of positive control carbendazim was 1.84 µg/mL (9.6 µM). Sporothriolide (90) at 200 µg/mL had a protective efficacy of 71.7% against Rice Sheath Blight in comparison with the protective efficacy 90.1% of the positive control of validamycin A at 200 µg/mL. Conidia of Magnaporthe oryzae could not form the germ tube and appressorium germinate in the sporothriolide with solution at the concentration of 1.5 µg/mL (5% DMSO). In vivo, sporothriolide at 50 µg/mL, compared to tricyclazole of 2.5 µg/mL, could control the developing of Rice Blast [54]. Sporothriolide (90) also showed antifungal activity against Sclerotinia sclerotiorum with EC50 of sporothriolide against was 2.78 µg/mL (10.7 µM) while the EC50 of positive control carbendazim was 0.17 µg/mL (0.89 µM). When it comes to the protective activity on rape leaves, the positive control carbendazim of 250 µg/mL was 57.6% and sporothriolide of 250 µg/mL was 41.5% [54].
Six phenolic bisabolane-type sesquiterpenoids (91, 92) (Figure 4) (93–96), along with a macrolide, pyrenophorin (97) (Figure 5) were isolated from Lopherdermium nitens DAOM 250027 endophyte of Pinus strobus (eastern white pine) near Sussex, NB, Canada. These compounds were characterized based on interpretation of spectroscopic data (NMR, OR, UV) and HRMS. All compounds were tested for antifungal activity. Pyrenophorin (97) significantly reduced the growth of Microbotryum violaceum and Saccharomyces cerevisiae at 5 µM, whereas sesquiterpenoids (91–96) were antifungal at 50 µM to both species tested [55]. Isocoumarin derivative exserolide C (98) and (12R)-12-hydroxymonocerin (99) (Figure 5) were isolated from endophyte Exserohilum sp. associated with Acer truncatum collected from Beijing, China. Compounds 98 and 99 displayed antifungal activity against Fusarium oxysporum, with MIC value of 20 µg/mL for both compounds, while Amphotericin B the positive control showed the MIC value of 0.63 µg/mL [56].
Endophytic fungus of Echinacea purpurea associated with Biscogniauxia mediterranea EPU38CA from the wild in Missouri, USA was the source of (−)-5-methylmellein (100) and (−)-(3R)-8-hydroxy-6-methoxy-3,5-dimethyl-3, 4-dihydroisocoumarin (101) (Figure 5). Compound 100 exhibited poor activity against Phomopsis obscurans, P. viticola, and Fusarium oxysporum, and stimulated the growth of Colletotrichum fragariae, C. acutatum, C. gloeosporioides, and B. cinerea. Compound 101 was acknowledged to be marginally more active in the microtiter method than 5-methylmellein [57]. Compound 101 was found to be slightly more active in the microtiter method than 5-methylmellein [57]. Trienylfuranol A (102) (Figure 5) was isolated from isolated from Hypoxylon submonticulosum the endophyte of Rubus idaeus collected from Jordan Station, ON, Canada. It was identified based on high-resolution LC-MS and 1- and 2-D NMR spectroscopy. Absolute stereochemical configurations of the compounds were confirmed by NOE NMR experiments and by the preparation of Mosher esters. Complete hydrogenation of I yielded THF 7 (103) (Figure 5) that was used for stereochemical characterization and assessment of antifungal activity. Compound THF 7 (103) significantly inhibited the growth of Saccharomyces cerevisiae (74 ± 4% inhibition) at a concentration of 250 µg/mL as compared with complete inhibition by nystatin at 10 µg/mL [58].
Endophytic fungus Phialophora mustea associated with Crocus sativus was the source of an unprecedented azaphilone derived skeleton, Phialomustin C (104) and D (105) (Figure 5). Compounds 104 and 105 showed potent activities against Candida albicans, with IC50 values of 14.3 and 73.6 µM respectively [59].
An unidentified ascomycete, associated with Melilotus dentatus was the source of two new polyketide metabolites cis-4-acetoxyoxymellein (106) and 8-deoxy-6-hydroxy-cis-4-acetoxyoxymellein (107) (Figure 5). Compounds 106 and 107 displayed potent antifungal activities toward Microbotryum violaceum and B. cinerea, with 8 mm zone of inhibition for both fungi tested (0.05 mg was pipetted onto 9 mm sterile filter paper disk). In the case of B. cinereal, there was some growth within the zone of inhibition [60].
(−)-Mycorrhizin A (108) (Figure 6) was isolated from Plectophomella sp. while cytochalasins E (109) and K (110) (Figure 5) were isolated from Physalospora sp. Similarly, radicinin (111) (Figure 5) was purified from the endophytic fungus Crataegus monogyna. (-)-Mycorrhizin A showed good antifungal activity towards Ustilago violacea and Eurotium repens. Cytochalasins E (109) and K (110) showed potent activity against E. repens and Mycotypha microspora. Radicinin (111) (Figure 5) showed good activity against E. repens and M. microspore [61].
Diepoxin ζ (112), palmarumycin C11 (113), palmarumycin C12 (114) (Figure 5), cladospirone B (115), palmarumycin C6 (116), 1,4,7β-trihydroxy-8-(spirodioxy-1′,8′-naphthyl)-7,8-dihydronaphthalene (117), and palmarumycin C8 (118) (Figure 6) were obtained from Berkleasmium sp., an endophyte associated with Dioscorea zingiberensis from Hubei Province, China. Compounds 112–118 were evaluated for their antifungal activity against the spore germination of M. oryzae. Compounds 112–118 inhibited spore germination of M. oryzae with IC50 values in the range 9.1–124.5 µg/mL. Palmarumycin C8 (118) showed the best inhibitory activity (IC50 9.1 µg/mL) among the compounds tested, although not as active as the positive control carbendazim (IC50 6.3 µg/mL) [62].
Bipolamide B (119) (Figure 6) was isolated from Bipolaris sp. MU34, the endophytic fungus associated with the leaves of Gynura hispida Thwaites collected from Mahidol University, Bangkok, Thailand. The compounds were characterized based on NMR and MS experiments. Bipolamide B (119) exhibited average antifungal activity with MIC values of 16, 32, 32, 64 and 64 µg/mL, against Cladosporium cladosporioides, C. cucumerinum, Saccharomyces cerevisiae, Aspergillus niger and Rhizopus oryzae respectively [63].
Altenusin (120) (Figure 6), a biphenyl derivative, was isolated from an endophytic fungus, Alternaria alternata Tche-153 of Terminalia chebula, collected from Bangkok, Thailand. Employing disk diffusion method and the microdilution checkerboard technique, altenusin (120) in amalgamation with each of three azole drugs, ketoconazole, fluconazole or itraconazole at their low sub-inhibitory concentrations displayed potent synergistic activity against C. albicans with the fractional inhibitory concentration index range of 0.078 to 0.188 [64]. It is reported that Schizosaccharomyces pombe cells treated with altenusin were more rounded in shape than untreated cells which suggest that altenusin could act through the inhibition of cell wall synthesis or assembly in S. pombe [65].
Cladosporin (121) and isocladosporin (122) (Figure 6) were isolated from endophytic fungus Cladosporium cladosporioides. Compound 121 exhibited growth inhibition against Colletotrichum acutatum, C. fragariae, C. gloeosporioides and Phomopsis viticola at 30 µM with 92.7%, 90.1%, 95.4%, and 79.9%, respectively. Similarly, compound 122 showed 50.4%, 60.2%, and 83.0% growth inhibition against C. fragariae, C. gloeosporioides, and P. viticola, respectively, at 30 µM [66].
Epicolactone (123) and epicoccolide A (124) and B (125) (Figure 6) polyoxygenated polyketides were obtained from an endophytic fungus, Epicoccum sp. CAFTBO, associated with stem bark and leaves of Theobroma cacao of Mount Kala, Republic of Cameroon. Compounds (123–125) showed good inhibitory effects on the mycelial growth of Pythium ultimum and Aphanomyces cochlioides and Rhizoctonia solani (MIC in the range of 20–80 µg per paper disc) [67].
5-methylmellein (100) (Figure 5) was isolated from endophytic fungus Biscogniauxia mediterranea Ohu 19B obtained from Opuntia humifusa (Cactaceae) of United States. Antifungal activity of compound 100 was evaluated using an in vitro microdilution broth assay against seven plant pathogens i.e., Colletotrichum acutatum, C. fragariae, C. gloeosporioides, Fusarium oxysporum, B. cinerea, Phomopsis obscurans, and P. viticola. Phomopsis obscurans was found to be being most susceptible (63.5% growth inhibition) at 150 µM at 120 h. The best growth inhibition (20.1%) to F. oxysporum was at 300 µM at 48 h. Lower doses (75 and 150 µM) of this compound caused stimulation of B. cinera and C. fragariae, while all doses caused stimulation of C. acutatum and C. gloeosporioides [68].
5-(undeca-3′,5′,7′-trien-1′-yl)furan-2-ol (126) and 5-(undeca-3′,5′,7′-trien-1′-yl)furan-2-carbonate (127) (Figure 6), two new alkylated furan derivatives, were recovered from the endophytic fungus Emericella sp. XL029 associated with the leaves of Panax notoginseng collected from Shijiazhuang, Hebei Province, China. Compound 126 displayed good antifungal activity against Rhizoctorzia solani, Verticillium dahliae, Helminthosporium maydis, Fusarium oxysporum, Fusarium tricinctum, Botryosphaeria dothidea, and Alternaria fragriae with MIC values ranging from 25 to 3.1 µg/mL, while compound 127 was found active against V. dahliae, H. maydis, F.tricinctum, B. dothidea, and A. fragriae with MIC values ranging from 50 to 12.5 µg/mL [69].
2.3. Compounds Produced by Hyphomycetes
5-hydroxy 2(3H)-benzofuranone (128), dehydrocostus lactone (129) and harpagoside (130) (Figure 6) were isolated from Fusarium fujikuroi, Penicilium chrysogenum and Penicillium expensum endophytes of Eleusine coracana grown under semi-hydroponic conditions Arkell Field Station, Arkell, ON, Canada. Compounds 128–130 exhibited antifungal activity against F. graminearum with the MIC of 31.25, 250.00 and 31.25 µg/mL, respectively. An in vitro interaction between each compound and Fusarium was investigated using light microscopy and vitality staining where the results proposed a mixed fungicidal/fungistatic mode of action [70].
Endophytic fungus Trichoderma koningiopsis YIM PH30002 harbored in Panax notoginseng collected from Wenshan, Yunnan Province, China was the source of koninginin O (131), koninginin Q (132) and 7-O-methylkoninginin D (133) (Figure 6). The antifungal activities of these compounds were tested against phytopathogenic fungi, Fusarium oxysporum, F. solani, F. flocciferum, Plectosphaerella cucumerina and Alternaria panax which are causes of pathogens of root rot diseases of P. notoginseng. Koninginin O (131) and koninginin Q (132) exhibited poor activity against F. oxysporum and P. cucumerina (MIC of 128 µg/mL). 7-O-methylkoninginin D (133) also showed poor activity against P. cucumerina (MIC 128 µg/mL). Nystatin positive control showed antifungal activity with MICs at 32 µg/mL [71]. Koningiopisin C (134) (Figure 6) was also isolated from the same fungus. Koningiopisin C showed antimicrobial activities against F. oxysporum, A. panax, F. solani and P. cucumerina with MICs at 32, 64, 32, and 16 µg/mL, respectively [72].
Dichlorodiaportinolide (135) and dichlorodiaportin (136) (Figure 6) were isolated from endophytic fungus Trichoderma sp. 09 obtained from the root of Myoporum bontioides A. Dichlorodiaportinolide (135) and dichlorodiaportin (136) showed weak to high antifungal activities with MIC values ranging from 6.25 to 150 µg/mL against Colletotrichum musae and Rhizoctonia solani and were inactive to Penicillium italic and Fusarium graminearum (MIC values > 200 µg/mL) [73].
Trichodermin (137) (Figure 6) was isolated from endophytic fungus strain, Trichoderma brevicompactum 0248 obtained from Allium sativum. Trichodermin showed potent inhibitory activity against Rhizoctonia solani, with an EC50 of 0.25 µg/mL and against B. cinerea, with an EC50 of 2.02 µg/mL but relatively poorly active against Colletotrichum lindemuthianum (EC50 = 25.60 µg/mL). Compound 137 exhibited good antifungal activity against the tested phytopathogens compared with the positive control Carbendazim [74].
Trichoderma koningiopsis YIM PH30002 collected at Wenshan, Yunnan Province of China was the source of two new metabolites koninginins R and S (138–139) (Figure 7). These isolated compounds showed certain antifungal activities against phytopathogens, Fusarium flocciferum and Fusarium oxysporum. Compound 138 possess the weak activity against F. oxysporum and F. flocciferum with the MICs at 128 µg/mL, while compound 139 displayed the poor activity against F. oxysporum with the MIC at 128 µg/mL [75].
Stigmasterol derivative (22E,24R)-stigmasta-5,7,22-trien-3-β-ol (140) and a new butyrolactones, aspernolide F (141) (Figure 7) were obtained from the endophytic fungus Aspergillus terreus associated with the roots of Carthamus lanatus collected at Assiut, Egypt. Compounds 140–141 exhibited good activity against C. neoformans with IC50 values of 4.38 and 5.19 µg/mL respectively, compared to amphotericin B (IC50 0.34 µg/mL) [76].
Fonsecinone A (142), and (R)-3-hydroxybutanonitrile (143) (Figure 7), were obtained from Aspergillus sp. KJ-9 an endophytic fungus associated with Melia azedarach which was collected at Yangling, Shaanxi Province, China, and identified by spectroscopic methods. Compounds 142 and 143 were active against Gibberella saubinetti, Magnaporthe grisea, B. cinerea, C. gloeosporioides and A. solani (MIC range of 6.25–50 µM) [77].
6-methyl-1,2,3-trihydroxy-7,8-cyclohepta-9,12-diene-11-one-5,6,7,8-tetralene-7-acetamide (KL-4) (144) (Figure 7) was isolated from Aspergillus sp. obtained from the seeds of Gloriosa superba which were collected from Tirupati, India. KL-4 (144) exhibited good antifungal activity against Saccharomyces cerevisiae, C. albicans and Cryptococcus gastricus with MIC 25, 12.5, and 50 µg/mL respectively [78].
Endophytic fungus Penicillium sp. R22 associated with Nerium indicum collected from Qinling Mountain, Shaanxi Province, China was the source of 5-hydroxy-8-methoxy -4-phenylisoquinolin-1(2H)-one (145) a new isoquinolone alkaloid along with 3-O-methylviridicatin (146) and viridicatol (147) (Figure 7) two known quinolinone alkaloids. Compound 145 exhibited good antifungal activity against Alternaria brassicae, A. alternata and Valsa mali with MIC value of 31.2 µg/mL, compound 146 against A. brassicae, B. cinerea and Valsa male with MIC value of 31.2 µg/mL, compound 147 against A. brassicae, A. alternata and B. cinerea with MIC value of 31.2 µg/mL [79].
Trisulfide gliovirin-like compound Outovirin C (148) (Figure 7), an epithiodiketopiperazine natural product, was identified from Penicillium raciborskii, an endophytic fungus associated with Rhododendron tomentosum were collected at the test site of University of Oulu, Finland. Outovirin C (148) showed antifungal activity when assayed by micro-spectrophotometry using a dose response growth inhibition assay. Outovirin C inhibited the growth of Fusarium oxysporum, B. cinerea, and Verticillium dahlia at a low concentration of 0.38 mM (207 µg/mL) but a more significant growth inhibition was observed at the higher concentration of 0.76 mM (413 µg/mL). Compound 148 was most active against B. cinerea (57% inhibition) and slightly less effective against V. dahliae (45% inhibition) [80].
Fusaripeptide A (149) (Figure 7), a new cyclodepsipeptide, was isolated from the culture of the endophytic fungus Fusarium sp. associated with roots of Mentha longifolia growing in Saudi Arabia. Its structure was elucidated based on 1D and 2D NMR and HRESI and GC-MS experiments. The absolute configuration of the amino acid residues of 149 was assigned by chiral GC-MS and Marfey’s analysis after acid hydrolysis. Compound 149 exhibited potent antifungal activity toward C. albicans, C. glabrata, C. krusei, and A. fumigates with IC50 values of 0.11, 0.24, 0.19, and 0.14 µM, respectively. Under similar condition control amphotericin B exhibited antifungal activity toward C. albicans, C. glabrata, C. krusei, and A. fumigates with IC50 values of 0.3, 0.6, 0.5, 0.7 µM, respectively [81].
Fusarithioamide A, a new benzamide derivative (150) (Figure 7) was isolated from Fusarium chlamydosporium associated with the leaves of Anvillea garcinii collected from Al-Azhar University, Saudi Arabia. Compound 150 exhibited good antifungal activity against C. albicans with inhibition zone diameters (IZD 16.2 mm and MIC 2.6 µg/mL which is comparable to the positive control substance clotrimazole (IZD 18.5 mm and MIC 3.7 µg/mL) [82].
A new helvolic acid derivative named helvolic acid methyl ester (151), together with two known helvolic acid compounds, helvolic acid (152) and hydrohelvolic acid (153) (Figure 7), were extracted from endophytic fungus Fusarium sp. associated with Ficus carica leaves collected from Qinling Mountain, Shaanxi Province, China. Compounds 151–153 exhibited good antifungal activity against B. cinerea, C. gloeosporioides, F. oxysporum f. sp. niveum, Fusarium graminearum and Phytophthora capsici with MIC value in the range of 12.5–25 µg/mL while Carbendazim the standard showed MIC value in the range of 32.2–62.5 µg/mL against the same fungi [83].
Colletorin B (154), colletochlorin B (155) (Figure 7), LL-Z1272β (llicicolin B) (156), and 4,5-dihydrodechloroascochlorin (157) (Figure 8) were extracted from endophytic fungus Fusarium sp. Colletorin B (154) and colletochlorin B (155) showed moderate antifungal activity towards Ustilago violacea and F. oxysporum. Compound 156 showed moderate antifungal activity towards U. violacea and F. oxysporum. Furthermore, 4,5-dihydrodechloroascochlorin (157) showed a very strong antifungal activity towards Eurotium repens [84].
Murranolide A (158), murranopyrone (159), curvularin (160), (S)-dehydrocurvularin (161), pyrenolide A (162), modiolide A (163), and 8-hydroxy-6-methoxy-3-methylisocoumarin (164) (Figure 8) were extracted from the endophytic fungus Curvularia sp., strain M12, associated with the leaf of Murraya koenigii were collected from Rajshahi University, Bangladesh. Pyrenolide A (162) was observed to impair the mobility of Phytophthora capsici zoospores in a short time (30 min) at a low concentration (100% at 0.5 µg/mL). Murranolide A (158), murranopyrone (159), curvularin (160), (S)-dehydrocurvularin (161), modiolide A (163), and 8-hydroxy-6-methoxy-3-methylisocoumarin (164) exhibited zoospore motility impairment activity at higher concentrations (IC50: 50–100 µg/mL) [85].
Two new isoaigialones, B (165) and C (166) (Figure 8), along with aigialone (167) (Figure 8), were obtained from Phaeoacremonium sp., an endophytic fungus associated with the leaves of Senna spectabilis was collected in the Araraquara Cerrado area, in June 2001, Araraquara, Sao Paulo state, Brazil. These compounds were evaluated against Cladosporium cladosporioides and C. sphaerospermum using direct bioautography. Compounds 165 and 167 exhibited antifungal activity, with a detection limit of 5 µg, for both fungi, while compound 166 displayed weak activity (detection limit > 5 µg), with a detection limit of 25 µg. Nystatin was used as a positive control, showing a detection limit of 1 µg [86].
Trichothecinol A (168) (Figure 8) was obtained from Trichothecium sp. an endophytic fungus isolated from Phyllanthus amarus collected from Pune India. Compound 101 showed activity against Cryptococcus albidus (NCIM 3372) up to 20 µg/mL [87]. Trichothecin (169) (Figure 8), a sesquiterpene, was isolated from endophytic fungus Trichothecium sp. residing inside the leaves of Phyllanthus sp. collected from Pune India. Compound 169 exhibited anti-fungal activity against Saccharomyces cerevisiae, Cryptococcus albidus var diffluens NCIM 3371, Cryptococcus albidus var diffluens NCIM 3372, Fusarium oxysporum, Penicillium expansum, Trichoderma viride, Paecilomyces varioti and Aspergillus niger with MIC of 6, 20, 12, 10, 30, 40, 20 and 12 µg/mL, respectively [88].
2.4. Compounds Produced by Basidiomycetes
Two lanostane triterpenoids, sclerodols A (170) and B (171), and a known related lanostane triterpenoid (172) (Figure 8) were isolated from Scleroderma UFSM Sc1 (Persoon) Fries an endophyte associated with Eucalyptus grandis. Both compounds were evaluated for their anti-candidal potential against Candida albicans, C. tropicalis, C. crusei, C. parapsiosis for activities. Compound 171 showed good anticandidal activity against C. albicans, C. tropicalis, C. crusei, C. parapsiosis with the MIC of 25.0, 25.0, 6.25 and 12.5 and MFC of 25.0, 25.0, 12.5 and 25.0 µg/mL respectively. Compounds 170 and 172 were less active against tested strain than compound 171 with the MIC in the range of 12.5–100 and MLC (minimal lethal concentratin) of >100.0 µg/mL. Control nystatin exhibited showed anti-candidal activities against tested strains with the MIC in the range of 0.77–1.52 µg/mL and MLC in the range of 3.12–6.25 µg/mL [89].
3. Antifungal Potential of Volatile Organic Compounds (VOCs) from Endophytic Fungi
Volatile organic compounds (VOCs) are generally carbon compounds which exist in the gaseous phase at normal/ambient temperatures and pressures. Over 250 different VOCs produced by fungi comprising different chemical classes such as aldehydes, ketones, alcohols, phenols, thioesters, and so forth, have been identified in the context of the deterioration of fruits, vegetables, indoor environments (sick building syndrome); as chemotaxonomic markers; and in the morphogenesis and development of fungi.
However, bioprospecting fungal endophytes for the production of volatile antimicrobials came into the limelight with the discovery of Muscodor albus from the plant Cinnamomum zeylanicum, from Honduras. M. albus was found to produce an admixture of VOCs which could effectively kill a variety of pathogenic bacteria and fungi associated with plants and animals. This research garnered much attention and drove people to explore the volatile antibiotic properties of endophytic fungi for varied applications [90,91].
The genus Muscodor comprises of an endophytic fungi which is predominantly sterile, does not possess true reproductive structures like other fungi, and emanates a characteristic smell which is largely attribute to its VOC composition [92]. Since the report of M. albus in the late 1990s, to date, 20 species have been added to this genus, which have largely been identified based on their volatile signatures, molecular phylogeny, and morphological characteristics (Table 2). The characteristic VOC profile, therefore, is helpful in delineating the species, as well as playing a significant role in its anti-fungal and anti-bacterial properties. In this section, we only be highlight the anti-fungal potential of VOCs produced by these endophytic fungi.
The majority of the VOCs produced by the endophytic fungi comprises of a mixture of volatile components which generally has either a synergistic effect or an additive effect that enhances their bioactivity against pathogenic microbes. However, in a couple of studies, the major components of the volatile mixture were independently evaluated to understand their true antimicrobial/anti-fungal potential. These are generally synthetically generated and converted into a volatile form and subsequently evaluated for their bioactivity against the test microorganisms. For instance, Sclerotina sclerotiorum was completely inhibited by 2-methyl-1-butanol and 3-methyl-1-butanol with an EC50 value of 0.8 µL/mL. 2-methyl-1-butanol also inhibited Penicillium digitatum with an EC50 value of 0.48 µL/mL and B. cinerea with a value of 1.38 µL/mL. However, the volatile admixture of the M. albus VOC exhibited an IC50 range between 0.08 and 1.13 µL/mL, which clearly confirms the hypothesis of the synergistic/additive effects of the volatile components [93].
Recently, ethyl acetate has been reported to be the main VOC of yeasts Wickerhamomyces anomalus, Metschnikowia pulcherrima, and Saccharomyces cerevisiae, which inhibit the decay causing mold, as well as B. cineria. All three yeasts exhibit excellent biological control properties and were used for checking the mold and pathogenic attack in sweet cherries and strawberries. W. anomalus induced the highest killing activity amongst the three which was attributed to the higher production of Ethyl acetate. The role of the ethyl acetate was re-affirmed by using synthetic ethyl acetate from strawberry fruits to affirm the anti-fungal action [94].
Similarly, Phaeosphaeria nodorum, which existed as an endophyte in plum leaves (Prunus domestica) was found to inhibit the pathogen Monilinia fruticola. The major component of the VOC produced by Phaeosphoran odorum comprised of 3-methyl-1-butanol, acetic acid, 2-propyn-1-ol, and 2-propenenitril [95]. Similarly, six VOCs from the endophytic fungus Hypoxylon anthochroum (that is, phenylethyl alcohol), 2-methyl-butanol and 3-methyl-1-butanol, eucalyptol, ocimene, and terpenoline were tested against Fusarium oxysporum. The results indicated that these compounds exhibited concentration-dependent anti-fungal activity individually but have better action and control synergistically. Thus, the mixture of six VOCs may be used for the control of Fusarium oxysporum in tomatoes [96].
The genus Muscodor is one of the best studied endophytic fungus which produces a synergistic mixture of VOCs having lethal effects against a wide variety of plant and human pathogenic fungi, nematodes, and bacteria as well as certain insects [97,98,99,100]. The volatility of the Muscodor species has been used to replace methyl bromide (MeBr)—a traditional soil fumigant—which has been globally banned as it causes the depletion of ozone layer. Different species of Muscodor, their major VOCs, and their anti-fungal spectrum are given in Table 2. Geographically, each Muscodor species has a characteristic signature volatility. For instance, the Indian Muscodor species invariably has 4-Octadecylmorpholine as a marker compound while 2-methyl propanoic acid is generally found in Muscodor isolated from North and South America.
The majority of the VOCs from the fungal endophytic fungi are used as biological control agents to prevent the fungal deterioration of crops, fruits, and vegetable, under both pre- and post-harvest conditions. However, the exploitation of these fungally volatile organic compounds (FVOCs) from endophytic fungi are not being actively applied to humans for the prevention of fungal infections.
There exists a huge scope in evaluating these FVOCs from endophytic fungi since they could be helpful in curing superficial skin infections, the sanitization of public toilets, and in night soil. They can also find applications in personal care products such as for the aroma/fragrance in deodorants and sprays. They could presumably be helpful in the development of sprays for inhalation to treat fungal diseases like Aspergillosis in lungs.
4. Methods Used for Activation of Silent Biosynthetic Genes
Several research studies confirm that most of the biosynthetic gene clusters are observed to be silent or expressed at a low (minimal) level upon employing conventional culturing conditions for growth/propagation of microorganisms [113]. To activate such silent biosynthetic genes, numerous strategies have been employed, such as the one strain many compounds (OSMAC) approach (activation mediated through modification in composition of medium, aeration, temperature or shape of culturing flask), co-culturing method (facilitating activation through interspecies crosstalk) and genomics based approaches (expression of orphan biosynthesis genes in a heterologous host). In recent times, the use of chemicals as modifiers to alter the epigenetic makeup/constitution of a microorganism to improve its biosynthetic potential has become a beneficial tool. The method uses a chemical that acts as DNA methyltransferase inhibitors (DNMTi) or histone deacetylase inhibitors (HDACi), thereby stimulating the transcription previously silent gene clusters and fostering the production of a spectrum of natural products. A comprehensive description of some of these methods are given below/highlighted in the subsequent section.
4.1. Epigenetic Modification
Endophytes have proven to be the prolific source of bioactive metabolites and offer a substitute and untapped reserve for the discovery of novel metabolites. Studies have led to findings that tell biosynthetic gene clusters of microorganisms are mostly silent or expressed at very low levels under standard culture conditions and are least expressed, but under stress condition may it be biological, chemical or physical their expression takes place. Epigenetic modulators lead to the expression of these silent or cryptic genes. Epigenetic gene regulation is mediated by covalent histone modification, DNA methylation chromatin modeling basically induced by DNA methyl transferase inhibitors such as 5-aza-2-deoxycytidine, 5-azacytidine, hydralazine, procaine and histone deacetylase [114]. Chromatic modification in fungi to enhance gene transcription has led to secondary metabolite production of anthraquinones, cladochromes, lunalides, mycotoxins, and nygerones [115]. Structural genes that control transcriptional factor regulates the synthesis of secondary metabolites in fungi, these genes mediate factors occupied in environmental signals like pH, nitrogen and carbon sources, temperature, light, etc. [116]. In lab condition, these gene clusters are mostly silent. Under which natural conditions these clusters become activated is still unexplained. As per genetic sequencing studies carried so far, it is estimated that the clusters of genes responsible for secondary metabolites have not yet been deciphered completely [117].
From endophytic fungi Aspergillus fumigatus (GA-L7) obtained from Grewia asiatica led to identification of seven metabolites namely pseurotin A (173), pseurotin D (174), pseurotin F2 (175), fumagillin (176), tryprostatin C (177), gliotoxin (178) and bis(methylthio)gliotoxin (179) (Figure 9). On addition of the valproic acid, increase in the production of fumiquinazoline C (180) up to 10 times was noticed along with a shift in the pattern of metabolite production. It was also observed that all the genes, i.e., Afua_6g 12040, Afua_6g 12050, Afua_6g 12060, Afua_6g 12070 and Afua_6g 12080, tangled in the biosynthesis of fumiquinazoline C (180), were upregulated significantly by 7.5, 8.8, 3.4, 5.6 and 2.1 folds, respectively [118].
The NAD+-dependent HDAC inhibitor, nicotinamide, enhanced the yield of eupenicinicol C (181), and D (182) (Figure 9), decalin containing metabolites together with eujavanicol A (183), and eupenicinicol A (184) (Figure 9), biosynthetically related compounds by endophytic Eupenicillium sp. LG41, identified from the Xanthium sibiricum, a Chinese medicinal plant [119]. Under a similar condition without HDAC inhibitor, nicotinamide Eupenicillium sp. LG41 produce different decalin- containing compounds: eupenicinicols A (184), and B (185); two new sirenin derivatives, eupenicisirenins A (186) and B (187); and other four known compounds, (2S)-butylitaconic acid (188), (2S)-hexylitaconic acid (189) (Figure 9), xanthomegnin (190), and viridicatumtoxin (191) (Figure 10) [120].
Vasanthakumari et al. [121] reported the attenuation of camptothecin (192) (Figure 10) yield in endophytic fungi obtained from camptothecin fabricating plants, Nothapodytes nimmoniana and Miquelia dentata incorporated with 5-azacytidine, a DNA methyltransferase inhibitor. It was indicated that the mode behind decrease of camptothecin yield in endophytic fungi could in principle be inverted by stimulating some signals from the tissue of the plant, probably the methylation or silencing of the genes liable for camptothecin production.
Metabolites such as (10′S)-verruculide B (193), vermistatin (194), and dihydrovermistatin (195) (Figure 10) were identified owing to the supplementation of HDAC inhibitor, suberoylanilide hydroxamic acid (SAHA) to endophytic fungi culture Phoma sp. nov. LG0217 isolated from Parkinsonia microphylla. However, in absence of SAHA, a novel metabolite (S,Z)-5-(3′,4′-dihydroxybutyldiene)-3-propylfuran-2(5H)-one (196), along with nafuredin (197) (Figure 10), was produced [122].
Four new meroterpenoids identified as (4S)-4-decarboxylflavipesolide C (198), 1-(2,2-dimethylchroman-6-yl)-3-(4-hydroxyphenyl)propan-2-one (199), (R,E)-3-(2,2-dimethyl chroman6-yl)-4-hydroxy-5-((2-(2-hydroxypropan-2-yl)-2,3-dihydrobenzofuran-5-yl)methylene)furan2(5H)-one (200), methyl (R)-2-(2-(2-hydroxypropan-2-yl)-2,3-dihydrobenzofuran-5-yl) acetate (201), along with nine known compounds flavipesolides A−C (202–204), rubrolide S (205), 5-[(3,4-dihydro-2,2-dimethyl-2H-1-benzopyran-6-yl)-methyl]-3-hydroxy-4(4-hydroxyphenyl)-2(5H)-furanone (206), (3R,4R)-3,4-dihydro-4,8-dihydroxy-6,7-dimethoxy-3-methylisocoumarin (207), (3R)-3,4-dihydro-6,8-dimethoxy-3-methylisocoumarin (208) (Figure 10), terretonin C (209), and ergosterol (210) (Figure 11) were obtained using chemically modified epigenetic culture of Aspergillus terreus OUCMDZ-2739 with 10 µM trichostatin A (TSA). Under the similar parameters without TSA, A. terreus OUCMDZ-2739 yielded many compounds, i.e., aspernolide B (211), butyrolactone II (212), butyrolactone IV (213), butyrolactone I (214), aspernolide A (215), asterrelenin (216) and (+)-terrein (217) (Figure 11), supporting that fungal metabolite enrichment and chemodiversity using epigenetic modifiers can be done to obtain new products [123]. Induction of isosulochrin (218) (Figure 11) was also witnessed when Chaetomium sp. was supplemented with 5-azacytidine or SAHA on solid rice medium [124].
Asai et al. [125] reported six new benzophenones, cephalanones A–F (219–224), and 2-(2,6-dihydroxy-4-methylbenzoyl)-6-hydroxybenzoic acid (225) (Figure 11) from culture of Graphiopsis chlorocephala, from Paeonia lactiflora in presence of HDAC inhibitors nicotinamide (10 µM) which resulted in significant increase in secondary metabolite production.
To express silent biosynthetic pathways, molecules such as HDAC and DNMT are used to enhance the fungal metabolites production. Different studies suggest an increase in chemical diversity of metabolites by induction with these epigenetic modifiers. For growth and acclimatization with the environment fungus are known to produce diverse secondary metabolites. Cross talk between microbes and plant lead to the expression of these pathways which stays silent in in vitro conditions. Metabolic profiles shift led by SMs induced modifier is due to expression of cryptic genes [126].
4.2. The Co-Culture Strategy
Interspecific interaction among different species leads to evolution and biodiversity, organism combines their genetic information for better adaptability. The cohabitation of different microorganisms that share similar niches competes with growth, morphology, adaptation, and development patterns [127,128]. The increased productions of metabolites in co-culture which are not produced in axenic culture are the result of competition or antagonism faced by the microorganism that leads to activation of cryptic genes. [129]. Co-cultivation is a way to provide natural habitat to fungi so that gene clusters become activated. In Aspergillus nidulans, the cryptic gene has been successfully activated leading to isolation of novel compounds [130].
In a study carried by Ola et al. [131], accumulation of secondary metabolites, i.e., lateropyrone (226), cyclic depsipeptides of the enniatin type (227–229), and the lipopeptide fusaristatin A (230) (Figure 12), was found to be enhanced by 78 folds by co-culturing B. subtilis 168 trpC2 with Fusarium tricinctum isolated from the Aristolochia paucinervis. This led to the identification of three new compounds macrocarpon C (231), 2-(carboxymethylamino)benzoic acid (232) and (−)-citreoisocoumarinol) (233), and a known compound, (−)-citreoisocoumarin (234), which was absent in axenic culture of bacterial or fungal control. On coculturing Alternaria sp. and Phomopsis sp. there was the enhancement of taxane (235) production by eight fold [132].
Aspergillus austroafricanus endophyte residing inside the leaves of Eichhornia crassipes was the source of two new metabolites, namely, xanthone dimer austradixanthone (236) and sesquiterpene (+)-austrosene (237), and five known compounds, (+)-sydowic acid (238), sydowinin B (239), oxaline (240), 4-hydroxymethyl-5-hydroxy-2H-pyran-2- one (241) (Figure 12), ergosterol (210) (Figure 13). However, the same endophyte grown in mixed cultures with Bacillus subtilis or Streptomyces lividans led to the identification of many diphenyl ethers i.e., violaceol I (242), violaceol II (243), and diorcinol (244) (Figure 12) along with new austramide (245) (Figure 12), increased up to 29 times [133].
Chaetomium sp. was isolated from Sapium ellipticum the Cameroonian medicinal plant. When Chaetomium sp. was cultured axenically on solid rice medium, average yields per culture flask were 2.8, 13.9, 132.7 and 14.6 mg of acremonisol A (246), SB236050 (247) (Figure 12), and SB238569 (248), respectively, and 1:1 mixture of 3- and 4-hydroxybenzoic acid methyl esters (249–250), respectively, (Figure 13) was observed. When Co-cultivation of Chaetomium sp. was undertaken with viable or autoclaved cultures of Bacillus subtilis there was a strong accumulation of the 1:1 mixture of (249), and (250), was observed, accounting for an 8.3 and 7.4-fold increase, respectively, compared to axenic fungal controls in both cases. SB236050 (247) and SB238569 (248), two major polyketides of Chaetomium sp., were not detected in co-cultures. Five new compounds, Shikimeran A (251), Bipherin A (252), Chorismeron (253), Quinomeran (254), and Serkydayn (255), and two known compounds, isosulochrin (218) and protocatechuic acid methyl ester (256) (Figure 12), were only detected in co-cultures of Chaetomium sp. with viable or autoclaved B. subtilis cultures, but were lacking in both fungal or bacterial controls when cultured axenically [124].
These studies indicate that co-culture generates a complex and promising environment to obtain new secondary metabolites as a response to the interaction between endophytic fungi. The above also indicates that the production of new natural products depends on stimuli.
5. Conclusions
Endophytic fungi are the ubiquitous source of novel chemical compounds having the potential to display antifungal activities. Interestingly, the active metabolites from endophytic fungi possess excellent antifungal activity not only against human fungal pathogens but also on plant fungal pathogens. In addition, the volatile organic compounds (VOCs) from genus Muscodor displayed significant antifungal as well as antibacterial properties and, therefore, they are used to prevent fungal deterioration of crops, fruits and vegetables. However, their application to control human fungal infection has not been explored. Fungal VOCs can be investigated for the development of sprays for inhalation to treat fungal diseases such as Aspergillosis in lungs, curing superficial skin infections and sanitization. Endophytic fungi are being studied to produce natural compounds which are originally produced from their host plants and, thus, emerging as an alternative and sustainable source of valuable natural products. It is important to investigate the interactions between endophytic fungi with the host plant and other endophytes which are very sensitive to the culture conditions and hence, provide an opportunity to tune the in vitro culture conditions to produce the desired range of secondary metabolites. It is possible to produce a compound of interest by varying the culture conditions such as media composition, aeration rate and temperature. In addition, cultivation of endophytic fungi in presence of bacteria or other fungi (co-cultivation) yield novel compounds which otherwise do not appear when fungi or bacteria are cultivated alone. Therefore, considerable research on endophytic fungi is required for the development of suitable co-culture system for the sustained production of the desired secondary metabolite.
Author Contributions
S.K.D., V.P., S.S. and M.K.G. reviewed the contents critically. V.P. and M.K.G. drew chemical structures and assisted in the preparation of Table 1. S.S. wrote the Antifungal potential of Volatile organic compounds of review. The manuscript has been read and approved by all named authors.
Funding
This research received no external funding.
Acknowledgments
The authors are thankful to Alok Adholeya, Senior Director, Sustainable Agriculture Division, The Energy and Resources Institute (India) for continuous support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hawksworth, D.L. The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C.; Draeger, S.; Römmert, A.K.; Krohn, K. Endophytic fungi: A source of novel biologically active secondary metabolites. Mycol. Res. 2002, 106, 996–1004. [Google Scholar] [CrossRef]
- Nicoletti, R.; Fiorentino, A. Plant bioactive metabolites and drugs produced by endophytic fungi of Spermatophyta. Agriculture 2015, 5, 918–970. [Google Scholar] [CrossRef]
- Strobel, G.; Daisy, B. Bioprospecting for microbial endophytes and their natural products. Microbiol. Mol. Biol. Rev. 2003, 67, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Gunatilaka, A.A.L. Natural products from plant-associated microorganisms: Distribution, structural diversity, bioactivity, and implications of their occurrence. J. Nat. Prod. 2006, 69, 509–526. [Google Scholar] [CrossRef] [PubMed]
- Kharwar, R.N.; Mishra, A.; Gond, S.K.; Stierle, A.; Stierle, D. Anticancer compounds derived from fungal endophytes: Their importance and future challenges. Nat. Prod. Rep. 2011, 28, 1208–1228. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, S.K.; Verekar, S.A. Fungal endophytes: A potential source of antifungal compounds. Front. Biosci. 2012, E4, 2045–2070. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Verekar, S.A.; Bhave, S.V. Endophytic fungi: A reservoir of antibacterials. Front. Microbiol. 2015, 5, 715. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Li, G.; Lou, H.X. Structural Diversity and Biological Activities of Novel Secondary metabolites from Endophytes. Molecules 2018, 23, 646. [Google Scholar] [CrossRef] [PubMed]
- Carroll, G. Fungal endophytes in stems and leaves: From latent pathogens to mutualistic symbionts. Ecology 1988, 69, 2–9. [Google Scholar] [CrossRef]
- Hallmann, J.; Sikora, R. Toxicity of fungal endophytic secondary metabolites to plant parasitic nematodes and soil borne plant pathogenic fungi. Eur. J. Plant Pathol. 1996, 102, 155–162. [Google Scholar] [CrossRef]
- Sturz, A.V.; Nowak, J. An endophytic community of rhizobacteria and the strategies requires to create yield enhancing associations with crops. Appl. Soil Ecol. 2000, 15, 183–190. [Google Scholar] [CrossRef]
- Azevedo, J.L.; Araujo, W.L. Diversity and applications of endophytic fungi isolated from tropical plants. In Fungi Multifaceted Microbes; Ganguli, B.N., Deshmukh, S.K., Eds.; Anamaya: New Delhi, India, 2007; pp. 189–207. [Google Scholar]
- Deshmukh, S.K.; Prakash, V.; Ranjan, N. Recent advances in the discovery of bioactive metabolites from Pestalotiopsis. Phytochem. Rev. 2017, 16, 883–920. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, B.Y.; Yang, X.L. Antifungal monoterpene derivatives from the plant endophytic fungus Pestalotiopsis foedan. Chem. Biodivers. 2016, 13, 1422–1425. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.S.; Jia, M.; Chen, L.; Zhu, B.; Dong, H.X.; Si, J.P.; Peng, W.; Han, T. Cytotoxic and antifungal constituents isolated from the metabolites of endophytic fungus DO14 from Dendrobium officinale. Molecules 2016, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, X.; Guo, L.; Che, Y.; Liu, L. 2H-Pyran-2-one and2H-Furan-2-one derivatives from the plant endophytic fungus Pestalotiopsis fici. Chem. Biodivers. 2013, 10, 2007–2013. [Google Scholar] [CrossRef] [PubMed]
- Subban, K.; Subramani, R.; Muthumary, J. A novel antibacterial and antifungal phenolic compound from the endophytic fungus Pestalotiopsis mangiferae. Nat. Prod. Res. 2013, 27, 1445–1449. [Google Scholar] [CrossRef] [PubMed]
- Chapla, V.M.; Zeraik, M.L.; Ximenes, V.F.; Zanardi, L.M.; Lopes, M.N.; Cavalheiro, A.J.; Silva, D.H.S.; Young, M.C.M.; da Fonseca, L.M.; Bolzani, V.S.; et al. Bioactive secondary metabolites from Phomopsis sp., an endophytic fungus from Senna spectabilis. Molecules 2014, 19, 6597–6608. [Google Scholar]
- Wu, S.H.; Huang, R.; Miao, C.P.; Chen, Y.W. Two new steroids from an endophytic fungus Phomopsis sp. Chem. Biodivers. 2013, 10, 1276–1283. [Google Scholar] [CrossRef] [PubMed]
- Tanney, J.B.; McMullin, D.R.; Green, B.D.; Miller, J.D.; Seifert, K.A. Production of antifungal and anti-insectan metabolites by the Picea endophyte Diaporthe maritima sp. nov. Fungal Biol. 2016, 120, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.; Deshmukh, P.; Gade, A.; Ingle, A.; Kövics, G.J.; Irinyi, L. Phoma Saccardo: Distribution, secondary metabolite production and biotechnological applications. Crit. Rev. Microbiol. 2009, 35, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.; Kock, I.; Al-Harras, A.; Al-Rawahi, A.; Abbas, G.; Green, I.R.; Shah, A.; Badshah, A.; Saleem, M.; Draeger, S.; et al. Antimicrobial chemical constituents from endophytic fungus Phoma sp. Asian. Pac. J. Trop. Med. 2014, 7, 699–702. [Google Scholar] [CrossRef]
- Mousa, W.K.; Schwan, A.; Davidson, J.; Auzanneau, F.I.; Strange, P.; Liu, H.; Zhou, T.; Raizada, M.N. An endophytic fungus isolated from finger millet (Eleusine coracana) produces anti-fungal natural products. Front. Microbiol. 2015, 6, 1157. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.; Wang, A.; Cao, Y.; Zhou, K.; Mao, Z.; Dong, X.; Tian, J.; Xu, D.; Dai, J.; Peng, Y.; et al. Bioactivedibenzo-α-pyrone derivatives from the endophytic fungus Rhizopycnis vagum Nitaf22. J. Nat. Prod. 2016, 79, 2022–2031. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.; Root, N.; Jabeen, F.; Al-Harrasi, A.; Ahmad, M.; Mabood, F.; Hassan, Z.; Shah, A.; Green, I.R.; Schulz, B.; et al. Microsphaerol and seimatorone: Two new compounds isolated from the endophytic fungi, Microsphaeropsis sp. and Seimatosporium sp. Chem. Biodivers. 2015, 12, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Chapla, V.M.; Zeraik, M.L.; Leptokarydis, I.H.; Silva, G.H.; Bolzani, V.S.; Young, M.C.M.; Pfenning, L.H.; Araujo, A.R. Antifungal compounds produced by Colletotrichum gloeosporioides, an endophytic fungus from Michelia champaca. Molecules 2014, 19, 19243–19252. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.; Root, N.; Jabeen, F.; Al-Harras, A.; Al-Rawahi, A.; Ahmad, M.; Hassan, Z.; Abba, G.; Mabood, F.; Shah, A.; et al. Seimatoric acid and colletonoic acid: Two new compounds from the endophytic fungi, Seimatosporium sp. and Colletotrichum sp. Chin. Chem. Lett. 2014, 25, 1577–1579. [Google Scholar] [CrossRef]
- Sun, P.; Huo, J.; Kurtan, T.; Mandi, A.; Antus, S.; Tang, H.; Draeger, S.; Schulz, B.; Hussain, H.; Krohn, K.; et al. Structural and stereochemical studies of hydroxyanthraquinone derivatives from the endophytic fungus Coniothyrium sp. Chirality 2013, 25, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Macias-Rubalcava, M.L.; Sanchez-Fernandez, R.E. Secondary metabolites of endophytic Xylaria species with potential applications in medicine and agriculture. World J. Microbiol. Biotechnol. 2017, 33, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Wu, S.H.; Zhai, Y.Z.; Xuan, Q.C.; Wang, T. Secondary metabolites from the genus Xylaria and their bioactivities. Chem. Biodivers. 2014, 11, 673–694. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; He, J.; Li, X.N.; Huang, R.; Song, F.; Chen, Y.W.; Miao, C.P. Guaianesesquiterpenes and isopimaranediterpenes from an endophytic fungus Xylaria sp. Phytochemistry 2014, 105, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Xie, X.S.; Fang, X.W.; Ma, K.X.; Wu, S.H. Five new guaianesesquiterpenes from the endophytic fungus Xylaria sp. YM 311647 of Azadirachta indica. Chem. Biodivers. 2015, 12, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Baraban, E.G.; Morin, J.B.; Phillips, G.M.; Phillips, A.J.; Strobel, S.A.; Handelsman, J. Xyolide, a bioactive nonenolide from an Amazonian endophytic fungus, Xylaria feejeensis. Tetrahedron Lett. 2013, 54, 4058–4060. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xiao, J.; Sun, Q.Q.; Qin, J.C.; Pescitelli, G.; Gao, J.M. Characterization of cytochalasins from the endophytic Xylaria sp. and their biological functions. J. Agric. Food. Chem. 2014, 62, 10962–10969. [Google Scholar] [CrossRef] [PubMed]
- Rukachaisirikul, V.; Buadam, S.; Sukpondma, Y.; Phongpaichit, S.; Sakayaroj, J.; Hutadilok-Towatana, N. Indanone and mellein derivatives from the Garcinia-derived fungus Xylaria sp. PSU-G12. Phytochem. Lett. 2013, 6, 135–138. [Google Scholar] [CrossRef]
- Sica, V.P.; Rees, E.R.; Tchegnon, E.; Bardsley, R.H.; Raja, H.A.; Oberlies, N.H. Spatial and temporal profiling of griseofulvin production in Xylaria cubensis using mass spectrometry mapping. Front. Microbiol. 2016, 7, 544. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.N.; Walker, A.K.; Nsiama, T.K.; McFarlane, J.; Sumarah, M.W.; Ibrahim, A.; Miller, J.D. Griseofulvin producing Xylaria endophytes of Pinus strobus and Vaccinium angustifolium: Evidence for a conifer-understory species endophyte ecology. Fungal Ecol. 2014, 11, 107–113. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, H.Q.; Zong, S.C.; Gao, J.M.; Zhang, A.L. Chemical and bioactive diversities of the genus Chaetomium secondary metabolites. Mini-Rev. Med. Chem. 2012, 12, 127–148. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Yan, X.; Fang, M.J.; Wu, Z.; Wang, D.; Qiu, Y.K. Two new cytochalasan derivatives from Chaetomium globosum SNSHI-5, a fungus derived from extreme environment. Nat. Prod. Res. 2017, 31, 1669–1675. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.S.; Zhang, Y.Y.; Yan, W.; Cao, L.L.; Xiao, Y.; Ye, Y.H.; Zhao, S.S.; Zhang, Y.Y.; Yan, W.; Cao, L.L.; et al. Chaetomium globosum CDW7, a potential biological control strain and its antifungal metabolites. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, X.; Yang, Y.; Duang, R.; Chen, G.; Li, X.; Li, Q.; Qin, S.; Li, S.; Zhao, L.; et al. Anti-phytopathogen, multi-target acetylcholinesterase inhibitory and antioxidant activities of metabolites from endophytic Chaetomium globosum. Nat. Prod. Res. 2016, 30, 2616–2619. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Y.Y.; Ding, D.D.; Yu, S.P.; Wang, L.W. A study on the secondary metabolites of endophytic fungus Chaetomium cupreum ZJWCF079 in Macleaya cordata. Health Res. 2013, 33, 94–96. [Google Scholar]
- Zhang, G.; Zhang, Y.; Qin, J.; Qu, X.; Liu, J.; Li, X.; Pan, H. Antifungal metabolites produced by Chaetomiumglobosum No.04, an endophytic fungus isolated from Ginkgo biloba. Indian J. Microbiol. 2013, 53, 175–180. [Google Scholar] [PubMed]
- Chen, Y.M.; Yang, Y.H.; Li, X.N.; Zou, C.; Zhao, P.J. Diterpenoids from the endophytic fungus Botryosphaeria sp. P483 of the Chinese herbal medicine Huperzia serrata. Molecules 2015, 20, 16924–16932. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Zhang, Q.; Gao, Y.Q.; Tang, J.J.; Zhang, A.L.; Gao, J.M. Secondary metabolites from the endophytic Botryosphaeria dothidea of Melia azedarach and their antifungal, antibacterial, antioxidant, and cytotoxic activities. J. Agric. Food Chem. 2014, 62, 3584–3590. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.B.; de Oliveira, D.M.; Hughes, A.F.S.; Kohlhoff, M.; Vieira, M.L.A.; Martins Vaz, A.B.; Ferreira, M.C.; Carvalho, C.R.; Rosa, L.H.; Rosa, C.A.; et al. Endophytic fungal compounds active against Cryptococcus neoformans and C. gattii. J. Antibiot. 2015, 68, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; de Sá, N.P.; Borelli, B.M.; Rosa, C.A.; Barbeira, P.J.S.; Cota, B.B.; Johann, S. Antifungal activity of eicosanoic acids isolated from the endophytic fungus Mycosphaerella sp. against Cryptococcus neoformans and C. gattii. Microb. Pathog. 2016, 100, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Li, T.X.; Yang, M.H.; Wang, X.B.; Wang, Y.; Kong, L.Y. Synergistic antifungal meroterpenes and dioxolanone derivatives from the endophytic fungus Guignardia sp. J. Nat. Prod. 2015, 78, 2511–2520. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Lai, D.; Liu, X.; Fu, X.; Meng, J.; Wang, A.; Wang, X.; Sun, W.; Liu, Z.L.; Zhou, L.; et al. Dibenzo-α-pyrones: A new class of larvicidal metabolites against Aedes aegypti from the endophytic fungus Hyalodendriella sp. Ponipodef12. Pest Manag. Sci. 2017, 73, 1478–1485. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Mao, Z.; Lou, J.; Xu, L.; Zhong, L.; Peng, Y.; Zhou, L.; Wang, M. Benzopyranones from the endophytic fungus Hyalodendriella sp. Ponipodef12 and their bioactivities. Molecules 2012, 17, 11303–11314. [Google Scholar] [CrossRef] [PubMed]
- Rosseto, P.; Costa, A.T.; Polonio, J.C.; da Silva, A.A.; Pamphile, J.A.; Azevedo, J.L. Investigation of mycoviruses in endophytic and phytopathogenic strains of Colletotrichum from different hosts. Genet. Mol. Res. 2016, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, G.; Zhang, Y.; Zheng, B.; Zhang, C.; Wang, L. Isolation and identification of an endophytic fungus Pezicula sp. in Forsythia viridissima and its secondary metabolites. World. J. Microbiol. Biotechnol. 2014, 30, 2639–2644. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.L.; Zhang, Y.Y.; Liu, Y.J.; Yang, T.T.; Zhang, J.L.; Zhang, Z.G.; Shen, L.; Liu, J.Y.; Ye, Y.H. Anti-phytopathogenic activity of sporothriolide, a metabolite from endophyte Nodulisporium sp. A21 in Ginkgo biloba. Pestic. Biochem. Physiol. 2016, 129, 7–13. [Google Scholar] [CrossRef] [PubMed]
- McMullin, D.R.; Green, B.D.; Miller, J.D. Antifungal sesquiterpenoids and macrolides from an endophytic Lophodermium species of Pinus strobus. Phytochem. Lett. 2015, 14, 148–152. [Google Scholar] [CrossRef]
- Li, R.; Chen, S.; Niu, S.; Guo, L.; Yin, J.; Che, Y. Exserolides A-F, new isocoumarin derivatives from the plant endophytic fungus Exserohilum sp. Fitoterapia 2014, 96, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.R.; Wedge, D.E.; Cantrell, C.L.; Silva-Hughes, A.F.; Pan, Z.; Moraes, R.M.; Madoxx, V.L.; Rosa, L.H. Molecular phylogeny, diversity, and bioprospecting of endophytic fungi associated with wild ethnomedicinal North American plant Echinacea purpurea (Asteraceae). Chem. Biodivers. 2016, 13, 918–930. [Google Scholar] [CrossRef] [PubMed]
- Burgess, K.M.N.; Ibrahim, A.; Soerensen, D.; Sumarah, M.W. Trienylfuranol A and trienylfuranone A.B: Metabolites isolated from an endophytic fungus, Hypoxylon submonticulosum, in the raspberry Rubus idaeus. J. Antibiot. 2017, 70, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Nalli, Y.; Mirza, D.N.; Wani, Z.A.; Wadhwa, B.; Mallik, F.A.; Raina, C.; Chaubey, A.; Riyaz-Ul-Hassan, S.; Ali, A. Phialomustin A-D, new antimicrobial and cytotoxic metabolites from an endophytic fungus, Phialophora mustea. RSC Adv. 2015, 5, 95307–95312. [Google Scholar] [CrossRef]
- Hussain, H.; Jabeen, F.; Krohn, K.; Al-Harras, A.; Ahmad, M.; Mabood, F.; Shah, A.; Badshah, A.; Rehman, N.U.; Green, I.R.; et al. Antimicrobial activity of two mellein derivatives isolated from an endophytic fungus. Med. Chem. Res. 2015, 24, 2111–2114. [Google Scholar] [CrossRef]
- Hussain, H.; Kliche-Spory, C.; Al-Harrasi, A.; Al-Rawahi, A.; Abbas, G.; Green, I.R.; Schulz, B.; Krohn, K.; Shah, A. Antimicrobial constituents from three endophyticfungi. Asian Pac. J. Trop. Med. 2014, 7, S224–S227. [Google Scholar] [CrossRef]
- Shan, T.; Tian, J.; Wang, X.; Mou, Y.; Mao, Z.; Lai, D.; Dai, J.; Peng, Y.; Zhou, L.; Wang, M. Bioactive spirobisnaphthalenes from the endophytic fungus Berkleasmium sp. J. Nat. Prod. 2014, 77, 2151–2160. [Google Scholar] [CrossRef] [PubMed]
- Siriwach, R.; Kinoshita, H.; Kitani, S.; Igarashi, Y.; Pansuksan, K.; Panbangred, W.; Nihira, T. Bipolamides A and B, triene amides isolated from the endophytic fungus Bipolaris sp. MU34. J. Antibiot. 2014, 67, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Phaopongthai, J.; Wiyakrutta, S.; Meksuriyen, D.; Sriubolmas, N.; Suwanborirux, K. Azole-synergistic anti-candidal activity of altenusin, a biphenyl metabolite of the endophytic fungus Alternaria alternata isolated from Terminalia chebula Retz. J. Microbiol. 2013, 51, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Johann, S.; Rosa, L.H.; Rosa, C.A.; Perez, P.; Cisalpino, P.S.; Zani, C.L.; Cota, B.B. Antifungal activity of altenusin isolated from the endophytic fungus Alternaria sp. against the pathogenic fungus Paracoccidioides brasiliensis. Rev. Iberoam. Micol. 2012, 29, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Radwan, M.M.; Tarawneh, A.H.; Gao, J.; Wedge, D.E.; Rosa, L.H.; Cutler, H.G.; Cutler, S.J. Antifungal activity against plant pathogens of metabolites from the endophytic fungus Cladosporium cladosporioides. J. Agric. Food Chem. 2013, 61, 4551–4555. [Google Scholar] [CrossRef] [PubMed]
- Talontsi, F.M.; Dittrich, B.; Schueffler, A.; Sun, H.; Laatsch, H. Epicoccolides: Antimicrobial and antifungal polyketides from an endophytic fungus Epicoccum sp. associated with Theobroma cacao. Eur. J. Org. Chem. 2013, 2013, 3174–3180. [Google Scholar] [CrossRef]
- Silva-Hughes, A.F.; Carvalho, C.R.; Wedge, D.E.; Cantrell, C.L.; Pan, Z.; Moraes, R.M.; Madoxx, V.L.; Rosa, L.H. Diversity and antifungal activity of the endophytic fungi associated with the native medicinal cactus Opuntia humifusa (Cactaceae) from the United States. Microbiol. Res. 2015, 175, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Pang, X.J.; Xu, L.L.; Zhao, T.; Long, X.Y.; Zhang, Q.Y.; Qin, H.L.; Yang, D.F.; Yang, X.L. Two new alkylated furan derivatives with antifungal and antibacterial activities from the plant endophytic fungus Emericella sp. XL029. Nat. Prod. Res. 2017, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mousa, W.K.; Schwan, A.L.; Raizada, M.N. Characterization of antifungal natural products isolated from endophytic fungi of finger millet (Eleusine coracana). Molecules 2016, 21, 1171. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Yang, Y.B.; Chen, J.L.; Miao, C.P.; Wang, Q.; Zhou, H.; Chen, Y.W.; Li, Y.Q.; Ding, Z.T.; Zhao, L.X. Koninginins N-Q, polyketides from the endophytic fungus Trichoderma koningiopsis Harbored in Panax notoginseng. Nat. Prod. Bioprospect. 2016, 6, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Yang, Y.; Miao, C.P.; Zheng, Y.K.; Chen, J.L.; Chen, Y.W.; Xu, L.H.; Guang, H.L.; Ding, Z.T.; Zhao, L.X. Koningiopisins A-H, polyketides with synergistic antifungal activities from the endophytic fungus Trichoderma koningiopsis. Planta Medica 2016, 82, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Xu, J.; Li, F.; Xu, L.; Li, C. A New antifungal isocoumarin from the endophytic fungus Trichoderma sp. 09 of Myoporum bontioides A. Gray. Pharmacogn. Mag. 2016, 12, 259–261. [Google Scholar] [PubMed]
- Shentu, X.; Zhan, X.; Ma, Z.; Yu, X.; Zhang, C. Antifungal activity of metabolites of the endophytic fungus Trichoderma brevicompactum from garlic. Braz. J. Microbiol. 2014, 45, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Li, Q.L.; Yang, Y.B.; Liu, K.; Miao, C.P.; Zhao, L.X.; Ding, Z.T. Koninginins R-S from the endophytic fungus Trichoderma koningiopsis. Nat. Prod. Res. 2017, 31, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Elkhayat, E.S.; Mohamed, G.A.; Khedr, A.I.M.; Fouad, M.A.; Kotb, M.H.R.; Ross, S.A. Aspernolides F and G, new butyrolactones from the endophytic fungus Aspergillus terreus. Phytochem. Lett. 2015, 14, 84–90. [Google Scholar] [CrossRef]
- Xiao, J.; Zhang, Q.; Gao, Y.Q.; Shi, X.W.; Gao, J.M. Antifungal and antibacterial metabolites from an endophytic Aspergillus sp. associated with Melia azedarach. Nat. Prod. Res. 2014, 28, 1388–1392. [Google Scholar] [PubMed]
- Budhiraja, A.; Nepali, K.; Sapra, S.; Gupta, S.; Kumar, S.; Dhar, K.L. Bioactive metabolites from an endophytic fungus of Aspergillus species isolated from seeds of Gloriosa superba Linn. Med. Chem. Res. 2013, 22, 323–329. [Google Scholar] [CrossRef]
- Ma, Y.M.; Qiao, K.; Kong, Y.; Li, M.Y.; Guo, L.X.; Miao, Z.; Fan, C. A new isoquinolone alkaloid from an endophytic fungus R22 of Nerium indicum. Nat. Prod. Res. 2017, 31, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Kajula, M.; Ward, J.M.; Turpeinen, A.; Tejesvi, M.V.; Hokkanen, J.; Tolonen, A.; Hakkanen, H.; Picart, P.; Ihalainen, J.; Sahl, H.G.; et al. Bridged epipolythiodiketopiperazines from Penicillium raciborskii, an endophytic fungus of Rhododendron tomentosum Harmaja. J. Nat. Prod. 2016, 79, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Abdallah, H.M.; Elkhayat, E.S.; Al Musayeib, N.M.; Asfour, H.Z.; Zayed, M.F.; Mohamed, G.A. Fusaripeptide A: New antifungal and anti-malarial cyclodepsipeptide from the endophytic fungus Fusarium sp. J. Asian Nat. Prod. Res. 2018, 20, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Elkhayat, E.S.; Mohamed, G.A.A.; Fat’hi, S.M.; Ross, S.A. Fusarithioamide A, a new antimicrobial and cytotoxic benzamide derivative from the endophytic fungus Fusarium chlamydosporium. Biochem. Biophys. Res. Commun. 2016, 479, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.A.; Ma, Y.M.; Zhang, H.C.; Liu, R.A. new helvolic acid derivative from an endophytic Fusarium sp. of Ficus carica. Nat. Prod. Res. 2016, 30, 2407–2412. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.; Drogies, K.H.; Al-Harrasi, A.; Hassan, Z.; Shah, A.; Ran, U.A.; Green, I.R.; Draeger, S.; Schulz, B.; Krohn, K. Antimicrobial constituents from endophytic fungus Fusarium sp. Asian Pac. J. Trop. Dis. 2015, 5, 186–189. [Google Scholar] [CrossRef]
- Mondol, M.A.M.; Farthouse, J.; Islam, M.T.; Schueffler, A.; Laatsch, H. Metabolites from the endophytic fungus Curvularia sp. M12 act as motility inhibitors against Phytophthora capsici Zoospores. J. Nat. Prod. 2017, 80, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.H.; Zeraik, M.L.; de Oliveira, C.M.; Teles, H.L.; Trevisan, H.C.; Pfenning, L.H.; Nicolli, C.P.; Young, M.C.M.; Mascarenhas, Y.P.; Abreu, L.M.; et al. Lactone derivatives produced by a Phaeoacremonium sp., an endophytic fungus from Senna spectabilis. J. Nat. Prod. 2017, 80, 1674–1678. [Google Scholar] [CrossRef] [PubMed]
- Taware, R.; Abnave, P.; Patil, D.; Rajamohananan, P.R.; Raja, R.; Soundararajan, G.; Kundu, G.C.; Ahmad, A. Isolation, purification and characterization of Trichothecinol-A produced by endophytic fungus Trichothecium sp. and its antifungal, anticancer and antimetastatic activities. Sustain. Chem. Process. 2014, 2, 8. [Google Scholar] [CrossRef]
- Taware, R.; Abnave, P.; Patil, D.; Rajamohananan, P.R.; Raja, R.; Soundararajan, G.; Kundu, G.C.; Kharat, M.K.D.; Pai, K.; Ahmad, A. Trichothecin from endophytic fungus Trichothecium sp. and its anticancer effect on murine melanoma and breast cancer cell lines. Curr. Biochem. Eng. 2015, 2, 73–80. [Google Scholar] [CrossRef]
- Morandini, L.M.B.; Neto, A.T.; Pedroso, M.; Antoniolli, Z.I.; Burrow, R.A.; Bortoluzzi, A.J.; Mostardeiro, M.A.; da Silva, U.F.; Dalcol, I.I.; Morel, A.F. Lanostane-type triterpenes from the fungal endophyte Scleroderma UFSMSc1 (Persoon) Fries. Bioorg. Med. Chem. Lett. 2016, 26, 1173–1176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.L.; Wang, G.P.; Mao, L.J.; Komon-Zelazowska, M.; Yuan, Z.L.; Lin, F.C.; Druzhinina, I.S.; Kubicek, C.P. Muscodor fengyangensis sp. nov. from southeast China: Morphology, physiology and production of volatile compounds. Fungal Biol. 2010, 114, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Woropong, J.; Strobel, G.A.; Ford, E.J.; Li, J.Y.; Baird, G.; Hess, W.M. Muscodor albusanam. nov., an endophyte from Cinnamomum zeylanicum. Mycotaxon 2001, 79, 67–79. [Google Scholar]
- Strobel, G.A. Muscodor species-endophytes with biological promise. Phytochem. Rev. 2011, 10, 165–172. [Google Scholar] [CrossRef]
- DiFranscesco, A.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of volatile organic compounds by Aureobasidium pullulans as a potential mechanism against post-harvest fruit pathogens. Biol. Control 2015, 81, 8–14. [Google Scholar] [CrossRef]
- Oro, L.; Feliziani, E.; Ciani, M.; Romanazzi, G.; Comitini, F. Volatile organic compounds of Wickerhamomyces anomalus, Metschinikowia pulcherrima and Saccharomyces cerevisiae inhibit growth of decay causing fungi and post-harvest disease of strawberries. J. Food Microbiol. 2018, 265, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, R.S.; da Silva, J.F.M.; Buyer, J.S.; Janiesiewicz, W.J. Endophytic fungi from Plums (Prunis domestica) and their antifungal activity against Monilinia fruticola. J. Food Prot. 2012, 75, 1883–1889. [Google Scholar] [CrossRef] [PubMed]
- Medina-Romaro, Y.M.; Roque-Flores, G.; Macias-Rubalcava, M.L. Volatile organic compounds from endophytic fungi as innovative post-harvest control of Fusarium oxysporum in cherry tomato fruits. Appl. Microbiol. Biotechnol. 2017, 101, 8209–8222. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.A.; Dirkse, E.; Sears, J.; Markworth, C. Volatile antimicrobials from Muscodor albus, a novel endophytic fungus. Microbiology 2001, 147, 2943–2950. [Google Scholar] [CrossRef] [PubMed]
- Daisy, B.H.; Strobel, G.A.; Castillo, U.; Ezra, D.; Sears, J.; Weaver, D.K.; Runyon, J.B. Naphthalene, an insect repellent, is produced by Muscodor vitigenus, a novel endophytic fungus. Microbiology 2002, 148, 3737–3741. [Google Scholar] [CrossRef] [PubMed]
- Grimme, E.; Zidack, N.K.; Sikora, R.A.; Strobel, G.A.; Jacobsen, B.J. Comparison of Muscodor albus volatiles with abiorational mixture for control of seedling diseases of sugar beet and root knot nematode on tomato. Plant Dis. 2007, 91, 220–225. [Google Scholar] [CrossRef]
- Strobel, G.A.; Knighton, B.; Kluck, K.; Ren, Y.; Livinghouse, T.; Griffin, M.; Spakowicz, D.; Sears, J. The production of myco-diesel hydrocarbons and their derivatives by the endophytic fungus Gliocladiumroseum (NRRL 50072). Microbiology 2008, 154, 3319–3328. [Google Scholar] [CrossRef] [PubMed]
- Worapong, J.; Strobel, G.; Daisy, B.; Castillo, U.F.; Baird, G.; Hess, M.W. Muscodor roseousanam. Sp. Nov. and endophyte from Grevillea pteridifolia. Mycotaxon 2002, 81, 463–475. [Google Scholar]
- González, M.C.; Anaya, A.L.; Glenn, A.E.; Macías-Rubalcava, M.L.; Hernández-Bautista, B.E.; Hanlin, R.T. Muscodor yucatanensis, a new endophytic ascomycete from Mexican chakah, Bursera simaruba. Mycotaxon 2009, 110, 363–372. [Google Scholar] [CrossRef]
- Mitchell, A.M.; Strobel, G.A.; Moore, E.; Robison, R.; Sears, J. Volatile antimicrobials from Muscodor crispans, a novel endophytic fungus. Microbiology 2010, 156, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Kudalkar, P.; Strobel, G.; Riyaz-Ul-Hassan, S.; Geary, B.; Sears, J. Muscodor sutura, a novel endophytic fungus with volatile antibiotic activities. Mycoscience 2012, 53, 319–325. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kumla, J.; Bussaban, B.; Hyde, K.D.; Matsui, K.; Lumyong, S. Molecular and morphological evidence support four new species in the genus Muscodor from northern Thailand. Ann. Microbiol. 2013, 63, 1341–1351. [Google Scholar] [CrossRef]
- Meshram, V.; Kapoor, N.; Saxena, S. Muscodor kashayum sp. nov.—A new volatile anti-microbial producing endophytic fungus. Mycology 2013, 4, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.; Meshram, V.; Kapoor, N. Muscodor darjeelingensis, a new endophytic fungus of Cinnamomum camphora collected from north eastern Himalayas. Sydowia 2014, 66, 55–67. [Google Scholar]
- Meshram, V.; Saxena, S.; Kapoor, N. Muscodor strobelii, a new endophytic species from South India. Mycotaxon 2014, 128, 93–104. [Google Scholar] [CrossRef]
- Saxena, S.; Meshram, V.; Kapoor, N. Muscodor tigerii sp. nov. Volatile antibiotic producing endophytic fungus from the North eastern Himalayas. Ann. Microbiol. 2015, 65, 47–57. [Google Scholar] [CrossRef]
- Siri-udom, S.; Suwannarach, N.; Lumyong, S. Existence of Muscodor vitigenus, M. equiseti and M. heveae sp. nov.in leaves of the rubber tree (Hevea brasiliensis Müll. Arg.), and their biocontrol potential. Ann. Microbiol. 2015, 66, 437–448. [Google Scholar] [CrossRef]
- Meshram, V.; Gupta, M.; Saxena, S. Muscodor ghoomensis and Muscodor indica: New endophytic species based on morphological features, molecular and volatile organic analysis from northeast India. Sydowia 2016, 67, 133–146. [Google Scholar]
- Meshram, V.; Kapoor, N.; Chopra, G.; Saxena, S. Muscodor camphora, a new endophytic species from Cinnamomum camphora. Mycosphere 2017, 8, 568–582. [Google Scholar] [CrossRef]
- Scherlach, K.; Hertweck, C. Triggering cryptic natural product biosynthesis in microorganisms. Org. Biomol. Chem. 2009, 7, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.B.; Henrikson, J.C.; Hoover, A.R.; Lee, A.E.; Cichewicz, R.H. Epigenetic remodeling of the fungal secondary metabolome. Org. Biomol. Chem. 2008, 6, 1895–1897. [Google Scholar] [CrossRef] [PubMed]
- Cichewicz, R.H. Epigenome manipulation as a pathway to new natural product scaffolds and their congeners. Nat. Prod. Rep. 2010, 27, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, E.M.; Howlett, B.J. Secondary metabolism, regulation and role in fungal biology. Curr. Opin. Microbiol. 2008, 11, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Fisch, K.M.; Gillaspy, A.F.; Gipson, M.; Henrikson, J.C.; Hoover, A.R.; Jackson, L.; Najar, F.Z.; Wägele, H.; Cichewicz, R.H. Chemical induction of silent pathway transcription in Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2009, 36, 1199–1213. [Google Scholar] [CrossRef] [PubMed]
- Magotra, A.; Kumar, M.; Kushwaha, M.; Awasthi, P.; Raina, C.; Gupta, A.P.; Shah, B.A.; Gandhi, S.G.; Chaubey, A. Epigenetic modifier induced enhancement of fumiquinazoline C production in Aspergillus fumigatus (GA-L7): An endophytic fungus from Grewia asiatica L. AMB Express 2017, 7, 43. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Kusari, S.; Golz, C.; Laatsch, H.; Strohmann, C.; Spiteller, M. Epigenetic modulation of endophytic Eupenicillium sp. LG41 by a histone deacetylase inhibitor for production of decalin-containing compounds. J. Nat. Prod. 2017, 80, 983–988. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Kusari, S.; Lamshöft, M.; Schüffler, A.; Laatsch, H.; Spiteller, M. Antibacterial secondary metabolites from an endophytic fungus, Eupenicillium sp. LG41. J. Nat. Prod. 2014, 77, 2335–2341. [Google Scholar] [CrossRef] [PubMed]
- Vasanthakumari, M.M.; Jadhav, S.S.; Sachin, N.; Vinod, G.; Singh, S.; Manjunatha, B.L.; Kumara, P.M.; Ravikanth, G.; Nataraja, K.N.; Uma Shaanker, R. Restoration of camptothecine production in attenuated endophytic fungus on re-inoculation into host plant and treatment with DNA methyltransferase inhibitor. World. J. Microbiol. Biotechnol. 2015, 31, 1629–1639. [Google Scholar] [CrossRef] [PubMed]
- Gubiani, J.R.; Wijeratne, E.M.K.; Shi, T.; Araujo, A.R.; Arnold, A.E.; Chapman, E.; Gunatilaka, A.A.L. An epigenetic modifier induces production of (10′ S)-verruculide B, an inhibitor of protein tyrosine phosphatases by Phoma sp. nov. LG0217, a fungal endophyte of Parkinsonia microphylla. Bioorg. Med. Chem. 2017, 25, 1860–1866. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Zhu, G.; Hao, J.; Wang, Y.; Zhu, W. Chemical-epigenetic Method to Enhance the Chemodiversity of the Marine Algicolous Fungus, Aspergillus terreus OUCMDZ-2739. Tetrahedron 2018, 74, 83–87. [Google Scholar] [CrossRef]
- Akone, S.H.; Mandi, A.; Kurtan, T.; Hartmann, R.; Lin, W.; Daletos, G.; Proksch, P. Inducing secondary metabolite production by the endophytic fungus Chaetomium sp. through fungal-bacterial co-culture and epigenetic modification. Tetrahedron 2016, 72, 6340–6347. [Google Scholar] [CrossRef]
- Asai, T.; Otsuki, S.; Sakurai, H.; Yamashita, K.; Ozeki, T.; Oshima, Y. Benzophenones from an endophytic fungus, Graphiopsis chlorocephala, from Cultivated in the Presence of an NAD+-Dependent HDAC Inhibitor. Org. Lett. 2013, 15, 2058–2061. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Menendez, V.; Perez-Bonilla, M.; Perez-Victoria, I.; Martin, J.; Munoz, F.; Reyes, F.; Tormo, J.R.; Genilloud, O. Multicomponent analysis of the differential induction of secondary metabolite profiles in fungal endophytes. Molecules 2016, 21, 234. [Google Scholar] [CrossRef] [PubMed]
- Rico-Gray, V. Interspecific interaction. Encycl. Life Sci. 2001. [Google Scholar] [CrossRef]
- Sandland, G.J.; Rodgers, J.K.; Minchella, D.J. Interspecific antagonism and virulence in hostsex-posed to two parasite species. J. Invertebr. Pathol. 2007, 96, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Marmann, A.; Aly, A.H.; Lin, W.; Wang, B.; Proksch, P. Co-Cultivation—A powerful emerging tool for enhancing the chemical diversity of microorganisms. Mar. Drugs 2014, 12, 1043–1065. [Google Scholar] [CrossRef] [PubMed]
- Brakhage, A.A.; Schroeckh, V. Fungal secondary metabolites–strategies to activate silent gene clusters. Fungal Genet. Biol. 2011, 48, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Ola, A.R.; Thomy, D.; Lai, D.; Brötz-Oesterhelt, H.; Proksch, P. Inducing secondary metabolite production by the endophytic fungus Fusarium tricinctum through co-culture with Bacillus subtilis. J. Nat. Prod. 2013, 76, 2094–2099. [Google Scholar] [CrossRef] [PubMed]
- Kusari, S.; Singh, S.; Jayabaskaran, C. Rethinking production of Taxol® (paclitaxel) using endophyte biotechnology. Trends Biotechnol. 2014, 32, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, W.; El-Neketi, M.; Lewald, L.I.; Orfali, R.S.; Lin, W.; Rehberg, N.; Kalscheuer, R.; Daletos, G.; Proksch, P. Metabolites from the Fungal Endophyte Aspergillus austroafricanus in Axenic Culture and in Fungal-Bacterial Mixed Cultures. J. Nat. Prod. 2016, 79, 914–922. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structures of metabolites isolated from Coelomycetes (1–24).

Figure 2.
Structures of metabolites isolated from Coelomycetes (25–37) and Ascomycetes (38–49).

Figure 3.
Structures of metabolites isolated from Ascomycetes (50–68).

Figure 4.
Structures of metabolites isolated from Ascomycetes (69–92).

Figure 5.
Structures of metabolites isolated from Ascomycetes (93–114).

Figure 6.
Structures of metabolites isolated from Ascomycetes (115–127) and Hyphomycetes (128–137).

Figure 7.
Structures of metabolites isolated from Hyphomycetes (138–155).

Figure 8.
Structures of metabolites isolated from Hyphomycetes (156–169) and Basidiomycetes (170–172).
Figure 8.
Structures of metabolites isolated from Hyphomycetes (156–169) and Basidiomycetes (170–172).

Figure 9.
Structures of metabolites isolated from Epigenetic modification in endophytic fungi (173–189).
Figure 9.
Structures of metabolites isolated from Epigenetic modification in endophytic fungi (173–189).

Figure 10.
Structures of metabolites obtained from Epigenetic modification in endophytic fungi (190–208).
Figure 10.
Structures of metabolites obtained from Epigenetic modification in endophytic fungi (190–208).

Figure 11.
Structures of metabolites obtained from Epigenetic modification in endophytic fungi (209–225).
Figure 11.
Structures of metabolites obtained from Epigenetic modification in endophytic fungi (209–225).

Figure 12.
Structures of metabolites obtained from co-culture in endophytic fungi (226–247).

Figure 13.
Structures of metabolites obtained from co-culture in endophytic fungi (248–256).


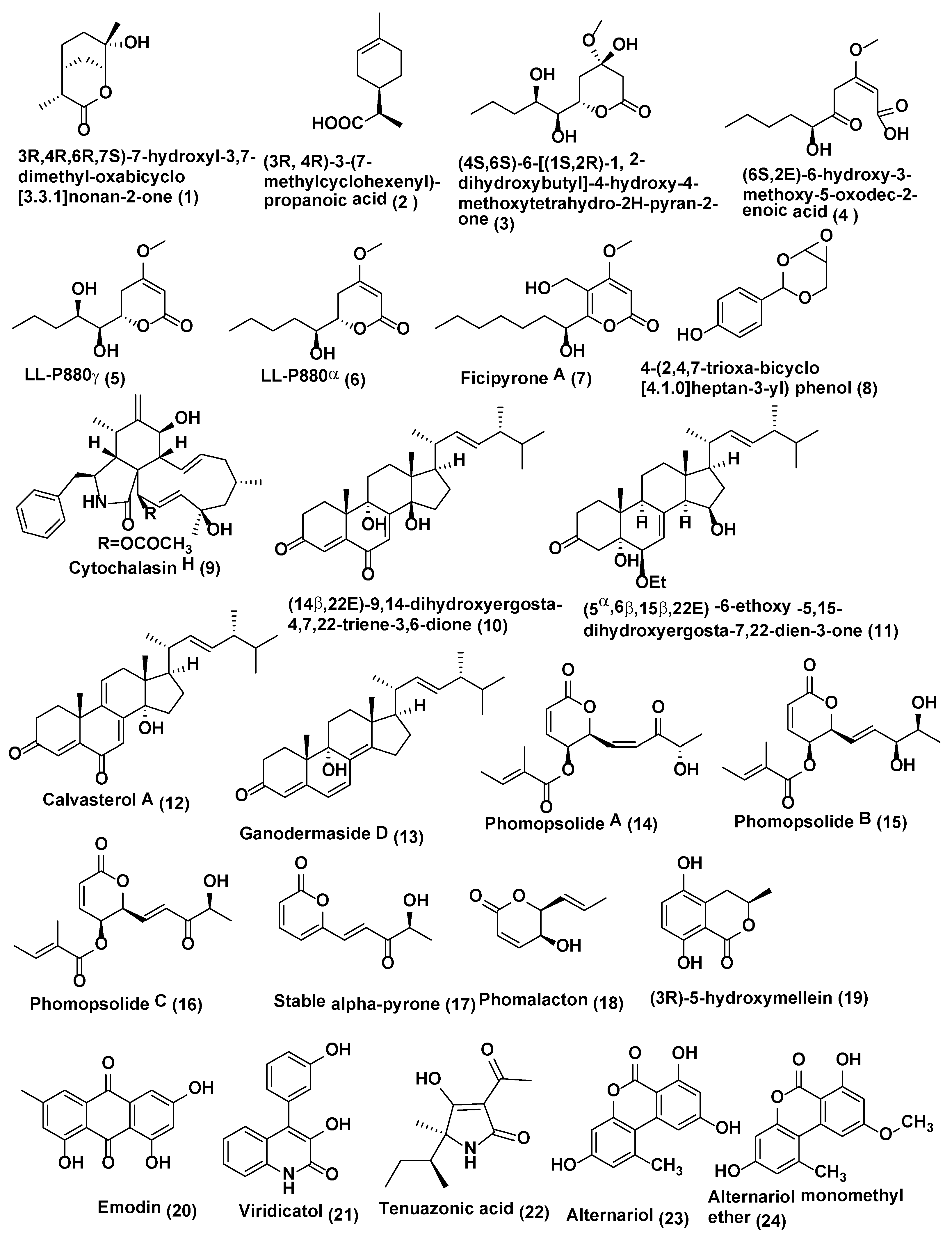{kind=link}
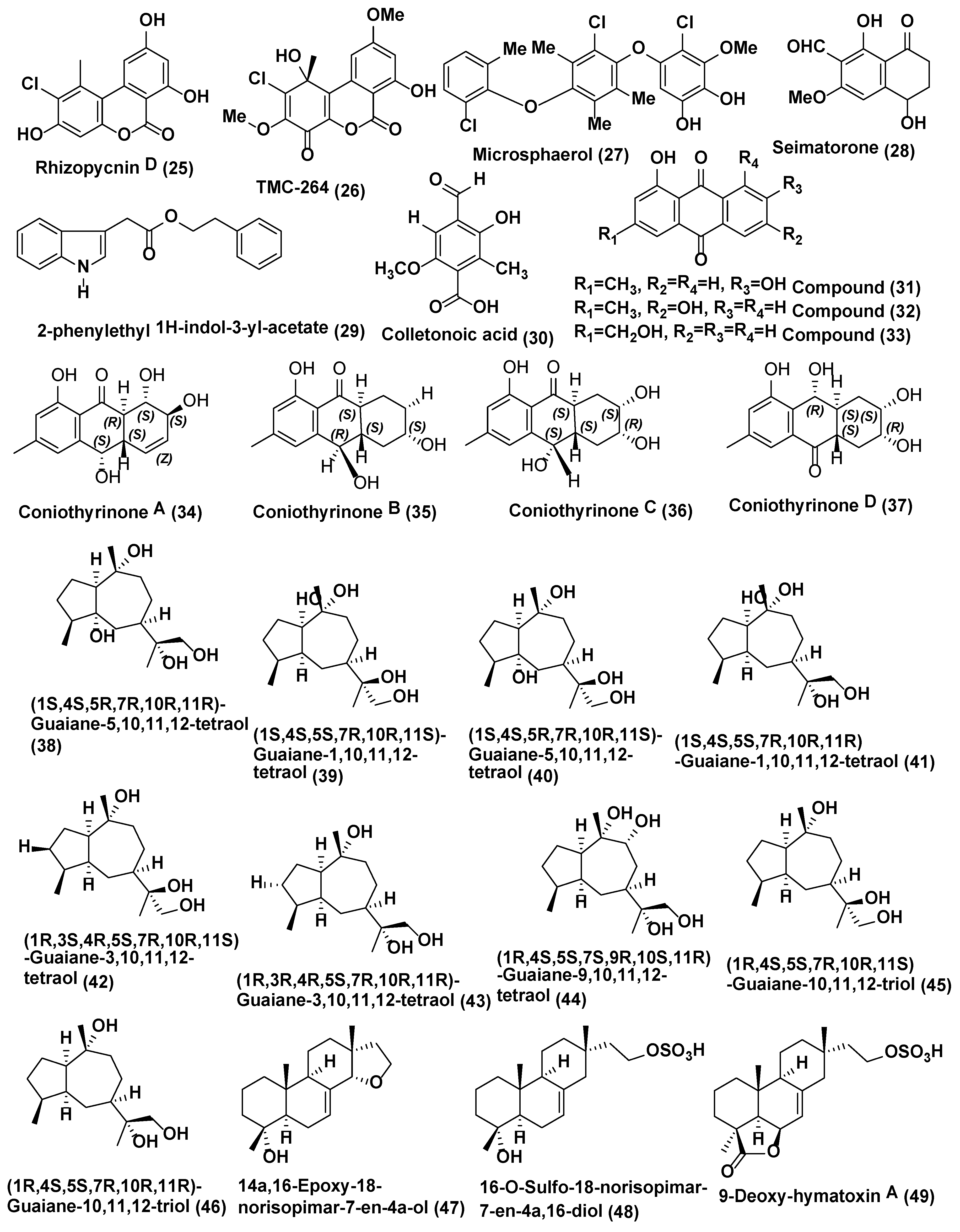{kind=link}
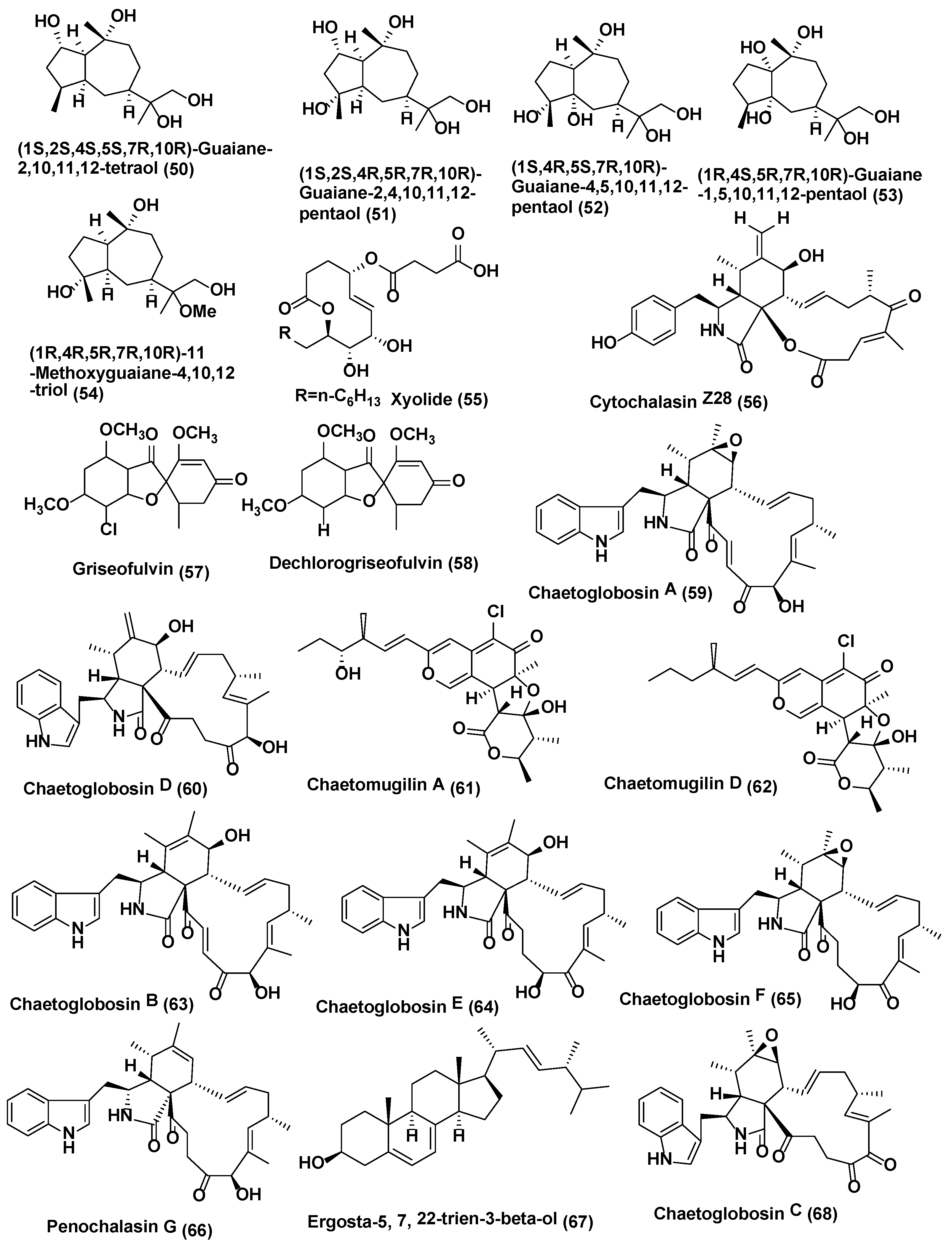{kind=link}
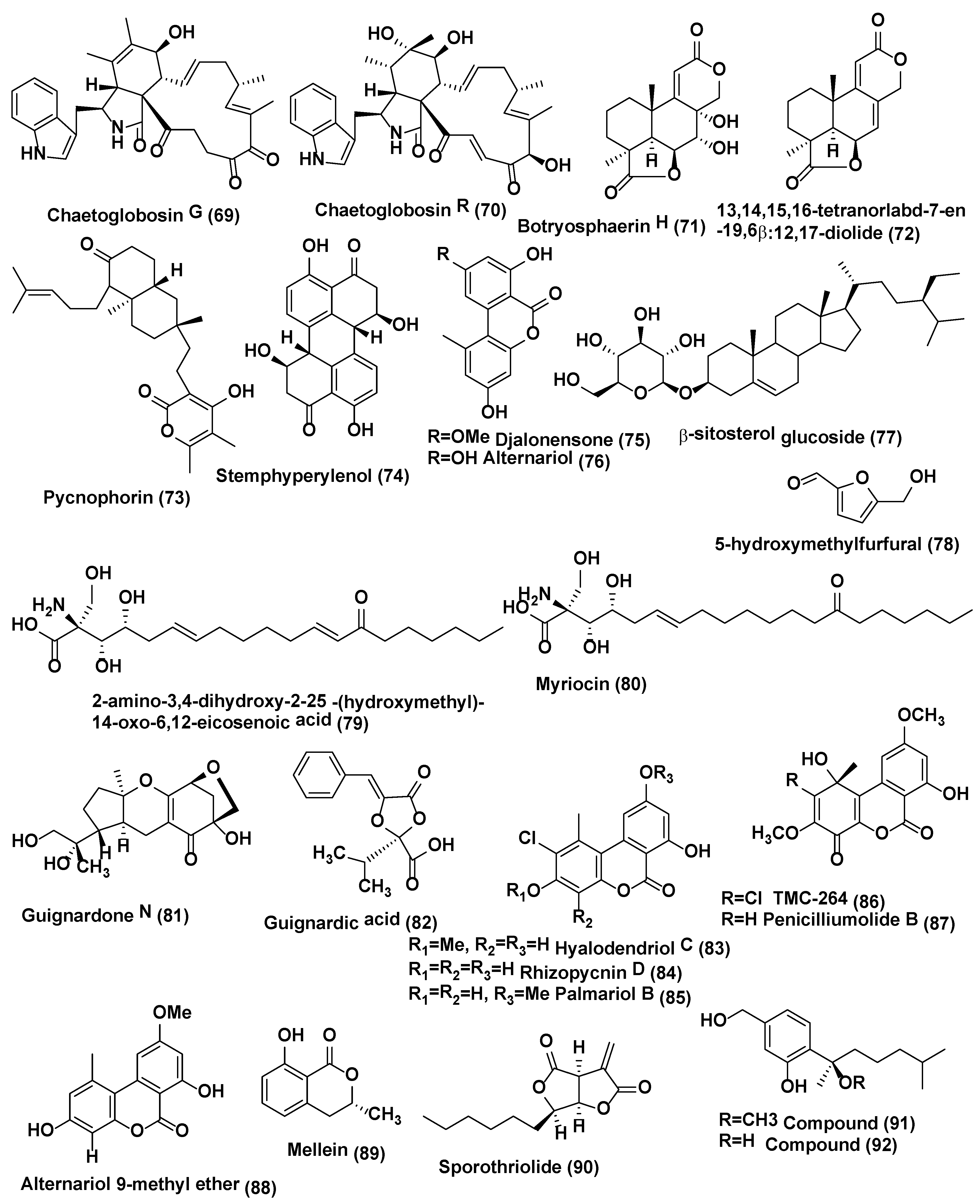{kind=link}
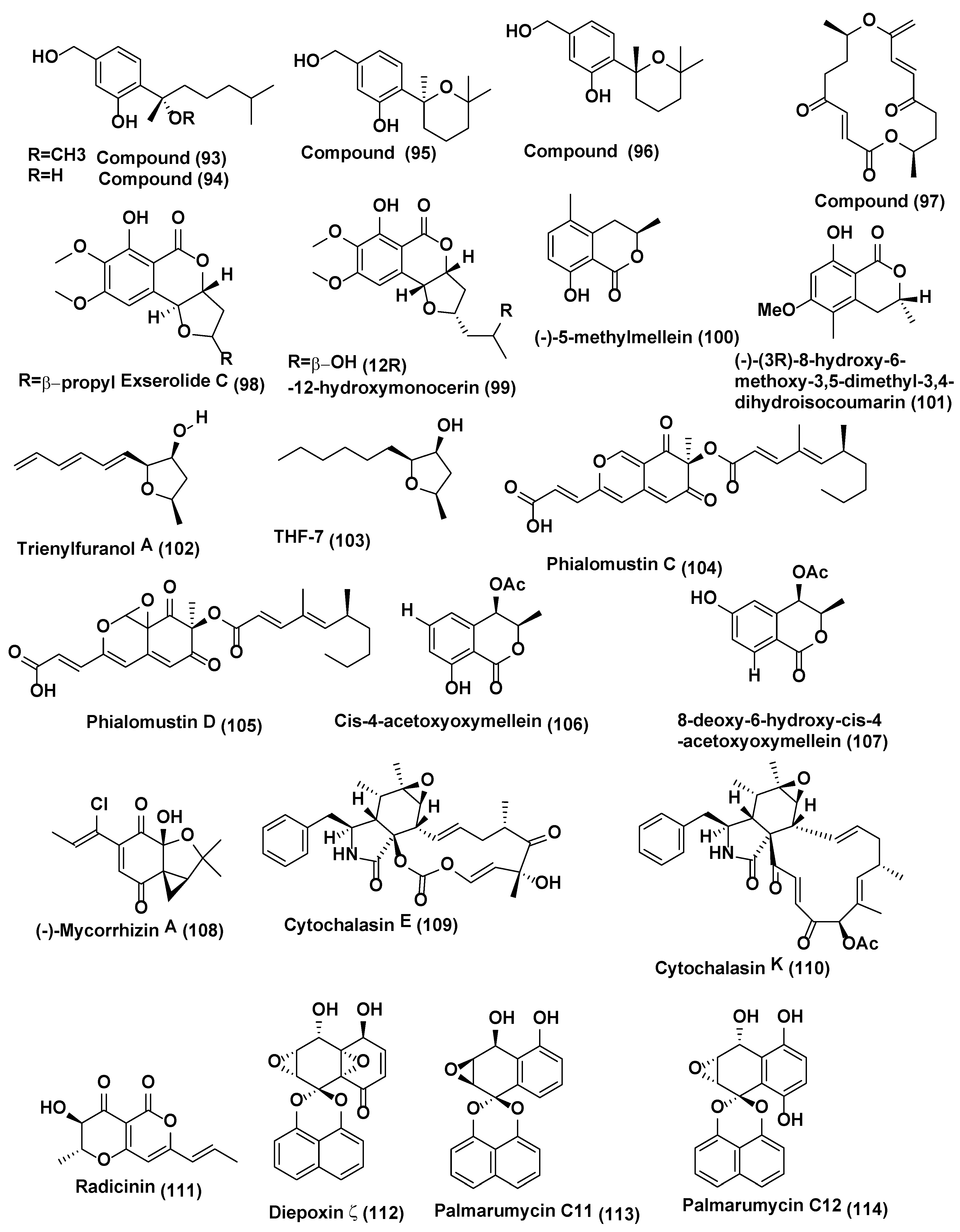{kind=link}
{kind=link}
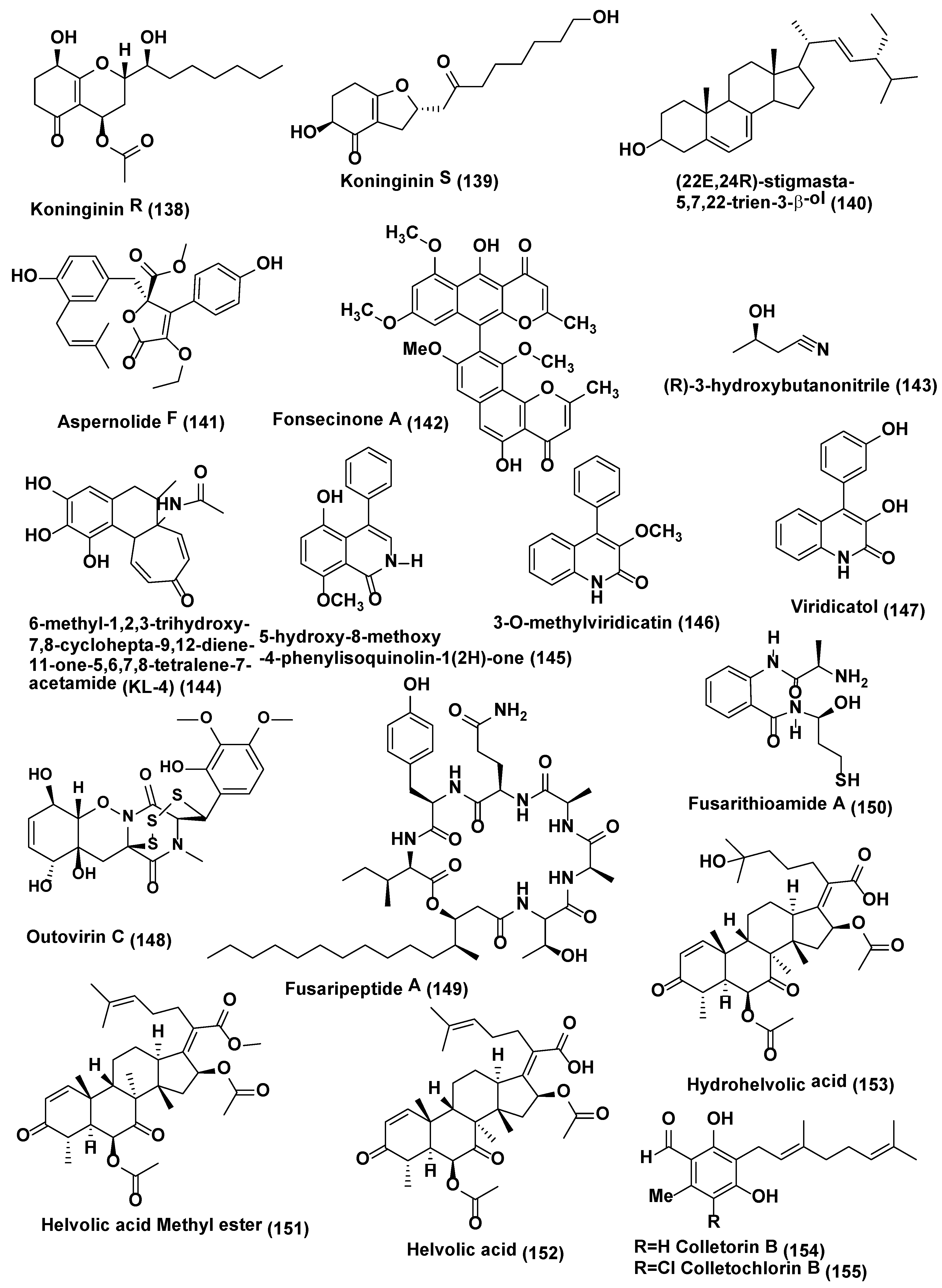{kind=link}
{kind=link}
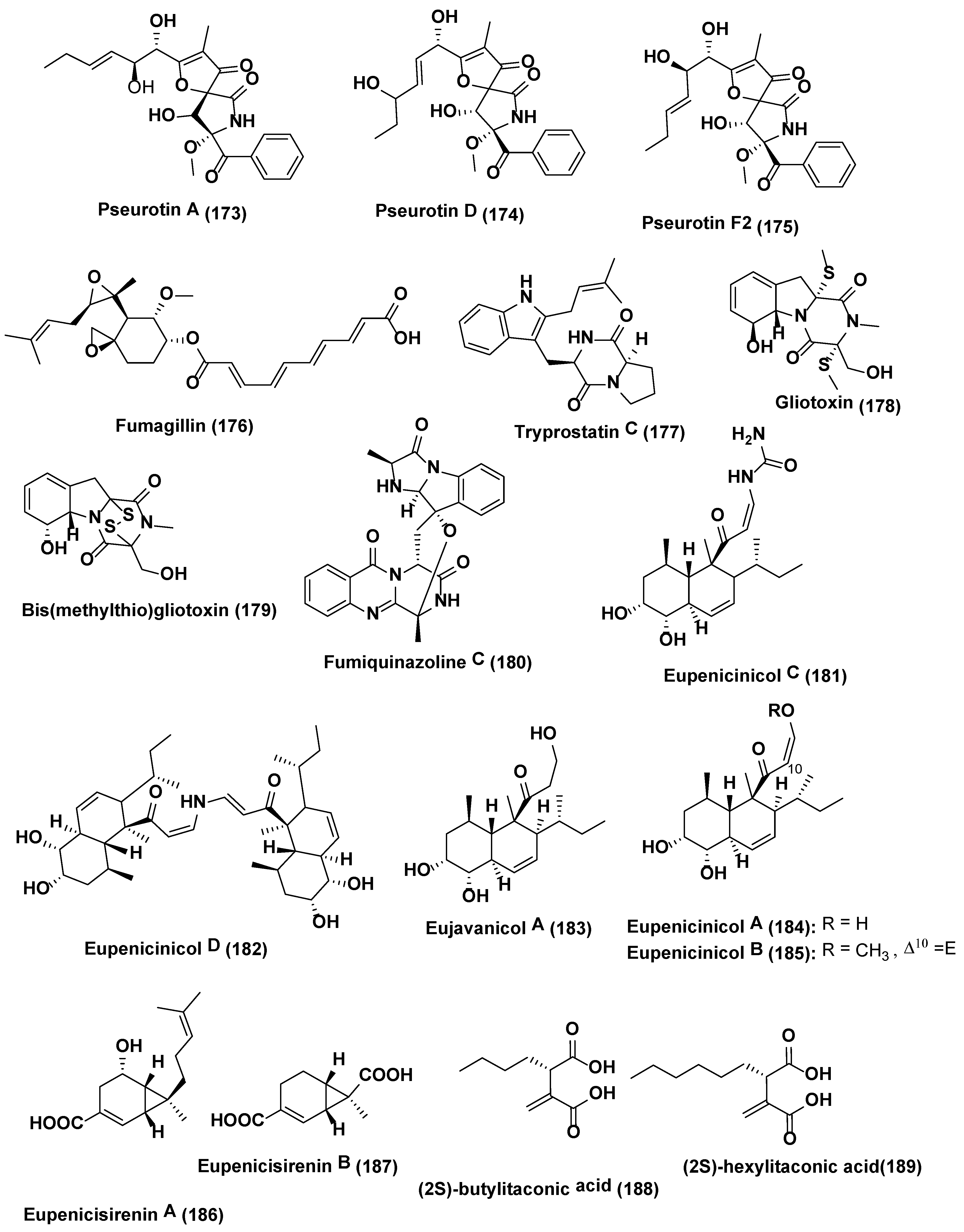{kind=link}
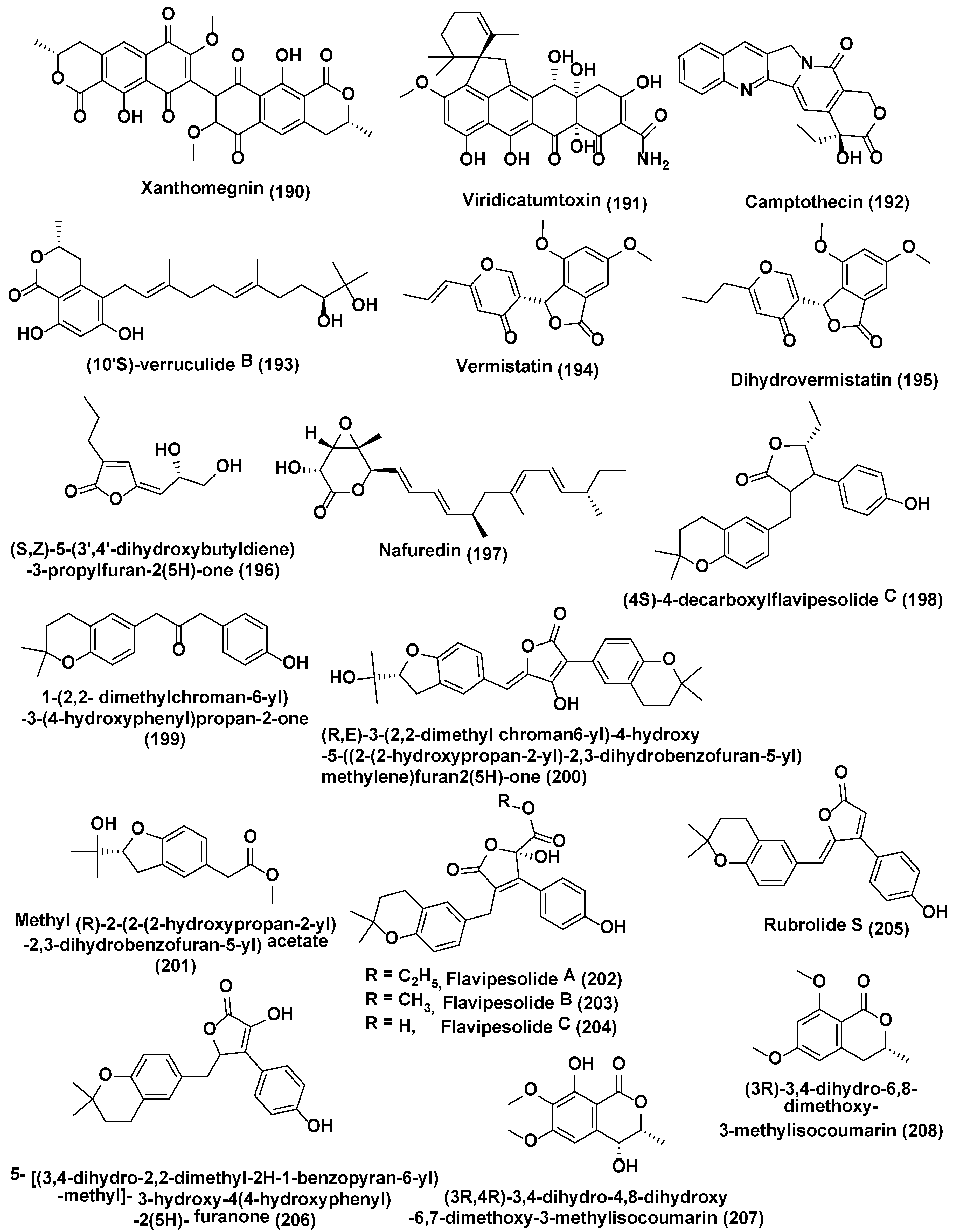{kind=link}
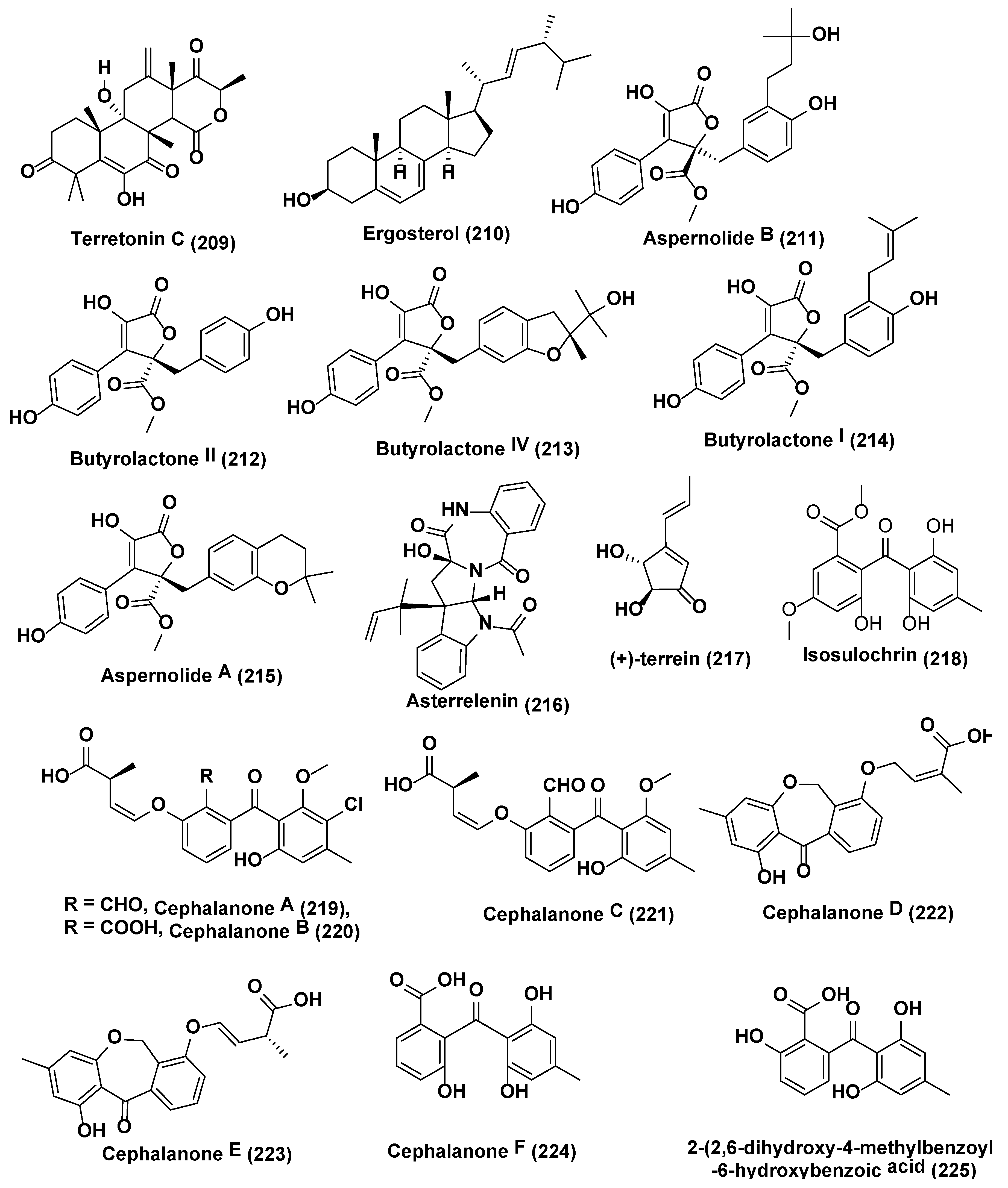{kind=link}
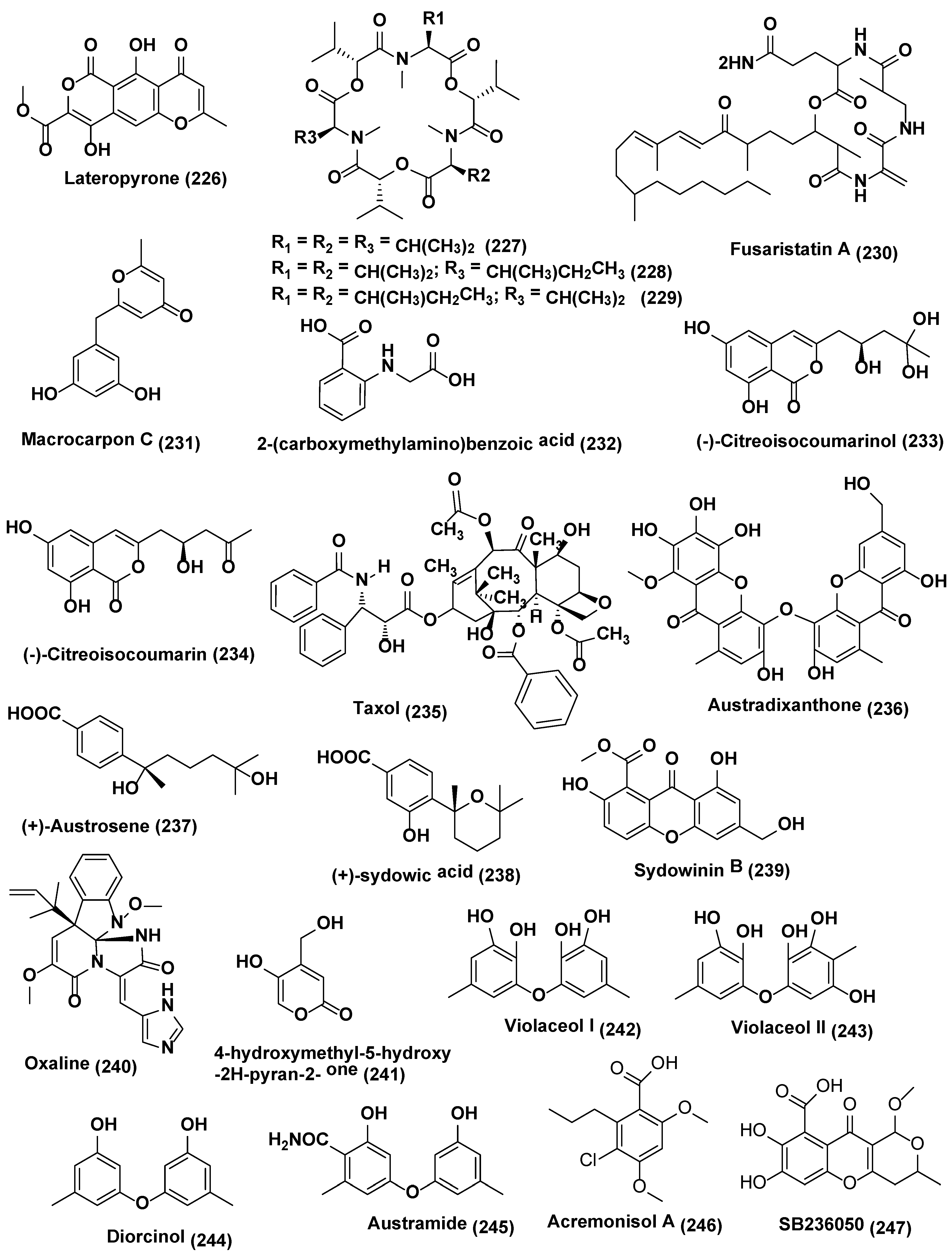{kind=link}
{kind=link}
Table 1.
Antifungal compounds reported from endophytic fungi.
| Sr. No. | Fungus | Plantsource | Compounds Isolated | Biologicalactivity * | Refs. |
|---|---|---|---|---|---|
| Comounds Produced by Coelomycetes | |||||
| 1 | Pestalotiopsis foedan | Bruguiera sexangula Hainan, China | (3R,4R,6R,7S)-7-hydroxyl-3,7-dimethyl-oxabicyclo[3.3.1]nonan-2-one (1), (3R,4R)-3-(7-methylcyclohexenyl)-propanoic acid (2) | Compound 1 B. cinerea and P. nicotianae (MIC 3.1 and 6.3 µg/mL), ketoconazole (MIC 3.1 µg/mL each) Compound 2 C. albicans MIC 50 µg/mL) ketoconazole (MIC 6.3 µg/mL) | [15] |
| 2 | Pestalotiopsis sp. DO14 | Dendrobium officinale, Yandang Mountain, Zhejiang Province, China. | (4S,6S)-6-[(1S,2R)-1,2-dihydroxybutyl]-4-hydroxy-4-methoxytetrahydro-2H-pyran-2-one (3) and (6S,2E)-6-hydroxy-3-methoxy-5-oxodec-2-enoic acid (4), LL-P880γ (5), LL-P880α (6) | Compounds 3–6 active against C. albicans, C. neoformans, T. rubrum, and A. fumigates (MIC ≤ 50 µg/mL) Compounds 3–4 active against C. albicans, C. neoformans, T. rubrum, and A. fumigatus (MIC, ≤ 25 µg/mL) | [16] |
| 3 | Pestalotiopsis fici | Camellia sinensis Hangzhou, China. | Ficipyrone A (7) | Compound 7 active against G. zeae (IC50 15.9 µM), ketoconazole (IC50 6.02 µM) | [17] |
| 4 | Pestalotiopsis mangiferae | Mangifera indica Maduravoyal, Tamil Nadu Province, India. | 4-(2,4,7-trioxa-bicyclo[4.1.0]heptan-3-yl) phenol (8) | Compound 8 active against C. albicans (MIC, 0.039 µg/mL), Nystatin (MIC 10.0 µg/mL) | [18] |
| 5 | Phomopsis sp. | Senna spectabilis São Paulo, Brazil | Cytochalasin H (9) | Compound 9 active against C. cladosporioides and C. sphaerosphermum (MIC 10.0 and 25.0 µg, respectively), nystatin (MIC = 1.0 µg) | [19] |
| 6 | Phomopsis sp. | Aconitum carmichaeli, Huize County, Yunnan Province, China. | (14β,22E)-9,14-dihydroxyergosta-4,7,22-triene-3,6-dione (10), (5α,6β,15β,22E)-6-ethoxy-5,15-dihydroxyergosta-7,22-dien-3-one (11), calvasterols A (12), and ganodermaside D (13) | Compound 10 active against C. albicans, H. compactum, and A. niger, (MIC, 64, 64, and 128 µg/mL, respectively). Compound 11 active against C. albicans and F. avenaceum (MIC = 128 µg/mL). Compounds 12 and 13 active against F. avenaceum. (MIC, 64 µg/mL), Compound 12 activie against P. oryzae and T. gypseum (MIC 128 and 256 µg/mL) | [20] |
| 7 | Diaporthe maritima | Picea sp., Acadian forest of Eastern Canada. | Phomopsolide A (14), B (15), and C (16), and a stable alpha-pyrone (17) | Compound 14 active against M. violaceum and S. cerevisiae at 25 µM, Compounds 15–17 demonstrated growth inhibition at 250 µM | [21] |
| 8 | Phoma sp. | Fucus serratus, | Phomalacton (18), (3R)-5-hydroxymellein (19) and emodin (20) | Compounds 18–20 active against M. violaceum with 5, 6 and 5mm zone of inhibition. | [23] |
| 9 | Phoma sp. WF4 | Eleusine coracana Arkell Field Station, Arkell, ON, Canada | Viridicatol (21), tenuazonic acid (22), alternariol (23), and alternariol monomethyl ether (24) | Compounds 21–24 caused dramatic breakage of F. graminearum hyphae in vitro | [24] |
| 10 | Rhizopycnis vagum Nitaf 22 | Nicotiana tabacum, China Agricultural University. Beijing 100193, China. | Rhizopycnin D (25) and TMC-264 (26) | Compounds 25–26 inhibited the spore germination of M. oryzae with IC50 values of 9.9 and 12.0 µg/mL, respectively | [25] |
| 11 | Microsphaeropsis sp. Seimatosporium sp. | Salsola oppositifolia, Playa del Ingles, Gomera, Spain | Microsphaerol (27) Seimatorone (28) | Compounds 27 and 28 active against M. violaceum with 9 and 5 mm zone of inhibition. In addition, there was some growth with in zone of inhibition | [26] |
| 12 | Colletotrichum gloeosporioides | Michelia champaca São Paulo State University, Araraquara, São Paulo, Brazil. | 2-phenylethyl 1H-indol-3-yl-acetate (29) | Compound 29 active against C. cladosporioides and C. sphaerospermum comparable to that of the positive control nystatin | [27] |
| 13 | Colletotrichum sp. | Gomera (Spain). | Colletonoic acid (30) | Compound 30 active against M. violaceum with 7 mm zone of inhibition | [28] |
| 14 | Coniothyrium sp., | Salsola oppostifolia Gomera in the Canary Islands. | 1,7-dihydroxy3-methyl-9,10-anthraquinone (31), 1,6-dihydroxy-3 -methyl-9,10-anthraquinone (phomarin) (32), and 1-hydroxy-3-hydroxymethyl-9,10-anthraquinone (33) coniothyrinones A-D (34–37) | Compounds 31–37 active against M. violaceum with 7, 10, 8, 7.5, 6, 8 and 7.5 mm zone of inhibition. Compounds 32–34 active against M. violaceum (10 and 9 mm zone of inhibition) and B. cinerea (7.5 and 12.5 mm zone of inhibition) when tested under similar conditions | [29,30] |
| Comounds Produced by Acsomycetes | |||||
| 15 | Xylaria sp. YM 311647 | Azadirachta indica,Yuanjiang County, Yunnan Province, China, | (1S,4S,5R,7R,10R,11R)-Guaiane-5,10,11,12-tetraol (38) (1S,4S,5S,7R,10R,11S) -Guaiane-1,10,11,12-tetraol (39) (1S,4S,5R,7R,10R,11S)-Guaiane-5,10,11,12-tetraol (40) (1S,4S,5S,7R,10R,11R)-Guaiane-1,10,11,12-tetraol (41) (1R,3S,4R,5S,7R,10R,11S) -Guaiane-3,10,11,12-tetraol (42) (1R,3R,4R,5S,7R,10R,11R)-Guaiane-3, 10,11,12-tetraol (43) (1R,4S,5S,7S,9R,10S,11R)-Guaiane-9,10,11,12-tetraol (44) (1R,4S,5S,7R,10R,11S) -Guaiane-10,11,12-triol (45) (1R,4S,5S,7R,10R,11R)-Guaiane-10,11,12-triol (46), 14a,16-Epoxy-18-norisopimar-7-en-4a-ol (47),16-O-Sulfo-18-norisopimar-7-en-4a,16-diol (48), and 9-Deoxy-hymatoxin A (49) | Compounds 38–46 active against C. albicans and H. compactum (MIC in the range of 32 to 256 µg/mL), compounds 47–49 active against C. albicans, A. niger, P. oryzae, F. avenaceum, and H. compactum (MIC in the range of 16 to 256 µg/mL). Compound 49 exhibited the potent inhibitory activity against C. albicans and P. oryzae with MIC values of 16 µg/mL | [32] |
| 16 | Xylaria sp. YM 31164 | Azadirachta indica, Yuanjiang County, Yunnan Province, China | (1S,2S,4S,5S,7R,10R)-Guaiane-2,10,11,12-tetraol (50), (1S,2S,4R,5R,7R,10R)-Guaiane-2,4,10,11,12-pentaol (51), (1S,4R,5S,7R,10R)-Guaiane-4,5,10,11,12-pentaol (52), (1R,4S,5R,7R,10R)-Guaiane-1,5,10,11,12-pentaol (53), (1R,4R,5R,7R,10R)-11-Methoxyguaiane-4,10,12-triol (54), | Compounds 50–54 active against P. oryzae and H. compactum (MIC in the range of 32–256 µg/mL). Compound 53 active against P. oryzae (MIC 32 µg/mL). Compounds 52 and 53 active against H. compactum with (MIC, 64 µg/mL), Compound 53 and 54 active against C. albicans (MIC 32 µg/mL). Compound 52 active against C. albicans, A. niger, and H. compactum (MIC, 64 µg/mL). | [33] |
| 17 | X. feejeensis | Croton lechleri. | Xyolide (55), | Compound 55 active against P. ultimum (MIC 425 µM) | [34] |
| 18 | Xylaria sp. XC-16 | Toona sinensis Yangling, Shaanxi Province, China | Cytochalasin Z28 (56) | Compound 56 active against G. saubinetti (MIC of 12.5 µM), Hymexazol (MIC = of 25 µM) | Zhang et al. [35] |
| 19 | Xylaria sp. strain F0010, Xylaria sp. PSU-G12X. cubensis, 13 strains of Xylaria sp. | Abies holophylla; Garcinia hombroniana ; Asimina triloba; Pinus strobus; Vaccinium angustifolium, New Brunswick and Nova Scotia, Canada | Griseofulvin (57) | Griseofulvin (57) Inhibits A. mali, B. cinerea, Colletotrichum gloeosporioides, Corticium sasaki, F. oxysporum and M. grisea in in vitro (IC50 values of 18.0, 5.0, 1.7, 11.0, 30.0, and 1.7 µg/mL, respectively, Compound 57 active against M. grisea, C. sasaki, B. cinerea, P. recondite and B .graminis f. sp. hordei in in vivo, with % of fungal control of 95, 100, 60, 90 and 90, respectively, at 150 µg/mL. | [36,37,38] |
| 20 | Xylaria sp. | Dechlorogriseofulvin (58) | Compound 58 showed weak antifungal activity, with an IC50 value, 200 µg/mL against M. grisea, C. sasaki, B. cinerea, P. recondite and B .graminis f. sp. hordei in in vivo, | [36,38] | |
| 21 | Chaetomium globosum CDW7 | Ginkgo biloba China | Chaetoglobosin A (59) and D (60) | Compounds 59–60 active against S. sclerotiorum with IC50 values of 0.35 and 0.62 µg/mL, respectively, carbendazim (0.17 µg/mL) | [41] |
| 22 | Chaetomium globosum | Seeds of Panax notoginseng collected at the Wenshan, Yunnan, China | Chaetoglobosin A (59), Chaetomugilin A (61), Chaetomugilin D (62), Chaetoglobosin B (63), Chaetoglobosin E (64), Chaetoglobosin F (65) and Penochalasin G (66) | Compounds 59 and 61–66 active against P. herbarum (MIC in the range of 16–128 µg/mL) and, E. nigrum (MIC in the range of <1–16 µg/mL). | [42] |
| 23 | Chaetomium cupreum ZJWCF079 | Macleaya cordata. | Ergosta-5, 7, 22-trien-3-beta-ol (67) | Compound 67 against S. sclerotiorum and B. cinerea with EC50 values of 125 µg/mL and 190 µg/mL respectively, | [43] |
| 24 | Chaetomium globosum No.04 | Barks of Ginkgo biloba, Linyi, Shandong Province, China. | Chaetoglobosin A (59), D (60), E (64), C (68), G (69), R (70) | Compounds 59–60, 64, and 68–70 active against R. stolonifer and C. diplodiella at a concentration of 20 µg/disk | [44] |
| 25 | Botryosphaeria sp. P483 | Huperzia serrata, Xichou County, Yunnan Province, China | Botryosphaerin H (71) 13,14,15,16-tetranorlabd-7-en-19,6β:12,17-diolide (72) | At 100 µg/disk, compound 71 showed zone of inhibition of 9, 7, 7, 8, and 8 mm, against G. graminis, F. solani, P. oryzae, F. moniliforme, and F. oxysporum; compound 72 showed zone of inhibition of 12, 10, 10, 11, and 13 mm against G. graminis, F. solani, P. oryzae, F. moniliforme, and F. oxysporum; carbendazim (50 µg/disk) showed the zone of inhibition of 14, 18, 15, 17, 15 mm against G. graminis F. solani P. oryzae F. moniliforme F. oxysporum, respectively | [45] |
| 26 | Botryosphaeriadothidea KJ-1, | Melia azedarach Yangling, Shaanxi Province, China. | Pycnophorin (73), stemphyperylenol (74), chaetoglobosin C (68), djalonensone (75), alternariol (76), β-sitosterol glucoside (77), 5-hydroxymethylfurfural (78) | Compound 74 active against A. solani (MICs of 1.57 µM) Compounds 68, 73, and 75–78 active against A. solani (MICs of 6.25−25 µM) | [46] |
| 27 | Mycosphaerella sp. | Eugenia bimarginata DC. Brazil (savannah). | 2-amino-3,4-dihydroxy-2-25-(hydroxymethyl)-14-oxo-6,12-eicosenoic acid (79), myriocin (80) | Compounds 79 active against several isolates of C. neoformans and C. gattii, with MIC values ranging from 1.3 to 2.50 µg/mL and 0.5 µg/mL, for compound 80 | [47] |
| Compounds 79 active against several isolates of C. neoformans and C. gattii, with MIC values ranging from 0.49 to 7.82 µM and 0.48–1.95 µM for compound 80. Compounds 79 and 80 cause deformities in cell shape, depressions on the surface, and withered cells. | [48] | ||||
| 28 | Guignardia sp., | Euphorbia sieboldiana collected from the campus of China Pharmaceutical University, Nanjing, Jiangsu, China | Guignardone N (81), guignardic acid (82) | At 6.3 µg/mL combined with 0.031 µg/mL of fluconazole, compounds 81 and 82 were found to have prominent inhibition on the growth of C. albicans with FIC index values of 0.23 and 0.19, respectively. Combined with fluconazole, both of them (40 µg/mL for (81) and 20 µg/mL for (82) could also inhibit C. albicans biofilms and reverse the tolerance of C. albicans biofilms to fluconazole | [49] |
| 29 | Hyalodendriella sp. Ponipodef 12 | “Neva” hybrid of Populus deltoides Marsh × P. nigra L., Longhua in Hebei Province of China. | hyalodendriol C (83), rhizopycnin D (84), palmariol B (85), TMC-264 (86), penicilliumolide B (87) and alternariol 9-methyl ether (88) | Compound 88 exhibited spore germination of M. oryzae with IC50 value of 11.6 µg/mL, positive control, carbendazim (IC50 6.9 µg/mL) Compounds 84–88 displayed antifungal effects against the spore germination of M. oryzae | [50,51,52] |
| 30 | Pezicula sp. | Forsythia viridissima, collected from Zhejiang Province, Southeast China | Mellein (89) | Compound 89 active against B. cinerea, P. ultimum, F. oxysporium f. sp. cucumerinum, C. orbiculare, V. dahliae, P. oryzae, P. diospyri, S. sclerotiorum and F. fulva, especially B. cinerea and F. fulva with EC50 values below 50 µg/mL | [53] |
| 31 | Nodulisporium sp. A21 | Leaves of Ginkgo biloba. Nanjing in Jiangsu Province, China | Sporothriolide, (90) | The EC50 of compound 90 against R.solani was 3.04 µg/mL (11.6 µM), while the EC50 of carbendazim was 1.84 µg/mL (9.6 µM). | [54] |
| 32 | Lopherdermium nitens DAOM 250027 | Pinus strobus Sussex, NB, Canada | Six phenolic bisabolane-type sesquiterpenoids (91–96), pyrenophorin (97) | Compound 97 significantly reduced the growth of M. violaceum and S. cerevisiae at 5 µM whereas sesquiterpenoids 91–96 active at 50 µM to both species tested | [55] |
| 33 | Exserohilum sp. | Acer truncatum Beijing, China. | Exserolide C (98), (12R)-12-hydroxymonocerin (99) | Compounds 98 and 99 active against F. oxysporum, both showing a MIC value of 20 µg/mL, Amphotericin B (MIC, 0.63 µg/mL) | [56] |
| 34 | Biscogniauxia mediterranea EPU38CA | Echinacea purpurea Missouri, USA, | (−)-5-methylmellein (100) and (−)-(3R)-8-hydroxy-6-methoxy-3,5-dimethyl-3, 4-dihydroisocoumarin (101) | Compound 100 active against P. obscurans, P. viticola, and F.oxysporum, and caused growth stimulation of C. fragariae, C. acutatum, C. gloeosporioides, and B. cinerea. Compound 101 was found to be slightly more active in the microtiter environment than 5-methylmellein | [57] |
| 35 | Hypoxylon submonticulosum | Rubus idaeus collected from Jordan Station, ON, Canada. | Trienylfuranol A (102) Complete hydrogenation of (102) yielded THF 7 (103) | THF 7 (103) inhibited the growth of S. cervisiae (74 ± 4% inhibition) at a concentration of 250 µg/mL as compared with complete inhibition by nystatin at 10 µg/mL | [58] |
| 36 | Phialophoramustea | Crocus sativus. | Phialomustin C-D (104) (105) | Compounds 104–105 active against C. albicans (IC50, 14.3 and 73.6 µM) | [59] |
| 37 | unidentified Ascomycete, | Melilotus dentatus. | cis-4-acetoxyoxymellein (106) and 8-deoxy-6-hydroxy-cis-acetoxyoxymellein (107) | Compounds 106 and 107 displayed activities toward M. violaceum, B. cinerea, with 8 mm zone of inhibition for both fungi. | [60] |
| 38 | Plectophomella sp. | (−)-Mycorrhizin A (108) | Compound 108 active against U. violacea and E. repens. | [61] | |
| 39 | Physalospora sp. | Cytochalasin E (109) and K (110) | Compound 109–110 active against E. repens and M. microspora | [61] | |
| 40 | Crataegus monogyna. | Radicinin (111) | Radicinin (111) active against E. repens and M. microspora | [61] | |
| 41 | Berkleasmium sp., | Dioscorea zingiberensis. Hubei Province, China. | Diepoxin ζ (112), palmarumycin C11 (113), palmarumycin C12 (114), cladospirone B (115), palmarumycin C6 (116), 1,4,7β-trihydroxy-8-(spirodioxy-1′,8′-naphthyl)-7,8-dihydronaphthalene (117) and palmarumycin C8 (118) | Compounds 112–118 inhibited spore germination of M. oryzae (IC50 values in the range 9.1−124.5 µg/mL). Compound 118 showed the best inhibitory activity (IC50, 9.1 µg/mL) among the compounds tested. Carbendazim (IC50 6.3 µg/mL) | [62] |
| 42 | Bipolaris sp. MU34 | Gynura hispida Bangkok, Thailand. | Bipolamide B (119) | Bipolamide B (119) active against C. cladosporioides, C. cucumerinum, S. cerevisiae, A. niger and R. oryzae, with MIC values of 16, 32, 32, 64 and 64 µg/mL, respectively | [63] |
| 43 | Alternaria alternata Tche-153 | Terminalia chebula Rezt. Suanluang Rama IX Public Park, Bangkok, Thailand. | Altenusin (120) | Altenusin (120) in combination with each of three azole drugs, ketoconazole, fluconazole or itraconazole at their low sub-inhibitory concentrations exhibited potent synergistic activity against C. albicans with the FIC index range of 0.078 to 0.188 | [64] |
| 44 | Alternaria sp. UFMGCB 55, | Leaves of Trixis vauthieri DC (Asteraceae). | Altenusin (120) | The altenusin (120) exhibited strong activity against 11 strains P. brasiliensis with MIC values ranging between 1.9 and 31.2 µg/mL MIC values found for amphotericin B were between 0.031 and 0.12 µg/mL. Additionally, S. pombe cells treated with altenusin were more rounded in shape than untreated cellssuggeststhat altenusin could act through the inhibition of cell wall synthesis or assembly in P. brasiliensis and S. pombe | [65] |
| 45 | Cladosporium cladosporioides | Cladosporin (121), Isocladosporin (122) | At 30 µM compound 121 exhibited 92.7, 90.1, 95.4, and 79.9% growth inhibition against C. acutatum, C. fragariae, C. gloeosporioides and P. viticola respectively. Compound 122 showed 50.4, 60.2, and 83.0% growth inhibition at 30 µM against C. fragariae, C. gloeosporioides, and P. viticola, respectively | [66] | |
| 46 | Epicoccum sp. CAFTBO, | Theobroma cacao (Sterculiaceae) Mount Kala, near Yaoundé, Centre Province, Republic of Cameroon | Epicolactone (123), Epicoccolide A (124) and B (125) | Compounds 123–125 showed inhibitory effects on the mycelial growth of P. ultimum and A. cochlioides and R. solani (MIC in the range of 20–80 µg per paper disc) | [67] |
| 47 | Biscogniauxiamediterranea Ohu 19B | Opuntia humifusa (Cactaceae) from the United States | 5-methylmellein (100) | Compound 100 5-methylmellein was evaluated for antifungal activity against seven plant pathogens (C. acutatum, C. fragariae, C. gloeosporioides, F. oxysporum, B. cinerea, P. obscurans, and P. viticola) using an in vitro microdilution broth assay. | [68] |
| 48 | Emericella sp. XL029 | Leaves of Panax notoginseng Shijiazhuang, Hebei Province, China. | 5-(undeca-3′,5′,7′-trien-1′-yl)furan-2-ol (126) and 5-(undeca-3′,5′,7′-trien-1′-yl)furan-2-carbonate (127) | Compound 126 active against R. solani, V. dahliae, H. maydis, F. oxysporum, F. tricinctum, B. dothidea, and A. fragriae (MIC values from 25 to 3.1 µg/mL), while compound 127 displayed activity against V. dahliae, H. maydis, F. tricinctum, B. dothidea, and A. fragriae (MIC values from 50 to 12.5 µg/mL) | [69] |
| Comounds Produced by Hyphomycetes | |||||
| 49 | Fusarium fujikuroi (WF5), Penicilium chrysogenum WF6, and P. expensum WF7 | Finger millet Plants Arkell Field Station, Arkell, ON, Canada. | 5-hydroxy 2(3H)-benzofuranone (128), dehydrocostus lactone (129) and harpagoside (130) | Compounds 128–130 active against F. graminearum with MIC of 31.25, 250.00 and 31.25 µg/mL, respectively. | [70] |
| 50 | Trichoderma koningiopsis YIM PH30002 | Panax notoginseng. Wenshan, Yunnan Province, China. | Koninginin O (131), koninginin Q (132), 7-O-methylkoninginin D (133) | Compounds 131–132 active against F. oxysporum and P. cucumerina, with an MIC of 128 µg/mL. Compound 133 showed activity against P. cucumerina with an MIC of 128 µg/mL. Nystatin was active with MICs at 32 µg/mL | [71] |
| 51 | Trichoderma koningiopsis YIM PH30002 | Panax notoginseng. Wenshan, Yunnan Province, China. | Koningiopisin C (134) | Compound 134 exhibited in vitro antifungal activity against F. oxysporum, A. panax, F. solani and P. cucumerina with MICs at 32, 64, 32, and 16 µg/mL, respectively | [72] |
| 52 | Trichoderma sp. 09 | Myoporum bontioides | Dichlorodiaportinolide (135), dichlorodiaportin (136) | Compounds 135–136 active against C. musae and Rhizoctoniasolani (MIC values from 6.25 to 150 µg/mL) | [73] |
| 53 | Trichoderma brevicompactum 0248 | Allium sativum | Trichodermin (137) | Compound 137 active against R. solani, B. cinereal, C. lindemuthianum with an EC50 of 0.25, 2.02 and 25.60 µg/mL respectively. Carbendazim showed, antifungal activity against R. solani, B. cinereal, with an EC50 of 0.36 and 10.35 µg/mL respectively | [74] |
| 54 | Trichoderma koningiopsis YIM PH30002 | Wenshan, Yunnan Province of China. | Koninginin R (138) and S (139) | Compound 138 active against F. oxysporum and F. flocciferum with MICs at 128 µg/mL, while compound 139 displayed activity against F. oxysporum with MIC at 128 µg/mL | [75] |
| 55 | Aspergillus terreus | Carthamus lanatus Al-Azhar University campus, Assiut Branch, Assiut, Egypt. | (22E,24R)-stigmasta-5,7,22-trien-3-β-ol (140), aspernolides F (141) | Compound 140 active against C. neoformans with IC50 values of 4.38 µg/mL, amphotericin B (IC50 0.34 µg/mL). Compound 141 showed good activity against C. neoformans (IC50 5.19 µg/mL). | [76] |
| 56 | Aspergillus sp. KJ-9, | Melia azedarach which was collected at Yangling, Shaanxi Province, China | Fonsecinone A (142), (R)-3-hydroxybutanonitrile (143) | Compounds 142 and 143 were active against G. saubinetti, M. grisea, B. cinerea, C. gloeosporioides and A. solani with MIC range of 6.25–50 µM | [77] |
| 57 | Aspergillus sp. | Gloriosa superba Tirupati, India. | 6-methyl-1,2,3-trihydroxy-7,8-cyclohepta-9,12-diene-11-one-5,6,7,8-tetralene-7-acetamide (KL-4) (144) | KL-4 (144) active against S. cerevisiae, C. albicans and C. gastricus with MIC 25, 12.5, and 50 µg/mL respectively | [78] |
| 58 | Penicillium sp. R22 | Nerium indicum collected from Qinling Mountain, Shaanxi Province, China. | 5-hydroxy-8-methoxy-4-phenylisoquinolin-1(2H)-one (145), 3-O-methylviridicatin (146) and viridicatol (147) | Compound 145 active against A. brassicae, A. alternata and V. mali with MIC value of 31.2 µg/mL, compound 146 against A. brassicae, B. cinerea and V. male with MIC value of 31.2 µg/mL, compound 147 against A. brassicae, A. alternata and B. cinerea with MIC value of 31.2 µg/mL | [79] |
| 59 | Penicillium raciborskii, | Rhododendron tomentosum were collected at the test site of University of Oulu, Finland. | Outovirin C (148) | Outovirin C (148) inhibited growth of F. oxysporum, B. cinerea, and V. dahlia at the concentration of 0.38 µM. Compound 148 active against B. cinerea (57% inhibition) and slightly less effective against V. dahliae (45% inhibition) | [80] |
| 60 | Fusarium sp. | Mentha longifolia Saudi Arabia. | Fusaripeptide A (149) | Compound 149 active against C. albicans, C. glabrata, C. krusei, and A. fumigates with IC50 values of 0.11, 0.24, 0.19, and 0.14 µM, respectively. Amphotericin B exhibited antifungal activity toward C. albicans, C. glabrata, C. krusei, and A. fumigates with IC50 values of 0.3, 0.6, 0.5, 0.7 µM, respectively | [81] |
| 61 | Fusarium chlamydosporium | Anvillea garcinii Al Madinah Al Munawwarah, Saudi Arabia. | Fusarithioamide A (150) | Compound 150 active with inhibition zone diameters 16.2 mm and MIC 2.6 µg/mL towards C. albicans. Clotrimazole (inhibition zone diameters 18.5 mm and MIC 3.7 µg/mL) | [82] |
| 62 | Fusarium sp. | Ficus carica Qinling Mountain, Shaanxi Province, China | Helvolic acid Methyl ester (151), helvolic acid (152) and hydrohelvolic acid (153) | Compounds 151–153 active against B. cinerea, C. gloeosporioides, F. oxysporum f. sp. niveum, F. graminearum and P. capsici (MIC in the range of 12.5–25 µg/mL), Carbendazim (MIC in the range of 32.2–62.5 µg/mL) | [83] |
| 63 | Fusarium sp. | Colletorin B (154), colletochlorin B (155), LL-Z1272β (llicicolin B) (156) and 4,5-dihydrodechloroascochlorin (157) | Compounds 154–156 showed antifungal against U. violacea and F. oxysporum. Compound 157 showed antifungal activity towards E. repens, | Hussain et al. [84] | |
| 64 | Curvularia sp., strain M12, | Murraya koenigii Rajshahi University, Bangladesh | Murranolide A (158), murranopyrone (159), Curvularin (160), (S)-dehydrocurvularin (161), pyrenolide A (162), modiolide A (163), and 8-hydroxy-6-methoxy-3-methylisocoumarin (164) | Pyrenolide A (162) showed a strong motility impairing activity against Phytophthora capsici zoospores at a low concentration (100% at 0.5 µg/mL) in a short time (30 min). Compounds 158–161 and 163–164 exhibited zoospore motility impairment activity at higher concentrations (IC50: 50–100 µg/mL) | [85] |
| 65 | Phaeoacremonium sp., | Senna spectabilis AraraquaraCerrado area, Araraquara, Sao Paulo state, Brazil. | Isoaigialone B (165), and C (166), aigialone (167) | Compounds 165 and 167 exhibited antifungal activity, with a detection limit of 5 µg, for C. cladosporioides and C. sphaerospermum, compound 166 exhibited weak activity (detection limit > 5 µg), with a detection limit of 25 µg. Nystatin, positive control, showing a detection limit of 1 µg | [86] |
| 66 | Trichothecium sp. | Phyllanthus amarus Pune India. | Trichothecinol A (168) | Compound 168 active against C. albidus up to 20 µg/mL | [87] |
| 67 | Trichothecium sp. | Phyllanthus sp. Pune India. | Trichothecin (169) | Trichothecin (169) active against S. cerevisiae, C. albidus var diffluens (NCIM 3371), C. albidus var diffluens (NCIM 3372), F. oxysporum, P. expansum, T. viride, P. varioti and A. niger with MIC of 6.0, 20.0, 12.0, 10.0, 30.0, 40.0, 20.0 and 12.0 µg/mL respectively | [88] |
| Comounds Produced by Basidiomycetes | |||||
| 68 | Scleroderma UFSM Sc1(Persoon) Fries | Eucalyptus grandis. | Sclerodol A (170) and B (171) and related lanostane triterpenoid (172) | Compound 170 active against C. albicans, C. tropicalis, C. crusei, C. parapsiosis (MIC of 25.0, 25.0, 6.25 and 12.5 MFC 25.0 25.0, 12.5, 25.0 µg/mL) Compound 170 and 172 were active against tested strain (MIC in range of 12.5–100 µg/mL). Nystatin active against test strains (MIC in the range of 0.77–1.52 µg/mL). | [89] |
Table 2.
Spectrum of VOCs emitted by Muscodor species and their anti-fungal activity predominantly against plant pathogenic fungi.
Table 2.
Spectrum of VOCs emitted by Muscodor species and their anti-fungal activity predominantly against plant pathogenic fungi.
| No. | Name of the Endophytic Fungi | Geographic Area of Isolation | Major VOCs Produced | Anti-Fungal Activity | Refs. |
|---|---|---|---|---|---|
| 1 | Muscodor albus | Central America (Honduras) | 2-methylpropanoic acid; 3-methyl-1-butanol; ethanol; acetic acid (methyl ester) | Rhizoctonia solani; Phytophthora cinnamomi; Sclerotinia sclerotiorum; Fusarium solani, Verticillium dahliae | [91] |
| 2 | M. vitigenus | South America (Peru) | naphthalene; caryophyllene; azulene | R. solani; Phoma sp.; C. coefficola | [98] |
| 3 | M. roseus | Australia | 2-butenoic acid (ethyl ester); 1,2,4-tri-methyl-benzene; 2-nonadiene | Antifungal spectrum not reported | [101] |
| 4 | M. yucatensis | South America (Mexico) | caryophyllene; aromadendrene | Botrytis cineria; R. solani; C. coefficola; Phoma sp. | [102] |
| 5 | M. fengyangensis | China | 2-methylpropionic acid; β-phellendrene | B. cineria; Aspergillus clavatus; Colletotrichum fragiae; Sclerotium rolfsii | [90] |
| 6 | M. crispans | South America (Bolivia) | 2-methylpropanoic acid; ethanol; ethyl acetate | B. cineria; Curvularia lunata; P. cinnamomi; S. sclerotiorum | [103] |
| 7 | M. sutura | USA (Columbia) | butylated hydroxytoleuene; octacecanoic acid; thujopsene; 2-methylpropanoic acid; naphthalene | Aspergillus fumigatus; Colletotrichum lagenarium; B. cineria; Cercospora beticola; Phytophthora palmivora; Fusarium solani | [104] |
| 8 | M. musae | Thailand | 3-methylbutanol acetate 2-methylpropanoic acid | Alternaria porri; Alternaria solani; Colletotrichum gloereosporioides; Nigrospora oryzae | [105] |
| 9 | M. oryzae | Thailand | 3-methylbutan-1-ol; 2-methylpropanoic acid | A. porri; A. solani; Aspergillus flavus; B. cineria; C. gloereosporioides; N. oryzae | [105] |
| 10 | M. suthepensis | Thailand | 3-methylpropanoic acid 3-methylbutan-1-ol | A. porri; Alternaria alternata; Aspergillus flavus; B. cineria; C. gloereosporioides; Fusarium oxysporum; Fusarium solani; N. oryzae | [105] |
| 11 | M. equiseti | Thailand | 3-methylbutan-1-ol; 3-methylbutanoyl acetate; 2-methylpropanoic acid | A. porri; A. solani; B. cineria; C. gloereosporioides; F. oxysporum; F. solani; N. oryzae | [105] |
| 12 | M. cinnamomi | Thailand | 2-methylpropanoic acid;2-methyl butanoic acid; azulene | Rhizoctonia solani | [105] |
| 13 | M. kashayum | India | 1-methyl-4(1-methylethhylidene)-cyclohexane; 2(4-morpholinyl)ethylamine; 9-octadecanoic acid (methyl ester); 4-octadecylmorpholine | Bionectria ochroleuca; Cercospora beticola; Chaetomium heterosporum; C. gloereosporioides; F. oxysporum; Fusarium equiseti; Curvularia lunata | [106] |
| 14 | M. darjeelingensis | India | 4-octadecylmorpholine; 2,6-bis(1,1-dimethylethyl)-4-(1-oxopropyl)phenol; beta-aminoethyl-morpholine | Lasiodiplodia theobromae; A. alternata; Rhizoctonia solani; Cercospora beticola | [107] |
| 15 | M. strobelii | India | 4-octadecylmorpholine; tetraoxapropellan; aspidofractanine-3-methanol;viridiflorol | Rhizoctonia solani; Colletotrichum gloereosporioides; Fusarium oxysporum; Lasiodiplodia theobromae | [108] |
| 16 | M. tigrii | India | 4-octadecylmorpholine; 1-tetradecamine n, n-dimethyl 1,2-benzidicarboxylic acid mono(2-ethylhexyl)ester | Alternaria alternate Cercospora beticola | [109] |
| 17 | M. heavae | Thailand | 2-phenylethanol; azulene | Aspergillus niger; Phellinus noxius; Rigidoporus microporus | [110] |
| 18 | M. ghoomensis | India | n,n-dimethyl-1-nonadecamine; 4-octadecylmorpholine | Cercospora beticola | [111] |
| 19 | M. indica | India | n, n-dimethyl-1-pentadecamine; 4-morpholinethanamine | Cercospora beticola Penicillium marnaeffi | [111] |
| 20 | M. camphora | India | tetracontane; 4-octadecylmorpholine; n, n-dimethyl-1-pentadecamine | Colletotrichum gloereosporioides; Lasiodiplodia theobromae | [112] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Deshmukh, S.K.; Gupta, M.K.; Prakash, V.; Saxena, S. Endophytic Fungi: A Source of Potential Antifungal Compounds. J. Fungi 2018, 4, 77. https://doi.org/10.3390/jof4030077
AMA Style
Deshmukh SK, Gupta MK, Prakash V, Saxena S. Endophytic Fungi: A Source of Potential Antifungal Compounds. Journal of Fungi. 2018; 4(3):77. https://doi.org/10.3390/jof4030077
Chicago/Turabian StyleDeshmukh, Sunil K., Manish K. Gupta, Ved Prakash, and Sanjai Saxena. 2018. "Endophytic Fungi: A Source of Potential Antifungal Compounds" Journal of Fungi 4, no. 3: 77. https://doi.org/10.3390/jof4030077
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.