Efficient Transduction and Expansion of Ovine Macrophages for Gene Therapy Implementations



1
Laboratory of Microbiology and Infectious Diseases, Faculty of Veterinary Medicine, Aristotle University of Thessaloniki, Thessaloniki 54124, Greece
2
Laboratory of Biology, School of Medicine, National and Kapodistrian University of Athens, Athens 11527, Greece
*
Authors to whom correspondence should be addressed.
Vet. Sci. 2018, 5(2), 57; https://doi.org/10.3390/vetsci5020057
Submission received: 6 May 2018
/
Revised: 5 June 2018
/
Accepted: 14 June 2018
/
Published: 18 June 2018
Abstract
:A number of bacteria provoking zoonotic diseases present intracellular survival and a host cell tropism limited to the monocyte/macrophage lineage. Thus, infection is rendered difficult to eradicate, causing chronic inflammatory reactions to the host and widespread prevalence. Although self-inactivating lentiviral vectors have been successfully tested in the clinic against virally-induced human infectious diseases, little is known about the transduction susceptibility of ruminant animal phagocytes that play a critical role in the outbreak of zoonotic diseases such as brucellosis. In view of the development of a lentiviral vector-based platform targeting and inactivating specific genetic features of intracellular bacteria, we have tested the transducibility of ovine macrophages in terms of transgene expression and vector copy number (VCN). We show that ovine macrophages are relatively resistant to transduction even at a high multiplicity of infection with a conventional lentiviral vector expressing the green fluorescence protein and that addition of transduction enhancers, such as polybrene, increases transgene expression even after a one-week culture of the transduced cells in vitro. Overall, we demonstrate that ovine macrophages may be efficiently expanded and transduced in culture, thus providing the benchmark for gene therapy applications for zoonotic diseases.
1. Introduction
Bacterial infections in livestock not only pose severe public health issues, but also translate into a substantial financial burden for stockbreeders, due to reduction of the quality and quantity of the downstream products. Especially when the microorganism is able to survive intracellularly and evade the antibiotics’ effect, therapy efforts towards infected animals is cost ineffective and has an ambiguous benefit. Moreover, no effective vaccine currently exists for preventing such diseases in humans, while antibiotic therapy is long-lasting and often extends beyond six months. In addition, treatment in humans does not exclude relapse or re-infection after de novo exposure to the microorganism [1].
An illustrative example are bacteria of the genus Brucella that present a host cell tropism limited to the monocyte/macrophage lineage. Although immunized animals destroy Brucella via cellular immunity mechanisms [1], vaccination of herds, as well as their supervision is often ineffective, as the vaccine may lead to abortions and infertility in animals [2]. In addition, since the vaccine partially preserves its virulent properties, it is not entirely safe for clinical practitioners who administer it [2]. Moreover, the fact that the wildtype strain in seropositive animals is not distinct from the strain used in the vaccine, along with the long-term immune reaction that the vaccine may provoke in certain instances, renders the discrimination between vaccinated and infected animals impossible [3].
Consequently, the necessity of massive sanitation of ruminant livestock is crucial and indispensable. The scientific gap described above could possibly be overcome by gene therapy, a novel therapeutic strategy that utilizes non-pathogenic viral vectors to deliver normal genes into malfunctioning cells, in order to correct the pathological phenotype of genetic diseases. Gene therapy may be achieved with two different approaches [4]. In the first approach, the target stem cells are collected, co-cultured in the presence of a non-pathogenic viral vector to allow gene transfer to take place and are subsequently re-infused as a phenotypically functional graft back into the same patient from whom they were initially harvested. In the second case, gene correction takes place in the patient’s body, via systemic administration of the vector and is usually implemented in tissues in which stem cells cannot be readily characterized or isolated (e.g., lungs, brain, liver, etc.).
Over the last fifteen years, gene therapy has been successfully applied in clinical trials for monogenic diseases [5,6,7,8,9,10], providing a potentially lifelong therapeutic benefit to patients. Although gene therapy has also been successfully implemented at a clinical level in the field of infectious diseases, against hepatitis B virus (HBV), aiding in its clearance [11], and against human immunodeficiency virus-1 (HIV-1) [12], by inactivating the expression and replication of its genes, its effectiveness remains to be further evaluated.
In order to facilitate massive sanitation of ruminant animal livestock by gene therapy, the development of a lentiviral vector-based platform targeting and inactivating specific genetic features of intracellular bacteria is considered essential. However, although the feasibility of gene transfer in human [13,14] or rodent animal [15] macrophages has been previously proven, little is known about the effectiveness of transduction in ruminant animal macrophages, for example of ovine origin. In the recent past, sheep have been effectively used as a model for plenty of gene transfer implementations targeting the nervous system [16,17], the respiratory tract [18], the eyes [19] or even for in utero gene therapy applications [20]. Moreover, lentiviral vectors have been extensively utilized to transduce small ruminant macrophages, but mostly in order to elucidate the immune responses against viral infections [21,22,23], rather than investigate the outcome of gene transfer itself.
For the first time here, we analyzed the transgene expression and vector copy number (VCN) incorporated in the target cell genome, after exposure of peripheral blood-derived ovine macrophages to a conventional vesicular stomatitis virus glycoprotein (VSV-G)-pseudotyped lentiviral vector expressing the green fluorescence protein (GFP). We show that by the addition of transduction enhancers, such as polybrene or protamine sulfate, transgene expression may be further increased to high levels and that GFP expression may be sustained, under certain conditions, even after a one-week culture of the transduced cells in vitro. Overall, our findings demonstrate the feasibility of transduction in terms of cells of the monocyte/macrophage lineage of ovine origin and that it may prove useful for any gene therapy application against zoonotic diseases in the future.
2. Materials and Methods
2.1. Isolation of Ovine Monocytes
Approximately 25 mL of peripheral blood were collected from healthy sheep in controlled herds into ethylenediaminetetraacetic acid (EDTA)-anticoagulant vacutainers (BD, Franklin Lakes, NJ, USA) and tested for sterility after seeding in blood/agar dishes (Oxoid, Hampshire, UK). Complete blood counts (CBC) were also performed to ensure animals’ health status. The mononuclear cell fraction containing monocytes was separated through a Biocoll (Merck, Berlin, Germany) density gradient centrifugation at a ratio of 1 blood to 0.75 ficoll. Mononuclear cells were isolated, washed once with phosphate-buffered saline (PBS) (Lonza, Basel, Switzerland) and seeded in macrophage serum-free medium (Macrophage-SFM, Invitrogen, Carlsbad, CA, USA) supplemented with heat-inactivated fetal bovine serum (FBS) (Invitrogen, Carlsbad, CA, USA) as previously described [24]. Three weeks post culture, adherent cells were harvested and checked for CD14 expression by flow cytometry (BD FACSCalibur, Franklin Lakes, NJ, USA), after staining with the anti-bovine CD14 monoclonal antibody, conjugated with fluorescein isothiocyanate (FITC) (Bio-Rad, Hercules, CA, USA). An FITC-conjugated mouse IgG1 antibody was used as an isotype control (BD, Biosciences, San Jose, CA, USA). Acquisition data were analyzed using the CellQuest Pro v6 software (BD Biosciences, San Jose, CA, USA) or the Cyflogic software (www.cyflogic.com, CyFlo Ltd., Turku, Finland).
2.2. Macrophage Phagocytosis Assay
To determine the macrophages’ phagocytic activity, cells were exposed to green fluorescent polystyrene microspheres (Flow Cytometry Sub-micron Particle Size Reference Kit, Invitrogen, Carlsbad, CA, USA), each with a known diameter (0.2 μm, 0.5 μm, 1 μm and 2 μm), that show an excitation and emission profile similar to FITC and are thus collected by the FL-1 channel of the cytometer. The beads were added to approximately 105 macrophages at a 1:1 ratio for 24 h. In order to quantify the phagocytosis events, the cells were washed 5–6 times with PBS, detached from the culture dishes with a cell scraper and analyzed by flow cytometry (BD FACSCalibur) using the CellQuest Pro v6 software (BD Biosciences, San Jose, CA, USA) or the Cyflogic software (www.cyflogic.com).
2.3. Lentiviral Vector Production and Titration
The vector used in this study was a self-inactivating (SIN) lentiviral vector that encodes GFP under the human phosphoglycerate kinase (PGK) promoter and also contains the 1.2-kb cHS4 insulator incorporated in the deleted region of U3. Details regarding the vector construct have been previously published [25]. Vector production and titration were performed as previously described [26]. Briefly, research-grade, VSV-G-pseudotyped lentiviral stocks of the GFP vector were generated by transfection of 293T cells with calcium phosphate precipitation using standard procedures. The final viral stocks were concentrated 100-fold by ultrafiltration. Serial dilutions of the concentrated viral stock were used to infect mouse erythroleukemia cells (MEL-585) for 3–4 days, and determination of the viral titer was documented by analyzing the cells for GFP expression by flow cytometry (BD FACSCalibur). Acquisition data were analyzed using the CellQuest Pro v6 software or the Cyflogic software (www.cyflogic.com).
2.4. Macrophage Transduction
Ovine macrophages were seeded in 24-well tissue culture plates at a concentration of ~0.1 × 106 cells/mL and were exposed, 24 h after initial seeding, to serial dilutions of the concentrated GFP vector stock corresponding to a multiplicity of infection (MOI) of 60, 6 and 0.6. To enhance transduction, 22 μg/mL hexadimethrine bromide (polybrene, Sigma-Adrich, St. Louis, MO, USA) or 37 μg/mL protamine sulfate (Sigma-Adrich, St. Louis, MO, USA) were added to the cells upon vector exposure. Two days later, cells were split and maintained in culture for another week. Transduction efficiency was assessed by monitoring the expression of GFP with flow cytometry, and vector-specific primers were utilized to determine the VCN per cell by quantitative real-time PCR. Mock-transduced cells and cells transduced in the absence of polybrene or protamine sulfate served as controls at all times.
2.5. Vector Copy Number Analysis
Genomic DNA from cultured cells was extracted using the Nucleospin Tissue kit (Macherey-Nagel, Düren, Germany) as per the manufacturer’s directions. Quantification of the GFP lentiviral VCN was performed by real-time PCR based on SYBR green using vector-specific primers against the Rev response element (RRE) as previously described [26]. Average VCN per cell was calculated by normalizing to the endogenous ovine prion protein (Prp) single-copy chromosomal gene [27,28], in order to adjust for equal loading of genomic DNA per reaction, by using the following primers: ovPrpF 5’-GCCAAAAACCAACATGAAGCAT-3’ and ovPrpR 5’-TGCTCATGGCACTTCCCAG-3’.
2.6. Statistics
Multiple comparisons were performed using one-way ANOVA. Values of p < 0.05 were considered statistically significant. All results are expressed as the means ± the standard deviation (SD).
3. Results
3.1. Characterization of Peripheral Blood-Derived Ovine Macrophages
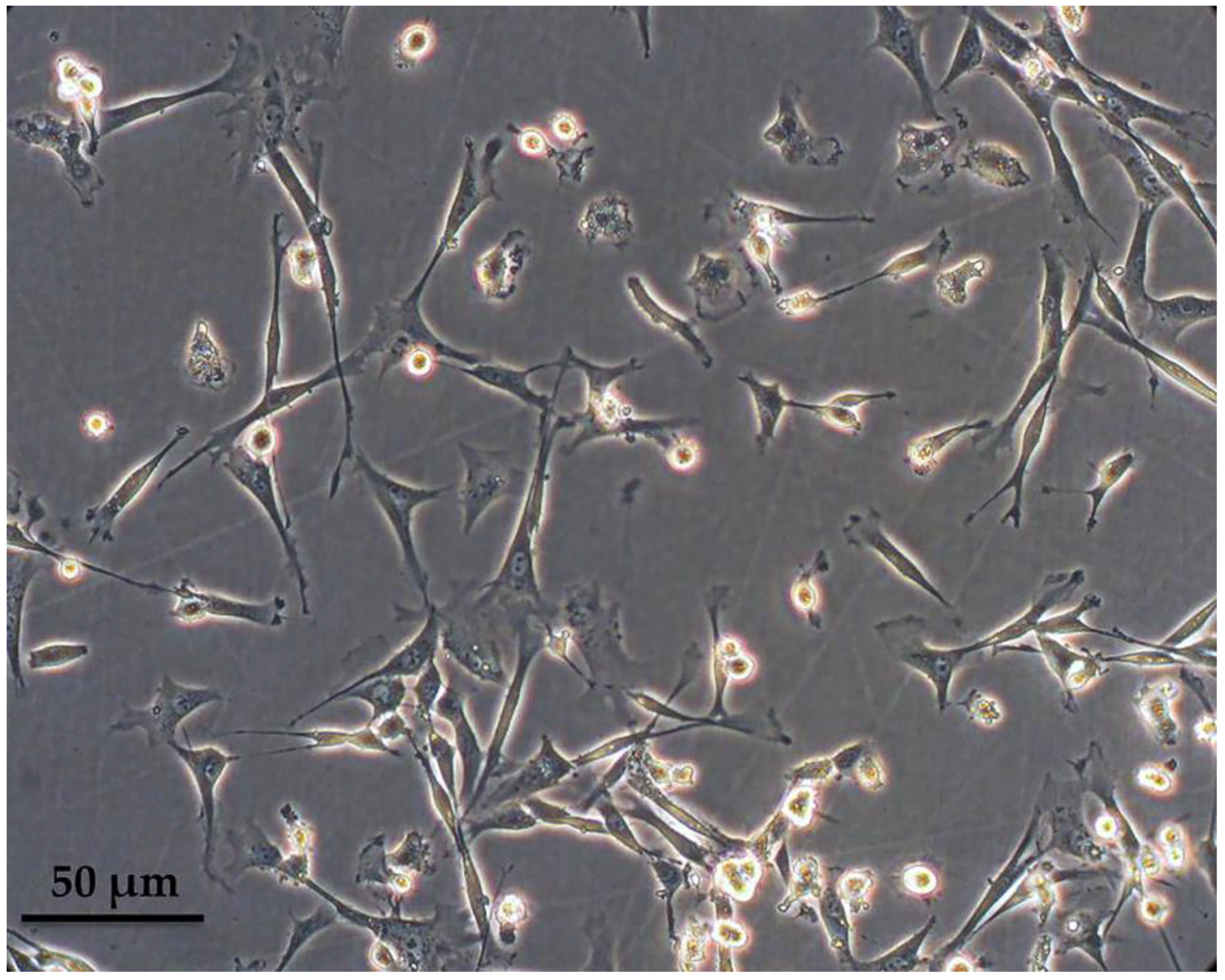
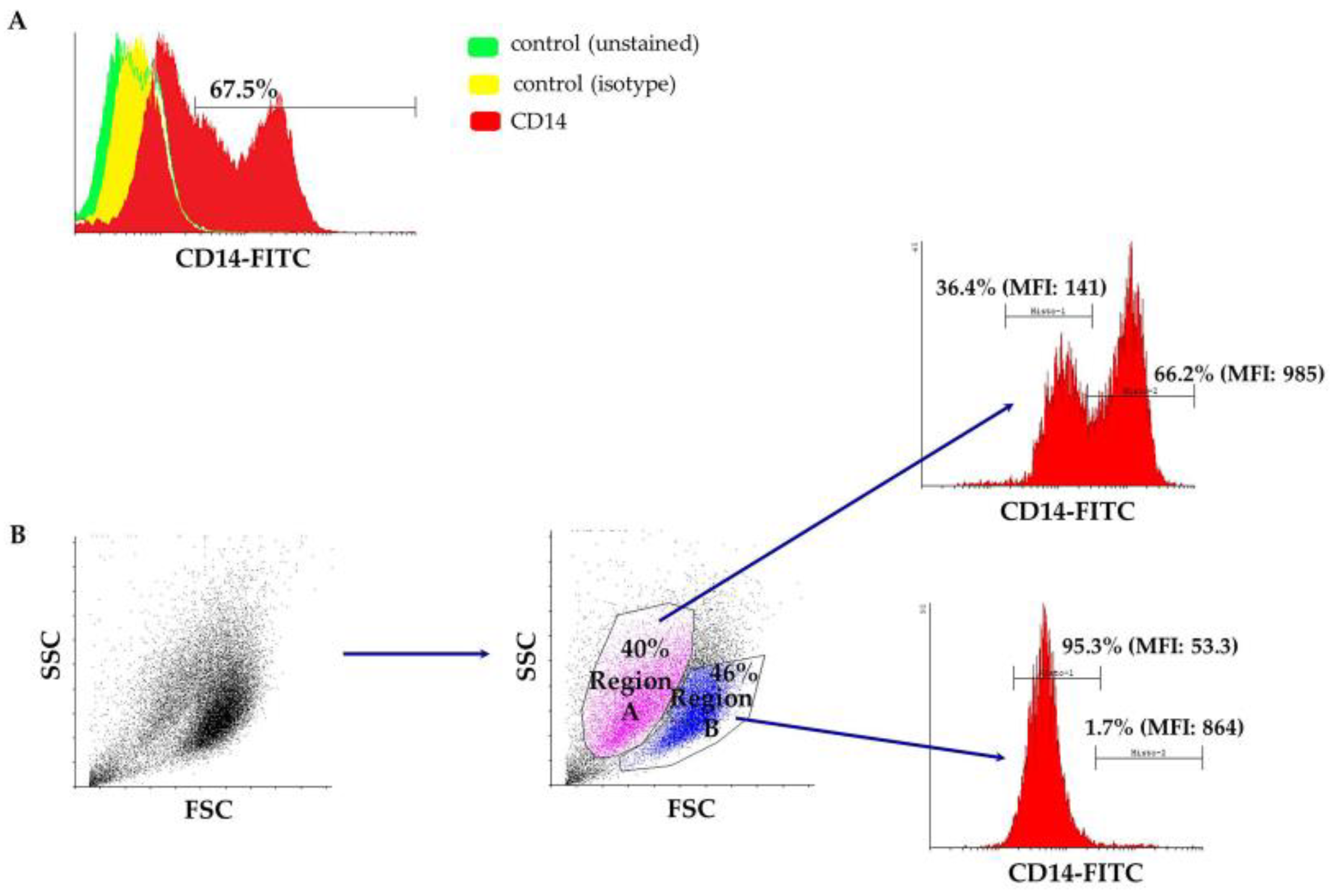
Peripheral blood-derived mononuclear cells were isolated from the buffy coat after a ficoll density gradient centrifugation and cultured in macrophage-specialized media for three weeks, in order to allow for adequate maturation of ovine monocytes (Figure 1). Macrophages were phenotypically identified after staining with anti-CD14 and subsequent flow cytometry analysis and a mean of 67.5% of the population was found to express CD14 (ranging from 55–80%), as shown in Figure 2A from one representative experiment (n = 4). Interestingly, the culture was comprised of two, proportionally semi-equal cell populations with distinct size and granularity that also expressed the CD14 antigen to a different extent. More specifically, as indicated in Figure 2B, in the forward scatter/side scatter (FSC/SSC) dot plot, we consistently observed two well-defined populations: one population containing smaller and more complex cells, that we classified as Region A, comprising approximately 40% of the entire population, and a second population that was comprised of 46% of the cells, consisting of larger and less complex cells, classified as Region B. The vast majority of cells within Region B, i.e., 95.3%, expressed low levels of CD14 (CD14low) as evidenced by the mean fluorescent intensity of the CD14+ population that was on average 53.3 (Figure 2B). On the contrary, the larger-sized cohort, i.e., Region A, mostly was comprised of CD14high cells (Figure 2B) since 66.2% of the cells were expressing high levels of CD14 (MFI: 985). To summarize, under the specified experimental conditions, we observed two different and distinct cell populations in terms of size and granularity. Larger and less complex cells (Region B) uniformly expressed low levels of CD14, whereas smaller and more complex cells (Region A) expressed both high levels of CD14 (66.2% of the cells in Region A, MFI: 985) and low levels of CD14 (36.4% of the cells in Region A, MFI: 141).
3.2. Phagocytosis Assay
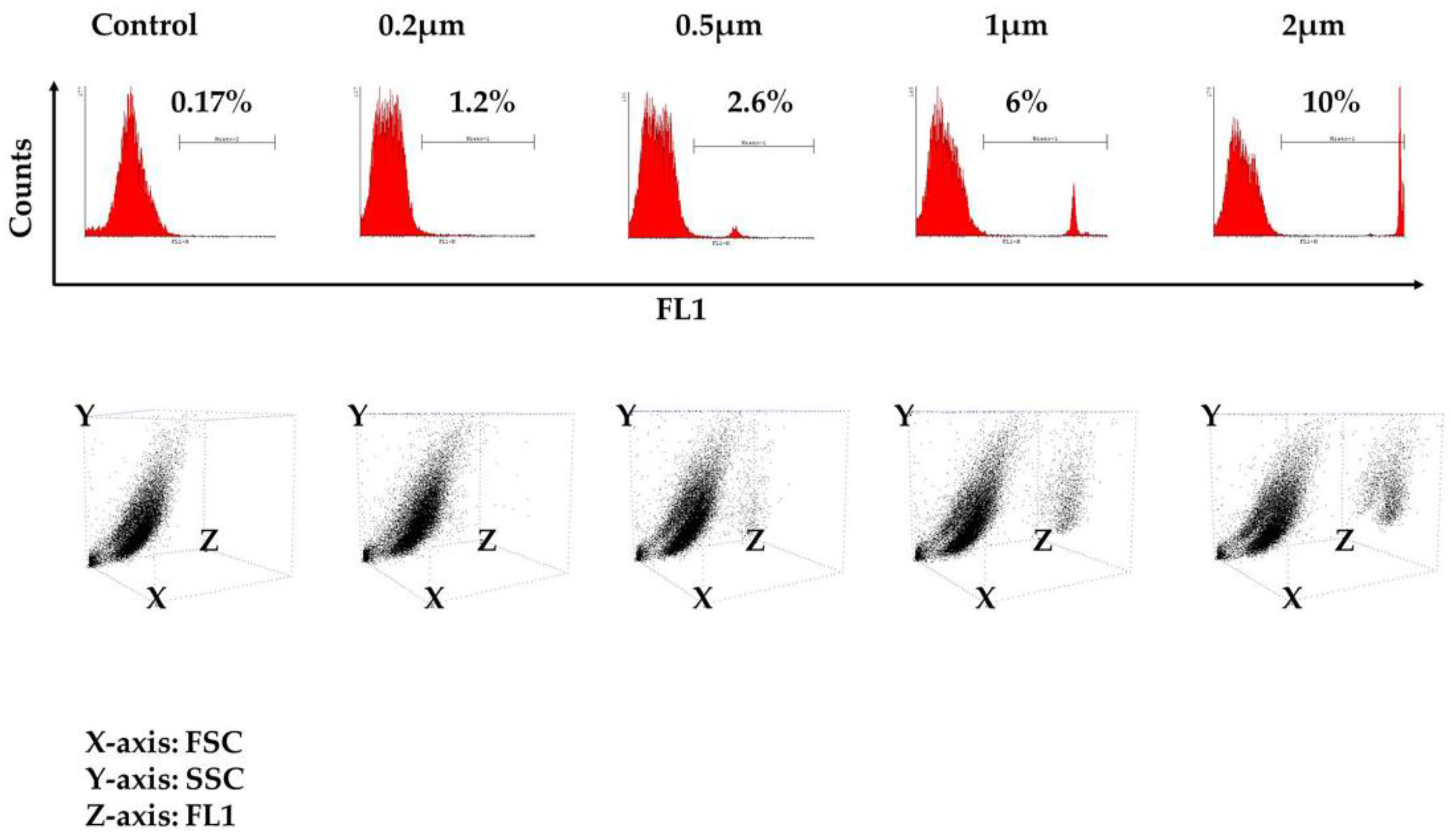
The kinetics of phagocytosis at the single-cell level was tested after exposure of the cells to variously-sized fluorescent beads ranging from 0.2 μm–2 μm in diameter that, upon engulfment, could be detected in the phagocytes’ cytoplasm with flow cytometry. The beads were added at a ratio of 1:1 (beads/cells), and as shown in Figure 3 from one representative experiment, 24 h after exposure to the beads, a certain percentage of the cells presented phagocytic properties, which proportionally increased with the diameter of the beads, as we noticed a higher percentage of phagocytosis when the 2-μm beads were utilized. A detailed description of the percentage of FITC-positive cells from each individual experiment is shown in Table 1 (control: 0.13% ± 0.04; 0.2 μm: 1.12% ± 0.11; 0.5 μm: 2.16% ± 0.45; 1 μm: 5.12% ± 1.32; 2 μm: 9.16% ± 0.74; number of experiments = 3). These findings confirmed that the cultured population is able to engulf beads of dimensions close to those found in bacteria, such as the 2-μm diameter, with greater efficiency. The 3D dot plot that is depicted in Figure 3 demonstrates that the ability to engulf is present in both populations of Region A and Region B.
3.3. Ovine Macrophages’ Resistance to Transduction May Be Averted with Transduction Enhancers
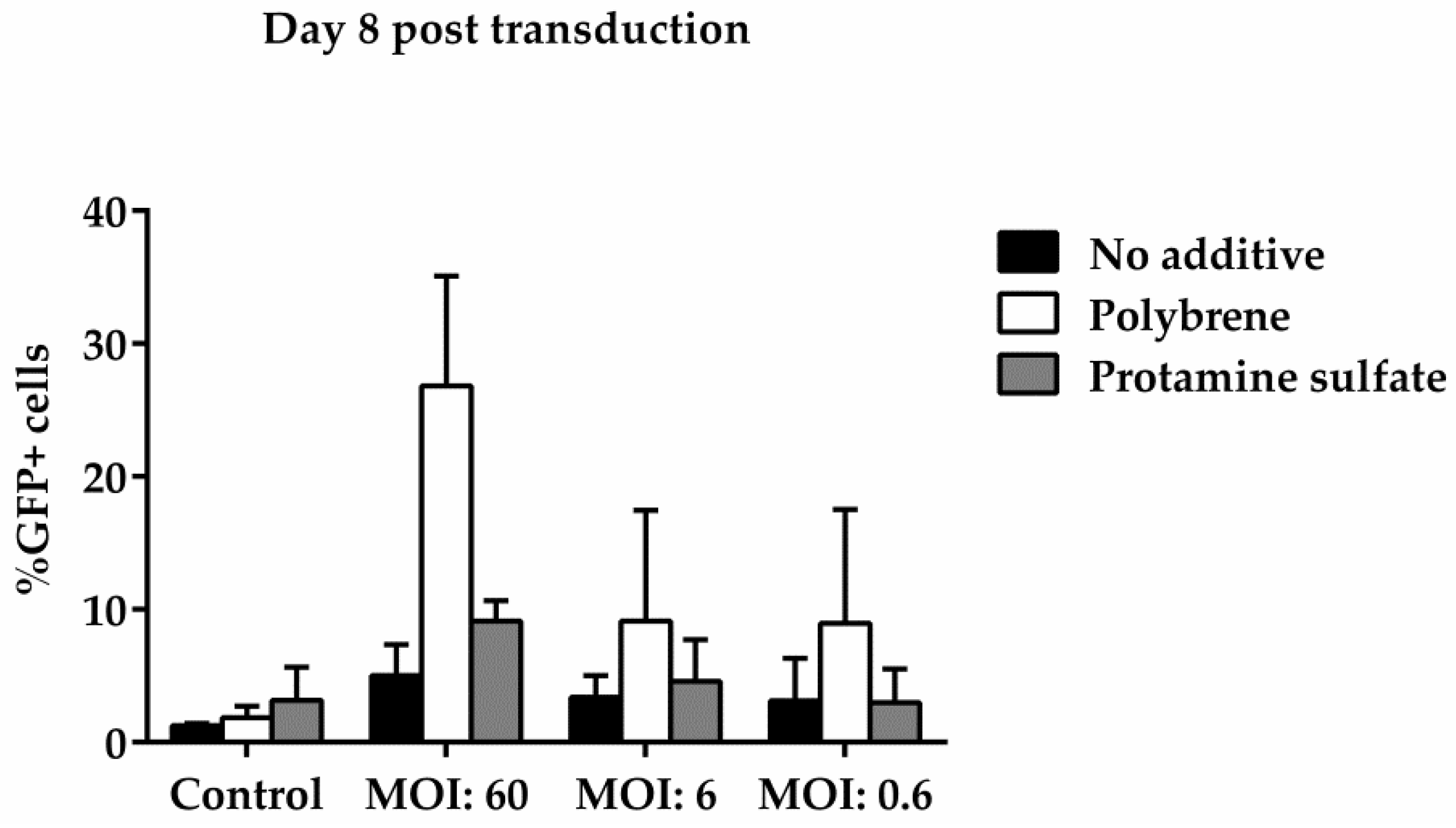
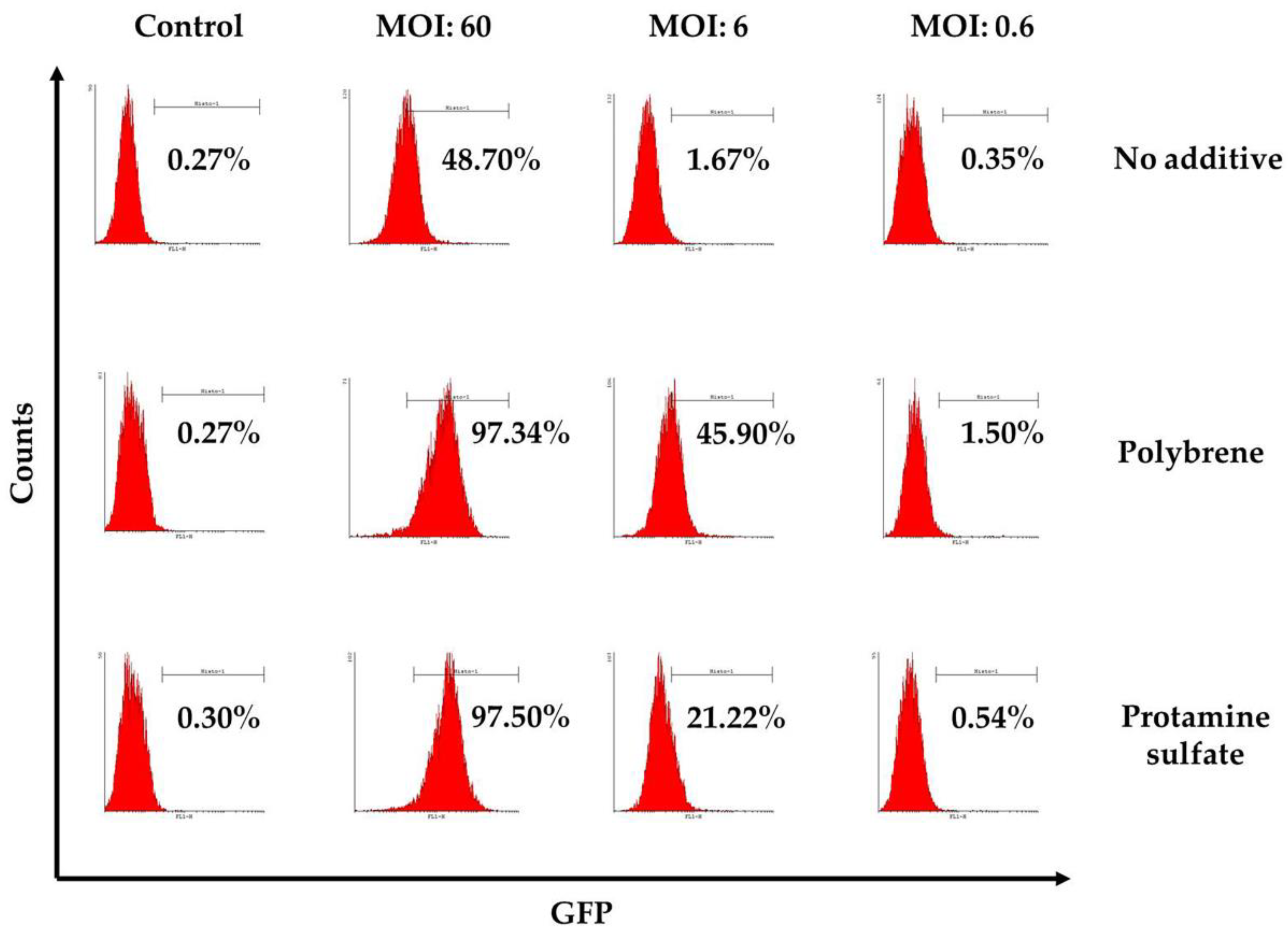
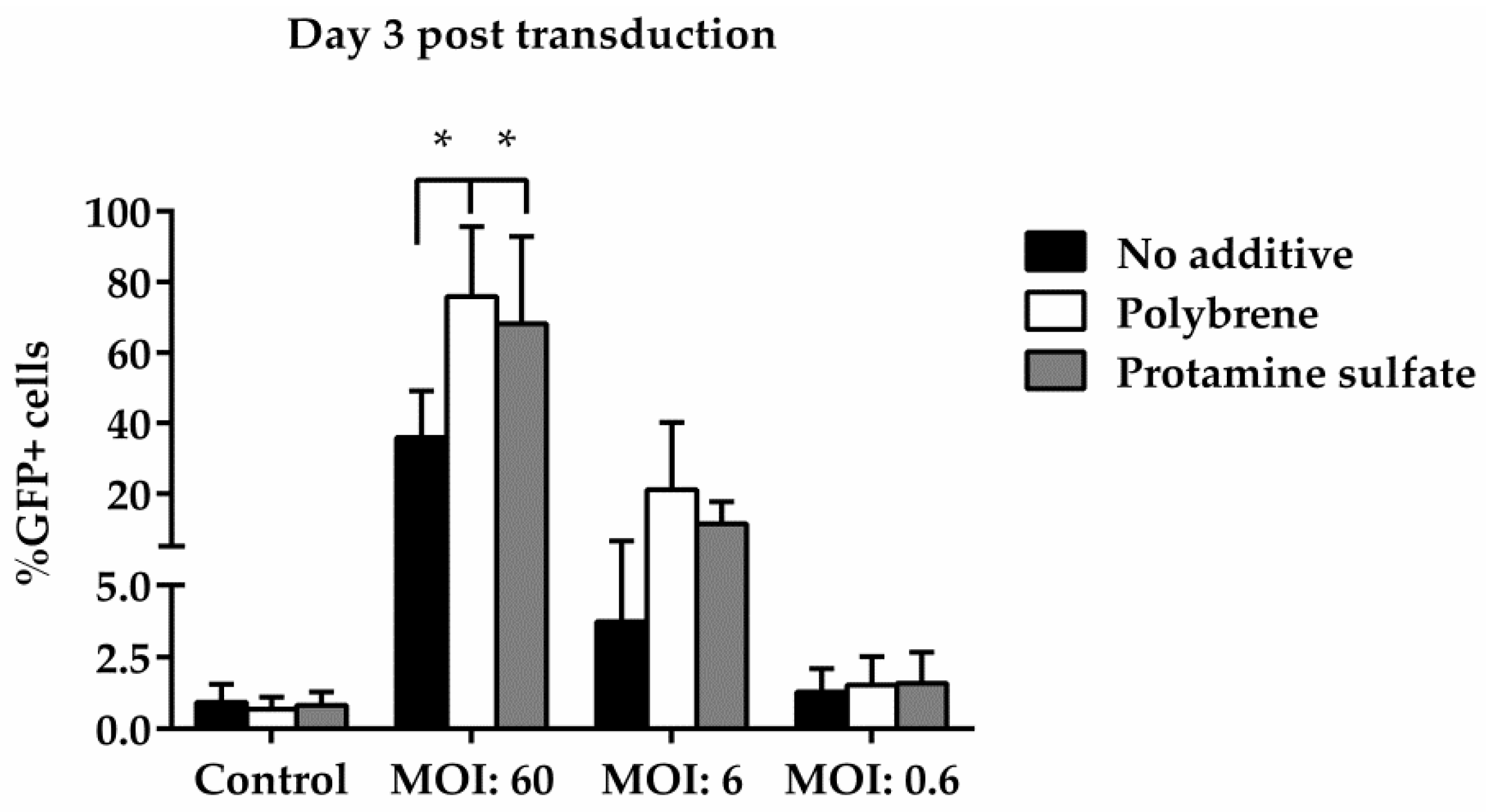
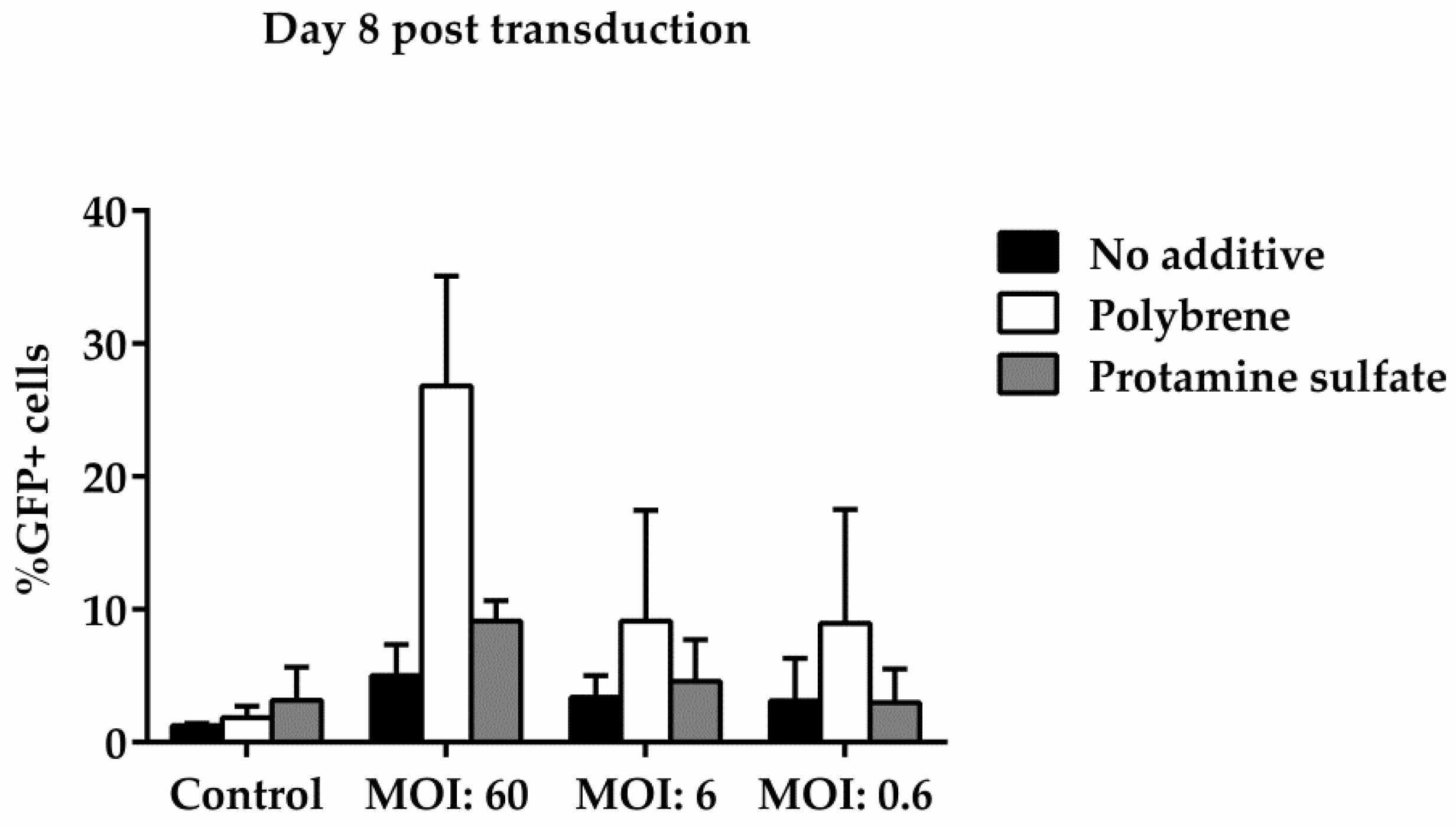
To examine ovine macrophages’ gene transfer efficiency, we exposed the cells to various concentrations of a GFP lentiviral vector and measured the expression of the green fluorescence protein, three and eight days post transduction. Our results demonstrate that even at the highest MOI of 60, ovine macrophages present a moderate transgene expression; a barrier, which, however, could be overcome by the addition of transduction enhancers, such as polybrene or protamine sulfate, at the time of vector exposure, as indicated in Figure 4 from one representative experiment (number of experiments = 5). A detailed analysis of the GFP-positive cells that were detected three days after transduction with the lentiviral vector is shown in Table 2 (no additive: control = 0.84% ± 0.65, MOI 60 = 38.6% ± 12.7, MOI 6 = 3.4% ± 3.03, MOI 0.6 = 1.12% ± 0.8; polybrene: control = 0.57% ± 0.38, MOI 60 = 79.9% ± 19, MOI 6 = 24.3% ± 19.3, MOI 0.6 = 1.14% ± 0.32; protamine sulfate: control = 0.74% ± 0.47, MOI 60 = 73.4% ± 23.5, MOI 6 = 12.6% ± 6.2, MOI 0.6 = 1.31% ± 0.99). Although both transduction enhancers showed a significant increase of gene transfer efficiency at the highest MOI on Day 3 (polybrene vs. no additive: 79.9% ± 19 vs. 38.6% ± 12.7, p = 0.01 and protamine sulfate vs. no additive: 73.4% ± 23.5 vs. 38.6% ± 12.7, p = 0.01) (Figure 5), only cells treated with polybrene displayed an increased GFP expression even at Day 8 post transduction, as compared with their no additive and protamine sulfate counterparts (Figure 6), demonstrating sustained transgene expression levels.
To complete the analysis, we calculated the vector copy number/cell (VCN) by real-time PCR. Our results showed that there was a dramatic decrease in VCN throughout the period of cultivation at the high MOI (MOI = 60). More specifically, although at Day 3, all experimental groups displayed high levels of gene marking, which was statistically significant particularly at the high MOI in comparison with the rest of the MOIs, we observed a severe decrease in VCN, at Day 8 post transduction (Table 3). With the exception of polybrene, which showed a VCN of two for MOI = 6 for Day 3 (data not shown), all of the MOIs and conditions failed to confer adequate gene marking eight days after transduction. This suggests that ovine macrophages, although they appear to be easily transducible by lentiviral vectors, fail to maintain a sustained VCN/cell after a certain period of time and are, thus, somewhat resistant to transduction. This feature precludes the possibility of gene addition via lentiviral vectors, but opens new perspectives for transient gene expression such as by employing genome editing tools, i.e., by transiently expressing components of the CRISPR/Cas9 system.
4. Discussion
Intracellular infectious agents such as bacteria of the genus Brucella, which affect ruminant animal populations, pose severe public health issues that are usually translated into a substantial financial burden for stockbreeders [29]. The only available means for eradicating the disease is based on vaccination programs, frequent monitoring and slaughtering of seropositive carriers [30]. In addition, the available animal vaccines are ambiguously effective, while, in addition, infectious for veterinarians [2]. Since application of programs for the eradication of brucellosis have been failing in southeastern Europe and the Mediterranean for almost four decades [31], massive sanitation of livestock is considered indispensable. Gene therapy, which has recently noted significant clinical successes in monogenic human disorders [5,6,7,8,9,10], as well as infectious diseases [11,12], could possibly constitute a novel, molecular-based platform for the development of a permanent cure not only for brucellosis, but for many other infectious diseases, where the microorganism presents an intracellular survival limited to the monocyte/macrophage lineage, thus effectively evading antibiotic treatment. For example, gene therapy could be utilized to render specific cell lineages resistant to infection, by deleting or inactivating genes that code for factors that play a critical role in the replication of the infectious agents in the host cytoplasm. This would be particularly feasible by introduction, via a lentiviral vector, of nuclease genetic information that would function specifically against the microbe genome, without harming the host DNA.
However, it is currently unknown whether macrophages of ruminant animal origin present a substantial susceptibility to transduction, so that gene therapy could be effectively implemented. Aiming at the development of a gene therapy approach targeting and inactivating specific genetic features of intracellular bacteria in our lab, we have tested the transducibility of ovine macrophages by lentiviral vectors in terms of transgene expression and VCN.
We first sought to determine whether the cells we isolated from the peripheral blood of healthy sheep could be cultured and expanded, so that an adequate number of macrophages could be generated for transduction. We show that the vast majority of cells in culture (~80%), in the presence of macrophage-specific media, did express the ruminant monocyte/macrophage CD14 differentiation antigen [32,33], to a various extent, which could be attributed to possible fluctuations in the differentiation status of the cells in culture. Extrinsic supplementation of growth factors, such as granulocyte-macrophage colony-stimulating factor (GM-CSF), to simulate in vivo maturation conditions [34] or the addition of interleukin 4/interleukin 13 to induce phagocytosis [35] may aid a more uniform CD14 expression. Further analysis of the surface antigens expressed or sorting of the populations may also elucidate the nature of these cells. Usually, phagocytosis is an effect that takes place within minutes [36] and is highly dependent on target shape and diameter [37]. Our results are in accordance with previously published literature that demonstrates that the maximal phagocytosis of polystyrene microspheres took place when their size was in the range of 1.0–3.0 μm. Thus, the absence of engulfment events in the cells exposed to the 0.2 μm-sized beads, along with the escalating percentage of phagocytic activity observed as the size of the beads increased, corroborates findings of other researchers [37].
It is widely acknowledged that macrophages, due to their germicidal nature, are particularly refractory to transduction [38,39]. Consequently, viral vector pseudotypes play a critical role in the transduction susceptibility of cells of the monocyte/macrophage lineage. In our study, we have utilized a classic VSV-G envelope, which allows infection to a wide range of cell types [40]. We report for the first time that sheep macrophages can be transduced with a conventional GFP lentiviral vector, but only under conditions of a high MOI and in the presence of cationic polymers, such as hexadimethrine bromide, which is commercially known as polybrene, or protamine sulfate. Both polybrene and protamine sulfate act by neutralizing the charge repulsion between virions and the cell surface [41,42]. Although they can be toxic to certain cell types, such as primary neurons [42], they are vastly used to increase the efficiency of transduction in cell cultures [43], if carefully titrated. In our case, we did not observe any toxic events associated with the use of polybrene or protamine sulfate, since, at all times, the viability of macrophages measured by trypan blue exclusion was over 90% (data not shown). This stands in accordance with a previous study from our group [26], where the same lentiviral vector, refined with an ultrafiltration concentration protocol as in our case, was used to transduce murine total bone marrow or human CD34+ hematopoietic stem cells, which resulted in equal transduction efficiency and reduced toxicity, compared to lentiviral vectors produced with standard ultracentrifugation-based methods.
Furthermore, polybrene-treated macrophages demonstrated a higher gene marking level even after culturing the cells for one week in vitro, although it should be noted that there was no statistical significance of the polybrene-treated cells against their no additive and protamine sulfate counterparts at the same time point, which most probably reflects the small number of samples analyzed. In addition, it is worth noting that the VCN per cell at the same time point was well correlated with the gene transfer rates measured by flow cytometry, since we observed higher GFP expression at the third day after transduction and a marked decrease both in VCN and GFP expression at Day 8 after transduction.
This finding is anticipated, in part, because lentiviral vector lots were derived from research-grade preparations, and the presence of transient plasmids contained in the vector stocks may lead to an overestimation of VCN at early time points post transduction. Thus, VCN is most often accurately measured when episomal particles are washed away upon subculturing of the cells. However, these results suggest that ovine macrophages although they appear to be easily transducible by lentiviral vectors, fail to maintain high VCN/cell after a sustained period of time and are, thus, somewhat resistant to transduction. This feature precludes the possibility of gene addition via lentiviral vectors, but opens new perspectives for transient gene expression such as by employing genome editing tools, i.e., by transiently expressing components of the CRISPR/Cas9 system in order to render the cells resistant to specific microorganisms.
In summary, the above data provide insights towards the generation of novel, target-specific molecular treatments against intracellular bacteria and are thus anticipated to provide the benchmark for gene therapy implementations of zoonoses.
5. Conclusions
We show that ovine macrophages are relatively resistant to transduction with lentiviral vectors even at a high multiplicity of infection and that addition of transduction enhancers, such as polybrene, increases transgene expression in vitro. Overall, we demonstrate that ovine macrophages may be efficiently expanded and transduced in culture, thus providing the benchmark for gene therapy applications for zoonotic diseases.
Author Contributions
G.K. designed and performed the experiments, i.e., isolation, culture and characterization of ovine macrophages, carried out transductions and performed DNA isolation and flow cytometric analyses G.K. also wrote the manuscript. S.K. coordinated the study and also helped to draft the manuscript. E.P. (Evanthia Petridou) conceived of and coordinated the study and the experiments and also helped to draft the manuscript. E.P. (Eleni Papanikolaou) conceived of the study, designed the experiments, made vector batches, carried out the initial transductions and performed the phagocytosis assay and the real-time PCR. Both E.P. (Evanthia Petridou) and E.P. (Eleni Papanikolaou) helped to draft the manuscript and have also given final approval of the version to be published. All authors read and approved the final manuscript.
Funding
This post-doctoral research was conducted with a scholarship from the Greek State Scholarships Foundation, which was funded by the Operation “Support for post-doctoral researchers” from the resources of the Operational Program “Human Resources Development, Education and Lifelong Learning” with Priority Axes 6, 8 and 9 and is co-funded by the European Social Fund (ESF) and the Greek state.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Von Bargen, K.; Gorvel, J.P.; Salcedo, S.P. Internal affairs: Investigating the Brucella intracellular lifestyle. FEMS Microbiol. Rev. 2012, 36, 533–562. [Google Scholar] [CrossRef] [PubMed]
- Blasco, J.M. A review of the use of B. melitensis Rev 1 vaccine in adult sheep and goats. Prev. Vet. Med. 1997, 31, 275–283. [Google Scholar] [CrossRef]
- Minas, A.; Minas, M.; Stournara, A.; Tselepidis, S. The "effects" of Rev-1 vaccination of sheep and goats on human brucellosis in Greece. Prev. Vet. Med. 2004, 64, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Kay, M.A. State-of-the-art gene-based therapies: The road ahead. Nat. Rev. Genet. 2011, 12, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Cavazzana-Calvo, M.; Hacein-Bey, S.; de Saint Basile, G.; Gross, F.; Yvon, E.; Nusbaum, P.; Selz, F.; Hue, C.; Certain, S.; Casanova, J.L.; et al. Gene therapy of human severe combined immunodeficiency (SCID)-X1 disease. Science 2000, 288, 669–672. [Google Scholar] [CrossRef] [PubMed]
- Cartier, N.; Hacein-Bey-Abina, S.; Bartholomae, C.C.; Veres, G.; Schmidt, M.; Kutschera, I.; Vidaud, M.; Abel, U.; Dal-Cortivo, L.; Caccavelli, L.; et al. Hematopoietic stem cell gene therapy with a lentiviral vector in X-linked adrenoleukodystrophy. Science 2009, 326, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Cavazzana-Calvo, M.; Payen, E.; Negre, O.; Wang, G.; Hehir, K.; Fusil, F.; Down, J.; Denaro, M.; Brady, T.; Westerman, K.; et al. Transfusion independence and HMGA2 activation after gene therapy of human b-thalassaemia. Nature 2010, 467, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Aiuti, A.; Biasco, L.; Scaramuzza, S.; Ferrua, F.; Cicalese, M.P.; Baricordi, C.; Dionisio, F.; Calabria, A.; Giannelli, S.; Castiello, M.C.; et al. Lentiviral hematopoietic stem cell gene therapy in patients with Wiskott-Aldrich syndrome. Science 2013, 341. [Google Scholar] [CrossRef] [PubMed]
- Biffi, A.; Montini, E.; Lorioli, L.; Cesani, M.; Fumagalli, F.; Plati, T.; Baldoli, C.; Martino, S.; Calabria, A.; Canale, S.; et al. Lentiviral hematopoietic stem cell gene therapy benefits metachromatic leukodystrophy. Science 2013, 341. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.A.; Walters, M.C.; Kwiatkowski, J.; Rasko, J.E.J.; Ribeil, J.A.; Hongeng, S.; Magrin, E.; Schiller, G.J.; Payen, E.; Semeraro, M.; et al. Gene therapy in patients with transfusion-dependent β-thalassemia. N. Engl. J. Med. 2018, 378, 1479–1493. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.R.; Yang, H.C.; Kuo, Y.T.; Liu, C.J.; Yang, T.Y.; Sung, K.C.; Lin, Y.Y.; Wang, H.Y.; Wang, C.C.; Shen, Y.C.; et al. The CRISPR/Cas9 system facilitates clearance of the intrahepatic HBV templates in vivo. Mol. Ther. Nucleic Acids 2014, 3. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Kaminski, R.; Yang, F.; Zhang, Y.; Cosentino, L.; Li, F.; Luo, B.; Alvarez-Carbonell, D.; Garcia-Mesa, Y.; Karn, J.; et al. RNA-directed gene editing specifically eradicates latent and prevents new HIV-1 infection. Proc. Natl. Acad. Sci. USA 2014, 111, 11461–11466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarrosson-Wuilleme, L.; Goujon, C.; Bernaud, J.; Rigal, D.; Darlix, J.L.; Cimarelli, A. Transduction of nondividing human macrophages with gammaretrovirus-derived vectors. J. Virol. 2006, 80, 1152–1159. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Planelles, V.; Sui, Z.; Gartner, S.; Maggirwar, S.B.; Dewhurst, S.; Ye, L.; Nerurkar, V.R.; Yanagihara, R.; Lu, Y. HIV-1-based defective lentiviral vectors efficiently transduce human monocytes-derived macrophages and suppress replication of wild-type HIV-1. J. Gene Med. 2006, 8, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Yang, S.; Wu, C.; Ye, L.; Lu, Y. Effective transduction of primary mouse blood- and bone marrow-derived monocytes/macrophages by HIV-based defective lentiviral vectors. J. Virol. Methods 2006, 134, 66–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linterman, K.S.; Palmer, D.N.; Kay, G.W.; Barry, L.A.; Mitchell, N.L.; McFarlane, R.G.; Black, M.A.; Sands, M.S.; Hughes, S.M. Lentiviral-mediated gene transfer to the sheep brain: Implications for gene therapy in Batten disease. Hum. Gene Ther. 2011, 22, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Gray-Edwards, H.L.; Randle, A.N.; Maitland, S.A.; Benatti, H.R.; Hubbard, S.M.; Canning, P.F.; Vogel, M.B.; Brunson, B.L.; Hwang, M.; Ellis, L.E.; et al. Adeno-associated virus gene therapy in a sheep model of Tay-Sachs disease. Hum. Gene Ther. 2018, 29, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wong, E.; Miller, D.; Smith, G.; Anson, D.; Parsons, D. Lentiviral airway gene transfer in lungs of mice and sheep: Successes and challenges. J. Gene Med. 2010, 12, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Banin, E.; Gootwine, E.; Obolensky, A.; Ezra-Elia, R.; Ejzenberg, A.; Zelinger, L.; Honig, H.; Rosov, A.; Yamin, E.; Sharon, D.; et al. Gene augmentation therapy restores retinal function and visual behavior in a sheep model of CNGA3 achromatopsia. Mol. Ther. 2015, 23, 1423–1433. [Google Scholar] [CrossRef] [PubMed]
- Porada, C.D.; Park, P.; Almeida-Porada, G.; Zanjani, E.D. The sheep model of in utero gene therapy. Fetal Diagn. Ther. 2004, 19, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Crespo, H.; Bertolotti, L.; Juganaru, M.; Glaria, I.; de Andrés, D.; Amorena, B.; Rosati, S.; Reina, R. Small ruminant macrophage polarization may play a pivotal role on lentiviral infection. Vet. Res. 2013, 44. [Google Scholar] [CrossRef] [PubMed]
- Jarczak, J.; Kaba, J.; Reczynska, D.; Bagnicka, E. Impaired expression of cytokines as a result of viral infections with an emphasis on small ruminant lentivirus infection in goats. Viruses 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- De Pablo-Maiso, L.; Glaria, I.; Crespo, H.; Nistal-Villán, E.; Andrésdóttir, V.; de Andrés, D.; Amorena, B.; Reina, R. Characterization of ovine A3Z1 restriction properties against small ruminant lentiviruses (SRLVs). Viruses 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.T.; Griffin, F.T. A comparison of ovine monocyte-derived macrophage function following infection with Mycobacterium avium ssp. avium and Mycobacterium avium ssp. paratuberculosis. Immunol. Cell Biol. 2006, 84, 349–356. [Google Scholar] [PubMed] [Green Version]
- Papanikolaou, E.; Georgomanoli, M.; Stamateris, E.; Panetsos, F.; Karagiorga, M.; Tsaftaridis, P.; Graphakos, S.; Anagnou, N.P. The new self-inactivating lentiviral vector for thalassemia gene therapy combining two HPFH activating elements corrects human thalassemic hematopoietic stem cells. Hum. Gene Ther. 2012, 23, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, E.; Kontostathi, G.; Drakopoulou, E.; Georgomanoli, M.; Stamateris, E.; Vougas, K.; Vlahou, A.; Maloy, A.; Ware, M.; Anagnou, N.P. Characterization and comparative performance of lentiviral vector preparations concentrated by either one-step ultrafiltration or ultracentrifugation. Virus Res. 2013, 175, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.Y.; Westaway, D.; Smit, A.F.A.; Wang, K.; Seto, J.; Chen, L.; Acharya, C.; Ankener, M.; Baskin, D.; Cooper, C.; et al. Complete genomic sequence and analysis of the prion protein gene region from three mammalian species. Genome Res. 1998, 8, 1022–1037. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Crespo, D.; Juste, R.A.; Hurtado, A. Selection of ovine housekeeping genes for normalisation by real-time RT-PCR; analysis of PrP gene expression and genetic susceptibility to scrapie. BMC Vet. Res. 2005, 1, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Donev, D.M. Brucellosis as priority public health challenge in South Eastern European countries. Croat. Med. J. 2010, 51, 283–284. [Google Scholar] [CrossRef] [PubMed]
- Avila-Calderón, E.; Lopez-Merino, A.; Sriranganathan, N.; Boyle, S.; Contreras-Rodríguez, A. A history of the development of Brucella vaccines. Biomed. Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Pappas, G.; Papadimitriou, P.; Akritidis, N.; Christou, L.; Tsianos, E.V. The new global map of human brucellosis. Lancet Infect. Dis. 2006, 6, 91–99. [Google Scholar] [CrossRef]
- Gupta, V.K.; McConnell, I.; Dalziel, R.G.; Hopkins, J. Identification of the sheep homologue of the monocyte cell surface molecule - CD14. Vet. Immunol. Immunopathol. 1996, 51, 89–99. [Google Scholar] [CrossRef]
- Berthon, P.; Hopkins, J. Ruminant cluster CD14. Vet. Immunol. Immunopathol. 1996, 52, 245–248. [Google Scholar] [CrossRef]
- May, K.F.; Jinushi, M.; Dranoff, G. Immunosurveillance: Innate and adaptive antitumor immunity. In Cancer Immunotherapy: Immune Suppression and Tumor Growth, 2nd ed.; Prendergast, G.C., Jaffee, E.M., Eds.; Academic Press: San Diego, CA, USA, 2013; pp. 101–113. ISBN 9780123942968. [Google Scholar]
- Gordon, S.; Martinez, F.O. Alternative activation of macrophages: Mechanism and functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Nagl, M.; Kacani, L.; Mullauer, B.; Lemberger, E.M.; Stoiber, H.; Sprinzl, G.M.; Schennach, H.; Dierich, M.P. Phagocytosis and killing of bacteria by professional phagocytes and dendritic cells. Clin. Vaccine Immunol. 2002, 9, 1165–1168. [Google Scholar] [CrossRef]
- Paul, D.; Achouri, S.; Yoon, Y.Z.; Herre, J.; Bryant, C.E.; Cicuta, P. Phagocytosis dynamics depends on target shape. Biophys. J. 2013, 105, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Edwards, J.P.; Mosser, D.M. The expression of exogenous genes in macrophages: Obstacles and opportunities. Methods Mol. Biol. 2009, 531, 123–143. [Google Scholar] [PubMed]
- Bobadilla, S.; Sunseri, N.; Landau, N.R. Efficient transduction of myeloid cells by an HIV-1-derived lentiviral vector that packages the Vpx accessory protein. Gene Ther. 2013, 20, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Cronin, J.; Zhang, X.Y.; Reiser, J. Altering the tropism of lentiviral vectors through pseudotyping. Curr. Gene Ther. 2005, 5, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.E.; Rosinski, M.; Morgan, J.R.; Yarmush, M.L. Charged polymers modulate retrovirus transduction via membrane charge neutralization and virus aggregation. Biophys. J. 2004, 86, 1234–1242. [Google Scholar] [CrossRef]
- Denning, W.; Das, S.; Guo, S.; Xu, J.; Kappes, J.C.; Hel, Z. Optimization of the transductional efficiency of lentiviral vectors: Effect of sera and polycations. Mol. Biotechnol. 2013, 53, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.E.; Morgan, J.R.; Yarmush, M.L. Polybrene increases retrovirus gene transfer efficiency by enhancing receptor-independent virus adsorption on target cell membranes. Biophys. Chem. 2002, 97, 159–172. [Google Scholar] [CrossRef]
Figure 1.
Ovine macrophages, after three weeks in culture. Cells were derived from the peripheral blood mononuclear fraction after ficoll separation and were left to mature in macrophage-specialized media supplemented with fetal bovine serum.
Figure 1.
Ovine macrophages, after three weeks in culture. Cells were derived from the peripheral blood mononuclear fraction after ficoll separation and were left to mature in macrophage-specialized media supplemented with fetal bovine serum.

Figure 2.
(A) A representative flow cytometry overlaid histogram showing the differentiation pattern of cultured cells towards the macrophage lineage (CD14+). Unstained cells (green histogram), as well as an FITC-conjugated mouse IgG1 isotypic antibody (yellow histogram) served as negative controls. (B) A representative flow cytometry dot plot depicting the two distinct cell populations that express different levels of the CD14 antigen. FITC: fluorescein isothiocyanate; FSC: forward scatter; SSC: side scatter.
Figure 2.
(A) A representative flow cytometry overlaid histogram showing the differentiation pattern of cultured cells towards the macrophage lineage (CD14+). Unstained cells (green histogram), as well as an FITC-conjugated mouse IgG1 isotypic antibody (yellow histogram) served as negative controls. (B) A representative flow cytometry dot plot depicting the two distinct cell populations that express different levels of the CD14 antigen. FITC: fluorescein isothiocyanate; FSC: forward scatter; SSC: side scatter.
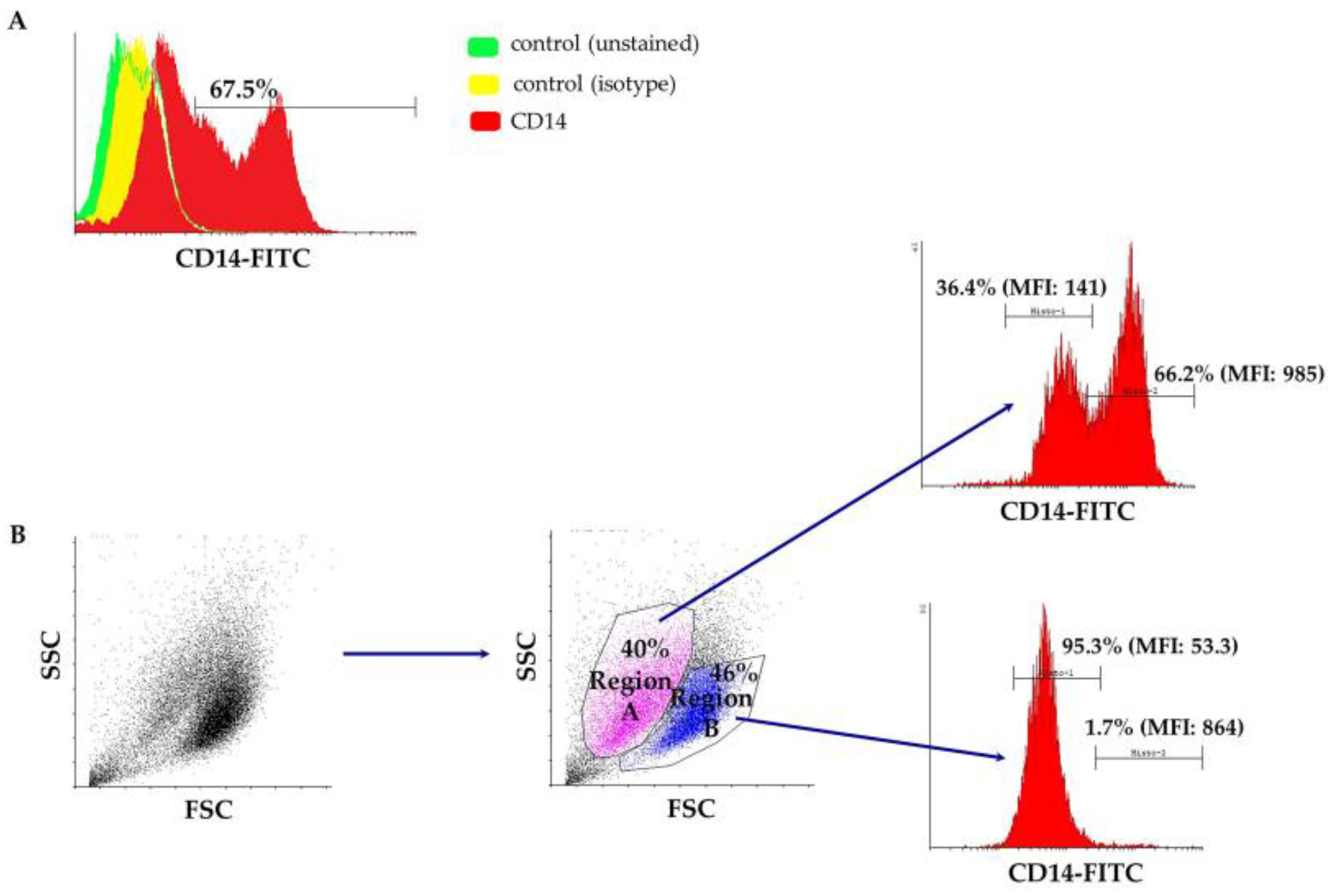
Figure 3.
Macrophage phagocytosis assay. Results shown after flow cytometry analysis from one representative experiment. Histograms on the top illustrate the percentage of macrophages presenting phagocytosis events after a 24-hour exposure to variously-sized fluorescent beads (0.2 μm, 0.5 μm, 1 μm and 2 μm). The respective 3D dot plots are depicted on the bottom of the page. Note that both cell populations (i.e., from Region A and Region B) appear to possess phagocytic properties. Macrophages not exposed to beads served as a negative control. FSC: forward scatter; SSC: side scatter.
Figure 3.
Macrophage phagocytosis assay. Results shown after flow cytometry analysis from one representative experiment. Histograms on the top illustrate the percentage of macrophages presenting phagocytosis events after a 24-hour exposure to variously-sized fluorescent beads (0.2 μm, 0.5 μm, 1 μm and 2 μm). The respective 3D dot plots are depicted on the bottom of the page. Note that both cell populations (i.e., from Region A and Region B) appear to possess phagocytic properties. Macrophages not exposed to beads served as a negative control. FSC: forward scatter; SSC: side scatter.

Figure 4.
Representative flow cytometry results, three days post transduction with a conventional lentiviral vector expressing the green fluorescence protein in the presence or absence of transduction enhancers (polybrene or protamine sulfate). The percentage of GFP-positive cells is displayed on each histogram. Mock-transduced cells served as a negative control. GFP: green fluorescence protein.
Figure 4.
Representative flow cytometry results, three days post transduction with a conventional lentiviral vector expressing the green fluorescence protein in the presence or absence of transduction enhancers (polybrene or protamine sulfate). The percentage of GFP-positive cells is displayed on each histogram. Mock-transduced cells served as a negative control. GFP: green fluorescence protein.
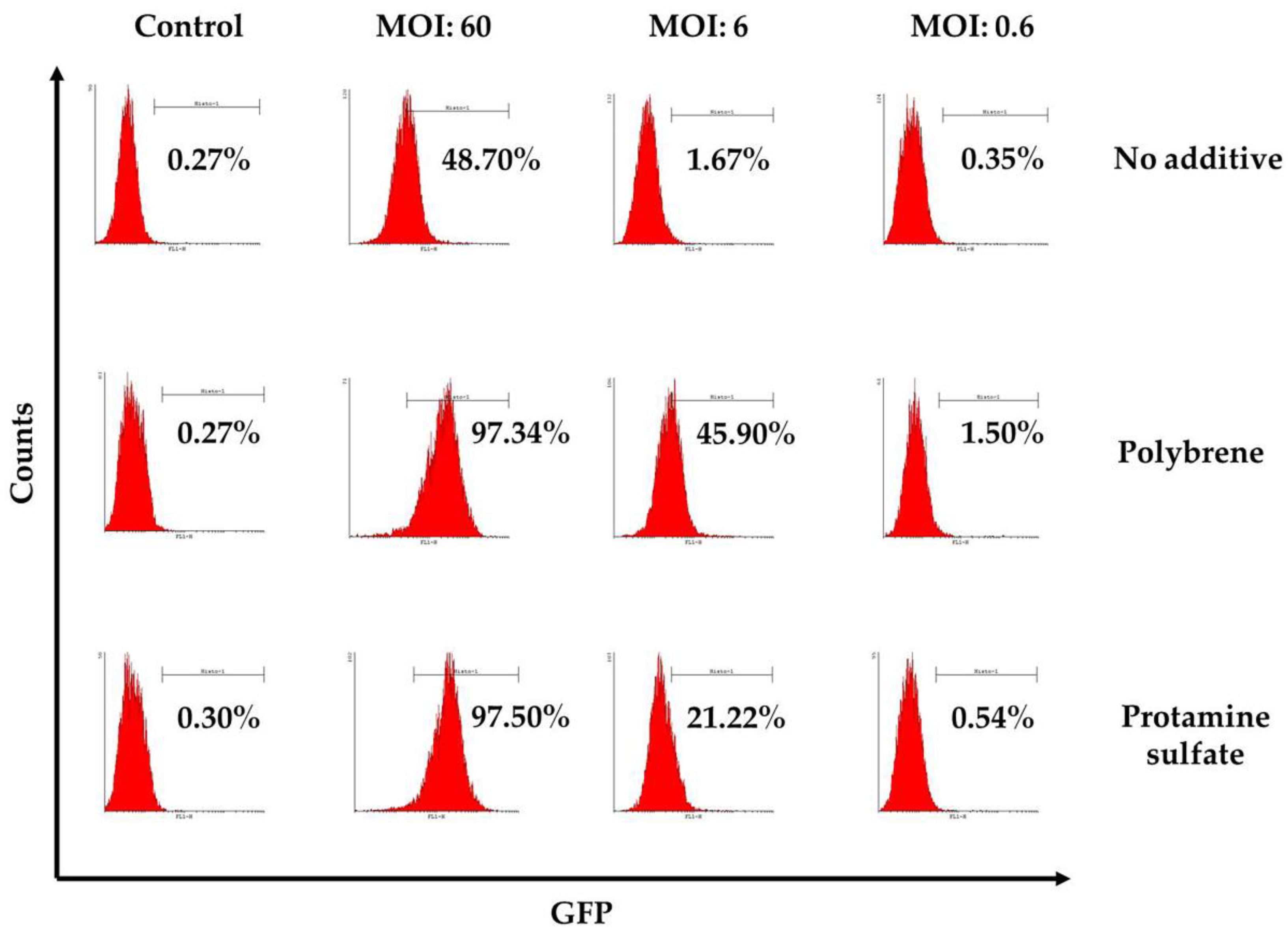
Figure 5.
Comparative analysis of GFP expression in ovine macrophages, by flow cytometry, at various multiplicities of infection on Day 3 post transduction. Each MOI condition was repeated five times. Mock-transduced cells served as a negative control. GFP: green fluorescence protein; MOI: multiplicity of infection.
Figure 5.
Comparative analysis of GFP expression in ovine macrophages, by flow cytometry, at various multiplicities of infection on Day 3 post transduction. Each MOI condition was repeated five times. Mock-transduced cells served as a negative control. GFP: green fluorescence protein; MOI: multiplicity of infection.
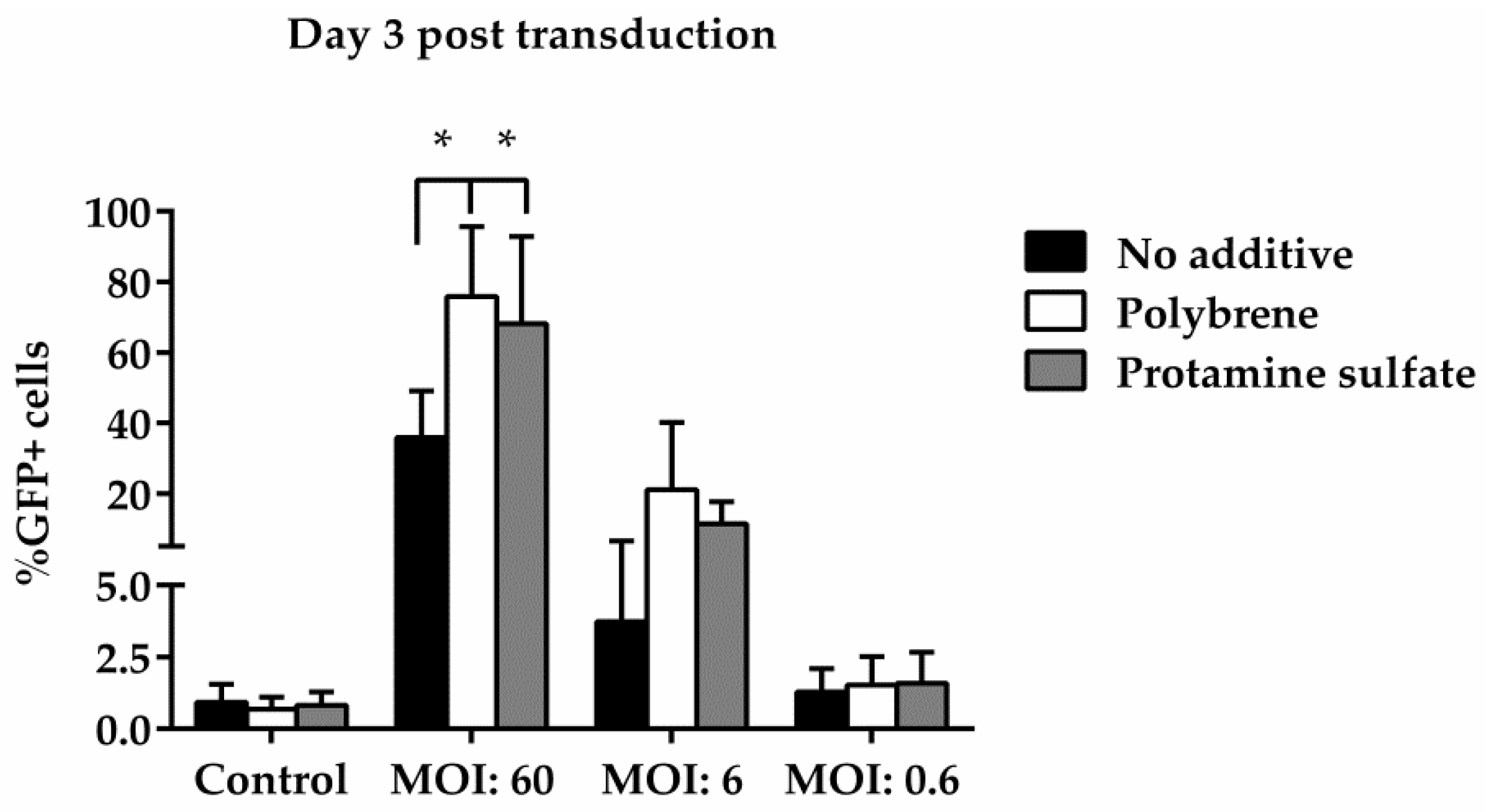
Figure 6.
Comparative analysis of GFP expression in ovine macrophages, by flow cytometry, at various multiplicities of infection on Day 8 post transduction. Each MOI condition was repeated two times. Mock-transduced cells served as a negative control. GFP: green fluorescence protein; MOI: multiplicity of infection.
Figure 6.
Comparative analysis of GFP expression in ovine macrophages, by flow cytometry, at various multiplicities of infection on Day 8 post transduction. Each MOI condition was repeated two times. Mock-transduced cells served as a negative control. GFP: green fluorescence protein; MOI: multiplicity of infection.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Percentage of FL-1 positive cells after 24 h of incubation with green fluorescent beads of different diameters.
Table 1.
Percentage of FL-1 positive cells after 24 h of incubation with green fluorescent beads of different diameters.
| 1st Experiment | 2nd Experiment | 3rd Experiment | |
|---|---|---|---|
| Control | 0.1 | 0.17 | 0.13 |
| 0.2 μm | 1.15 | 1.21 | 1 |
| 0.5 μm | 1.7 | 2.6 | 2.2 |
| 1 μm | 3.6 | 6 | 5.75 |
| 2 μm | 8.9 | 8.6 | 10 |
Table 2.
Detailed results from all the experiments performed to determine the transduction efficiency. Numbers indicate the percentage of GFP-positive cells 3 days after transduction.
Table 2.
Detailed results from all the experiments performed to determine the transduction efficiency. Numbers indicate the percentage of GFP-positive cells 3 days after transduction.
| 1st Experiment | 2nd Experiment | 3rd Experiment | 4th Experiment | 5th Experiment | ||
|---|---|---|---|---|---|---|
| No additive | control | 0.27 | 0.09 | 1.4 | 0.9 | 1.54 |
| MOI *:60 | 48.7 | 48.1 | 43.7 | 34.2 | 18.3 | |
| MOI:6 | 1.67 | 1.66 | 8.8 | 2.7 | 2.37 | |
| MOI:0.6 | 0.35 | 0.16 | 1.7 | 1.9 | 1.47 | |
| Polybrene | control | 0.27 | 0.09 | 0.86 | 0.7 | 0.95 |
| MOI:60 | 97.34 | 97.46 | 78 | 75.6 | 51.4 | |
| MOI:6 | 45.9 | 42.7 | 14.55 | 14.7 | 2.75 | |
| MOI:0.6 | 1.5 | 0.71 | 1.34 | 1.25 | 0.91 | |
| Protamine sulfate | control | 0.3 | 0.15 | 1.04 | 1.05 | 1.14 |
| MOI:60 | 97.5 | 94.7 | 70 | 64.7 | 40.3 | |
| MOI:6 | 21.2 | 16.8 | 6.8 | 10.8 | 7.55 | |
| MOI:0.6 | 0.54 | 0.41 | 1.06 | 2.8 | 1.76 |
* MOI: multiplicity of infection.
Table 3.
Comparative vector copy number analysis of transduced macrophages, by quantitative PCR, at two different time points.
Table 3.
Comparative vector copy number analysis of transduced macrophages, by quantitative PCR, at two different time points.
| Days Post Transduction | MOI: 60 | |
|---|---|---|
| No additive | 3 | 10.46 |
| 8 | 0.13 | |
| Polybrene | 3 | 11.33 |
| 8 | 0.29 | |
| Protamine sulfate | 3 | 10.94 |
| 8 | 0.21 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Karponi, G.; Kritas, S.; Petridou, E.; Papanikolaou, E. Efficient Transduction and Expansion of Ovine Macrophages for Gene Therapy Implementations. Vet. Sci. 2018, 5, 57. https://doi.org/10.3390/vetsci5020057
AMA Style
Karponi G, Kritas S, Petridou E, Papanikolaou E. Efficient Transduction and Expansion of Ovine Macrophages for Gene Therapy Implementations. Veterinary Sciences. 2018; 5(2):57. https://doi.org/10.3390/vetsci5020057
Chicago/Turabian StyleKarponi, Garyfalia, Spyridon Kritas, Evanthia Petridou, and Eleni Papanikolaou. 2018. "Efficient Transduction and Expansion of Ovine Macrophages for Gene Therapy Implementations" Veterinary Sciences 5, no. 2: 57. https://doi.org/10.3390/vetsci5020057
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.