
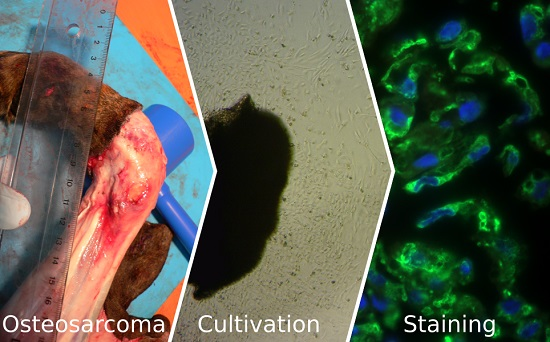
Establishment and Characterization of New Canine and Feline Osteosarcoma Primary Cell Lines

Abstract
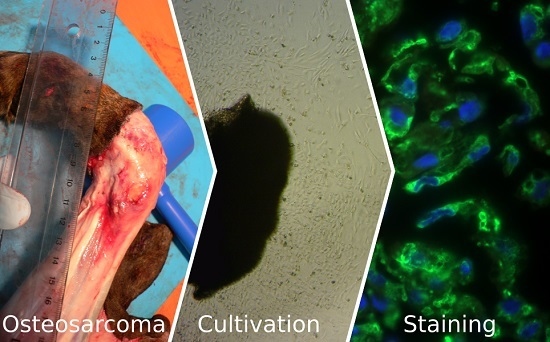
:
1. Introduction
2. Experimental Section
2.1. Animals
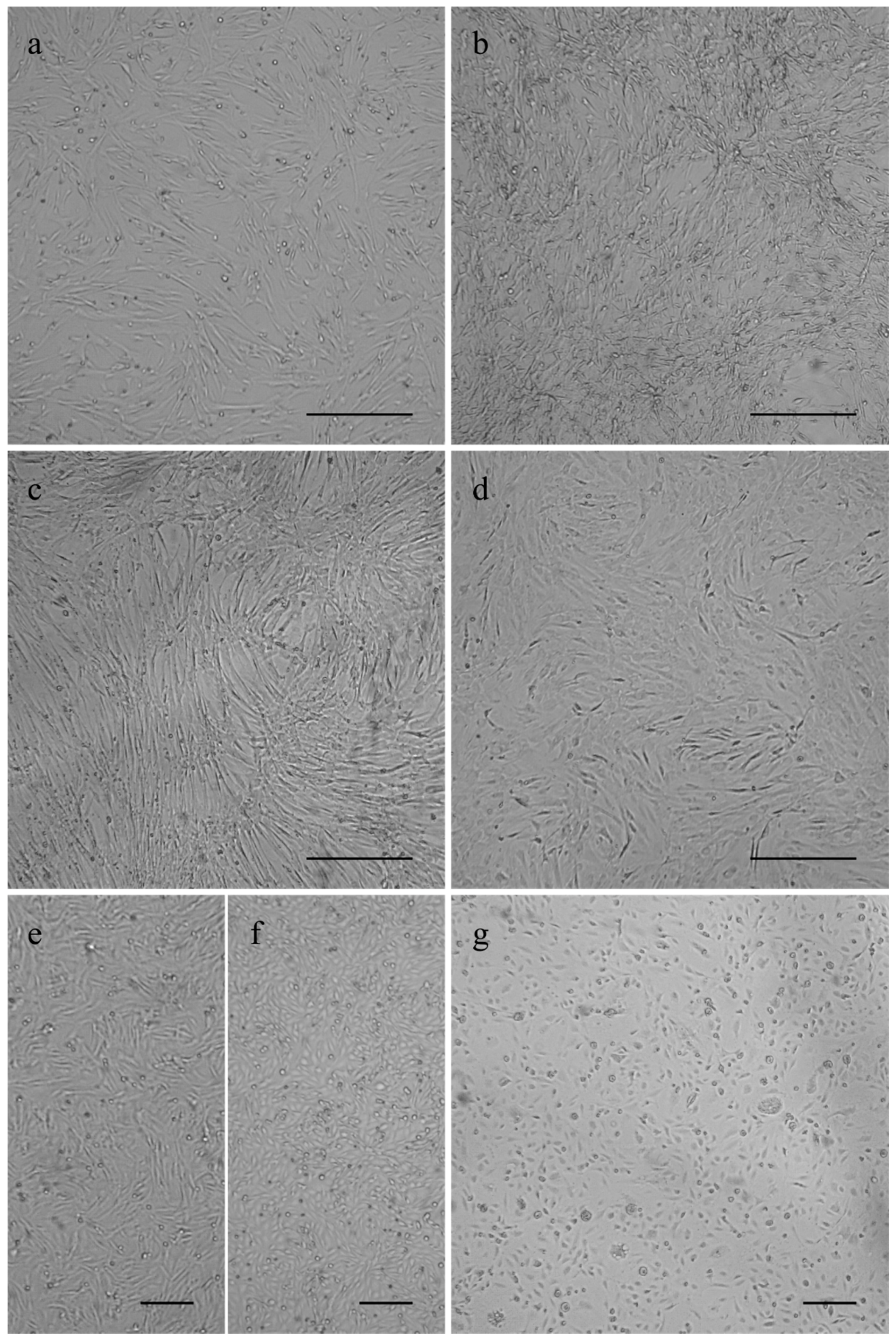
2.2. Cell Culture
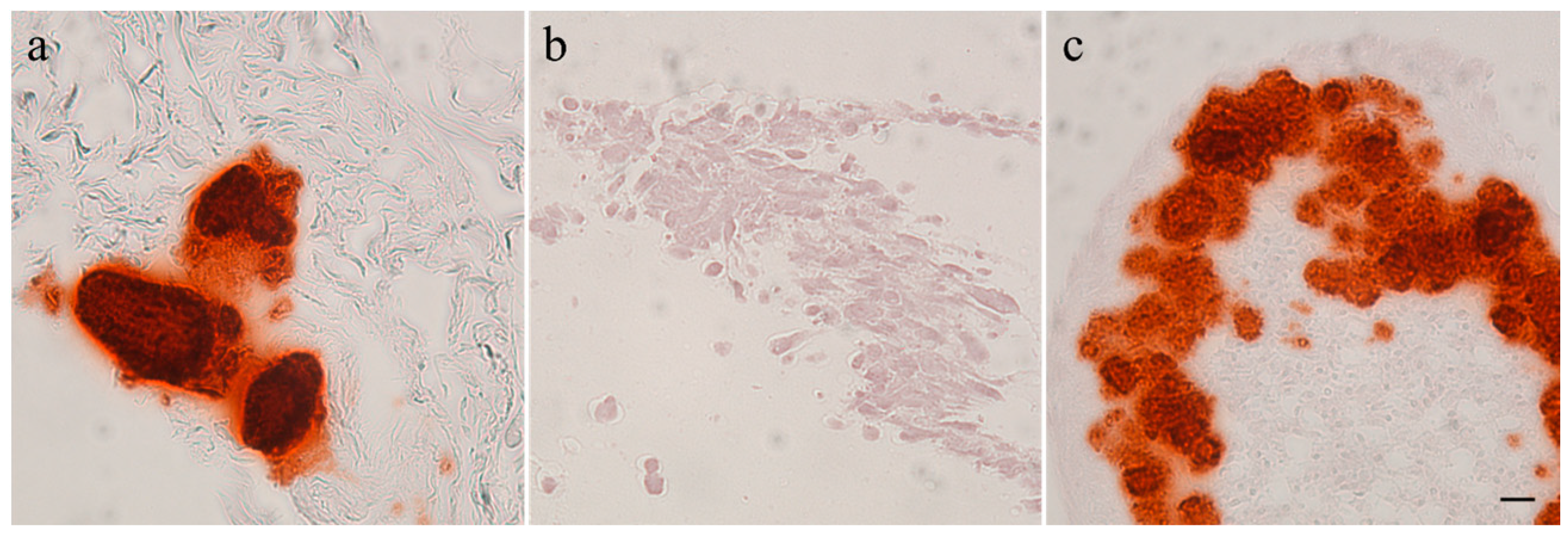
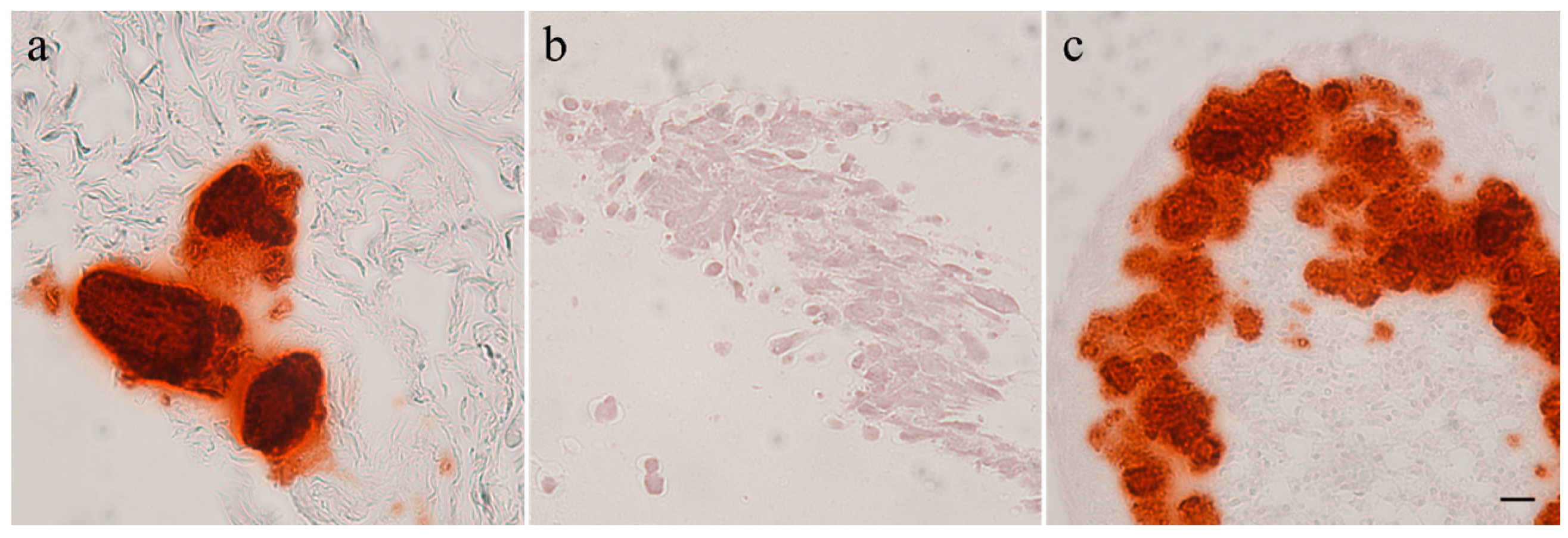
2.3. Histology
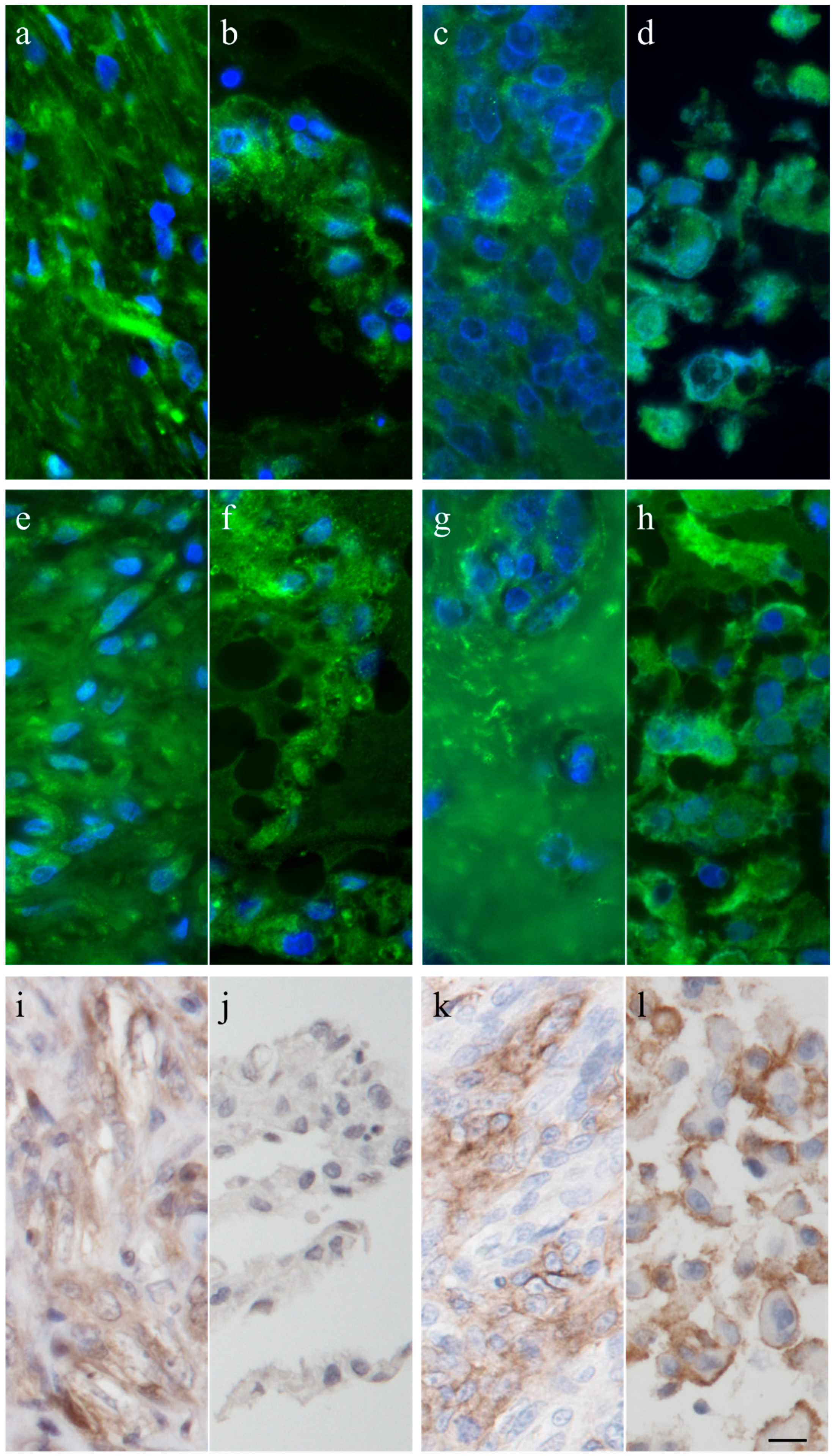
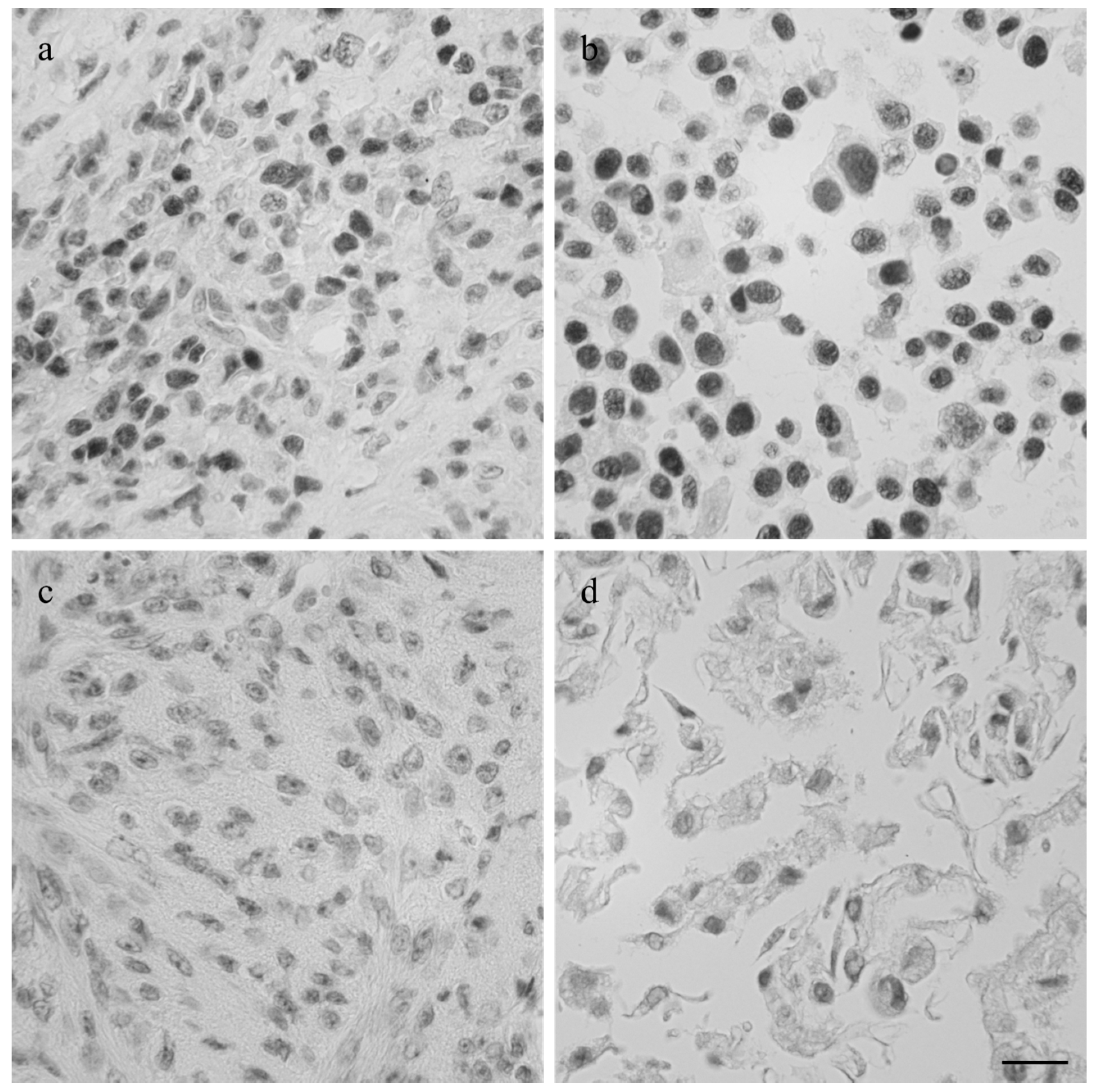
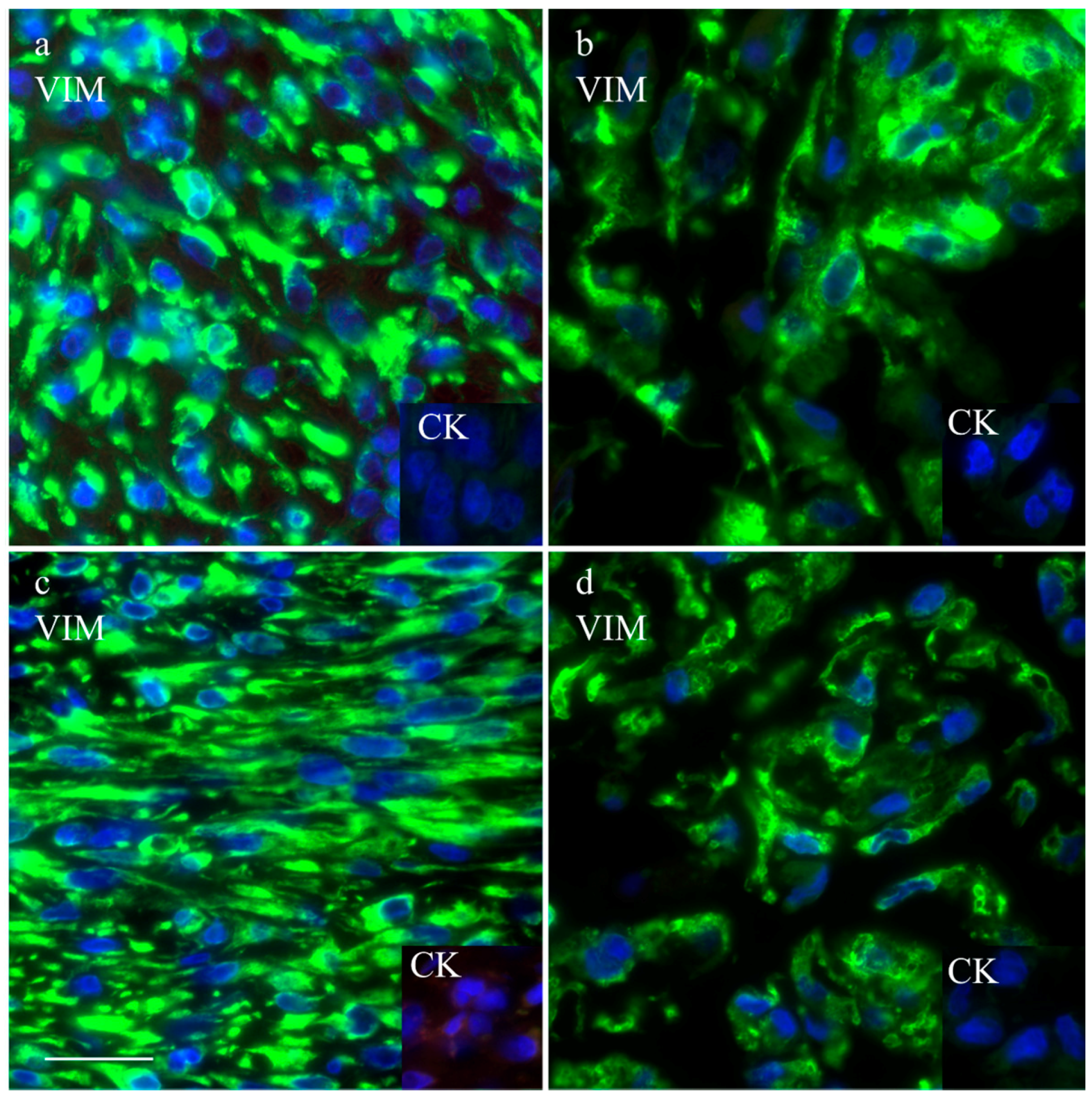
2.4. Immunohistochemistry
2.5. PCR
2.6. DNA-Fingerprint Assay
3. Results
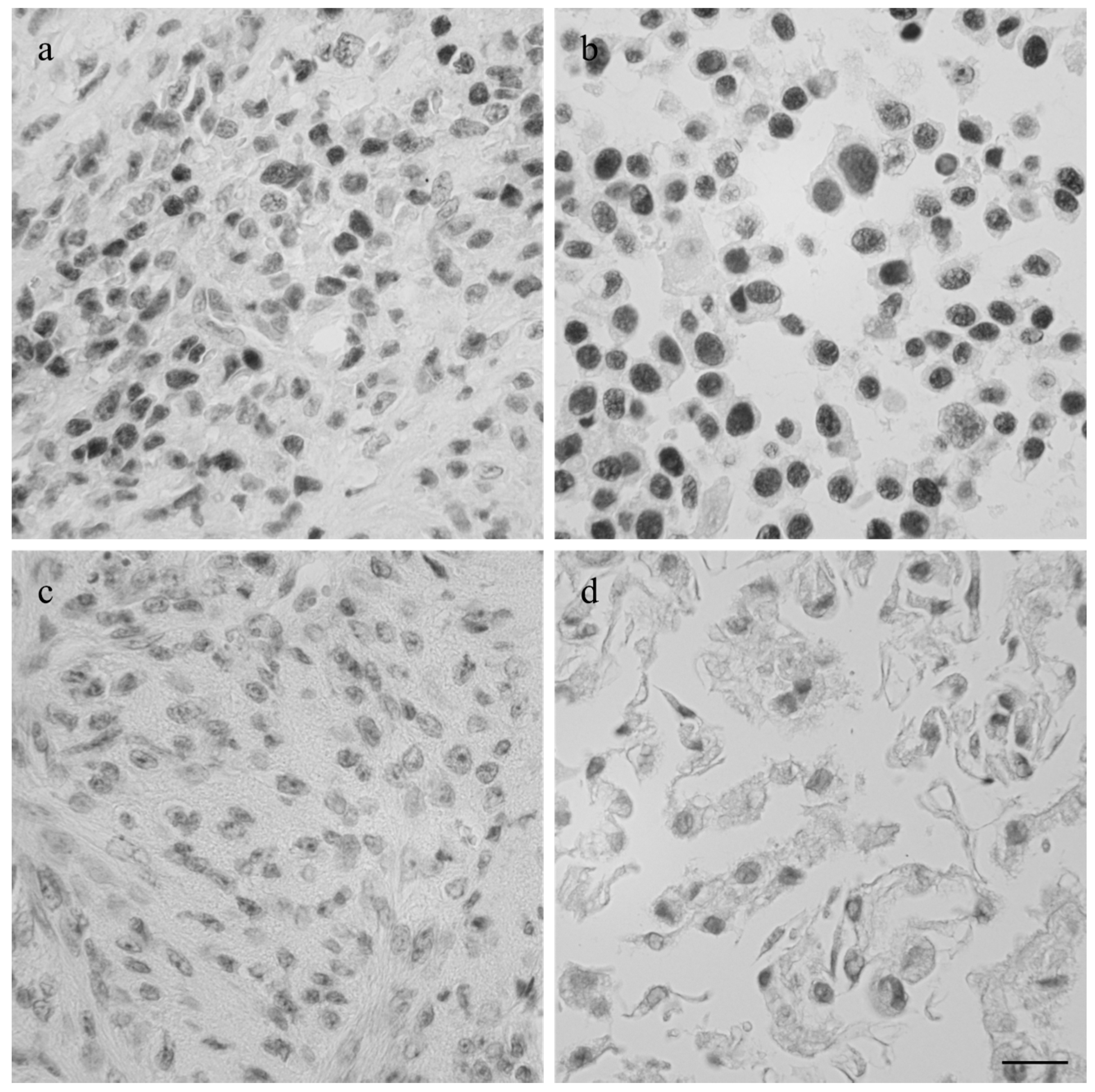
3.1. Immunohistochemistry
3.2. PCR
3.3. DNA Fingerprint Assay
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Cancer Research Fund International. Available online: https://www.wcrf.org/int/cancer-facts-figures/data-cancer-frequency-country (accessed on 31 May 2016).
- Dorn, C.R.; Taylor, D.O.; Schneider, R.; Hibbard, H.H.; Klauber, M.R. Survey of animal neoplasms in Alameda and Contra Costa Counties, California. II. Cancer morbidity in dogs and cats from Alameda County. J. Natl. Cancer Inst. 1968, 40, 307–318. [Google Scholar] [PubMed]
- Boerma, M.; Burton, G.R.; Wang, J.; Fink, L.M.; McGehee, R.E.; Hauer-Jensen, M. Comparative expression profiling in primary and immortalized endothelial cells: Changes in gene expression in response to hydroxy methylglutaryl-coenzyme A reductase inhibition. Blood Coagul. Fibrinolysis 2006, 17, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Lidington, E.; Moyes, D.; McCormack, A.; Rose, M. A comparison of primary endothelial cells and endothelial cell lines for studies of immune interactions. Transpl. Immunol. 1999, 7, 239–246. [Google Scholar] [CrossRef]
- Rockwell, S. In vivo-in vitro tumour cell lines: Characteristics and limitations as models for human cancer. Br. J. Cancer. Suppl. 1980, 4, 118–122. [Google Scholar] [PubMed]
- Wilson, H.; Huelsmeyer, M.; Chun, R.; Young, K.M.; Friedrichs, K.; Argyle, D.J. Isolation and characterisation of cancer stem cells from canine osteosarcoma. Vet. J. 2008, 175, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Levings, P.P.; McGarry, S.V.; Currie, T.P.; Nickerson, D.M.; McClellan, S.; Ghivizzani, S.C.; Steindler, D.A.; Gibbs, C.P. Expression of an exogenous human Oct-4 promoter identifies tumor-initiating cells in osteosarcoma. Cancer Res. 2009, 69, 5648–5655. [Google Scholar] [CrossRef] [PubMed]
- Di Fiore, R.; Santulli, A.; Drago Ferrante, R.; Giuliano, M.; de Blasio, A.; Messina, C.; Pirozzi, G.; Tirino, V.; Tesoriere, G.; Vento, R. Identification and expansion of human osteosarcoma-cancer-stem cells by long-term 3-aminobenzamide treatment. J. Cell. Physiol. 2009, 219, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Vail, D.M.; Macewen, E.G. Spontaneously occurring tumors of companion animals as models for human cancer. Cancer Investig. 2000, 18, 781–792. [Google Scholar] [CrossRef]
- Withrow, S.J.; Powers, B.E.; Straw, R.C.; Wilkins, R.M. Comparative aspects of osteosarcoma. Dog versus man. Clin. Orthop. 1991, 159–168. [Google Scholar]
- Bruland, O.S. Hematogenous micrometastases in osteosarcoma patients. Clin. Cancer Res. 2005, 11, 4666–4673. [Google Scholar] [CrossRef] [PubMed]
- Mirabello, L.; Troisi, R.J.; Savage, S.A. Osteosarcoma incidence and survival rates from 1973 to 2004: Data from the surveillance, epidemiology, and end results program. Cancer 2009, 115, 1531–1543. [Google Scholar] [CrossRef] [PubMed]
- Misdorp, W.; Hart, A.A. Some prognostic and epidemiologic factors in canine osteosarcoma. J. Natl. Cancer Inst. 1979, 62, 537–545. [Google Scholar] [PubMed]
- Gelberg, K.H.; Fitzgerald, E.F.; Hwang, S.; Dubrow, R. Growth and development and other risk factors for osteosarcoma in children and young adults. Int. J. Epidemiol. 1997, 26, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Longhi, A.; Pasini, A.; Cicognani, A.; Baronio, F.; Pellacani, A.; Baldini, N.; Bacci, G. Height as a risk factor for osteosarcoma. J. Pediatr. Hematol. Oncol. 2005, 27, 314–318. [Google Scholar] [CrossRef] [PubMed]
- Kelsey, J.L.; Moore, A.S.; Glickman, L.T. Epidemiologic studies of risk factors for cancer in pet dogs. Epidemiol. Rev. 1998, 20, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Fodstad, O.; Brøgger, A.; Bruland, O.; Solheim, O.P.; Nesland, J.M.; Pihl, A. Characteristics of a cell line established from a patient with multiple osteosarcoma, appearing 13 years after treatment for bilateral retinoblastoma. Int. J. Cancer 1986, 38, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Kadosawa, T.; Nozaki, K.; Sasaki, N.; Takeuchi, A. Establishment and characterization of a new cell line from a canine osteosarcoma. J. Vet. Med. Sci. 1994, 56, 1167–1169. [Google Scholar] [CrossRef] [PubMed]
- Pautke, C.; Schieker, M.; Tischer, T.; Kolk, A.; Neth, P.; Mutschler, W.; Milz, S. Characterization of osteosarcoma cell lines MG-63, Saos-2 and U-2 OS in comparison to human osteoblasts. Anticancer Res. 2004, 24, 3743–3748. [Google Scholar] [PubMed]
- Séguin, B.; Zwerdling, T.; McCallan, J.L.; DeCock, H.E.V.; Dewe, L.L.; Naydan, D.K.; Young, A.E.; Bannasch, D.L.; Foreman, O.; Kent, M.S. Development of a new canine osteosarcoma cell line. Vet. Comp. Oncol. 2006, 4, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Kadosawa, T.; Mochizuki, M.; Matsunaga, S.; Nishimura, R.; Sasaki, N. Establishment and characterization of two cell lines derived from canine spontaneous osteosarcoma. J. Vet. Med. 1998, 60, 757–760. [Google Scholar] [CrossRef]
- McAllister, R.M.; Gardner, M.B.; Greene, A.E.; Bradt, C.; Nichols, W.W.; Landing, B.H. Cultivation in vitro of cells derived from a human osteosarcoma. Cancer 1971, 27, 397–402. [Google Scholar] [CrossRef]
- Ponten, J.; Saksela, E. Two established in vitro cell lines from human mesenchymal tumours. Int. J. Cancer 1967, 2, 434–447. [Google Scholar] [CrossRef] [PubMed]
- Fogh, J.; Trempe, G. New human tumor cell lines. In Human Tumor Cells in Vitro; Fogh, J., Ed.; Springer U.S.: New York, NY, USA, 1975; pp. 115–159. [Google Scholar]
- Billiau, A.; Edy, V.G.; Heremans, H.; Van Damme, J.; Desmyter, J.; Georgiades, J.A.; de Somer, P. Human interferon: Mass production in a newly established cell line, MG-63. Antimicrob. Agents Chemother. 1977, 12, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Riggs, J.L.; McAllister, R.M.; Lennette, E.H. Immunofluorescent studies of RD-114 virus replication in cell culture. J. Gen. Virol. 1974, 25, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulou, M.; Kirpensteijn, J.; Moens, H.; Kik, M. Histologic prognosticators in feline osteosarcoma: A comparison with phenotypically similar canine osteosarcoma. Vet. Surg. 2008, 37, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Heldmann, E.; Anderson, M.A.; Wagner-Mann, C. Feline osteosarcoma: 145 Cases (1990–1995). J. Am. Anim. Hosp. Assoc. 2000, 36, 518–521. [Google Scholar] [CrossRef] [PubMed]
- Holmes, K.E.; Thompson, V.; Piskun, C.M.; Kohnken, R.A.; Huelsmeyer, M.K.; Fan, T.M.; Stein, T.J. Canine osteosarcoma cell lines from patients with differing serum alkaline phosphatase concentrations display no behavioural differences in vitro: OSA cell lines differing in serum ALP. Vet. Comp. Oncol. 2015, 13, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Legare, M.E.; Bush, J.; Ashley, A.K.; Kato, T.; Hanneman, W.H. Cellular and phenotypic characterization of canine osteosarcoma cell lines. J. Cancer 2011, 2, 262. [Google Scholar] [CrossRef] [PubMed]
- Fanburgsmith, J.; Bratthauer, G.; Miettinen, M. Osteocalcin and osteonectin immunoreactivity in extraskeletal osteosarcoma: A study of 28 cases. Hum. Pathol. 1999, 30, 32–38. [Google Scholar] [CrossRef]
- Loukopoulos, P.; O’Brien, T.; Ghoddusi, M.; Mungall, B.; Robinson, W. Characterisation of three novel canine osteosarcoma cell lines producing high levels of matrix metalloproteinases. Res. Vet. Sci. 2004, 77, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Mohseny, A.B.; Machado, I.; Cai, Y.; Schaefer, K.-L.; Serra, M.; Hogendoorn, P.C.W.; Llombart-Bosch, A.; Cleton-Jansen, A.-M. Functional characterization of osteosarcoma cell lines provides representative models to study the human disease. Lab. Investig. 2011, 91, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Martins-Neves, S.R.; Lopes, Á.O.; do Carmo, A.; Paiva, A.A.; Simões, P.C.; Abrunhosa, A.J.; Gomes, C.M. Therapeutic implications of an enriched cancer stem-like cell population in a human osteosarcoma cell line. BMC Cancer 2012, 12, 139. [Google Scholar] [CrossRef] [PubMed]
- Meyer, F.R.L.; Steinborn, R.; Grausgruber, H.; Wolfesberger, B.; Walter, I. Expression of platelet-derived growth factor BB, erythropoietin and erythropoietin receptor in canine and feline osteosarcoma. Vet. J. 2015, 206, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Morello, E.; Martano, M.; Buracco, P. Biology, diagnosis and treatment of canine appendicular osteosarcoma: Similarities and differences with human osteosarcoma. Vet. J. 2011, 189, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Pool, R.R.; Thompson, K.G. Tumors of joints. In Tumors in Domestic Animals; Meuten, D.J., Ed.; Iowa State Press: Ames, IA, USA, 2002; pp. 199–243. [Google Scholar]
- Mueller, F.; Fuchs, B.; Kaser-Hotz, B. Comparative biology of human and canine osteosarcoma. Anticancer Res. 2007, 27, 155–164. [Google Scholar] [PubMed]
- Chang, Y.S.; Graves, B.; Guerlavais, V.; Tovar, C.; Packman, K.; To, K.-H.; Olson, K.A.; Kesavan, K.; Gangurde, P.; Mukherjee, A.; Baker, T.; et al. Stapled α-helical peptide drug development: A potent dual inhibitor of MDM2 and MDMX for p53-dependent cancer therapy. Proc. Natl. Acad. Sci. USA 2013, 110, 3445–3454. [Google Scholar] [CrossRef] [PubMed]
- Couto, J.I.; Bear, M.D.; Lin, J.; Pennel, M.; Kulp, S.K.; Kisseberth, W.C.; London, C.A. Biologic activity of the novel small molecule STAT3 inhibitor LLL12 against canine osteosarcoma cell lines. BMC Vet. Res. 2012, 8, 244. [Google Scholar] [CrossRef] [PubMed]
- Lourda, M.; Trougakos, I.P.; Gonos, E.S. Development of resistance to chemotherapeutic drugs in human osteosarcoma cell lines largely depends on up-regulation of Clusterin/Apolipoprotein J. Int. J. Cancer 2007, 120, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Montanini, L.; Lasagna, L.; Barili, V.; Jonstrup, S.P.; Murgia, A.; Pazzaglia, L.; Conti, A.; Novello, C.; Kjems, J.; Perris, R.; et al. MicroRNA cloning and sequencing in osteosarcoma cell lines: Differential role of miR-93. Cell. Oncol. Dordr. 2012, 35, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Sharili, A.-S.; Allen, S.; Smith, K.; Price, J.; McGonnell, I.M. Snail2 promotes osteosarcoma cell motility through remodelling of the actin cytoskeleton and regulates tumor development. Cancer Lett. 2013, 333, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Rimann, M.; Laternser, S.; Gvozdenovic, A.; Muff, R.; Fuchs, B.; Kelm, J.M.; Graf-Hausner, U. An in vitro osteosarcoma 3D microtissue model for drug development. J. Biotechnol. 2014, 189, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Majeska, R.J.; Rodan, S.B.; Rodan, G.A. Parathyroid hormone-responsive clonal cell lines from rat osteosarcoma. Endocrinology 1980, 107, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Mohseny, A.B.; Hogendoorn, P.C.W.; Cleton-Jansen, A.-M. Osteosarcoma Models: From cell lines to Zebrafish. Sarcoma 2012. [Google Scholar] [CrossRef] [PubMed]
- Mather, J.P.; Roberts, P.E. Introduction to Cell and Tissue Culture: Theory and Technique; Springer Science & Business Media: Berlin, Germany, 2007. [Google Scholar]
- Hiddemann, W.; Roessner, A.; Wörmann, B.; Mellin, W.; Klockenkemper, B.; Bösing, T.; Büchner, T.; Grundmann, E. Tumor heterogeneity in osteosarcoma as identified by flow cytometry. Cancer 1987, 59, 324–328. [Google Scholar] [CrossRef]
- Kunz, P.; Fellenberg, J.; Moskovszky, L.; Sápi, Z.; Krenacs, T.; Poeschl, J.; Lehner, B.; Szendrõi, M.; Ewerbeck, V.; Kinscherf, R.; et al. Osteosarcoma microenvironment: Whole-slide imaging and optimized antigen detection overcome major limitations in immunohistochemical quantification. PLoS ONE 2014, 9, e90727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalla-Torre, C.A.; Yoshimoto, M.; Lee, C.-H.; Joshua, A.M.; de Toledo, S.R.; Petrilli, A.S.; Andrade, J.A.; Chilton-MacNeill, S.; Zielenska, M.; Squire, J.A. Effects of THBS3, SPARC and SPP1 expression on biological behavior and survival in patients with osteosarcoma. BMC Cancer 2006, 6, 237. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, E.; Hirayama, K.; Matsuda, K.; Okamoto, M.; Ohmachi, T.; Kadosawa, T.; Taniyama, H. Diversity of histologic patterns and expression of cytoskeletal proteins in canine skeletal osteosarcoma. Vet. Pathol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Kastan, M.B.; Canman, C.E.; Leonard, C.J. P53, cell cycle control and apoptosis: Implications for cancer. Cancer Metastasis Rev. 1995, 14, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Donehower, L.A.; Harvey, M.; Slagle, B.L.; McArthur, M.J.; Montgomery, C.A.; Butel, J.S.; Bradley, A. Mice deficient for p53 are developmentally normal but susceptible to spontaneous tumours. Nature 1992, 356, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Bongiovanni, L.; Mazzocchetti, F.; Malatesta, D.; Romanucci, M.; Ciccarelli, A.; Buracco, P.; de Maria, R.; Palmieri, C.; Martano, M.; Morello, E.; et al. Immunohistochemical investigation of cell cycle and apoptosis regulators (survivin, β-catenin, p53, caspase 3) in canine appendicular osteosarcoma. BMC Vet. Res. 2012, 8, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasir, L.; Rutteman, G.R.; Reid, S.W.; Schulze, C.; Argyle, D.J. Analysis of p53 mutational events and MDM2 amplification in canine soft-tissue sarcomas. Cancer Lett. 2001, 174, 83–89. [Google Scholar] [CrossRef]
- Mayr, B.; Reifinger, M.; Loupal, G. Polymorphisms in feline tumour suppressor gene p53. Mutations in an osteosarcoma and a mammary carcinoma. Vet. J. 1998, 155, 103–106. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Resulting Cell Line | Species | Sex a | Age | Breed | OS Subtype | Location |
|---|---|---|---|---|---|---|---|
| 1033 | COS_1033 | Dog | F | 7y 1m | Boxer | Teleangiectatic OS with giant cell participation | Lung metastasis |
| 1077 | FOS_1077 | Cat | F | 10y | Domestic shorthair | FibroblasticOS | Costa |
| 1140 | FOS_1140 | Cat | M ° | n/s | Domestic shorthair | Osteoblastic OS | Tibia |
| 1186 | COS_1186w COS_1186h | Dog | M ° | 11y 2m | Dachshund | Osteoblastic OS | Scapula |
| 1189 | COS_1189 | Dog | M ° | 7y 3m | Mixed breed | Poorly defined OS/ signs of osteo-or synovialsarkom | Humerus |
| 1220 | COS_1220 | Dog | F ° | 5y 3m | Boxer | Fibroblastic OS | Radius |
| Primary Antibody | Clone | Dilution | Antigen Retrieval | Source | |
|---|---|---|---|---|---|
| Osteopontin | polyclonal | 1:75 | Tissue | 30 min steamed in citric acid buffer pH 6.0 | Biogenesis, Poole, UK |
| rabbit | Cell pellet | 2x 5 min microwaved in citric acid buffer pH 6.0 | |||
| Osteocalcin | polyclonal | 1:400 | Tissue | 30 min steamed in citric acid buffer pH 6.0 | Biogenesis |
| rabbit | Cell pellet | 2x 5 min microwaved in citric acid buffer pH 6.0 | |||
| Cytokeratin | monoclonal | 1:250 | Tissue | 30 min steamed in Tris-EDTA buffer pH 9.0 | Cell Marque, Rocklin, CA, USA |
| mouse | Cell pellet | 2x 5 min microwaved in Tris-EDTA buffer pH 9.0 | |||
| Vimentin | monoclonal | 1:200 | Tissue | 20 min steamed in citric acid buffer pH 6.0 | Dako, Glostrup, Denmark |
| mouse | Cell pellet | 2x 5 min microwaved in citric acid buffer pH 6.0 | |||
| Alkaline Phosphatase | polyclonal | Dog: 1:100 | Tissue | 30 min steamed in citric acid buffer pH 6.0 | Genetex, Irvine, CA, USA |
| rabbit | Cat: 1:250 | Cell pellet | 2x 5 min microwaved in citric acid buffer pH 6.0 | ||
| Osteonectin | polyclonal | 1:1500 | Tissue | none | Millipore, Billerica, MA, USA |
| rabbit | Cell pellet | none | |||
| p53 | monoclonal | 1:90 | Tissue | 3x 5 min microwaved in Tris-EDTA buffer pH 9.0 | Enzo Life Sciences, Lausen, Switzerland |
| mouse | Cell pellet | 2x 5 min microwaved in Tris-EDTA buffer pH 9.0 | |||
| Gene / Symbol | Species | NCBI Ref Nr (Dog/Cat) | Foward Primer | Reverse Primer | Length (bp) |
|---|---|---|---|---|---|
| Osteocalcin / | Dog | XM_547536.4 | GCTGGTCCAGCAGATGCAA | CCCAGCCCAGAGTCCAGGTA | 125 |
| BGLAP | Cat | XM_003999711.3 | GCCCGGCAGATGCAAAG | CCCTCCTGCTTGGACACGA | 70 |
| Osteopontin/ | Dog | XM_003434023.2 | ACTGACATTCCAGCAACCCAA | CACAAGTGATGTGAAGTCCTCCTCT | 168 |
| SPP1 | Cat | XM_006930977.2 | CAATTTTTCACCCCAGCTGTC | CACAAGTGATGTGAAGTCCTCCTCT | 150 |
| Osteonectin / | Dog | XM_005619272.1 | CACCCTGGAAGGCACCAA | CGCAGAGGGAATTCAGTCAGC | 108 |
| SPARC | Cat | XM_003981374.3 | CCAAGAAGGGCCACAAACTC | GGAATTCGGTCAGCTCGGA | 87 |
| Alkaline phosphatase / | Dog | XM_005617214.1 | GGCCTGAACCTCATCGACAT | GCGGTTCCAGACGTAGTGAGA | 72 |
| ALPL | Cat | NM_001042563.1 | GGACGGCCTGAACCTCG | GAGTTCGGT GCGGTTCCA | 85 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meyer, F.R.L.; Walter, I. Establishment and Characterization of New Canine and Feline Osteosarcoma Primary Cell Lines. Vet. Sci. 2016, 3, 9. https://doi.org/10.3390/vetsci3020009
Meyer FRL, Walter I. Establishment and Characterization of New Canine and Feline Osteosarcoma Primary Cell Lines. Veterinary Sciences. 2016; 3(2):9. https://doi.org/10.3390/vetsci3020009
Chicago/Turabian StyleMeyer, Florian R. L., and Ingrid Walter. 2016. "Establishment and Characterization of New Canine and Feline Osteosarcoma Primary Cell Lines" Veterinary Sciences 3, no. 2: 9. https://doi.org/10.3390/vetsci3020009