3.1. Optimization of β-Glucosidase Extraction from Germinated Soybean Epicotyls
The linear effect of the variable
X1 (temperature) on the response function
y (β-glucosidase activity) was not significant, whereas the linear effect of the variable
X2 (pH) was significant. In contrast, the quadratic effects of the variables
X1 and
X2 were significant but the interaction between the variables
X1 and
X2 was not significant (
Table 3). These results indicated that variable
X2 (pH between 3.6 and 6.4) was essential to obtaining extracts with high β-glucosidase activity from epicotyls from germinated soybeans.
The determination coefficient (
R2) of 0.94 indicates that 94% of the experimental data fitted the model. The polynomial model (
Y) representing the activity of β-glucosidase of extracts from EGS is described below:
where
x1 and
x2 are the coded variables representing the temperature and pH, respectively, for the optimum recovery of high-activity β-glucosidase extracts from EGS.
According to the results in
Table 4, extracts obtained at 30 or 35 °C and low pH values of 3.6 or 4.0 (assays 10 and 3, respectively) showed lower β-glucosidase activity. In contrast, higher β-glucosidase activity was found in extracts obtained at 23 °C and pH 5.0 (assay 8), 25 °C and pH 6.0 (assay 2), 30 °C and pH 6.4 (assay 11), and 37 °C and pH 5.0 (assay 9). Finally, the response surface (
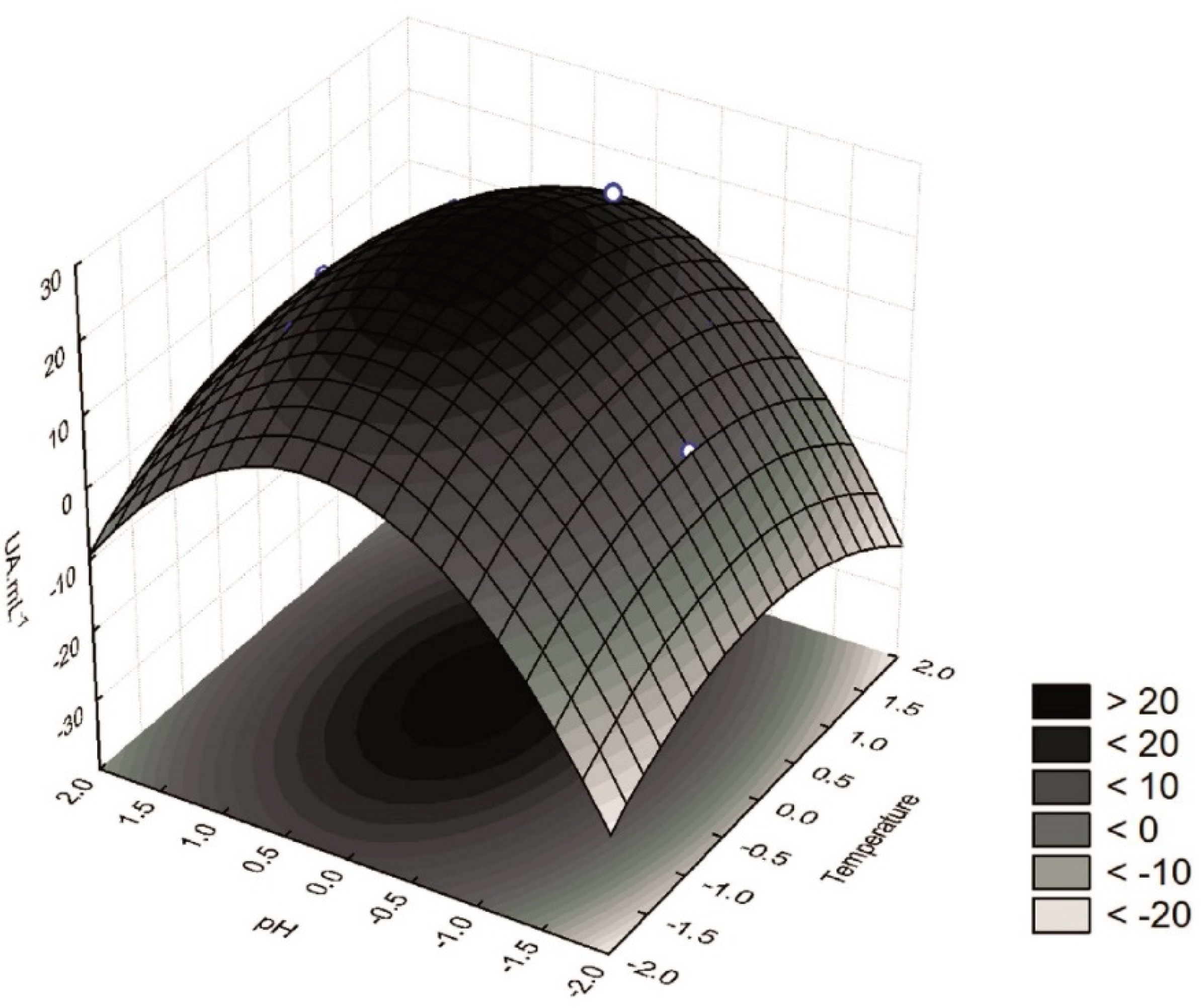
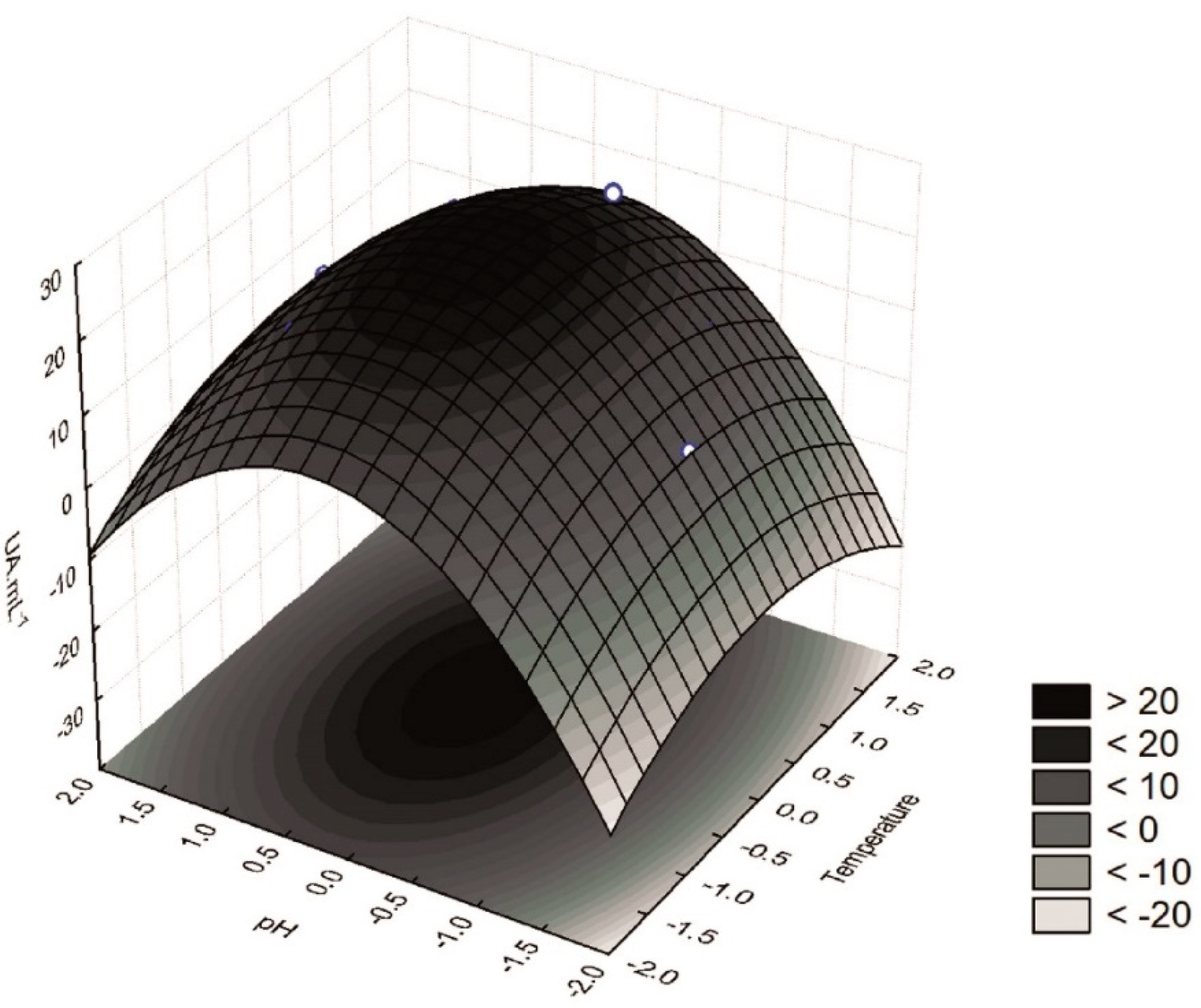
Figure 1) shows that the highest β-glucosidase activity was obtained when the extractions were performed at the central point, i.e., at 30 °C and pH 5.0 (assays 5, 6, and 7). Furthermore, extracts with high β-glucosidase activity could also be obtained at temperatures ranging from 27.5 to 33.5 °C and pH from 4.9 to 5.5. The conditions used as the central point were also in good agreement with the desirability parameter of the proposed model (
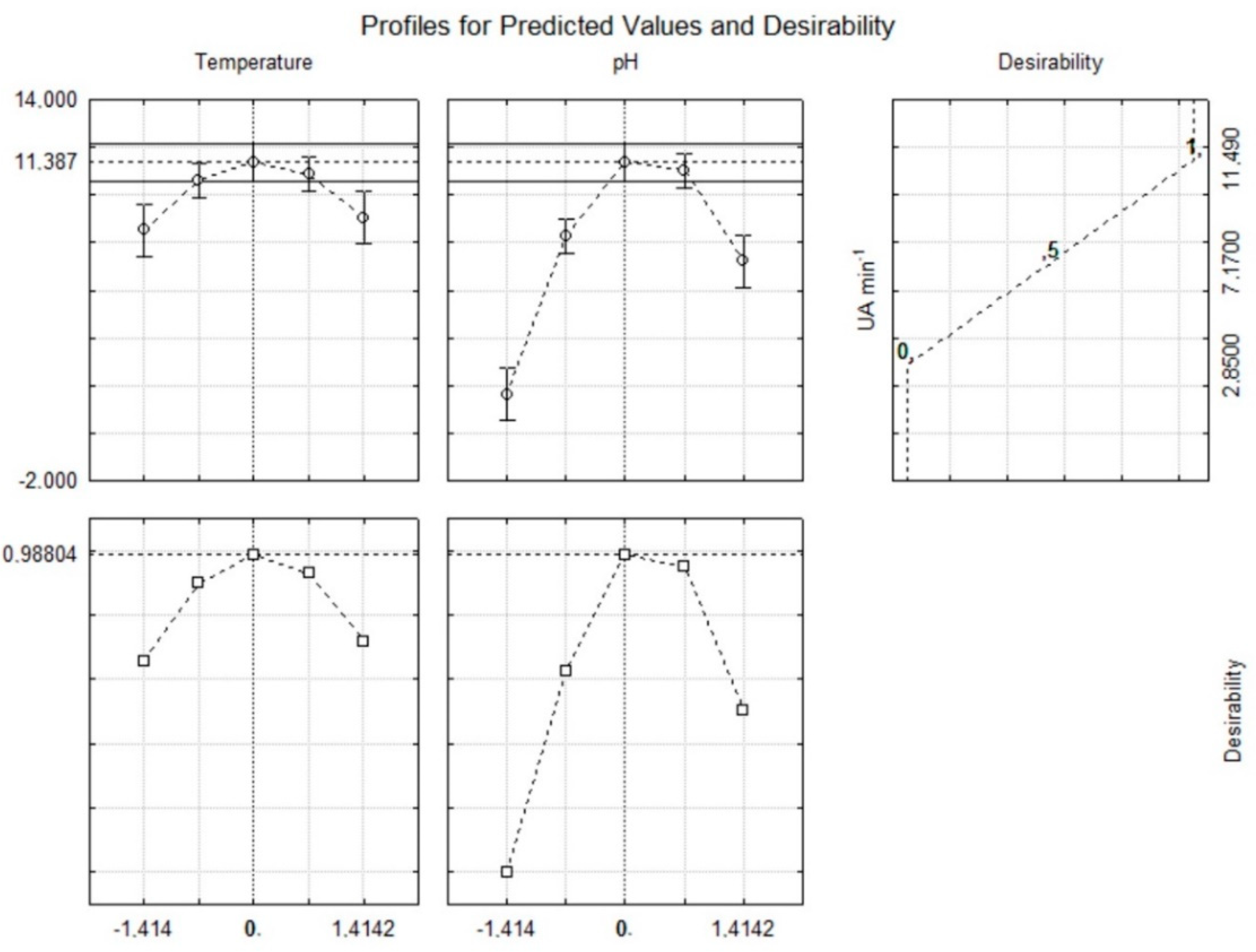
Figure 2), thus supporting the procurement of extracts with a high β-glucosidase activity at 30 °C and pH 5.0.
Matsuura and Obata [
26] extracted, partially purified, and characterized β-glucosidases from soybean cotyledons under different conditions including lower temperatures (between −10 and 5 °C). Although the extraction of β-glucosidase from cotyledons has been studied by these authors, to the best of our knowledge, the procurement of extracts with high β-glucosidase activity from germinated soybean epicotyls has not been reported in the literature. Furthermore, our proposed model proved to be advantageous to establish the best conditions to obtain extracts with high enzyme activity. In addition, working at 30 °C offers more operational advantages compared to the lower temperatures (between −10 and 5 °C) reported in the literature [
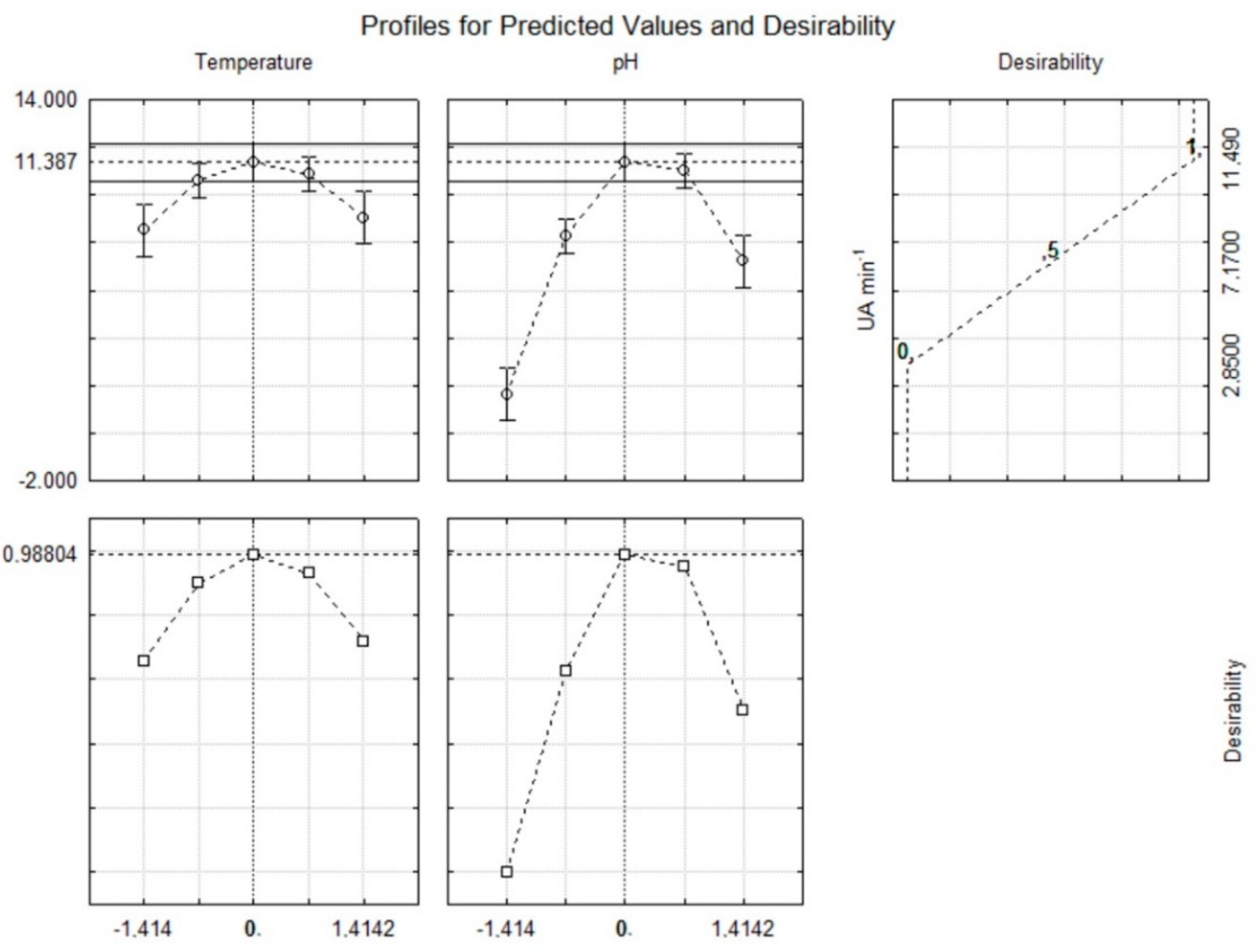
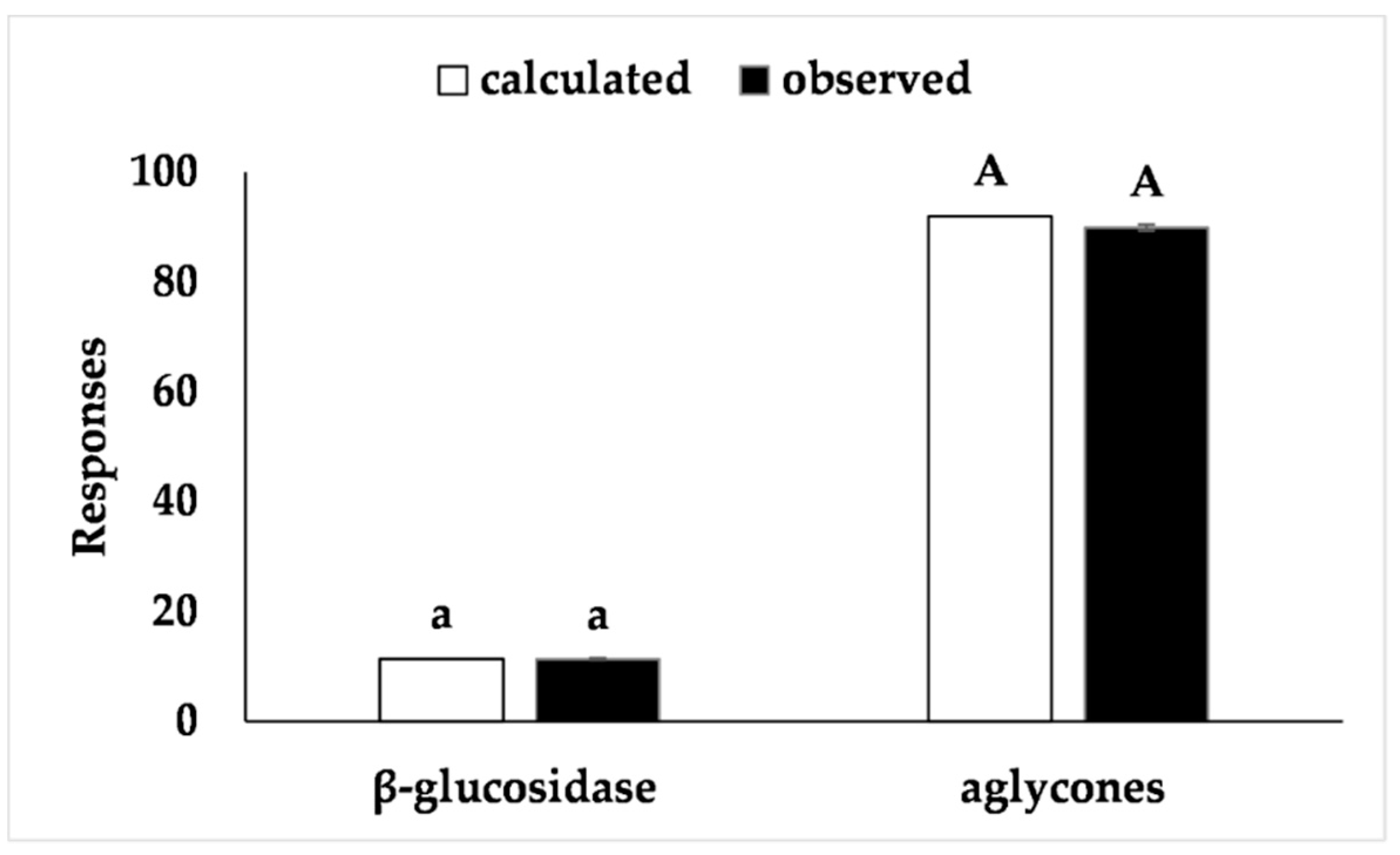
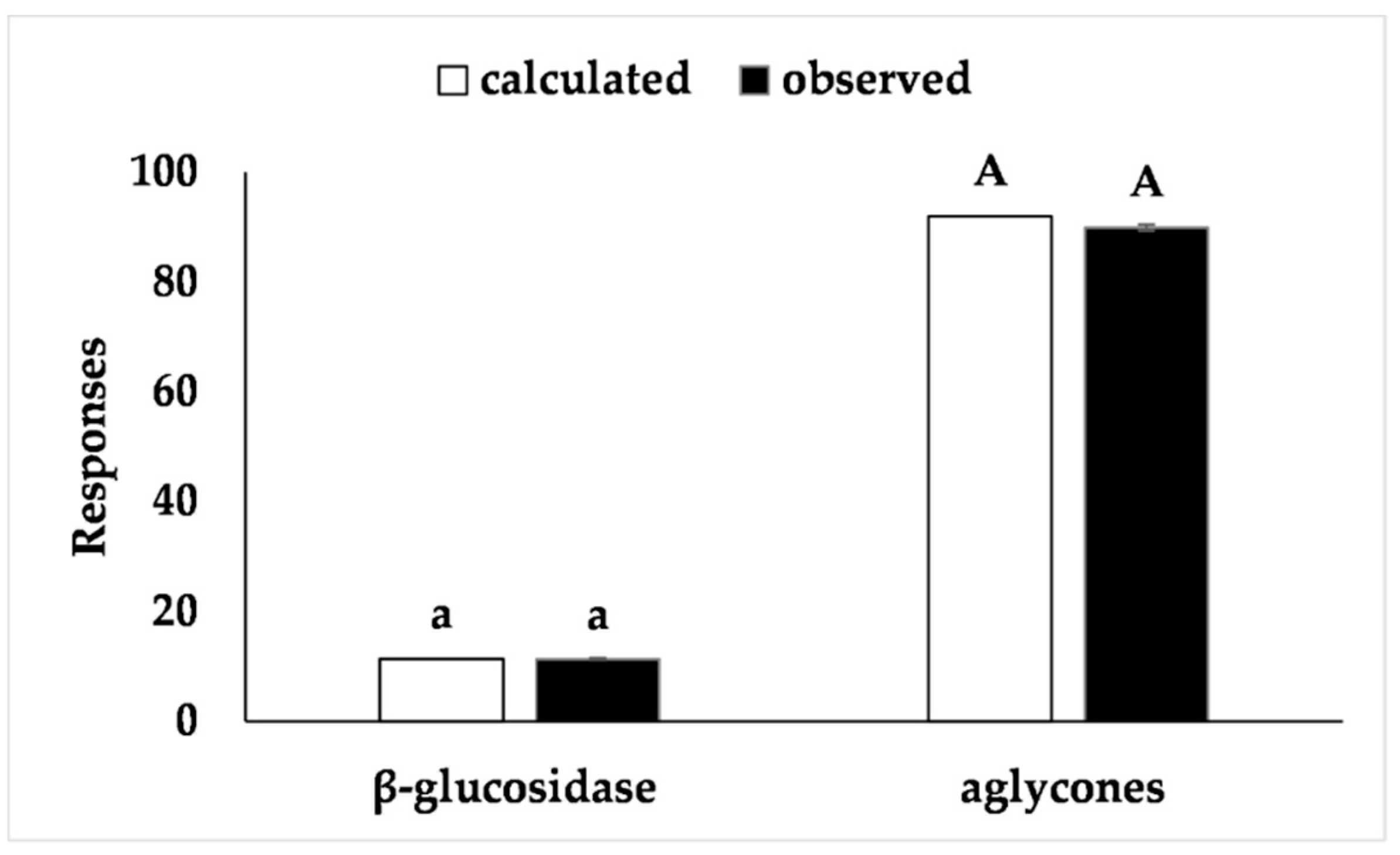
26]. The proposed model was validated with an additional experiment under optimal conditions and the results (Figure 4) fell within the confidence interval of the estimated response, thus confirming the validity of the model.
3.2. Optimization of the Recovery of Aglycones Using β-Glucosidase from Germinated Soybean Epicotyls
According to variance analysis (ANOVA), the linear effect of the variable
X4 (pH) and quadratic effects of the variables
X3 (°C) and
X4 (pH) were significant. However, the effects of the block, linear variable
X3 (°C), and the interaction of the variables
X3 (°C) and
X4 (pH) on the response (
W = % aglycones) were not significant (
Table 5). Therefore, nonsignificant terms were excluded from the model which did not affect the
R2. The coefficient of determination (
R2) was 0.86, in other words, 86% of experimental data can be explained by the model. Therefore, Equation (3) is described as follows:
where
W = % aglycones and
x3 and
x4 are the coded variables representing the temperature and pH, respectively.
The aglycone content of the soybean cotyledon flour (BRS 257) devoid of β-glucosidase addition was 2.9% and, regardless of the temperature (
X3), reached values higher than 47% (
Table 6) with greater pH (
X4) values (0, +1, and +1.41). This suggests a greater pH influence on the response function (
W = % of aglycones), thus supporting the screening of the main effects.
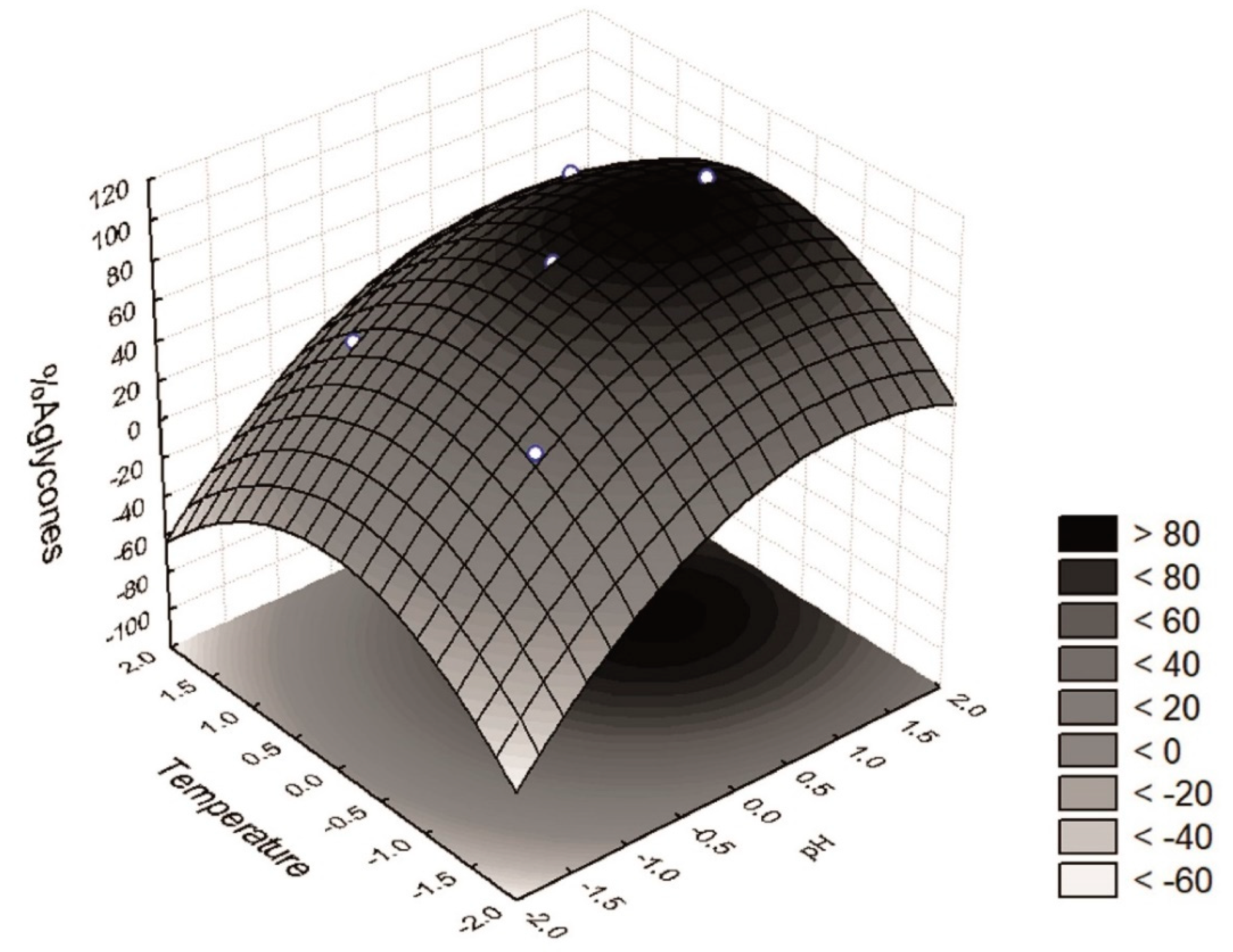
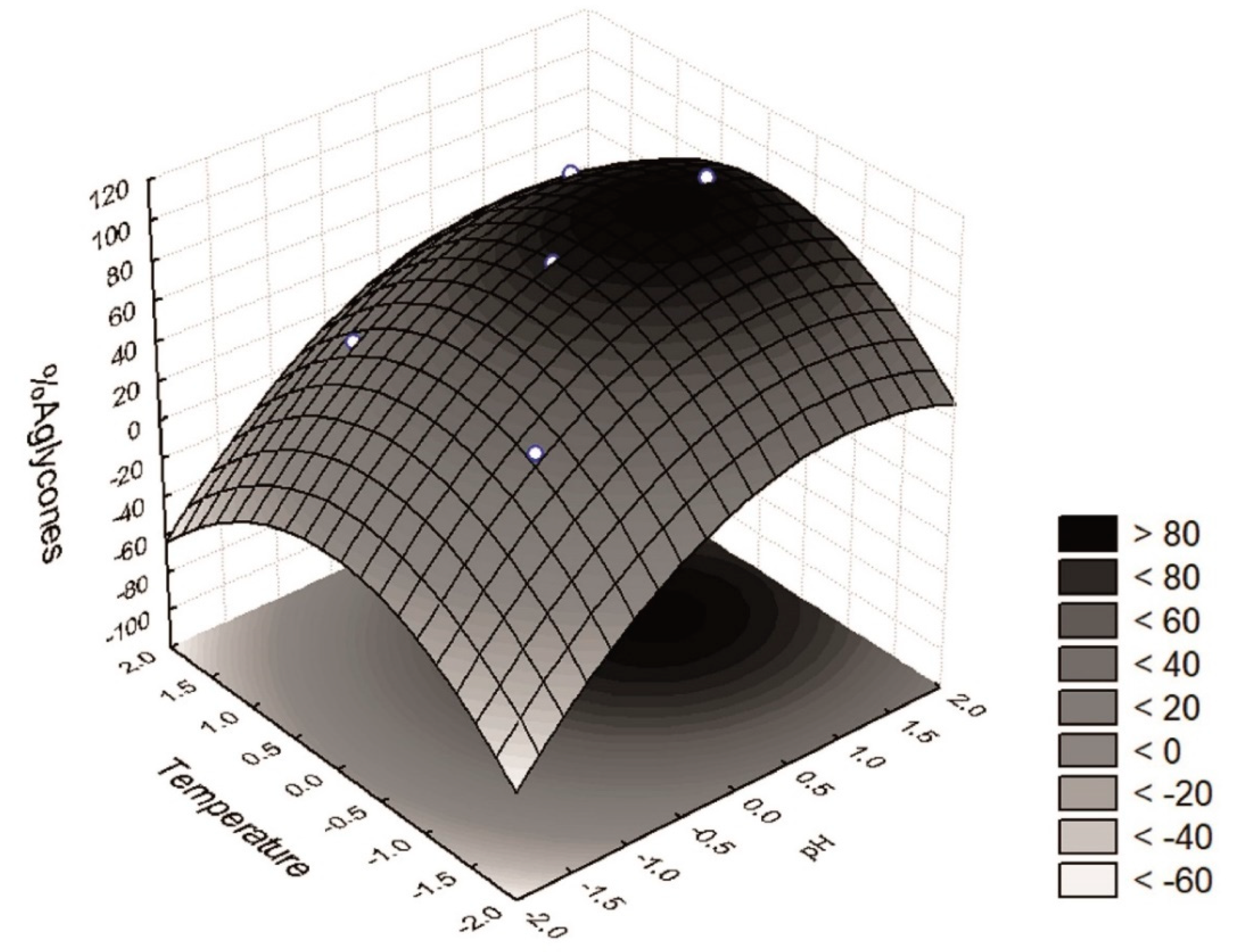
The response surface (
Figure 3) shows a region at which more than 80% of the isoflavones recovered were in the aglycone form, i.e., a pH range between +0.5 and +1.5 (6.25 < pH < 7.75) and a temperature between −0.5 and +0.5 (27.5 < T < 42.5). Furthermore, an optimum region at which the maximum percentage of isoflavones in the aglycone form was recovered could be determined. The desirability parameter indicates two optimal points of maximum (
W = 98.7% aglycones), i.e., when
x3 = 0 (35 °C) and
x4 = +1 or +1.41 (pH = 7.00 and 7.61). One of these test points coincides with assay 9 (
Z = 98.7% aglycones). These results are distinct from the optimum conditions of pH (between 5.2 and 6.0) and temperature (50 °C) for β-glucosidase activity from soybean cell tissue described by Hosel and Todenhagen [
29] and by Matsuura and Obata [
30] who reported optimal activity at pH 4.5 and a temperature of 45 °C for a soybean β-glucosidase. However, the proposed models were validated for both response functions which were within the confidence interval of the model (
Figure 4).
A complete conversion of glycosidic isoflavones into their aglycones has been described by other authors [
31]. However, their study was conducted with β-glucosidase from microbial origin (
Lactobacillus (L.)
rhamnosus CRL981). Furthermore, Song and Yu [
32] applied β-glucosidase from
Thermotoga maritima to recombinant soybean flour and obtained an almost complete conversion of all isoflavone glycosides. Endogenous β-glucosidases represent a more secure source of β-glucosidases than microbial sources. However, endogenous β-glucosidases have been poorly explored when compared with the microbial conversion of conjugated isoflavones into their aglycone forms. As mentioned before, in the present study, the maximum aglycone isoflavones obtained was 98.7% when endogenous β-glucosidase from EGS was used under the two conditions described above, therefore providing strong evidence on the potential use of EGS as a great source of endogenous β-glucosidase.
Several pieces of evidence have demonstrated that different plant parts may present different phenolic compounds [
33,
34,
35,
36,
37,
38,
39], therefore suggesting that the expression of genes associated with the production of some enzymes may be tissue specific. Likewise, not only the absorption of the phenolics is important but also their structure/activity [
40,
41,
42]. Perera et al. [
40] demonstrated that epigallocatechin gallate derivatives procured via lipophilization showed enhanced antioxidant properties compared with those of epigallocatechin gallate. Likewise, Oh and Shahidi [
42] reported that several lipophilized resveratrol derivatives had better hydrogen peroxide scavenging activity than resveratrol. In addition, Oldoni et al. [
41] demonstrated that minor changes such as the position of the hydroxyl group in flavonoids may be responsible for major changes in their final effects. According to their study, the concentration of procyanidin A2 necessary to scavenge 50% of the DPPH radical was 1.7-fold higher than that of procyanidin A1. Besides the antioxidant activity, which may be related to the inhibition of low-density lipoprotein cholesterol oxidation and DNA damage [
43,
44], phenolic compounds have been regarded as potential inhibitors of enzymes related to the absorption of carbohydrates and lipids [
38,
45]. In a bioactivity-guided isolation and purification study to identify α-glucosidase inhibitors, Sun et al. [
46] suggested that C
1-OH of the saccharide moiety in phenolic glycosides are necessary for a potent inhibition of intestinal α-glucosidases. According to Bustanji et al. [
47], the inhibitory effect towards lipase activity was in the order of gallic acid > caffeic acid > chlorogenic acid > rosmarinic acid. A recent study [
37] also demonstrated that whereas proanthocyanidin-rich extracts showed higher antioxidant activity, the extracts containing only phenolic acids showed higher antimicrobial effects. Finally, inflammation has been linked to several health issues, including those related to oxidative stress. A myriad of phenolic compounds have been reported to act as potential anti-inflammatory and antioxidant compounds [
39,
48,
49,
50] and the link between inflammatory responses and several diseases is well recognized. According to Lee et al. [
51], fermented soymilk with greater contents of isoflavones in the aglycone form also showed higher antioxidant properties. The importance of natural product characterization in studies related to their bioactivity has long been discussed [
52,
53]. Therefore, by understanding the best conditions to produce β-glucosidase from EGS as well as its further application in the procurement of high-aglycone defatted soybean cotyledon flours, the present study contributes to both basic and applied science related to food bioactives and health.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}