
Precise Authenticity of Quinoa, Coix Seed, Wild Rice and Chickpea Components Using Optimized TaqMan Real-Time PCR
 and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Materials
2.2. Extraction of Genome DNA
2.3. qPCR Primer and Probe Design
2.4. Real-Time qPCR
2.5. Specificity Test
2.6. Sensitivity and Amplification Efficiency Test
2.7. Robustness Evaluation
2.8. Detection of Commercial Samples
2.9. Quantification Performance of qPCR Methods
3. Result and Discussion
3.1. Specific Analysis of Primer and Probe
3.2. Specificity Tests
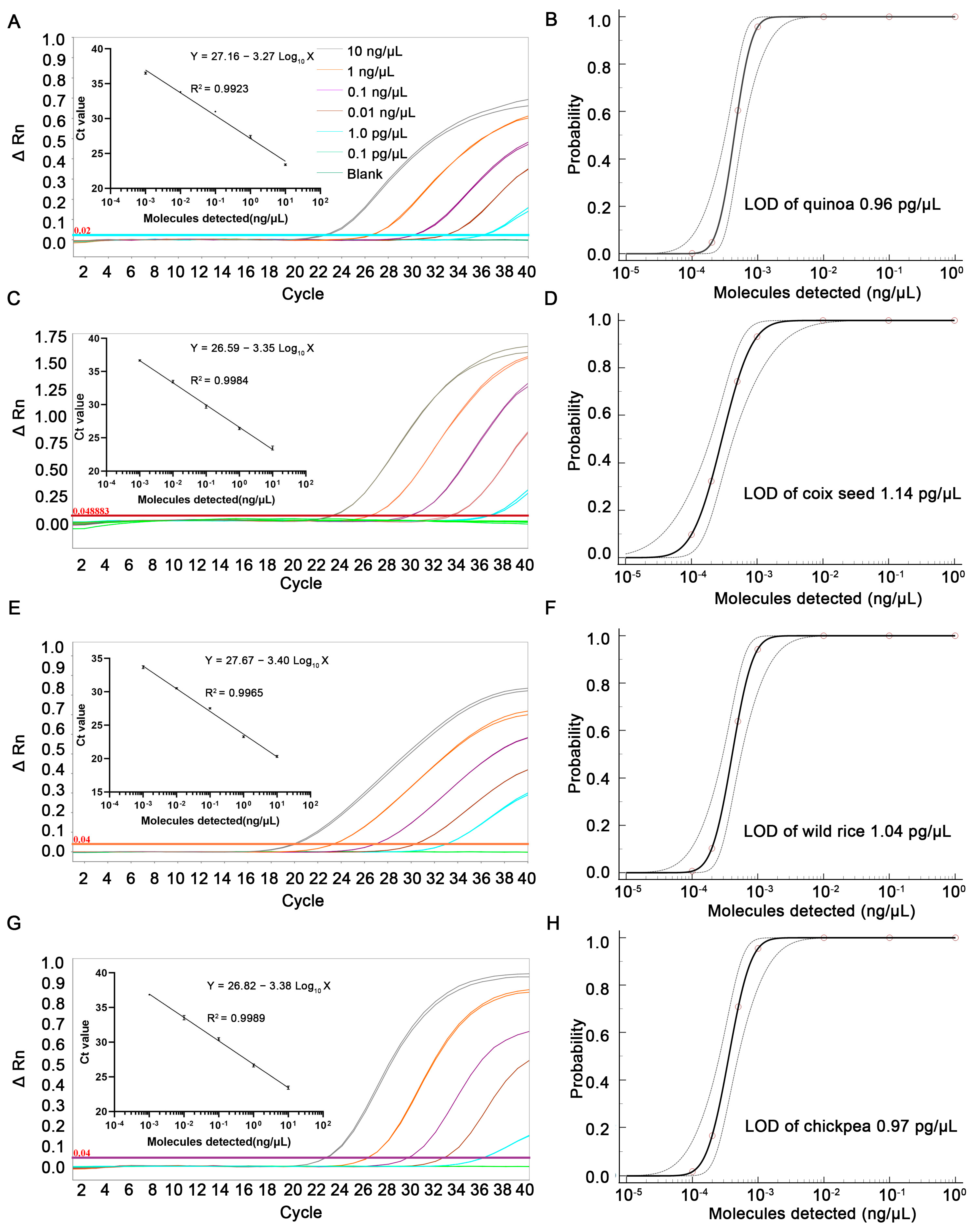
3.3. Sensitivity Test
3.4. Quantification Performance of qPCR Methods
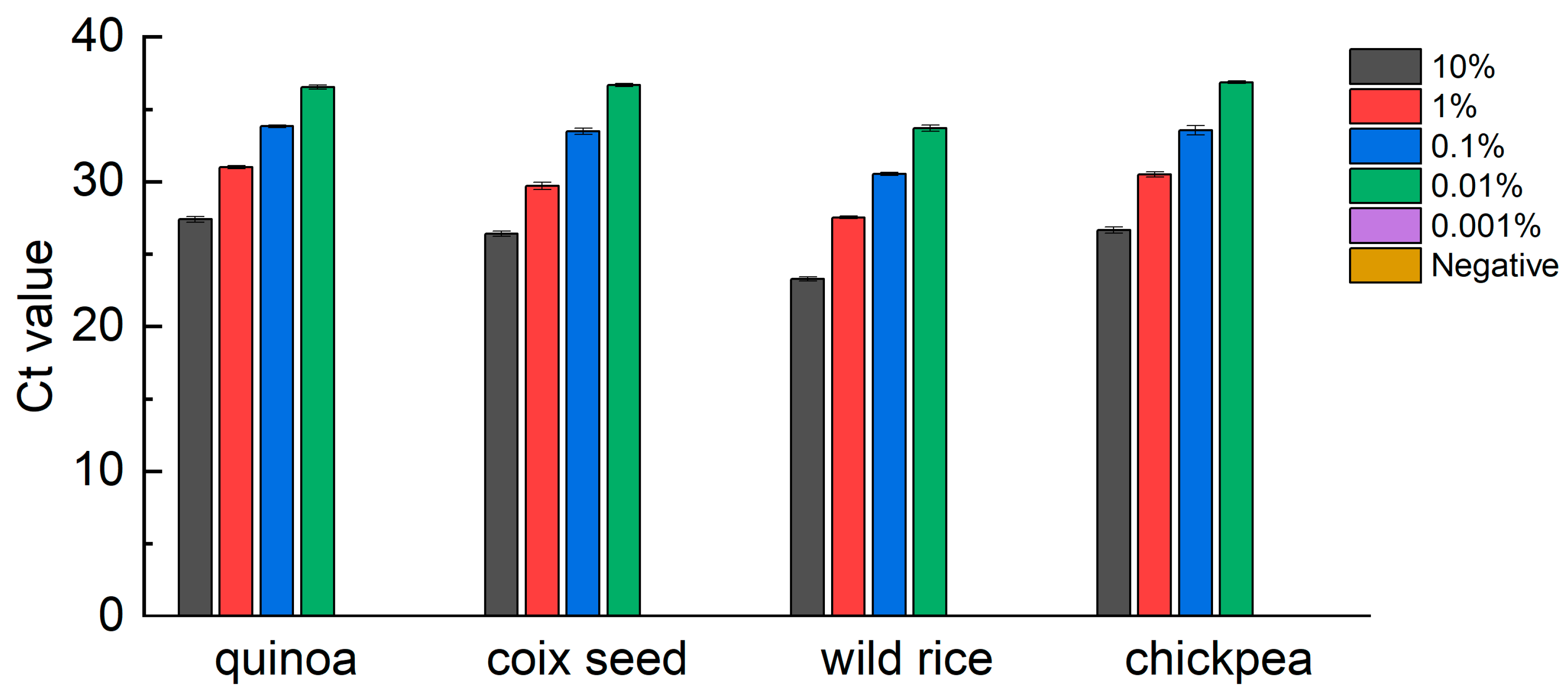
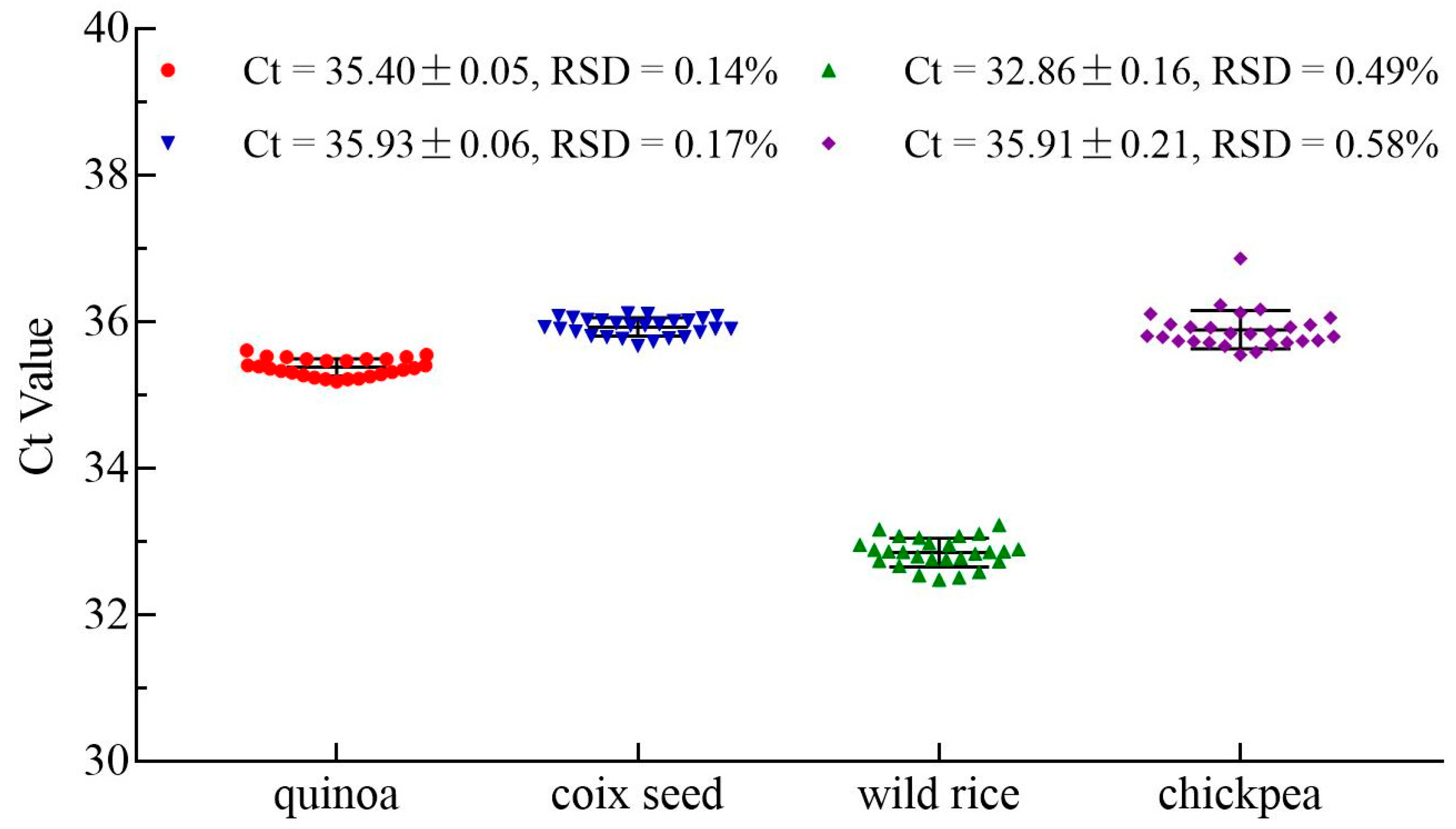
3.5. Robustness Test
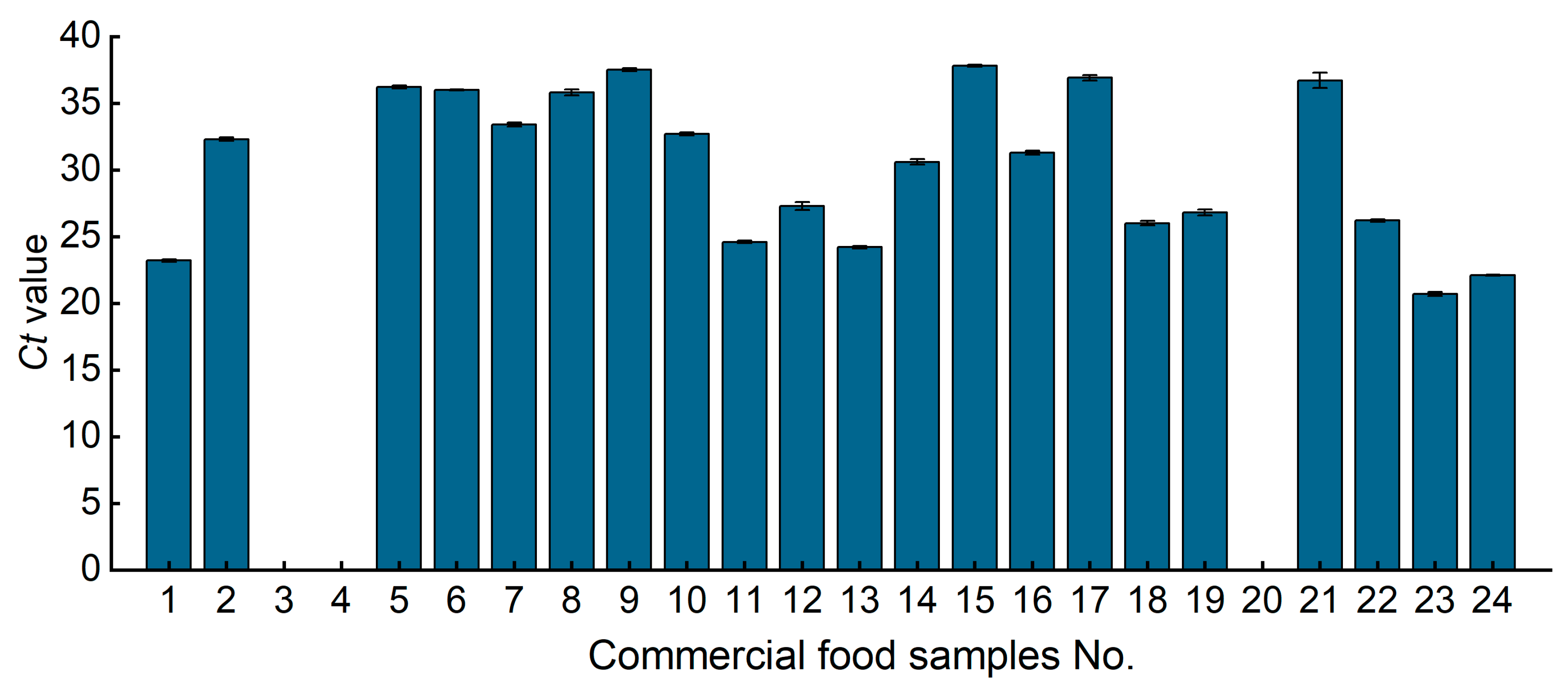
3.6. Analysis of Commercial Samples
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Filho, A.M.; Pirozi, M.R.; Borges, J.T.; Pinheiro Sant’Ana, H.M.; Chaves, J.B.; Coimbra, J.S. Quinoa: Nutritional, functional, and antinutritional aspects. Crit. Rev. Food Sci. Nutr. 2017, 57, 1618–1630. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Han, P.; Li, Y.; Wang, W.; Lai, D.; Zhou, L. Quinoa Secondary Metabolites and Their Biological Activities or Functions. Molecules 2019, 24, 2512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, E.; Cadavez, V.; Barros, L.; Encina-Zelada, C.; Stojkovic, D.; Sokovic, M.; Calhelha, R.C.; Gonzales-Barron, U.; Ferreira, I. Chenopodium quinoa Willd. (quinoa) grains: A good source of phenolic compounds. Food Res. Int. 2020, 137, 109574. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, J.A.; Reganold, J.P.; Murphy, K.M.; Carpenter-Boggs, L.A. A Plant-Fungus Bioassay Supports the Classification of Quinoa (Chenopodium quinoa Willd.) as Inconsistently Mycorrhizal. Microb. Ecol. 2021, 82, 135–144. [Google Scholar] [CrossRef]
- Abugoch James, L.E. Chapter 1 Quinoa (Chenopodium quinoa Willd.): Composition, chemistry, nutritional, and functional properties. Adv. Food Nutr. Res. 2009, 58, 1–31. [Google Scholar] [CrossRef]
- Angeli, V.; Miguel Silva, P.; Crispim Massuela, D.; Khan, M.W.; Hamar, A.; Khajehei, F.; Graeff-Honninger, S.; Piatti, C. Quinoa (Chenopodium quinoa Willd.): An Overview of the Potentials of the “Golden Grain” and Socio-Economic and Environmental Aspects of Its Cultivation and Marketization. Foods 2020, 9, 216. [Google Scholar] [CrossRef] [Green Version]
- Miao, G.; Qin, Y.; Guo, J.; Zhang, Q.; Bao, Y. Transcriptome characterization and expression profile of Coix lacryma-jobi L. in response to drought. PLoS ONE 2021, 16, e0256875. [Google Scholar] [CrossRef]
- Kang, S.H.; Kim, B.; Choi, B.S.; Lee, H.O.; Kim, N.H.; Lee, S.J.; Kim, H.S.; Shin, M.J.; Kim, H.W.; Nam, K.; et al. Genome Assembly and Annotation of Soft-Shelled Adlay (Coix lacryma-jobi Variety ma-yuen), a Cereal and Medicinal Crop in the Poaceae Family. Front. Plant Sci. 2020, 11, 630. [Google Scholar] [CrossRef]
- Fu, Y.H.; Yang, C.; Meng, Q.; Liu, F.; Shen, G.; Zhou, M.; Ao, M. Genetic Diversity and Structure of Coix lacryma-jobi L. from Its World Secondary Diversity Center, Southwest China. Int. J. Genom. 2019, 2019, 9815697. [Google Scholar] [CrossRef] [Green Version]
- Han, S.F.; Zhang, H.; Zhai, C.K. Protective potentials of wild rice (Zizania latifolia (Griseb) Turcz) against obesity and lipotoxicity induced by a high-fat/cholesterol diet in rats. Food Chem. Toxicol. 2012, 50, 2263–2269. [Google Scholar] [CrossRef]
- Cho, S.; Kays, S.J. Aroma-active compounds of wild rice (Zizania palustris L.). Food Res. Int. 2013, 54, 1463–1470. [Google Scholar] [CrossRef]
- Yan, N.; Du, Y.; Liu, X.; Chu, C.; Shi, J.; Zhang, H.; Liu, Y.; Zhang, Z. Morphological Characteristics, Nutrients, and Bioactive Compounds of Zizania latifolia, and Health Benefits of Its Seeds. Molecules 2018, 23, 1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tikapunya, T.; Henry, R.J.; Smyth, H. Evaluating the sensory properties of unpolished Australian wild rice. Food Res. Int. 2018, 103, 406–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acevedo Martinez, K.A.; Yang, M.M.; Gonzalez de Mejia, E. Technological properties of chickpea (Cicer arietinum): Production of snacks and health benefits related to type-2 diabetes. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3762–3787. [Google Scholar] [CrossRef] [PubMed]
- Nisa, Z.U.; Arif, A.; Waheed, M.Q.; Shah, T.M.; Iqbal, A.; Siddiqui, A.J.; Choudhary, M.I.; El-Seedi, H.R.; Musharraf, S.G. A comparative metabolomic study on desi and kabuli chickpea (Cicer arietinum L.) genotypes under rainfed and irrigated field conditions. Sci. Rep. 2020, 10, 13919. [Google Scholar] [CrossRef]
- Lord, N.; Flores Elizondo, C.J.; Spencer, J. The dynamics of food fraud: The interactions between criminal opportunity and market (dys)functionality in legitimate business. Criminol. Crim. Justic. 2017, 17, 605–623. [Google Scholar] [CrossRef] [Green Version]
- Goh, M.S.; Lam, S.D.; Yang, Y.; Naqiuddin, M.; Addis, S.N.K.; Yong, W.T.L.; Luang-In, V.; Sonne, C.; Ma, N.L. Omics technologies used in pesticide residue detection and mitigation in crop. J. Hazard. Mater. 2021, 420, 126624. [Google Scholar] [CrossRef]
- von Bargen, C.; Brockmeyer, J.; Humpf, H.-U. Meat Authentication: A New HPLC–MS/MS Based Method for the Fast and Sensitive Detection of Horse and Pork in Highly Processed Food. J. Agric. Food Chemistr. 2014, 62, 9428–9435. [Google Scholar] [CrossRef]
- Shah, S.M.S.; Ullah, F.; Munir, I. Biochemical characterization for determination of genetic distances among different indigenous chickpea (Cicer arietinum L.) varieties of North-West Pakistan. Braz. J. Biol. 2021, 81, 977–988. [Google Scholar] [CrossRef]
- Feng, Z.; Ding, C.; Li, W.; Wang, D.; Cui, D. Applications of metabolomics in the research of soybean plant under abiotic stress. Food Chem. 2020, 310, 125914. [Google Scholar] [CrossRef]
- Farag, M.A.; Gad, H.A.; Heiss, A.G.; Wessjohann, L.A. Metabolomics driven analysis of six Nigella species seeds via UPLC-qTOF-MS and GC-MS coupled to chemometrics. Food Chem. 2014, 151, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Petrakis, E.A.; Cagliani, L.R.; Polissiou, M.G.; Consonni, R. Evaluation of saffron (Crocus sativus L.) adulteration with plant adulterants by (1)H NMR metabolite fingerprinting. Food Chem. 2015, 173, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Spanoghe, M.; Godoy Jara, M.; Riviere, J.; Lanterbecq, D.; Gadenne, M.; Marique, T. Development and application of a quantitative real-time PCR assay for rapid detection of the multifaceted yeast Kazachstania servazzii in food. Food Microbiol. 2017, 62, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Tichy, H.V.; Bruhs, A.; Palisch, A. Development of Real-Time Polymerase Chain Reaction Systems for the Detection of So-Called “Superfoods” Chia and Quinoa in Commercial Food Products. J. Agric. Food Chem. 2020, 68, 14334–14342. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, Y.; Liu, M.; Wang, B.; Li, M.; Chen, Y. Molecular Tracing of the Origin of Six Different Plant Species in Bee Honey Using Real-Time PCR. J. AOAC Int. 2017, 100, 744–752. [Google Scholar] [CrossRef]
- Garrido-Maestu, A.; Azinheiro, S.; Fucinos, P.; Carvalho, J.; Prado, M. Highly sensitive detection of gluten-containing cereals in food samples by real-time Loop-mediated isothermal AMPlification (qLAMP) and real-time polymerase chain reaction (qPCR). Food Chem. 2018, 246, 156–163. [Google Scholar] [CrossRef]
- Mano, J.; Nishitsuji, Y.; Kikuchi, Y.; Fukudome, S.I.; Hayashida, T.; Kawakami, H.; Kurimoto, Y.; Noguchi, A.; Kondo, K.; Teshima, R.; et al. Quantification of DNA fragmentation in processed foods using real-time PCR. Food Chem. 2017, 226, 149–155. [Google Scholar] [CrossRef]
- Broeders, S.; Huber, I.; Grohmann, L.; Berben, G.; Taverniers, I.; Mazzara, M.; Roosens, N.; Morisset, D. Guidelines for validation of qualitative real-time PCR methods. Trends Food Sci. Tech. 2014, 37, 115–126. [Google Scholar] [CrossRef]
- Mazzara, M.; Savini, C.; Delobel, C.; Broll, H.; Damant, A.; Paoletti, C.; Van den Eede, G. Definition of Minimum Performance Requirements for Analytical Methods of GMO Testing, European Network of GMO Laboratories(ENGL). JRC Sci. Tech. Rep. 2008. [Google Scholar] [CrossRef]
- Bhunia, R.K.; Chakraborty, A.; Kaur, R.; Gayatri, T.; Bhattacharyya, J.; Basu, A.; Maiti, M.K.; Sen, S.K. Seed-specific increased expression of 2S albumin promoter of sesame qualifies it as a useful genetic tool for fatty acid. metabolic engineering and related transgenic intervention in sesame and other oil seed crops. Plant Mol. Biol. 2014, 86, 351–365. [Google Scholar] [CrossRef]
- Hsiao, E.S.; Lin, L.J.; Li, F.Y.; Wang, M.M.; Liao, M.Y.; Tzen, J.T. Gene Families Encoding Isoforms of Two Major Sesame Seed. Storage Proteins, 11S Globulin and 2S Albumin. J. Agric. Food Chem. 2006, 54, 9544–9550. [Google Scholar] [CrossRef]
- Kachroo, A.; Shanklin, J.; Whittle, E.; Lapchyk, L.; Hildebrand, D.; Kachroo, P. The Arabidopsis stearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis. Plant Mol. Biol. 2007, 63, 257–271. [Google Scholar] [CrossRef]
- Zaborowska, Z.; Starzycki, M.; Femiak, I.; Swiderski, M.; Legocki, A.B. Yellow lupine gene encoding stearoyl-ACP desaturase-organization, expression and potential application. Acta Biochim. Pol. 2002, 49, 29–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.; Zhang, P.; Liu, X.; Liang, J.; Li, W. Genetic structure and diversity of Glehnia littoralis, an endangered medicinal plant in China. Biochem. Syst. Ecol. 2016, 66, 265–271. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Ruwe, H.; Melzer, M.; Junker, A.; Hensel, G.; Tschiersch, H.; Schwenkert, S.; Chamas, S.; Schmitz-Linneweber, C.; Borner, T.; et al. The Arabidopsis AAC Proteins CIL and CIA2 Are Sub-functionalized Paralogs Involved in Chloroplast Development. Front. Plant Sci. 2021, 12, 681375. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Target Gene | Primer and Probe | Sequence (5′ → 3′) | Position | Size (bp) |
|---|---|---|---|---|---|
| quinoa | 2S albumin | forward primer | CCAAGGAAGGGAGCAATACTTAG | 319–341 | 357 |
| reverse primer | ACACTCTCTTAAGCAAATTAGACTTAAC | 648–675 | |||
| probe | FAM-ATGAAGATGATGGTGGAAGATAAAGGGATGATGCA-BHQ1 | 395–429 | |||
| coix seed | SAD | forward primer | GTCTGGGAGCATTTTGCTTGC | 1455–1475 | 134 |
| reverse primer | TCCATGTCCACGCCCAGTTC | 1569–1588 | |||
| probe | FAM-TCTCATAGAGTGATAGGAGAATGTCCGTCTGTGTTC-BHQ1 | 1484–1519 | |||
| wild rice | ITS | forward primer | CGAGAGTCGTGTGGATGTTGT | 227–247 | 238 |
| reverse primer | TGCGGAAGGATCATTGTCGT | 10–29 | |||
| probe | FAM-CGGCGGTCGGTAAGAGGTGTTCC-BHQ1 | 175–197 | |||
| chickpea | CIA-2 | forward primer | AGAAGAAGGTTGTTACGGTGGAG | 1422–1444 | 170 |
| reverse primer | CGGTGCGTCGGAGATAGGA | 1572–1591 | |||
| probe | FAM-GAAGGCGTTCGGAATGCTTGGTCT GATAAA-BHQ1 | 1520–1548 | |||
| Reference | 18SrRNA | forward primer | TCTGCCCTATCAACTTTCGATGGTA | 233–257 | 137 |
| reverse primer | AATTTGCGCGCCT GCTGCCTTCCTT | 345–369 | |||
| probe | FAM-CCGTTTCTCAGGCTCCCTCTCCGGAATCGAACC-BHQ1 | 290–322 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Q.; Yin, X.; Yang, A.; Yu, N.; Xing, R.; Chen, Y.; Deng, R.; Cao, J. Precise Authenticity of Quinoa, Coix Seed, Wild Rice and Chickpea Components Using Optimized TaqMan Real-Time PCR. Foods 2023, 12, 852. https://doi.org/10.3390/foods12040852
Zheng Q, Yin X, Yang A, Yu N, Xing R, Chen Y, Deng R, Cao J. Precise Authenticity of Quinoa, Coix Seed, Wild Rice and Chickpea Components Using Optimized TaqMan Real-Time PCR. Foods. 2023; 12(4):852. https://doi.org/10.3390/foods12040852
Chicago/Turabian StyleZheng, Qiuyue, Xinying Yin, Aifu Yang, Ning Yu, Ranran Xing, Ying Chen, Ruijie Deng, and Jijuan Cao. 2023. "Precise Authenticity of Quinoa, Coix Seed, Wild Rice and Chickpea Components Using Optimized TaqMan Real-Time PCR" Foods 12, no. 4: 852. https://doi.org/10.3390/foods12040852