
Perturbation Analysis of Calcium, Alkalinity and Secretion during Growth of Lily Pollen Tubes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
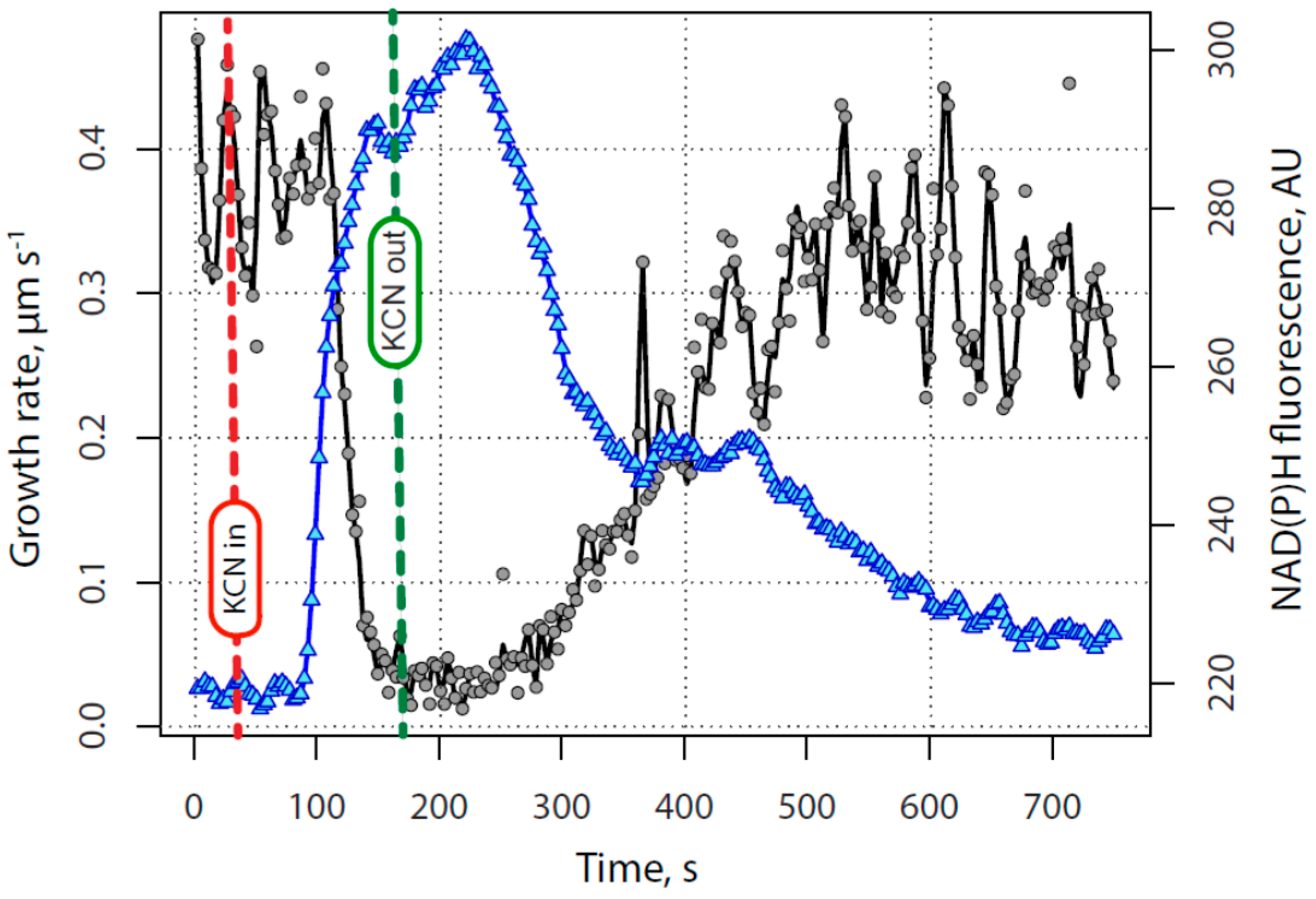
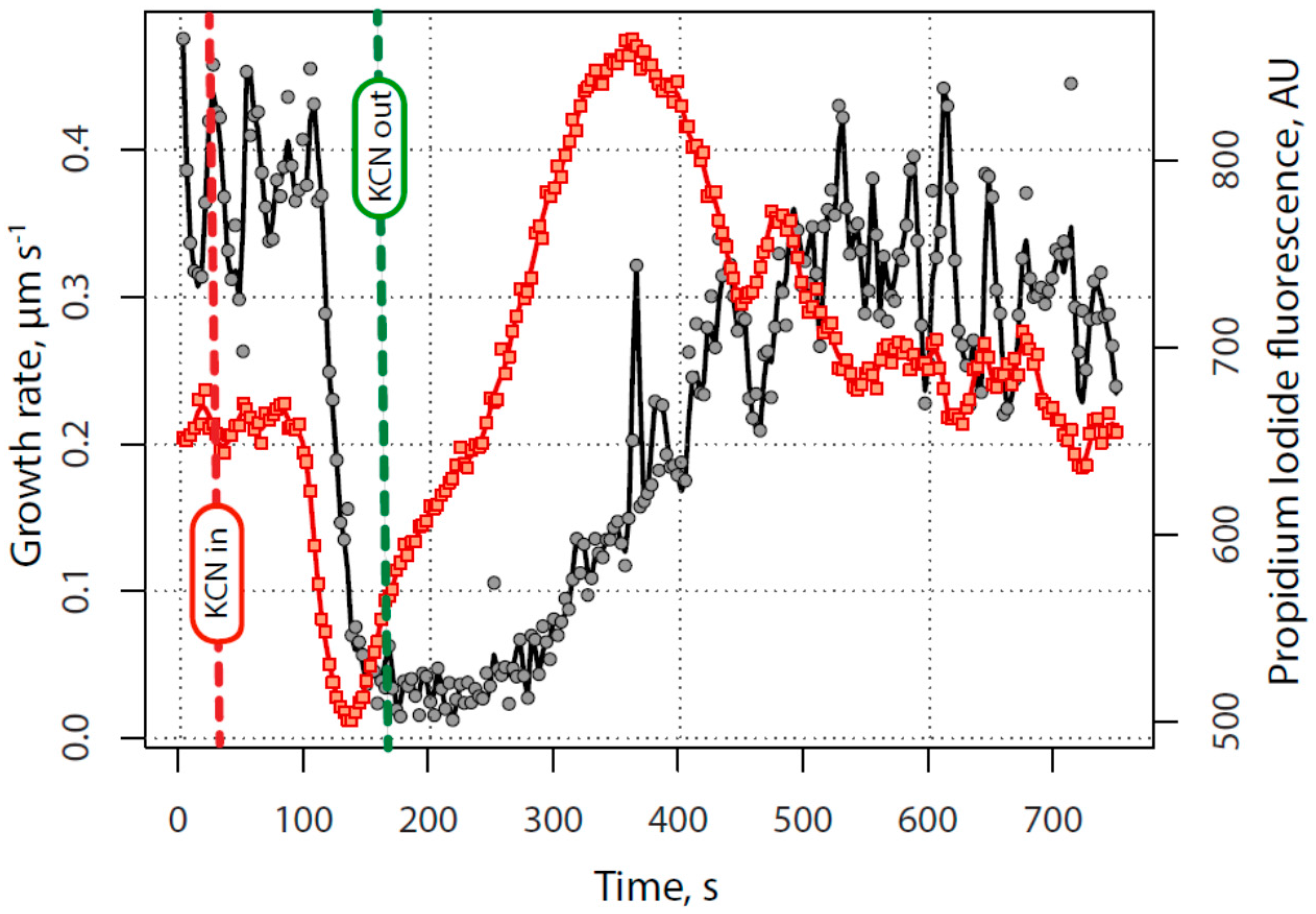
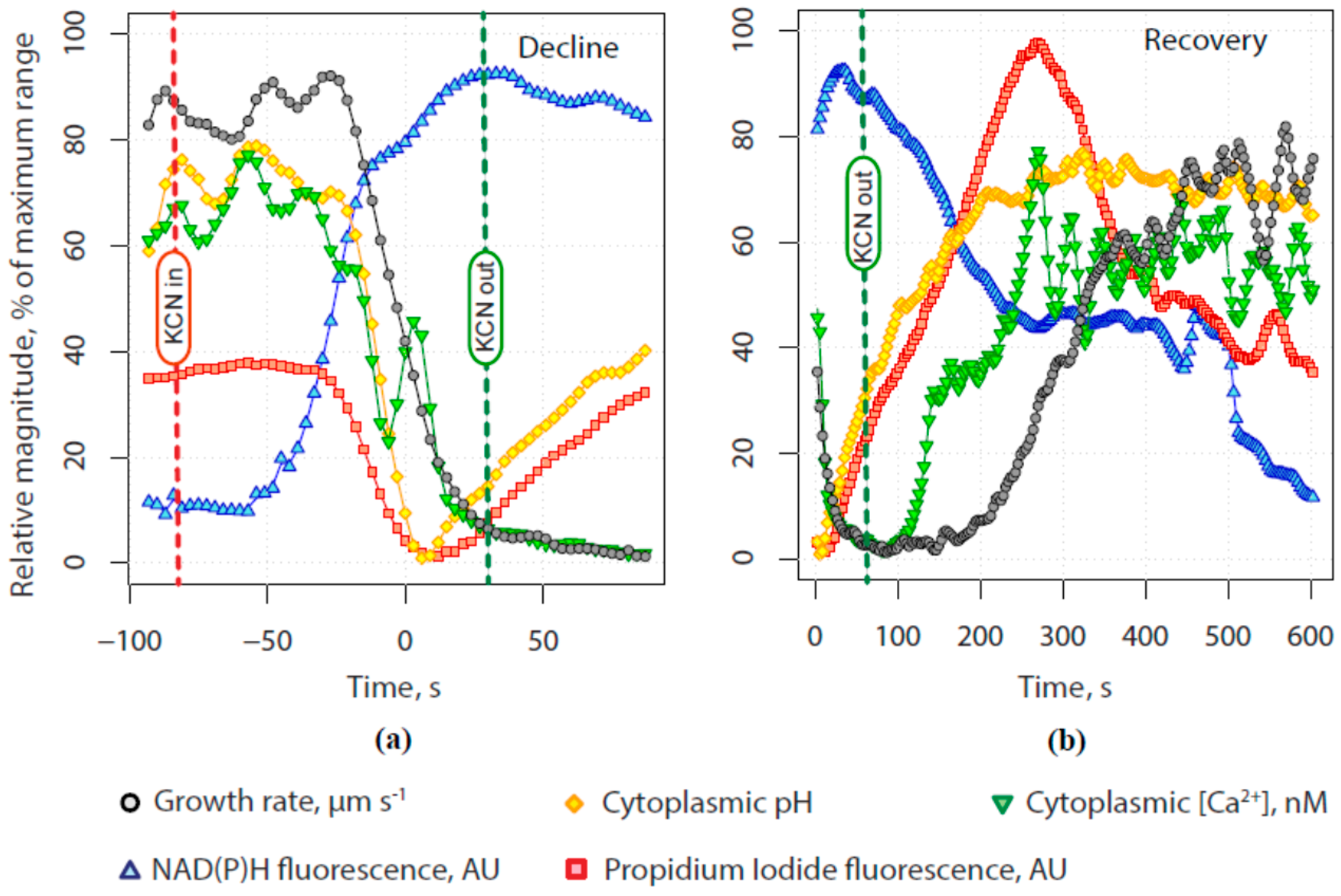
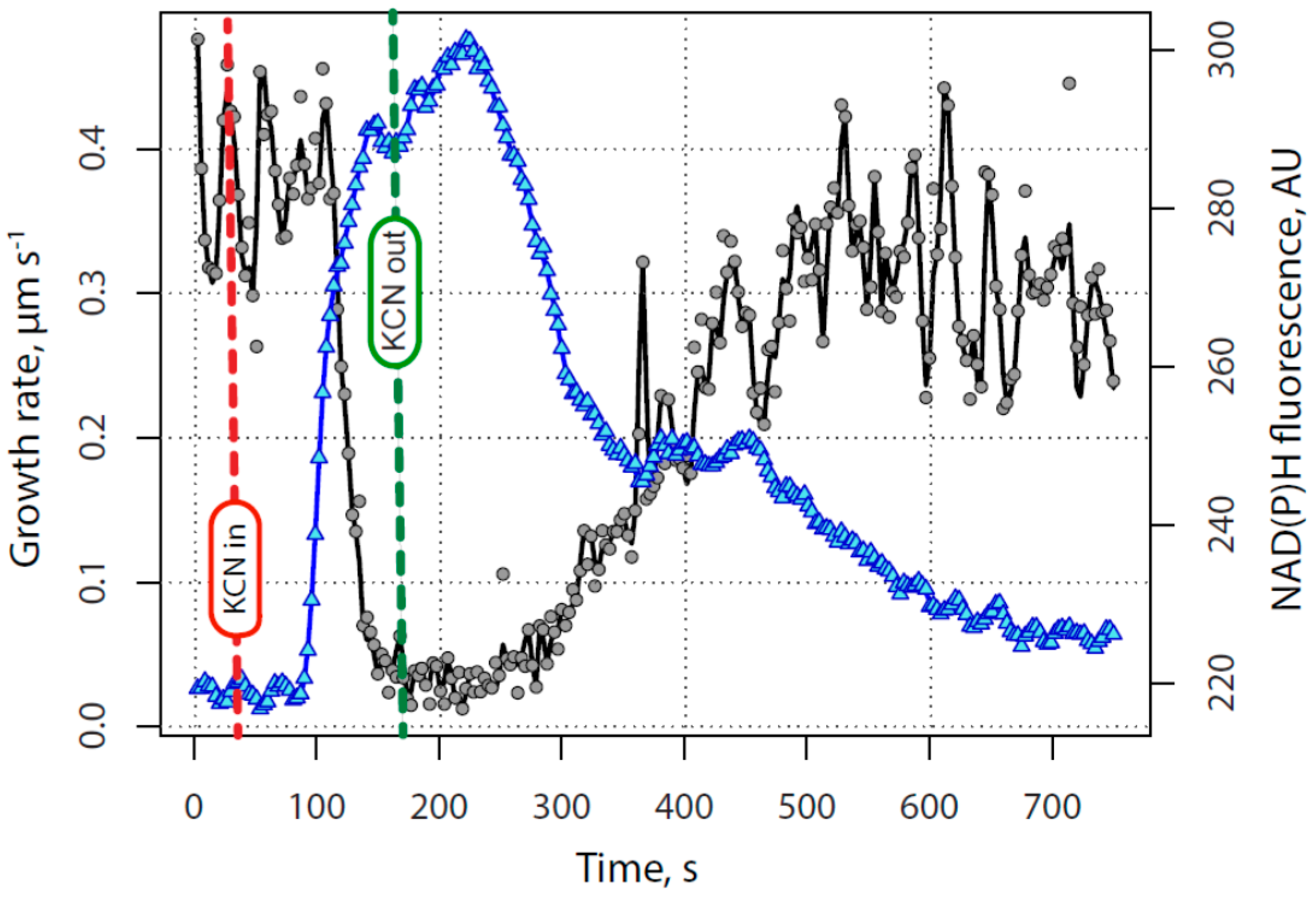
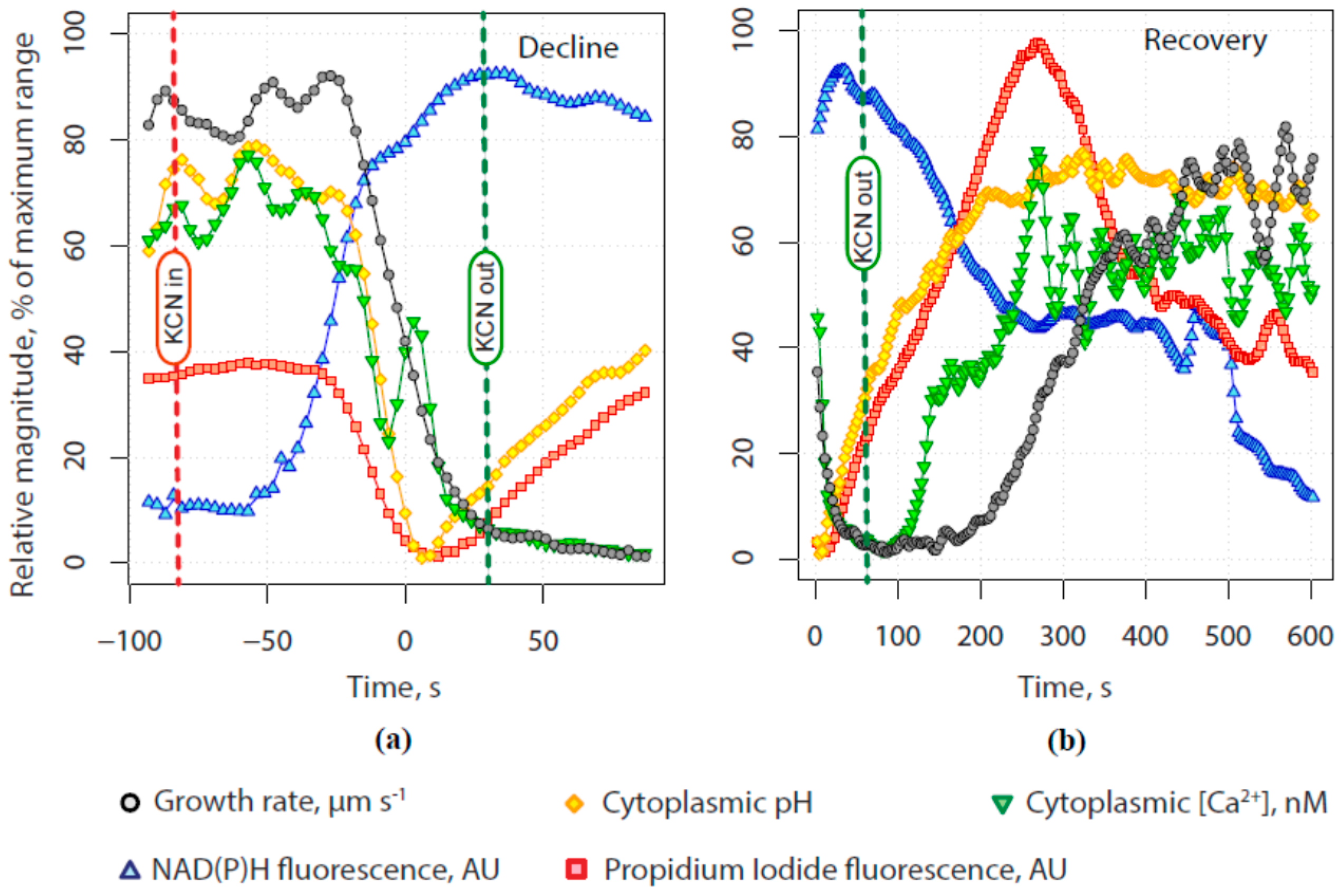
2.1. KCN Reversibly Inhibits Pollen Tube Growth by Blocking the Electron Transport Chain
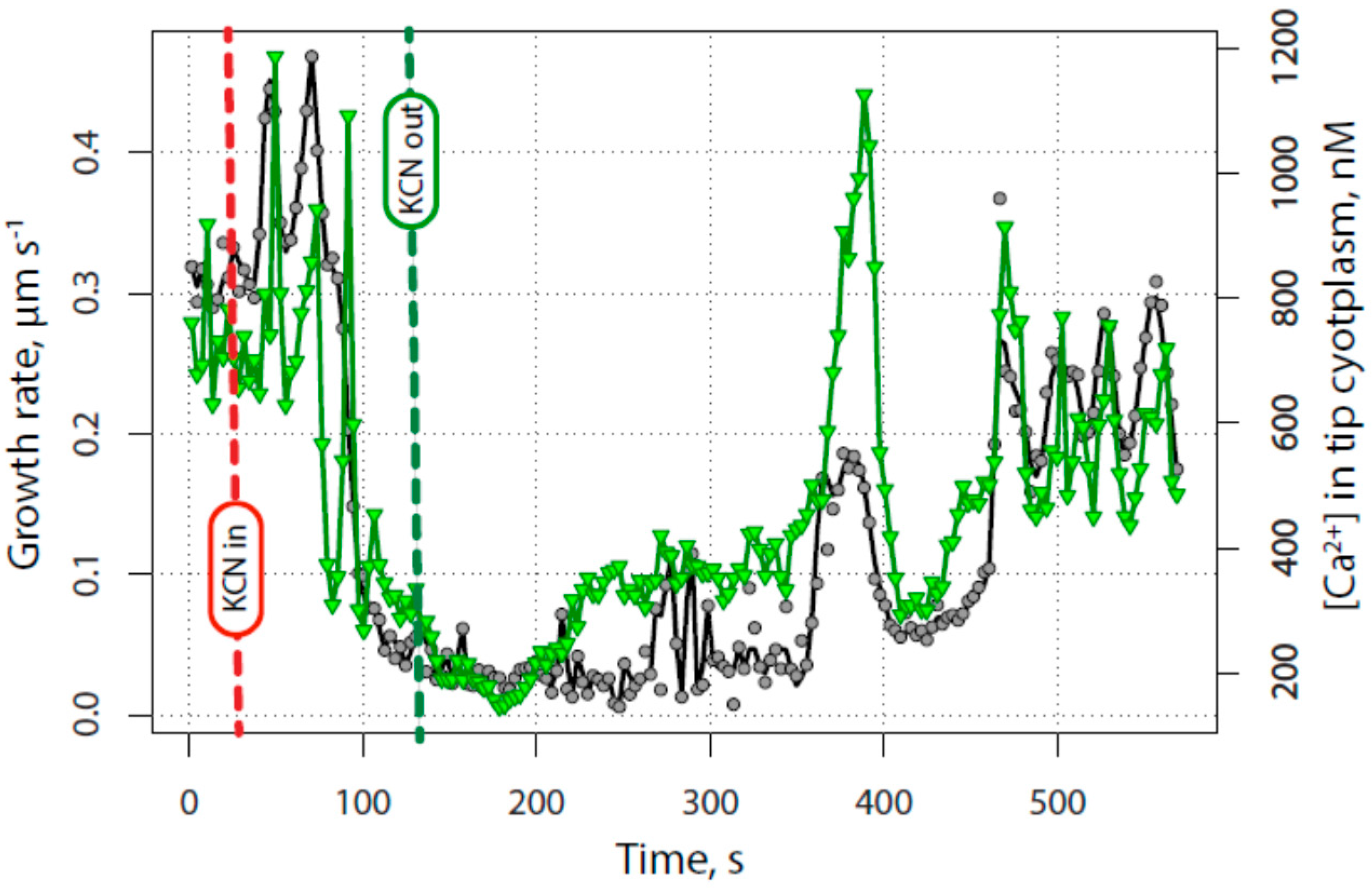
2.2. The Tip-Focused Calcium Gradient Reappears before Pollen Tube Growth Restarts.
2.3. The Alkaline Band Also Appears before Growth and Anticipates Calcium
2.4. Secretion Also Appears before Tube Growth, and before Calcium Increases
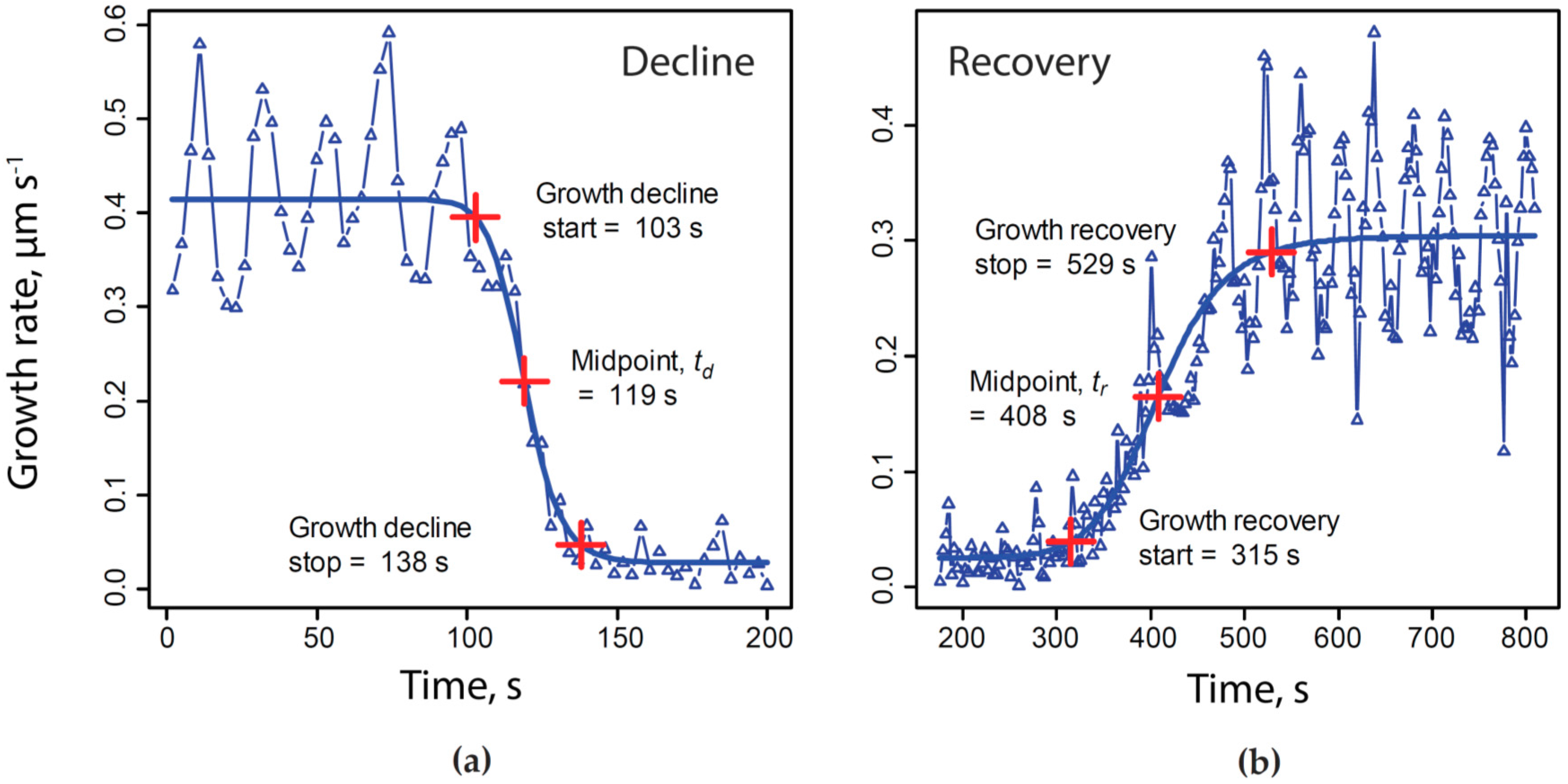
2.5. Quantitative Assessment of the Relationship between Calcium, Alkalinity and Secretion with Growth
3. Discussion
4. Materials and Methods
4.1. Pollen Tube Growth Procedures, and Culture Chamber Construction
4.2. Cell Observation
4.3. Growth Inhibition and Recovery
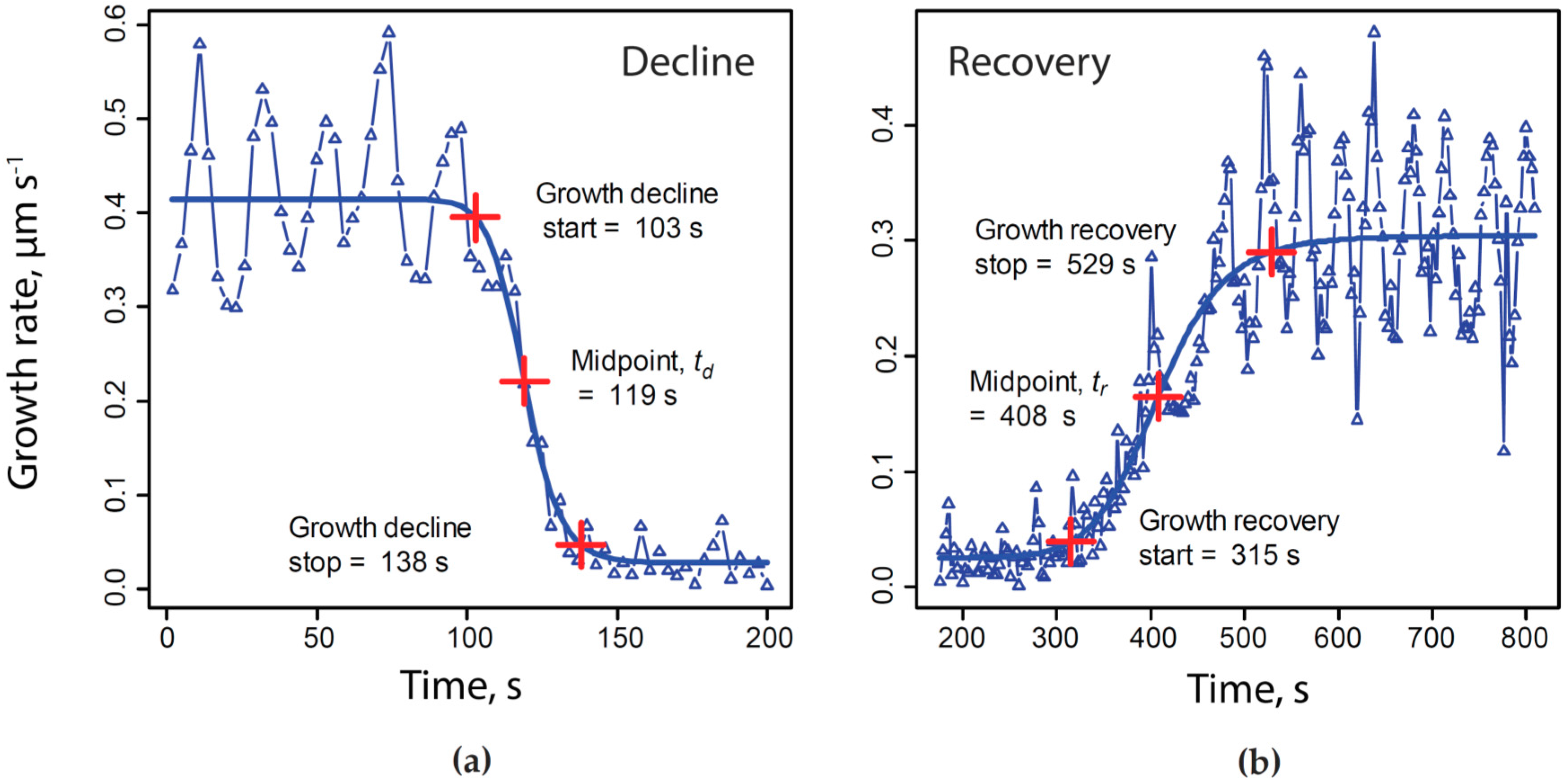
4.4. Normalizing Time Series to a Common Time Base
- where t = elapsed time, s;
- G = growth rate, µm·s−1;
- A = maximum asymptote of growth rate, µm·s−1;
- B = Hill slope;
- C = inflection point;
- D = minimum asymptote of growth rate, µm·s−1.
- = time normalized to growth decline and recovery, s;
- = actual elapsed time during experiment, s;
- = midpoint of KCN-induced growth rate decline, s;
- = midpoint of growth rate recovery following KCN removal, s;
- = mean of across all experiments included in our analysis, e.g., average duration of growth decline due to exposure to 200 µM KCN, s.
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brewbaker, J.L.; Kwack, B.H. The essential role of calcium ion in pollen germination and pollen tube growth. Am. J. Bot. 1963, 50, 859–865. [Google Scholar] [CrossRef]
- Steinhorst, L.; Kudla, J. Calcium—A central regulator of pollen germination and tube growth. Biochim. Biophys. Acta 2013, 1833, 1573–1581. [Google Scholar] [CrossRef] [PubMed]
- Bosch, M.; Hepler, P.K. Pectin methylesterases and pectin dynamics in pollen tubes. Plant Cell 2005, 17, 3219–3226. [Google Scholar] [CrossRef] [PubMed]
- Battey, N.; James, N.; Greenland, A.; Brownlee, C. Exocytosis and endocytosis. Plant Cell 1999, 11, 643–660. [Google Scholar] [CrossRef] [PubMed]
- Sutter, J.U.; Homann, U.; Thiel, G. Ca2+-stimulated exocytosis in maize coleoptile cells. Plant Cell 2000, 12, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Pierson, E.S.; Miller, D.D.; Callaham, D.A.; van Aken, J.; Hackett, G.; Hepler, P.K. Tip-localized calcium entry fluctuates during pollen tube growth. Dev. Biol. 1996, 174, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Holdaway-Clarke, T.L.; Feijo, J.A.; Hackett, G.R.; Kunkel, J.G.; Hepler, P.K. Pollen tube growth and the intracellular cytosolic calcium gradient oscillate in phase while extracellular calcium influx is delayed. Plant Cell 1997, 9, 1999–2010. [Google Scholar] [CrossRef] [PubMed]
- Hepler, P.K.; Lovy-Wheeler, A.; McKenna, S.T.; Kunkel, J.G. Ions and Pollen Tube Growth; Springer: Berlin/Heidelberg, Germany, 2006; pp. 47–69. [Google Scholar]
- Messerli, M.A.; Créton, R.; Jaffe, L.F.; Robinson, K.R. Periodic increases in elongation rate precede increases in cytosolic Ca2+ during pollen tube growth. Dev. Biol. 2000, 222, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas, L.; Lovy-Wheeler, A.; Kunkel, J.G.; Hepler, P.K. Pollen tube growth oscillations and intracellular calcium levels are reversibly modulated by actin polymerization. Plant Physiol. 2008, 146, 1611–1621. [Google Scholar] [CrossRef] [PubMed]
- Rounds, C.M.; Lubeck, E.; Hepler, P.K.; Winship, L.J. Propidium iodide competes with Ca2+ to label pectin in pollen tubes and Arabidopsis root hairs. Plant Physiol. 2011, 157, 175–187. [Google Scholar] [CrossRef] [PubMed]
- McKenna, S.T.; Kunkel, J.G.; Bosch, M.; Rounds, C.M.; Vidali, L.; Winship, L.J.; Hepler, P.K. Exocytosis precedes and predicts the increase in growth in oscillating pollen tubes. Plant Cell 2009, 21, 3026–3040. [Google Scholar] [CrossRef] [PubMed]
- Holdaway-Clarke, T.L.; Weddle, N.M.; Kim, S.; Robi, A.; Parris, C.; Kunkel, J.G.; Hepler, P.K. Effect of extracellular calcium, pH and borate on growth oscillations in Lilium formosanum pollen tubes. J. Exp. Bot. 2003, 54, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Feijó, J.A.; Sainhas, J.; Hackett, G.R.; Kunkel, J.G.; Hepler, P.K. Growing pollen tubes possess a constitutive alkaline band in the clear zone and a growth-dependent acidic tip. J. Cell Biol. 1999, 144, 483–496. [Google Scholar] [CrossRef] [PubMed]
- Lovy-Wheeler, A.; Kunkel, J.G.; Allwood, E.G.; Hussey, P.J.; Hepler, P.K. Oscillatory increases in alkalinity anticipate growth and may regulate actin dynamics in pollen tubes of lily. Plant Cell 2006, 18, 2182–2193. [Google Scholar] [CrossRef] [PubMed]
- Rounds, C.M.; Winship, L.J.; Hepler, P.K. Pollen tube energetics: Respiration, fermentation and the race to the ovule. AoB PLANTS 2011. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, B.; Arango, M.; Oufattole, M.; Crouzet, J.; Purnelle, B.; Boutry, M. Identification of a nicotiana plumbaginifolia plasma membrane H+-ATPase gene expressed in the pollen tube. Plant Mol. Biol. 2005, 58, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Certal, A.C.; Almeida, R.B.; Carvalho, L.M.; Wong, E.; Moreno, N.; Michard, E.; Carneiro, J.; Rodriguéz-Léon, J.; Wu, H.-M.; Cheung, A.Y.; et al. Exclusion of a proton ATPase from the apical membrane is associated with cell polarity and tip growth in Nicotiana tabacum pollen tubes. Plant Cell 2008, 20, 614–634. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, D.B. Inhibition of Pollen Respiration by Oligomycin. Nature 1966, 210, 1362–1363. [Google Scholar] [CrossRef]
- Pertl, H.; Pöckl, M.; Blaschke, C.; Obermeyer, G. Osmoregulation in Lilium pollen grains occurs via modulation of the plasma membrane H+-ATPase activity by 14-3-3 proteins. Plant Physiol. 2010, 154, 1921–1928. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas, L.; McKenna, S.T.; Kunkel, J.G.; Hepler, P.K. NAD(P)H oscillates in pollen tubes and is correlated with tip growth. Plant Physiol. 2006, 142, 1460–1468. [Google Scholar] [CrossRef] [PubMed]
- Paital, B.; Sablok, G.; Kumar, S.; Singh, S.K.; Chainy, G.B.N. Investigating the conformational structure and potential site interactions of SOD Inhibitors on Ec-SOD in marine mud crab scylla serrata: A molecular modeling approach. Interdiscip. Sci. Comput. Life Sci. 2016, 8, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Potocký, M.; Jones, M.A.; Bezvoda, R.; Smirnoff, N.; Zárský, V. Reactive oxygen species produced by NAD(P)H oxidase are involved in pollen tube growth. New Phytol. 2007, 174, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Speranza, A.; Crinelli, R.; Scoccianti, V.; Geitmann, A. Reactive oxygen species are involved in pollen tube initiation in kiwifruit. Plant Biol. Stuttg. Ger. 2012, 14, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Rounds, C.M.; Hepler, P.K.; Fuller, S.J.; Winship, L.J. Oscillatory growth in lily pollen tubes does not require aerobic energy metabolism. Plant Physiol. 2010, 152, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Selinski, J.; Scheibe, R. Pollen tube growth: Where does the energy come from? Plant Signal. Behav. 2014, 9, e977200. [Google Scholar] [CrossRef] [PubMed]
- Stael, S.; Wurzinger, B.; Mair, A.; Mehlmer, N.; Vothknecht, U.C.; Teige, M. Plant organellar calcium signalling: An emerging field. J. Exp. Bot. 2012, 63, 1525–1542. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.; Behera, S.; Bortoli, S.D.; Logan, D.C.; Fuchs, P.; Carraretto, L.; Teardo, E.; Cendron, L.; Nietzel, T.; Füßl, M.; et al. The EF-Hand Ca2+ binding protein MICU choreographs mitochondrial Ca2+ dynamics in Arabidopsis. Plant Cell 2015, 27, 3190–3212. [Google Scholar] [CrossRef] [PubMed]
- Rounds, C.M.; Hepler, P.K.; Winship, L.J. The apical actin fringe contributes to localized cell wall deposition and polarized growth in the lily pollen tube. Plant Physiol. 2014, 166, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Dutta, R.; Robinson, K.R. Identification and characterization of stretch-activated ion channels in pollen protoplasts. Plant Physiol. 2004, 135, 1398–1406. [Google Scholar] [CrossRef] [PubMed]
- Bucher, M.; Brander, K.A.; Sbicego, S.; Mandel, T.; Kuhlemeier, C. Aerobic fermentation in tobacco pollen. Plant Mol. Biol. 1995, 28, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Colaço, R.; Moreno, N.; Feijó, J.A. On the fast lane: Mitochondria structure, dynamics and function in growing pollen tubes. J. Microsc. 2012, 247, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Sze, H.; Li, X.; Palmgren, M.G. Energization of plant cell membranes by H+-pumping ATPases. Regulation and biosynthesis. Plant Cell 1999, 11, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Palmgren, M.G. Plant plasma membrane H+-ATPases: Powerhouses for Nutrient Uptake. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 817–845. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E. Plant Physiology, 4th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2006. [Google Scholar]
- Chen, C.Y.; Wong, E.I.; Vidali, L.; Estavillo, A.; Hepler, P.K.; Wu, H.; Cheung, A.Y. The regulation of actin organization by actin-depolymerizing factor in elongating pollen tubes. Plant Cell 2002, 14, 2175–2190. [Google Scholar] [CrossRef] [PubMed]
- Lovy-Wheeler, A.; Wilsen, K.L.; Baskin, T.I.; Hepler, P.K. Enhanced fixation reveals the apical cortical fringe of actin filaments as a consistent feature of the pollen tube. Planta 2005, 221, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Lenartowska, M.; Michalska, A. Actin filament organization and polarity in pollen tubes revealed by myosin II subfragment 1 decoration. Planta 2008, 228, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Carlier, M.F.; Laurent, V.; Santolini, J.; Melki, R.; Didry, D.; Xia, G.X.; Hong, Y.; Chua, N.H.; Pantaloni, D. Actin depolymerizing factor (ADF/cofilin) enhances the rate of filament turnover: Implication in actin-based motility. J. Cell Biol. 1997, 136, 1307–1322. [Google Scholar] [CrossRef] [PubMed]
- Gungabissoon, R.A.; Jiang, C.-J.; Drøbak, B.K.; Maciver, S.K.; Hussey, P.J. Interaction of maize actin-depolymerising factor with actin and phosphoinositides and its inhibition of plant phospholipase C. Plant J. 1998, 16, 689–696. [Google Scholar] [CrossRef]
- Allwood, E.G.; Anthony, R.G.; Smertenko, A.P.; Reichelt, S.; Drobak, B.K.; Doonan, J.H.; Weeds, A.G.; Hussey, P.J. Regulation of the pollen-specific actin-depolymerizing factor LIADF1. Plant Cell 2002, 14, 2915–2927. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Pei, W.; Haiyun, R. Actin fringe is correlated with tip growth velocity of pollen tubes. Mol. Plant 2012, 5, 1160–1162. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Qu, X.; Zhang, R. Plant villins: Versatile actin regulatory proteins. J. Integr. Plant Biol. 2015, 57, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Kovar, D.R.; Drøbak, B.K.; Staiger, C.J. Maize profilin isoforms are functionally distinct. Plant Cell 2000, 12, 583–598. [Google Scholar] [CrossRef] [PubMed]
- Vidali, L.; Rounds, C.M.; Hepler, P.K.; Bezanilla, M. Lifeact-mEGFP reveals a dynamic apical F-actin network in tip growing plant cells. PLoS ONE 2009, 4, e5744. [Google Scholar] [CrossRef] [PubMed]
- Vos, J.W.; Valster, A.H.; Hepler, P.K. Methods for studying cell division in higher plants. Methods Cell Biol. 1999, 61, 413–437. [Google Scholar] [PubMed]







© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winship, L.J.; Rounds, C.; Hepler, P.K. Perturbation Analysis of Calcium, Alkalinity and Secretion during Growth of Lily Pollen Tubes. Plants 2017, 6, 3. https://doi.org/10.3390/plants6010003
Winship LJ, Rounds C, Hepler PK. Perturbation Analysis of Calcium, Alkalinity and Secretion during Growth of Lily Pollen Tubes. Plants. 2017; 6(1):3. https://doi.org/10.3390/plants6010003
Chicago/Turabian StyleWinship, Lawrence J., Caleb Rounds, and Peter K. Hepler. 2017. "Perturbation Analysis of Calcium, Alkalinity and Secretion during Growth of Lily Pollen Tubes" Plants 6, no. 1: 3. https://doi.org/10.3390/plants6010003