Thai Fermented Foods as a Versatile Source of Bioactive Microorganisms—A Comprehensive Review


Innovation Center for Holistic Health, Nutraceuticals, and Cosmeceuticals, Faculty of Pharmacy, Chiang Mai University, Chiang Mai 50200, Thailand
*
Authors to whom correspondence should be addressed.
Sci. Pharm. 2018, 86(3), 37; https://doi.org/10.3390/scipharm86030037
Submission received: 25 July 2018
/
Revised: 8 September 2018
/
Accepted: 9 September 2018
/
Published: 11 September 2018
Abstract
:Fermented foods are known for several health benefits, and they are generally used among the Asian people. Microorganisms involved in the fermentation process are most responsible for the final quality of the food. Traditional fermented (spontaneous fermentation) foods are a versatile source of bioactive molecules and bioactive microbes. Several reports are available regarding the isolation and characterization of potent strains from traditional fermented foods. A collection of information for easy literature analysis of bioactive microbes derived from Thai fermented food is not yet available. The current manuscript compiled information on bioactive (antimicrobial- and enzyme-producing probiotic) microbes isolated from naturally fermented Thai foods.
1. Introduction
Fermented foods have been commonly used among the Asian people, especially Southeast Asian people, for centuries. Natural or spontaneous fermentation was used by our ancestors for the preparation and preservation of foods (e.g., wine) without having any knowledge of microbes involved in the process. Microorganisms involved in the fermentation process are most responsible for the final food quality regarding taste, texture, nutrition value, etc. The use of a desirable starting culture for the making of fermented food may prevent contamination and helps retain consistent food quality.
Fermented vegetable (Inziangsang, Kimchi, Pak-Gard-Dong, Khalpi, Dakguadong, Jiang-gua, Burong mustala, Gundruk, Ca muoi, Soidon, Suan-tsai, Sayur asin, Paocai, Sauerkraut, Nozawana-Zuke, Yan-jiang, and Yan-tsai-shin), fruit (Yan-taozih, Olive, Sunki, Tempoyak), fish (Bagoong, Funazushi, Hongeohoe, Pla ra, and Sikhae), and meat (Chartayshya, Arjia, Jamma or Geema, satchu, kargyong, Kheuri, Suka Ko Masu, Jangpangnatsu, and Jang Kap) products are habitually consumed by Thai, Indian, Chinese, Vietnamese, Japanese, Korean, and Taiwanese people [1,2,3,4,5,6,7,8,9,10,11,12,13].
Many fermented foods are prepared by following traditional methods to retain the reputation of the food. Conventional or secret seed cultures and processing conditions for food preparation also vary, and they play a critical role in the eminence of food. The lactic-acid-bacteria-mediated fermentation of vegetable, meats, and fruits is a standard method for preserving and advancing the dietary and sensory characteristics of food commodities [14,15].
Traditional fermented foods are prepared by spontaneous fermentation. In general, fermented foods of Asian countries are fermented using lactic-acid bacteria (LAB). Commonly used probiotic LAB strains are Lactobacillus casei, L. plantarum, L. brevis, L. pentosus, L. fermentum, L. paracasei, Weissella koreenis, W. confusa, W. cibaria, Leuconostoc kimchi, L. mesenteroides, L. fallax, Enterococcus and Pediococcus spp. The complex nutritional substance of food raw materials like meat and vegetables are a rich source of necessary vitamins and minerals for the growth of LAB strains, which facilitates the microbial production of enzymes and other metabolites [13]. Only little is known about the microorganisms involved in the fermentation process of various traditional fermented foods due to the lack of studies on them. Several studies revealed the effect of supplementation of foods containing active microbes, which are known as probiotics that deliver health benefits such as anti-diarrhea and anti-colitis effects, as well as activity against hypercholesterolemia, urinary tract infections, allergies, etc. [16,17,18,19,20].
Several reports are available on the isolation and characterization of potent LAB strains from traditional fermented foods [21,22,23]. Samples of fermented foods were serially diluted and spread on de Man, Rogosa, and Sharpe (MRS) agar plates and incubated at 37 °C for 48 h. After incubation, colonies were randomly selected for further study based on the different morphology of the colonies. The colonies were streaked to get pure colonies, and further phenotypic identification was carried out by observing the colony morphology, and through microscopical examination by Gram staining and endospore staining. Biochemical characterization was carried out, and isolates were then subjected to screening tests such as acid tolerance tests, growth rates at different NaCl concentrations and pH, antagonistic activity tests, artificial gastrointestinal juice tolerance tests, mucus adhesion tests, and screening for the production of enzymes and their activities. Finally, the isolates were identified using molecular techniques including DNA extraction, DNA amplification using PCR, and DNA sequencing [21,22,23]. The current manuscript compiles information on bioactive (antimicrobial- and enzyme-producing probiotic) microbes isolated from naturally fermented Thai foods.
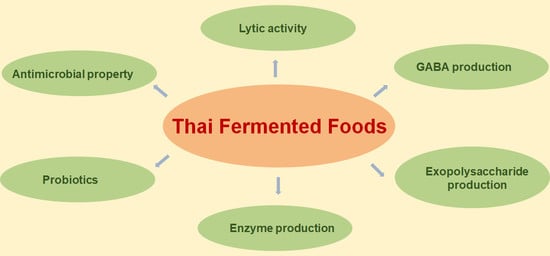
2. Bioactive Microbes from Thai Fermented Foods
2.1. Strains with Antimicrobial Properties
The antimicrobial activities of LAB isolated from Thai fermented foods are listed in Table 1. A total of 43 lactic acid bacteria (LAB) were isolated from fermented fish, and the isolates were found to exhibit anti-listerial activity (inhibiting the growth of Listeria monocytogenes). Furthermore, the isolates were challenged against Vibrio spp. and Aeromonas spp. and about 37 strains were active against the tested pathogens. Most of the isolates are Lactobacillus species. The antimicrobial properties of the isolates were attributed to the production of bacteriocin-like compounds and lactic acid [24]. L. plantarum N014, isolated from fermented pork, was also reported for anti-listerial activity. The bacteriocin, an 8-kDa protein, which is sensitive to proteolytic enzymes and resistant to lipase A, α-amylase, and lysozyme, is produced by the strain N014, which inhibited the growth of representative Gram-positive (Lactobacillus spp. and Streptococcus spp.), and Gram-negative (Escherichia coli, Shigella dysenteriae, and Pseudomonas aeruginosa) pathogens. Antimicrobial activity was retained at various levels of temperature and pH [25].
Bacteriocin-producing W. cibaria 110 was isolated from Plaa-Som (Thai fermented fish product), which was active against L. sakei, L. sanfranciscensis, L. acetotolerans, L. homohiochii, L. coryniformis subsp. coryniformis, W. kandleri, W. halotolerans, W. paramesenteroides, and Leuconostoc lactis. The purified bacteriocin was about 2.5 kDa in size with a peptide mass of 3487.8 Da. The N-terminal amino-acid sequence analysis suggested that the bacteriocin of W. cibaria 110 was novel, and was named as weissellicin 110 at the time of the report [26]. Another Weissella strain, W. hellenica BCC 7239 isolated from Nham, was reported for the production of bacteriocins (7293A and 7293B) with antimicrobial activity against pathogens like E. coli, Aeromonas hydrophila, P. aeruginosa, and Salmonella Typhimurium. Bacteriocin 7293A exhibited higher antimicrobial activity than 7293B. Both bacteriocins were inactive after proteolytic enzyme treatment, while their functionality was not affected by organic solvents and other enzymes. The molecular weights of the novel bacteriocins of W. hellenica BCC 7239 (7293A and 7293B) were found as 6249.302 and 6489.716 Da, respectively [27].
Prachyakij et al. [28] screened LAB isolates for anti-yeast activity (food contaminants) and found that several L. plantarum strains showed better antagonistic activity against Rhodotorula species. A new variant of nukacin-131 (a bacteriostatic antibiotic) producing Staphylococcus hominis KQU-131 strain was isolated from the fermented fish product of Thailand. Based on the genetic and structural analysis, the bacteriocin was termed as nukacin KQU-131 (only three amino-acid residues differentiate KQU-131 from ISK-1) [29].
About 23 Bacillus strains were isolated from Thua-nao (a Thai-style fermented soybean) and screened against an aflatoxin-producing food isolate of Aspergillus flavus and a reference strain A. westerdijkiae NRRL3174. Among the isolates, one strain showed promising inhibition of Aspergillus growth and reduced 74 and 92.5% of myotoxicity of aflatoxin B1 and ochratoxin A, respectively. Another isolate was identified as B. subtilis, which also showed about 85% aflatoxin B1 detoxification. The authors claimed that the food isolate, B. subtilis, could be used as a biocontrol agent to detoxify aflatoxin and ochratoxin [30].
LAB strains isolated from several fermented foods were screened for anti-Vibrio activity, especially against the pandemic strains of V. parahaemolyticus O3:K6. L. plantarum, isolated from fermented pork, showed strong inhibition of the growth of V. parahaemolyticus and V. cholerae, and the inhibitory property was attributed to acid production. L. plantarum (PSU-LAB71) showed high affinity to the Caco-2 cell line, and perhaps this strain can be used as probiotic to control pandemic infections [31].
About 529 LAB were isolated from Thai fermented foods and screened for antimicrobial activity. Among the isolates, KKU 170, later identified as Pediococcus acidilactici, was selected for further study. The antagonistic compound produced by KKU 170 was bacteriocin and was found to be active against tested microbial pathogens including Listeria species. The optimal conditions for producing bacteriocin by KKU 170 were found as a pH of 6.5, a temperature of 45 °C, and 0.2% glucose in the medium. Even though the optimal temperature was 45 °C, the enzyme could withstand up to 121 °C for 30 min [32].
Heat- and pH-stable bacteriocin-producing Lactococcus lactis KTH0-1S was isolated from Kung-som (Thai traditional fermented shrimp product), and it was observed that the bacteriocin KTH0-1S was active against food-related pathogens and effectively reduced the viability and growth of Staphylococcus aureus. The chromatography and spectrometry analyses revealed that bacteriocin KTH0-1S was nisin Z with a molecular mass of 3346 Da. L. lactis KTH0-1S was non-virulent and able to inhibit contaminating microbes. Thus, L. lactis KTH0-1S could be used as an antagonistic strain in the production of fermented products [33]. Another strain, L. lactis WNC20 was isolated from Thai fermented sausage (Nham). The heat- and pH-stable bacteriocin produced by the WNC20 strain was active against S. aureus, B. cereus, L. monocytogenes, and Clostridium perfringens. The detailed molecular studies suggested that the bacteriocin WNC20 was identical to nisin Z [34].
Probiotic strain, Pediococcus pentosaceus NP6, was isolated from Nam-pla (fermented fish paste), which was tolerable to acids and bile salt, and was able to utilize carbohydrates and produce amylase and lipase. P. pentosaceus NP6 was antagonistic against foodborne pathogens. The co-cultivation study revealed that P. pentosaceus NP6 could potentially inhibit the growth of Salmonella Typhimurium in Nile tilapia by-products. The authors claimed that NP6 could be used as a biocontrol agent to protect from microbial contamination and extend the shelf life of the raw meats [35].
2.2. Lytic Activity
The lytic activities of LAB isolated from Thai fermented foods are listed in Table 2. The proteolytic bacterial strains were isolated from Thai fermented foods (Thua Nao, Sa La Pao, and Kha Nhom Jeen), and potent strains were identified as Bacillus subtilis strains DB and SR, and B. licheniformis. The antiallergic nature of crude enzymes of B. subtilis strains DB and SR was determined, and it was found that the enzyme diminished the allergenicity of gliadin, which was attributed to the hydrolyzing ability of the enzyme. B. subtilis DB enzyme also reduced β-lactoglobulin allergenicity by hydrolyzing the allergenic epitope of β-lactoglobulin. The enzyme of B. subtilis DB and SR strain could be used to produce low-allergic wheat and milk products [36].
About six Bacillus strains (B. pumilus, B. methylotropicus, B. subtilis, B. flexus, B. anthracis, and B. cereus), and a Paenibacillus strain (P. pasadenensis) were isolated from fermented pork (Nham), fish (Pla-ra), and shrimp (Kung-chom) products of Thailand. The strains were found to be susceptible to tested common antibiotics. All strains produce lipase in the presence of Tween 20, 40, 60, and 80 in nutrient broth. B. subtilis SS48-5 produced relatively high lipase activity (3.231 ± 0.087 U/mL) in the presence of Tween 80 at pH 7.5. These isolates could be used for the lipase production [37].
2.3. Probiotic Strains
Three L. fermentum strains were found by screening 563 strains isolated from Thai fermented products such as fermented fish, fermented pork, and fermented tea leaves. The strains were able to grow in a wide range of pH (2–7) and bile-salt concentrations (0.3–1.0%), and had potent adhesive properties. The fermented pork isolates (no. 3007 and 3010) showed relatively high adhesive potential compared to other strains. All strains exhibited antimicrobial activity against E. coli TISTR 780, S. aureus TISTR029, and S. Typhi DMST5784. The strains were resistant to vancomycin and ciprofloxacin, and were sensitive to erythromycin, kanamycin, chloramphenicol, tetracycline quinupristin, linezolid, streptomycin, and rifampicin [38].
B. siamensis B44v isolated from Phak-dong (Thai pickled vegetables) was carefully evaluated for probiotic activity. The strain B44v exhibited antimicrobial activity against a wide range of Gram-positive and Gram-negative pathogens, which was attributed to the production of bacteriocin-like compounds. B. siamensis B44v could produce cellulase and protease enzymes, and survived in gastrointestinal conditions. The strain was susceptible to several common antibiotics and showed high hydrophobicity (a measure to determine the adherence property of the strain), co-aggregation, auto-agglutination, and mucin-binding capacity. The strain B44v was found to protect the hybrid catfish from fish pathogens A. hydrophila FW52 effectively, and also found to promote fish growth compared to the control. The results of the study suggested that B. siamensis B44v could be used as a probiotic for catfish farming and for other aquaculture applications [39].
About 119 strains were isolated from various fermented foods, and the strains were screened for antibacterial activity against fish pathogens. The isolate CR1T5 exhibited the best antagonistic activity against the tested pathogens (Streptococcus agalactiae, A. hydrophila, and A. caviae), and it was identified as L. plantarum. The strain CR1T5 was resistant to simulated gastrointestinal conditions, and had high adherence capacity. The supplementation of L. plantarum CR1T5 promoted the growth of tilapia fish and protected them from A. hydrophila infection (87.5%). The results suggested that L. plantarum CR1T5 could be a potent probiotic for farmed fish [40].
2.4. Enzyme Production
γ-Glutamyl-hydrolase-producing Bacillus spp., designated as Bacillus CMU29, were isolated from Thai fermented soybean food (Thua-nao). The enzyme γ-glutamyl hydrolase was separated from the culture supernatant of CMU29. The molecular mass of the enzyme was 28 kDa, and the enzyme could hydrolyze oligo-γ-l-glutamates and γ-polyglutamic acid. The enzyme acts on the γ-glutamyl linkage of γ-polyglutamic acid. The results proved that the purified enzyme was γ-glutamyl hydrolase [41].
Raw and fermented pork (Nham and Saigongyisan) were screened for bioactive LAB strains that were active against E. coli DMST 4212, L. monocytogenes DMST 11256, Pseudomonas fluorescens DMST 20076, S. Typhimurium DMST 0562, and S. aureus TISTR 118. The isolate P0805 was found as a potent bioactive strain with antagonistic activity against tested pathogens, and the isolate was identified as P. pentosaceus. The strain was sensitive to erythromycin, ampicillin, and chloramphenicol, and resistant to gentamicin, cefuroxime, and rifampicin. The antagonistic substance produced by P. pentosaceus P0805 was partially purified and the fraction 409,600 AU/mL showed high activity against S. Typhimurium. The isolate P0805 was nitrate reductase and catalase positive. Further evaluation proved that the catalase of P0805 was heme-independent catalase. The authors claimed that P. pentosaceus P0805 could be used as a starter for the production of fermented meat product [42].
The β-glucosidase-enzyme-producing LAB strain, L. plantarum SC359, was isolated from Thai pickled soybean. L. plantarum SC359 survived in bile salt (0.30%) and at various pH (pH 2–5), and exhibited antagonistic activity against tested Gram-positive and Gram-negative bacterial pathogens. L. plantarum SC359 was susceptible to all common antibiotics and displayed lipolytic and proteolytic activities. L. plantarum SC359 actively prevented the adherence of S. Typhi, E. coli, and S. sonnei to Caco-2 cells. The β-glucosidase-producing probiotic strain L. plantarum SC359 may be considered as the suitable starter for fermented foods [43].
About 338 bacterial strains were isolated from different Thai fermented foods and were screened for l-glutaminase and glutamic acid decarboxylase production. The isolates G507/1 and G254/2 exhibited high glutaminase and glutamic acid decarboxylase activity, respectively. The isolates G507/1 and G254/2 were identified as L. brevis and L. fermentum, respectively. The influence of pH, temperature, and incubation on enzyme activity were studied, and the results suggested that pH values of 6.0 and 6.5, temperatures of 35 and 30 °C, and 10 days of incubation were the optimal condition for l-glutaminase and glutamic acid decarboxylase activity, respectively [44].
B. licheniformis RKK-04 was isolated from Thai fermented fish sauce. The strain exhibited proteolytic activity in vitro. The partially purified enzyme was 31 kDa in mass and had an isoelectric point of >9.3. Further characterization revealed that the optimal values of temperature and pH for enzyme activity were 50 °C and 10.0, respectively. The enzyme was sensitive to phenylmethanesulfonyl fluoride and diisopropyl fluorophosphate, which proved that the RKK-04 enzyme may be a subtilisin-like alkaline serine proteinase. The high salt concentration (30% NaCl) did not significantly affect the enzyme activity at 30 °C, while reduced enzyme activity was observed at 50 °C. The RKK-04 enzyme differed from other subtilisin-like proteases regarding its cleavage sites in the insulin B-chain. The halotolerant B. licheniformis RKK-04 proteinase could digest the myosin heavy chain of fish protein completely. The results suggested that B. licheniformis RKK-04 proteinase could play a critical role in fish sauce fermentation [45].
2.5. Gamma-Aminobutyric Acid (GABA) Production
A GABA (Gamma-Aminobutyric Acid)-producing LAB strain was screened from bacterial isolates of various fermented Thai foods. About 602 isolates were screened and four potent GABA-producing isolates such as HN8, NH116, NH102, and NH2 were found producing GABA contents of about 9.060, 8.411, 8.386, and 7.339 g per L, respectively. The pH of culture supernatants was in the range of 3.8–4.2. All four isolates exhibited good antagonistic activity against tested foodborne pathogens. There were no significant changes in survival in up to 2 h of exposure to both intestinal juice and simulated gastric juice, but increased exposure time reduced the survival of the isolates. The antibiotic sensitivity pattern varied among the isolates, and ampicillin and tetracycline inhibited the growth of all four isolates. The strains survived in the presence of bile salt with no hemolysis. The isolates HN8, NH116, and NH102 were closely related to P. pentosaceus, and the isolate NH2 was found to be Lactobacillus namurensis. Based on the fermentation study, HN8 and NH2 were identified as potential starters for the preparation of fermented meat products [46].
The GABA-producing bacterial isolates CS3 and CS5 were isolated from Thai fermented shrimp (Kung-Som), and later identified as Lactobacillus futsaii using morphological and molecular methods. L. futsaii CS3 was the most potent GABA producer, and it could convert monosodium glutamate to GABA with 99% yield. The molecular studies proved that strain CS3 had glutamate decarboxylase, with 469 amino acids containing the protein-coding gene. The predicted molecular mass and isoelectric point of the protein were 53.64 kDa and 5.56, respectively. The author suggested that the strain L. futsaii CS3 could be used for the development of GABA-rich fermented foods [47].
2.6. Exopolysaccharide (EPS) Production
LAB strains were isolated from various Thai traditional fermented foods such as fermented fish, fermented shrimp, fermented pork sausage, and fermented vegetables. LAB isolates were screened for exopolysaccharide (EPS) production and it was found that some of the cocci were able to produce EPS in both solid and liquid media, later identified as P. pentosaceus, while some of the rod-shaped strains produced EPS only on solid media. About 6.0 and 2.5 g per L of EPS production were observed in sucrose-containing liquid media by P. pentosaceus AP-1 and AP-3, respectively. The secreted EPS belongs to the dextran class and has branches, and was proven based on the methylation analysis, sugar configuration, and nuclear magnetic resonance spectroscopic examinations. It was the first report on dextran-producing P. pentosaceus [48].
3. Conclusions and Future Prospects
Thai fermented foods are versatile sources of bioactive microorganisms. So far, several studies reported the antagonistic and probiotic properties, as well as the enzyme- and exopolysaccharide-producing abilities of the bacterial strains, which were isolated from traditional Thai fermented foods. Several studies reported antimicrobial- and bacteriocin-producing LAB in fermented Thai foods. Even though many potent bioactive microbes and metabolites were isolated and identified from Thai fermented foods, their applicability or sustainable utilization was questionable, except in some cases.
The probiotic LAB strain, E. faecalis (G414/1), isolated from Thai fermented foods was used for the development of l-glutamine- and l-glutamic-acid-rich fermented Hericium erinaceus juice [49]. In another study, L. fermentum HP3 was used for the preparation of H. erinaceus juice (FHJ), and the antidiabetic property of FHJ was reported [50].
Further studies are necessary to explore several untouched Thai traditional fermented foods and their functional properties. The application of identified and characterized bioactive microbes of Thai fermented foods and their metabolites in food industries may be ideal for the development of functional foods.
Author Contributions
B.S.S. contributed to conception and design, acquisition, manuscript preparation, and critical revision of the manuscript. P.K. and C.C. were involved in the review and finalization of the manuscript. All authors agreed with the content of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The author gratefully acknowledges the Chiang Mai University Grant (CMU-grant), Chiang Mai University, and the Faculty of Pharmacy, Chiang Mai University, Chiang Mai, Thailand. Bhagavathi Sundaram Sivamaruthi wishes to acknowledge the CMU Post-Doctoral Fellowship (Ref: No. 6592(11)/1379, dated 26 February 2018), Chiang Mai University, Chiang Mai, Thailand.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tamang, J.P. Himalayan Fermented Foods: Microbiology, Nutrition and Ethnic Values, 1st ed.; CRC Press: New Delhi, India, 2009; pp. 1–216. ISBN 9781420093247. [Google Scholar]
- Yan, P.M.; Xue, W.T.; Tan, S.S.; Zhang, H.; Chang, X.H. Effect of inoculating lactic acid bacteria starter cultures on the nitrite concentration of fermenting Chinese Paocai. Food Control 2008, 19, 50–55. [Google Scholar] [CrossRef]
- Puspito, H.; Fleet, G.H. Microbiology of sayur asin fermentation. Appl. Microbiol. Biotechnol. 1985, 22, 442–445. [Google Scholar] [CrossRef]
- Viander, B.; Maki, M.; Palva, A. Impact of low salt concentration, salt quality on natural large-scale sauerkraut fermentation. Food Microbiol. 2003, 20, 391–395. [Google Scholar] [CrossRef]
- Chen, Y.S.; Yanagida, F.; Hsu, J.S. Isolation and characterization of lactic acid bacteria from suan-tsai (fermented mustard), a traditional fermented food in Taiwan. J. Appl. Microbiol. 2006, 101, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Wu, H.C.; Lo, H.Y.; Lin, W.C.; Hsu, W.H.; Lin, C.W.; Lin, P.Y.; Yanagida, F. Isolation and characterization of lactic acid bacteria from jiang-gua (fermented cucumbers), a traditional fermented food in Taiwan. J. Sci. Food Agric. 2012, 92, 2069–2075. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Wu, H.C.; Pan, S.F.; Lin, B.; Lin, Y.H.; Tung, W.C.; Li, Y.; Chiang, C.; Yanagida, F. Isolation and characterization of lactic acid bacteria from yan-taozih (pickled peaches) in Taiwan. Ann. Microbiol. 2013, 63, 607–614. [Google Scholar] [CrossRef]
- Chen, Y.S.; Wu, H.C.; Wang, C.M.; Lin, C.C.; Chen, Y.T.; Jhong, Y.J.; Yanagida, F. Isolation and characterization of lactic acid bacteria from pobuzihi (fermented cummingcordia), a traditional fermented food in Taiwan. Folia Microbiol. 2013, 58, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; Chen, Y.S.; Yanagida, F. Isolation and characterization of lactic acid bacteria from yan-jiang (fermented ginger), a traditional fermented food in Taiwan. J. Sci. Food Agric. 2011, 91, 1746–1750. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.T.; van Hoorde, K.; Cnockaert, M.; de Brandt, E.; Aerts, M.; Binh Thanh, L.; Vandamme, P. A description of the lactic acid bacteria microbiota associated with the production of traditional fermented vegetables in Vietnam. Int. J. Food Microbiol. 2013, 163, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Karovicova, J.; Kohajdova, Z.; Greif, G. The use of PCA, CA, FA for evaluation of vegetable juices processed by lactic acid fermentation. Czech J. Food Sci. 2002, 20, 135–143. [Google Scholar] [CrossRef]
- Tanasupawat, S.; Komagata, K. Lactic acid bacteria in fermented foods in Thailand. World J. Microbiol. Biotechnol. 1995, 11, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Swain, M.R.; Anandharaj, M.; Ray, R.C.; Rani, R.P. Fermented fruits and vegetables of Asia: A potential source of probiotics. Biotechnol. Res. Int. 2014, 2014, 19. [Google Scholar] [CrossRef] [PubMed]
- Demir, N.; Bahceci, K.S.; Acar, J. The effects of different initial Lactobacillus plantarum concentrations on some properties of fermented carrot juice. J. Food Process. Preserv. 2006, 30, 352–363. [Google Scholar] [CrossRef]
- Di Cagno, R.; Coda, R.; de Angelis, M.; Gobbetti, M. Exploitation of vegetables and fruits through lactic acid fermentation. Food Microbiol. 2013, 33, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Doron, S.I.; Hibberd, P.L.; Gorbach, S.L. Probiotics for prevention of antibiotic-associated diarrhea. J. Clin. Gastroenterol. 2008, 42, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Grin, P.M.; Kowalewska, P.M.; Alhazzan, W.; Fox-Robichaud, A.E. Lactobacillus for preventing recurrent urinary tract infections in women: Meta-analysis. Can. J. Urol. 2013, 20, 6607–6614. [Google Scholar] [PubMed]
- Dylag, K.; Hubalewska-Mazgaj, M.; Surmiak, M.; Szmyd, J.; Brzozowski, T. Probiotics in the mechanism of protection against gut inflammation and therapy of gastrointestinal disorders. Curr. Pharm. Des. 2014, 20, 1149–1155. [Google Scholar] [CrossRef] [PubMed]
- Tomaro-Duchesneau, C.; Saha, S.; Malhotra, M.; Jones, M.L.; Labbe, A.; Rodes, L.; Kahouli, I.; Prakash, S. Effect of orally administered L. fermentum NCIMB 5221 on markers of metabolic syndrome: An in vivo analysis using ZDF rats. Appl. Microbiol. Biotechnol. 2014, 98, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Sivamaruthi, B.S. A comprehensive review on clinical outcome of probiotic and synbiotic therapy for inflammatory bowel diseases. Asian Pac. J. Trop. Biomed. 2018, 8, 179–186. [Google Scholar] [CrossRef]
- Bansal, S.; Singh, A.; Mangal, M.; Sharma, S.K. Isolation and characterization of lactic acid bacteria from fermented Foods. Vegetos 2013, 26, 325–330. [Google Scholar] [CrossRef]
- Ho, J.C.K.; Sze, LY. Isolation, identification and characterization of enzyme-producing lactic acid bacteria from traditional fermented foods. Biosci. Horiz. 2018, 11, hzy004. [Google Scholar] [CrossRef]
- Linh, N.T.H.; Sakai, K.; Taoka, Y. Screening of lactic acid bacteria isolated from fermented food as potential probiotics for aquacultured carp and amberjack. Fish. Sci. 2018, 84, 101–111. [Google Scholar] [CrossRef]
- Ostergaard, A.; Embarek, P.K.B.; Wedell-Neergaard, C.; Huss, H.H.; Gram, L. Characterization of anti-listerial lactic acid bacteria isolated from Thai fermented fish products. Food Microbiol. 1998, 15, 223–233. [Google Scholar] [CrossRef]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Isolation and preliminary characterization of a bacteriocin produced by Lactobacillus plantarum N014 isolated from Nham, a traditional Thai fermented pork. J. Food Prot. 2006, 69, 1937–1943. [Google Scholar] [CrossRef] [PubMed]
- Srionnual, S.; Yanagida, F.; Lin, L.; Hsiao, K.; Chen, Y. Weissellicin 110, a newly discovered bacteriocin from Weissella cibaria 110, isolated from Plaa-Som, a fermented fish product from Thailand. Appl. Environ. Microbiol. 2007, 73, 2247–2250. [Google Scholar] [CrossRef] [PubMed]
- Woraprayote, W.; Pumpuang, L.; Tosukhowong, A.; Roytrakul, S.; Perez, R.H.; Zendo, T.; Sonomoto, K.; Benjakul, S.; Visessanguan, W. Two putatively novel bacteriocins active against Gram-negative food borne pathogens produced by Weissella hellenica BCC 7293. Food Control 2015, 55, 176–184. [Google Scholar] [CrossRef]
- Prachyakij, P.; Schnurer, J.; Charernjiratrakul, W.; Kantachote, D. Selection and identification of lactic acid bacteria that inhibit yeast contaminants isolated from fermented plant beverages. Songklanakarin J. Sci. Technol. 2007, 29, 211–218. [Google Scholar]
- Wilaipun, P.; Zendo, T.; Okuda, K.; Nakayama, J.; Sonomoto, K. Identification of the Nukacin KQU-131, a new type-A(II) lantibiotic produced by Staphylococcus hominis KQU-131 isolated from Thai fermented fish product (Pla-ra). Biosci. Biotechnol. Biochem. 2008, 72, 2232–2235. [Google Scholar] [CrossRef] [PubMed]
- Petchkongkaew, A.; Taillandier, P.; Gasaluck, P.; Lebrihi, A. Isolation of Bacillus spp. from Thai fermented soybean (Thua-nao): Screening for aflatoxin B1 and ochratoxin A detoxification. J. Appl. Microbiol. 2008, 104, 1495–1502. [Google Scholar] [CrossRef] [PubMed]
- Charernjiratragul, W.; Bhoopong, P.; Kantachote, D.; Jomduang, S.; Kong-Ngoen, R.; Nair, G.B.; Vuddhakul, V. Inhibitory activity of lactic acid bacteria isolated from Thai fermented food against pandemic strains of Vibrio parahaemolyticus. J. Food Saf. 2010, 30, 67–82. [Google Scholar] [CrossRef]
- Leelavatcharamas, V.; Arbsuwan, N.; Apiraksakorn, J.; Laopaiboon, P.; Kishida, M. Thermotolerant bacteriocin-producing lactic acid bacteria isolated from Thai local fermented foods and their bacteriocin productivity. Biocontrol Sci. 2011, 16, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Saelao, S.; Maneerat, S.; Kaewsuwan, S.; Rabesona, H.; Choiset, Y.; Haertle, T.; Chobert, J.M. Inhibition of Staphylococcus aureus in vitro by bacteriocinogenic Lactococcus lactis KTH0-1S isolated from Thai fermented shrimp (Kung-som) and safety evaluation. Arch. Microbiol. 2017, 199, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Noonpakdee, W.; Santivarangkna, C.; Jumriangrit, P.; Sonomoto, K.; Panyim, S. Isolation of nisin-producing Lactococcus lactis WNC 20 strain from nham, a traditional Thai fermented sausage. Int. J. Food Microbiol. 2003, 81, 137–145. [Google Scholar] [CrossRef]
- Tasaku, S.; Siripornadulsil, S.; Siripornadulsil, W. Inhibitory activity of food-originated Pediococcus pentosaceus NP6 against Salmonella enterica serovar Typhimurium in Nile Tilapia by-products. Chiang Mai J. Sci. 2017, 44, 383–393. [Google Scholar]
- Phromraksa, P.; Nagano, H.; Boonmars, T.; Kamboonruang, C. Identification of proteolytic bacteria from Thai traditional fermented foods and their allergenic reducing potentials. J. Food Sci. 2008, 73, M189–M195. [Google Scholar] [CrossRef] [PubMed]
- Phoottosavako, M.; Keeratipibul, S.; Techo, S.; Tanasupawat, S. Identification and characterization of lipolytic bacteria from Thai fermented foods. Malays. J. Microbiol. 2015, 11, 231–239. [Google Scholar] [CrossRef]
- Klayraung, S.; Viernstein, H.; Sirithunyalug, J.; Okonogi, S. Probiotic properties of Lactobacilli isolated from Thai traditional food. Sci. Pharm. 2008, 76, 485–503. [Google Scholar] [CrossRef]
- Meidong, R.; Doolgindachbaporn, S.; Jamjan, W.; Sakai, K.; Tashiro, Y.; Okugawa, Y.; Tongpim, S. A novel probiotic Bacillus siamensis B44v isolated from Thai pickled vegetables (Phak-dong) for potential use as a feed supplement in aquaculture. J. Gen. Appl. Microbiol. 2017, 63, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Meidong, R.; Doolgindachbaporn, S.; Sakai, K.; Tongpim, S. Isolation and selection of lactic acid bacteria from Thai indigenous fermented foods for use as probiotics in tilapia fish Oreochromis niloticus. AACL Bioflux 2017, 10, 455–463. [Google Scholar]
- Chunhachart, O.; Itoh, T.; Sukchotiratana, M.; Tanimoto, H.; Tahara, Y. Characterization of γ-glutamyl hydrolase produced by Bacillus sp. isolated from Thai thua-nao. Biosci. Biotechnol. Biochem. 2006, 70, 2779–2782. [Google Scholar] [CrossRef] [PubMed]
- Nanasombat, S.; Treebavonkusol, P.; Kittisrisopit, S.; Jaichalad, T.; Phunpruch, S.; Kootmas, A.; Nualsri, I. Lactic acid bacteria isolated from raw and fermented pork products: Identification and characterization of catalase producing Pediococcus pentosaceus. Food Sci. Biotechnol. 2017, 26, 173–179. [Google Scholar] [CrossRef]
- Sirilun, S.; Chaiyasut, C.; Kantachote, D.; Luxananil, P. Functional properties of β-glucosidase-producing Lactobacillus plantarum SC 359 isolated from Thai fermented soybean food. Acta Aliment. 2012, 41, 451–464. [Google Scholar] [CrossRef]
- Woraharn, S.; Lailerd, N.; Sivamaruthi, B.S.; Wangcharoen, W.; Sirisattha, S.; Chaiyasut, C. Screening and kinetics of glutaminase and glutamate decarboxylase producing lactic acid bacteria from fermented Thai foods. Food Sci. Technol. 2014, 34, 793–799. [Google Scholar] [CrossRef] [Green Version]
- Toyokawa, Y.; Takahara, H.; Reungsang, A.; Fukuta, M.; Hachimine, Y.; Tachibana, S.; Yasuda, M. Purification and characterization of a halotolerant serine proteinase from thermotolerant Bacillus licheniformis RKK-04 isolated from Thai fish sauce. Appl. Microbiol. Biotechnol. 2010, 86, 1867–1875. [Google Scholar] [CrossRef] [PubMed]
- Ratanaburee, A.; Kantachote, D.; Charernjiratrakul, W.; Sukhoom, A. Selection of c-aminobutyric acid-producing lactic acid bacteria and their potential as probiotics for use as starter cultures in Thai fermented sausages (Nham). Int. J. Food Sci. Technol. 2013, 48, 1371–1382. [Google Scholar] [CrossRef]
- Sanchart, C.; Rattanaporn, O.; Haltrich, D.; Phukpattaranont, P.; Maneerat, S. Lactobacillus futsaii CS3, a new GABA-producing strain isolated from Thai fermented shrimp (Kung-Som). Indian J. Microbiol. 2017, 57, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Smitinont, T.; Tansakul, C.; Tanasupawat, S.; Keeratipibul, S.; Navarini, L.; Bosco, M.; Cescutti, P. Exopolysaccharide-producing lactic acid bacteria strains from traditional Thai fermented foods: Isolation, identification and exopolysaccharide characterization. Int. J. Food Microbiol. 1999, 51, 105–111. [Google Scholar] [CrossRef]
- Woraharn, S.; Lailerd, N.; Sivamaruthi, B.S.; Wangcharoen, W.; Peerajan, S.; Sirisattha, S.; Chaiyasut, C. Development of fermented Hericium erinaceus juice with high content of l-glutamine and l-glutamic acid. Int. J. Food Sci. Technol. 2015, 50, 2104–2112. [Google Scholar] [CrossRef]
- Chaiyasut, C.; Woraharn, S.; Sivamaruthi, B.S.; Kesika, P.; Lailerd, N.; Peerajan, S. Lactobacillus fermentum HP3 mediated fermented Hericium erinaceus juice as a health promoting food supplement to manage diabetes mellitus. J. Evid. Based Integr. Med. 2018, 23, 1–9. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
Table 1.
Antimicrobial activities of lactic-acid bacteria isolated from Thai fermented foods.
| S. No. | Source | Name of the Isolate | Bioactivity | Other Observations | Ref. |
|---|---|---|---|---|---|
| 1 | Fermented fish | Lactobacillus spp. | Anti-listerial activity | Displayed antimicrobial activity against Vibrio cholerae, V. parahaemolyticus, Aeromonas sp. | [24] |
| 2 | Fermented pork (Nham) | L. plantarum N014 | Anti-listerial activity, Bacteriocin production | Displayed antimicrobial activity against several Gram-positive and Gram-negative microbes. | [25] |
| 3 | Fermented fish (Plaa-Som) | Weissella cibaria 110 | Bacteriocin production | The bacteriocin, weissellicin 110, was active against some Gram-positive bacteria. Peptide mass was 3487.8 Da. | [26] |
| 4 | Fermented pork meat (Nham) | Weissella hellenica BCC 7293 | Bacteriocin production (7293A and 7293B) | Active against Gram-negative foodborne pathogens. | [27] |
| 5 | Fermented foods | L. plantarum | Anti-yeast activity | The isolated strains were tested against Rhodotorula spp., which were isolated from contaminated fermented plant beverages | [28] |
| 6 | Fermented fish (Pla-ra) | Staphylococcus hominis KQU-131 | Bacteriocin production | Nukacin-KQU-131, a new variant of nukacin ISK-1, was identified. | [29] |
| 7 | Fermented soybean (Thua-nao) | Bacillus spp. | Anti-Aspergillus activity | The isolate inhibited the growth of Aspergillus flavus and A. westerdijkiae, and detoxified aflatoxin B1 and ochratoxin A. | [30] |
| 8 | Fermented pork | L. plantarum (PSU-LAB71) | Inhibits V. parahaemolyticus, and V. cholerae | The isolate PSU-LAB71 was active against pandemic strains of Vibrio and found as a probiotic. | [31] |
| 9 | Fermented foods | Pediococcus acidilactici | Active against Gram-positive bacteria | P. acidilactici produced heat-tolerant bacteriocin. | [32] |
| 10 | Fermented shrimp(Kung-som) | Lactococcus lactis KTH0-1S | Anti-Staphylococcus aureus activity | Bacteriocin production (Nisin Z). The isolate KTH0-1S was safe and could be used to protect fermented seafood. | [33] |
| 11 | Fermented pork meat (Nham) | L. lactis WNC 20 | Active against foodborne pathogens | Heat-stable bacteriocin production (Nisin Z like). | [34] |
| 12 | Fermented fish (Nam-Pla) | P. pentosaceus NP6 | Anti-Salmonella enterica serovar Typhimurium activity | Active against both Gram-positive and Gram-negative pathogens. The isolate NP6 exhibited some probiotic properties. | [35] |
Table 2.
Functional properties of lactic-acid bacteria isolated from Thai fermented foods.
| Source | Name of the Isolate | Bioactivity | Other Observations | Ref. |
|---|---|---|---|---|
| Lytic Activity | ||||
| Fermented soybean (Thua Nao), fermented rice-noodle (Kha Nhom Jeen) | Bacillus subtilis, B. licheniformis | Proteolytic activity | The crude extract of culture supernatant showed in vitro anti-allergic activity. | [36] |
| Fermented meat and seafood | Bacillus spp. Paenibacillus spp. | Lipolytic activity | [37] | |
| Probiotic Strains | ||||
| Fermented pork, and tea leaves, | Lactobacillus fermentum | Probiotic | Strains were sensitive to some antibiotics and active against Escherichia coli, Salmonella Typhi, and Staphylococcus aureus. | [38] |
| Thai pickled vegetables (Phak-dong) | B. siamensis B44v | Probiotic in hybrid catfish culture | Bacteriocin production and activity against Gram-positive and Gram-negative pathogens. Sensitive to tested antibiotics. Protected the hybrid catfish from Aeromonas hydrophila FW52 infection. | [39] |
| Thai indigenous fermented foods | L. plantarum CR1T5 | Probiotic in tilapia fish culture | Activity against fish pathogens (Aeromonas caviae, A. hydrophila, and Streptococcus agalactiae). The supplementation of CR1T5 enhanced the survivability of infected fish, and also enhanced the growth. | [40] |
| Enzyme Production | ||||
| Fermented soybean (Thua-nao) | Bacillus spp. | γ-Glutamyl hydrolase production | The enzyme was about 28 kDa. The enzyme acted on the γ-glutamyl linkage of γ-polyglutamic acid. | [41] |
| Fermented pork | Pediococcus pentosaceus | Catalase production | Activity against S. Typhimurium. Hematin enhanced the catalase production, but was not necessary for production (i.e., heme-independent catalase). | [42] |
| Fermented soybean | L. plantarum SC359 | β-glucosidase production | Activity against representative microbial pathogens. The strain competed for pathogenic adherence to Caco-2 cells. | [43] |
| Many fermented foods | L. brevis, and L. fermentum | Glutaminase and glutamate decarboxylase production | The optimal conditions for the enzyme production were found at pH 6.5, temperature 40 °C, and 10 days of incubation time. | [44] |
| Fish sauce | Bacillus licheniformis RKK-04 | Serine proteinase production | Thermotolerant subtilisin-like alkaline serine proteinase enzyme. pH 10.0 and 50 °C as optimal conditions for enzyme production. The enzyme from RKK-04 could be used in fish sauce fermentation. | [45] |
| Gamma-Aminobutyric Acid Production | ||||
| Many fermented foods | L. namurensis, P. pentosaceus | γ-Aminobutyric acid (GABA) production | Four isolates were isolated and found to produce GABA (7339–9060 mg/L) in the culture medium. Two isolates (HN8 and NH2) were recommended for use in fermented pork as a starter culture. | [46] |
| Fermented shrimp (Kung-Som) | L. futsaii CS3 | GABA production | Reported primary GABA producer, and had the ability to convert 99% monosodium glutamate to GABA. | [47] |
| Exopolysaccharide Production | ||||
| Many fermented foods | P. pentosaceus (AP-1, and AP-3) | Exopolysaccharide Production | Dextran class of exopolysaccharide. | [48] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sivamaruthi, B.S.; Kesika, P.; Chaiyasut, C. Thai Fermented Foods as a Versatile Source of Bioactive Microorganisms—A Comprehensive Review. Sci. Pharm. 2018, 86, 37. https://doi.org/10.3390/scipharm86030037
AMA Style
Sivamaruthi BS, Kesika P, Chaiyasut C. Thai Fermented Foods as a Versatile Source of Bioactive Microorganisms—A Comprehensive Review. Scientia Pharmaceutica. 2018; 86(3):37. https://doi.org/10.3390/scipharm86030037
Chicago/Turabian StyleSivamaruthi, Bhagavathi Sundaram, Periyanaina Kesika, and Chaiyavat Chaiyasut. 2018. "Thai Fermented Foods as a Versatile Source of Bioactive Microorganisms—A Comprehensive Review" Scientia Pharmaceutica 86, no. 3: 37. https://doi.org/10.3390/scipharm86030037