Known and Unknown Features of Hair Cuticle Structure: A Brief Review

Molecular and Biomedical Sciences, University of Adelaide, Adelaide, SA 5005, Australia
Cosmetics 2019, 6(2), 32; https://doi.org/10.3390/cosmetics6020032
Submission received: 26 March 2019
/
Revised: 8 April 2019
/
Accepted: 9 April 2019
/
Published: 9 May 2019
(This article belongs to the Special Issue Recent Advance in Hair Science and Hair Care Technologies)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The cuticle is the outermost layer of overlapping flattened cells of hair and has been subjected to many years of study to understand its structure and how it develops in the follicle. The essential function of the cuticle with its tough inelastic protein content is to protect the inner cortex that provides the elastic properties of hair. Progress in our knowledge of hair came from studies with the electron microscope, initially transmission electron microscopy (TEM) for internal structure and later the scanning electron microscope (SEM) for cuticle surface shape and for investigating changes caused by various environmental influences such as cosmetic treatments and industrial processing of wool. Other physical techniques have been successfully applied in conjunction with proteomics. The outstanding internal features of the cuticle cells are the internal layers consisting of keratin filament proteins and the keratin-associated proteins. The stability and physical toughness of the cuticle cell is partly accounted for by the high content of disulphide crosslinking. The material between the cells that holds them tightly together, the cell membrane complex, consists of a layer of lipid on both sides of a central protein layer. The lipid contains 18-methyleicosanoic acid that is part of the hydrophobic lipid surface of hair. For the past decade there have been aspects that remained unanswered because they are difficult to study. Some of these are discussed in this brief review with suggestions for experimental approaches to shed more light.
1. Introduction
The cuticle of hair is vital in protecting the inner structure, the cortex, from damage caused by natural environmental factors, cosmetic treatments, industrial processes and the invention of new ones. It is a multiple layer in human hair (single layer in wool) of flattened overlapping and physically hard cells covering the hair cortex (Figure 1).
The outward orientation of the edges of the cells provides self-cleaning of the continuous desquamation of the epidermis.
Since the hair cuticle was studied by early histologists [1] a large literature has developed. It is briefly reviewed here with relevant references to our understanding of cuticle structure achieved through the use of techniques of cell biology. These include transmission electron microscopy (TEM), scanning EM (SEM), X-ray photoelectron spectroscopy (XPS), secondary ion mass spectroscopy (SIMS) and atomic force microscopy (AFM) in conjunction with protein analytical techniques. Research over some 70 years has revealed the major details of the internal structure of cuticle cells. It is important to also emphasise that all of these techniques have contributed to the knowledge of hair structure and properties generally.
2. The Three Layers in a Cuticle Cell
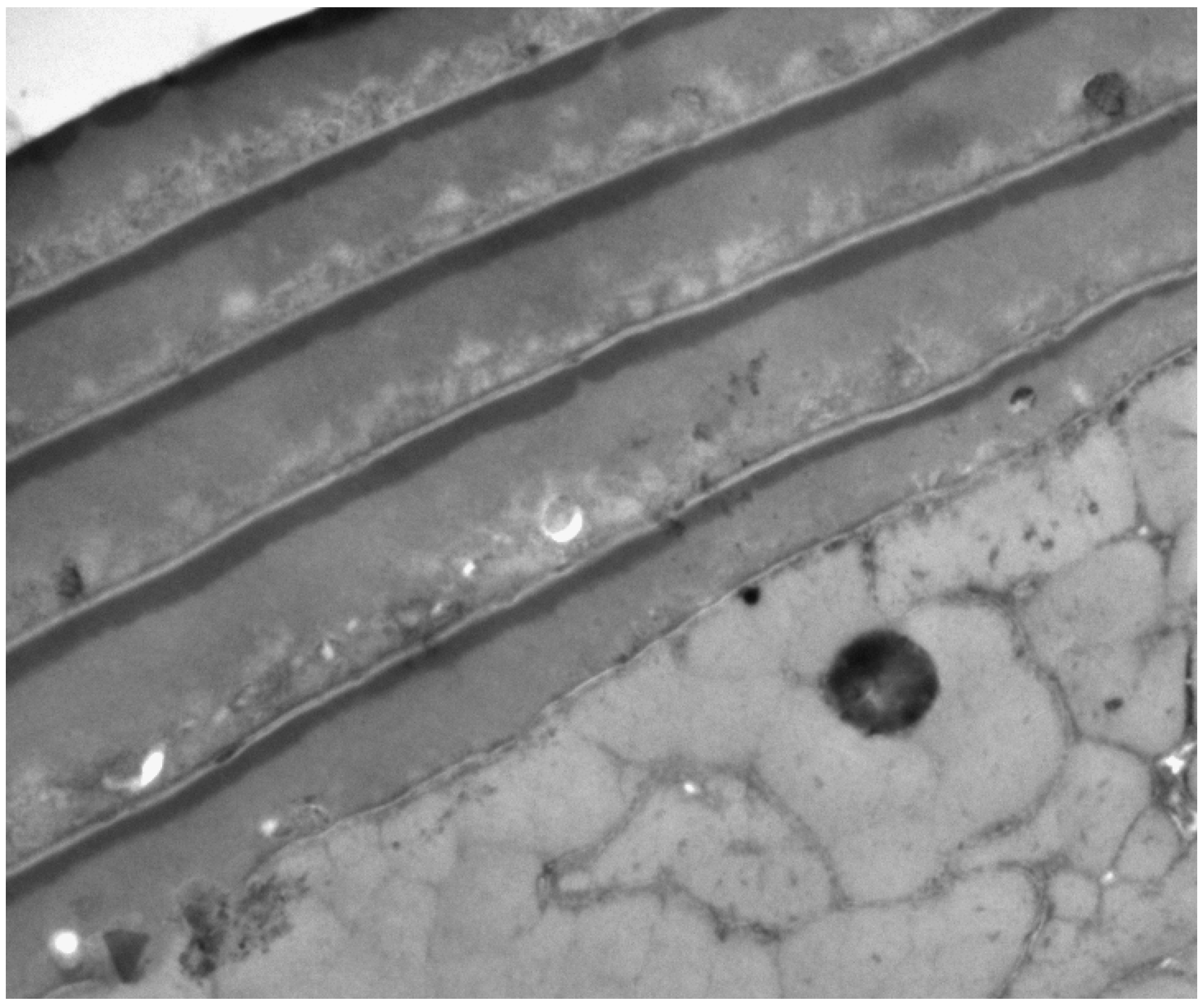
The outstanding feature is the three layers, A-layer, exocuticle and endocuticle (Figure 2) that were first distinguished in the TEM by their electron densities and produced according to their relative sulphur contents binding the metal stains of osmium, lead and mercury for TEM imaging [2,3].
The layer closest to the upper surface of each cuticle cell is the densely-stained A-layer then the exocuticle and beneath that the endocuticle.
During hair growth and protein synthesis, the cuticle and cortex proteins are synthesised with the sulphur in the reduced state as cysteinyl residues. As a consequence the metal stains, osmic acid, lead citrate and mercuric iodide readily react to increase electron density of the keratin and associated keratin proteins until cross-linking occurs about half way along the follicle whence the staining diminishes. A TEM study of the developing cuticle cell [4,5] showed that the exocuticle and A-layer develop as small (Type 1) and large droplets (Type 2) of protein. The small droplets appear to be the precursors of the A-layer and the larger ones, the exocuticle (Figure 3).
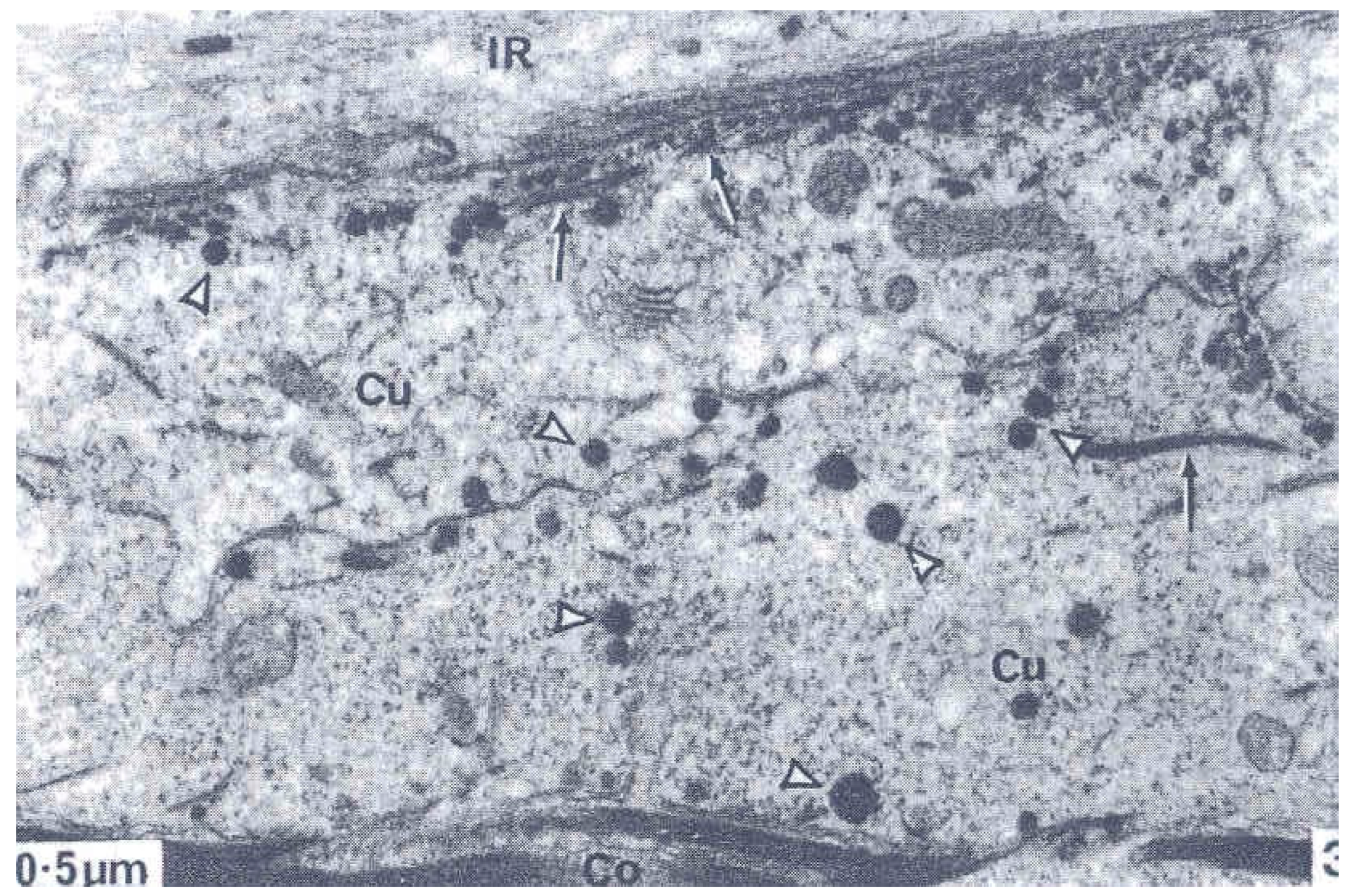
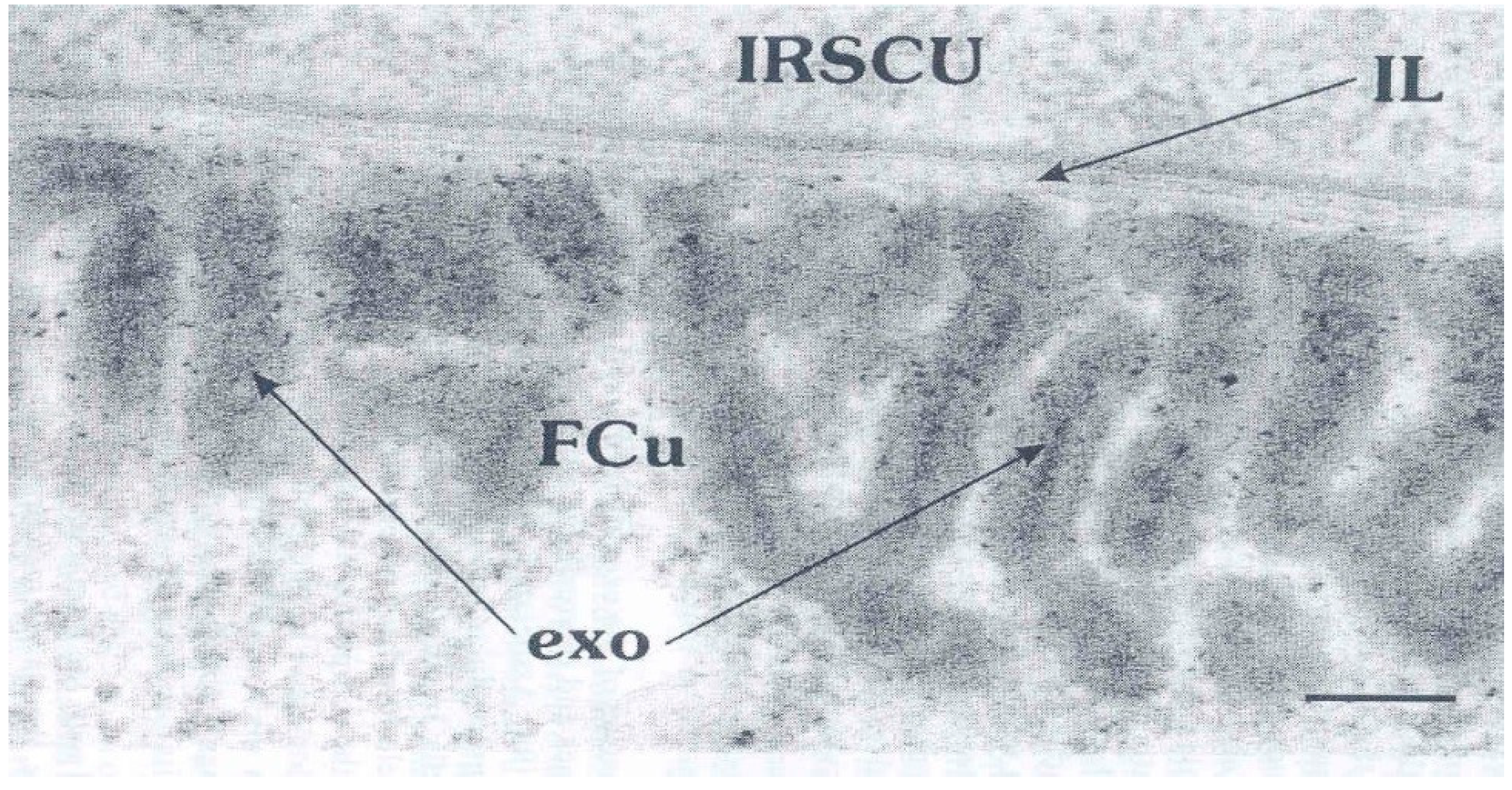
These dynamic events imaged in the TEM as static changes suggested that the smaller droplets form the outermost A-layer. They migrate and aggregate including with keratin filaments to form the amorphous A-layer beneath the outermost plasma membrane of the cell. The larger droplets as precursors of the exocuticle increase in size, they also migrate outwards to form a layer of complex lamellae. In some orientations a sparse central denser layer is discernible in the lamellae (Figure 4) [6], suggesting that some dense A-layer protein is incorporated into the exocuticle laminae. There is a gap in our knowledge here because there are no TEMs showing the development of this event.
These structures finally move to abut the A-layer and fuse with it to form a uniform mass, the exocuticle (Figure 5).
An important discovery in these processes was the presence of fatty acids covalently linked to the cuticle surface. They include stearic and palmitic acids but especially an unusual branched chain fatty acid, anteiso-18-methyleicosanoic acid (18-MEA) [7,8,9]. This finding explained the hydrophobic properties of the hair surface. According to the best evidence 18-MEA can constitute up to 70% of the covalently bound fatty acids that entirely cover the fibre surface. The abundance of 18-MEA relative to the straight chain fatty acids is indicative of a particular function such as stability of the hydrophobic surface or influencing the friction on the surface but answers to these questions need further research. The fatty acids are attached by thioester bonds [9] to a protein underlay, yet to be certainly identified. The surface lipid of 18-MEA was estimated to be 0.9 nm thick [10] and was also imaged by TEM as a ‘fuzzy’ surface layer [11]. The MEA molecules are longer than the hydrophobic layer and could be folded to be accommodated in the 0.9 nm space [12].
The endocuticle, compared with the exocuticle, is a completely different layer, a loose amorphous mass that appears to be cytoplasmic residues of protein except for the presence of the calcium-binding protein S100E [13]. Its function is unknown.
3. The Epicuticle
The epicuticle was the first sub-structure of the cuticle to be studied in the TEM [14]. Its discovery came about when hairs or wool were treated with chlorine saturated water and was called the Allworden reaction [15] after the author who first described it. Chlorine was known to reduce the frictional properties of wool; its effect is to produce microscopic membranous bubbles on the surface but does not otherwise destroy the fibre structure. The epicuticle is a derived membranous structure estimated to be 50–70 nm thick. Initially the epicuticle was thought to cover only the hair surface. Further study indicated it that it totally surrounds the surface of individual cuticle cells [16]. The origin of its composition is uncertain. It is mainly protein [17] and the covalently bound lipids, mainly 18-MEA, are cleaved from the surface by the chlorine [9]. The A-layer is thought to be involved because of the sulphur content that could provide the sulphydryl groups for the covalent anchorage of the fatty acids. The A-layer is variable in thickness so if it totally cleaved from the exocuticle its contribution to the epicuticle thickness from wool would be of the order of 70 nm [5]. Measurements of epicuticle thickness provided a value of 5 nm [12] and given that figure, only part of the A-layer could be involved. The stability of the epicuticle protein to vigorous reagents has been attributed to isopeptide cross-linking [6]. This is not necessarily the case because the inner root sheath and the medulla proteins have a high isopeptide content and yet are degraded by proteases to peptides [18,19,20]. More likely, resistance to degradation is the result of collaborative forces of hydrogen bonds, disulphide bonds and isopeptide cross-links in a beta pleated sheet structure.
4. The Cell Membrane Complex (CMC)
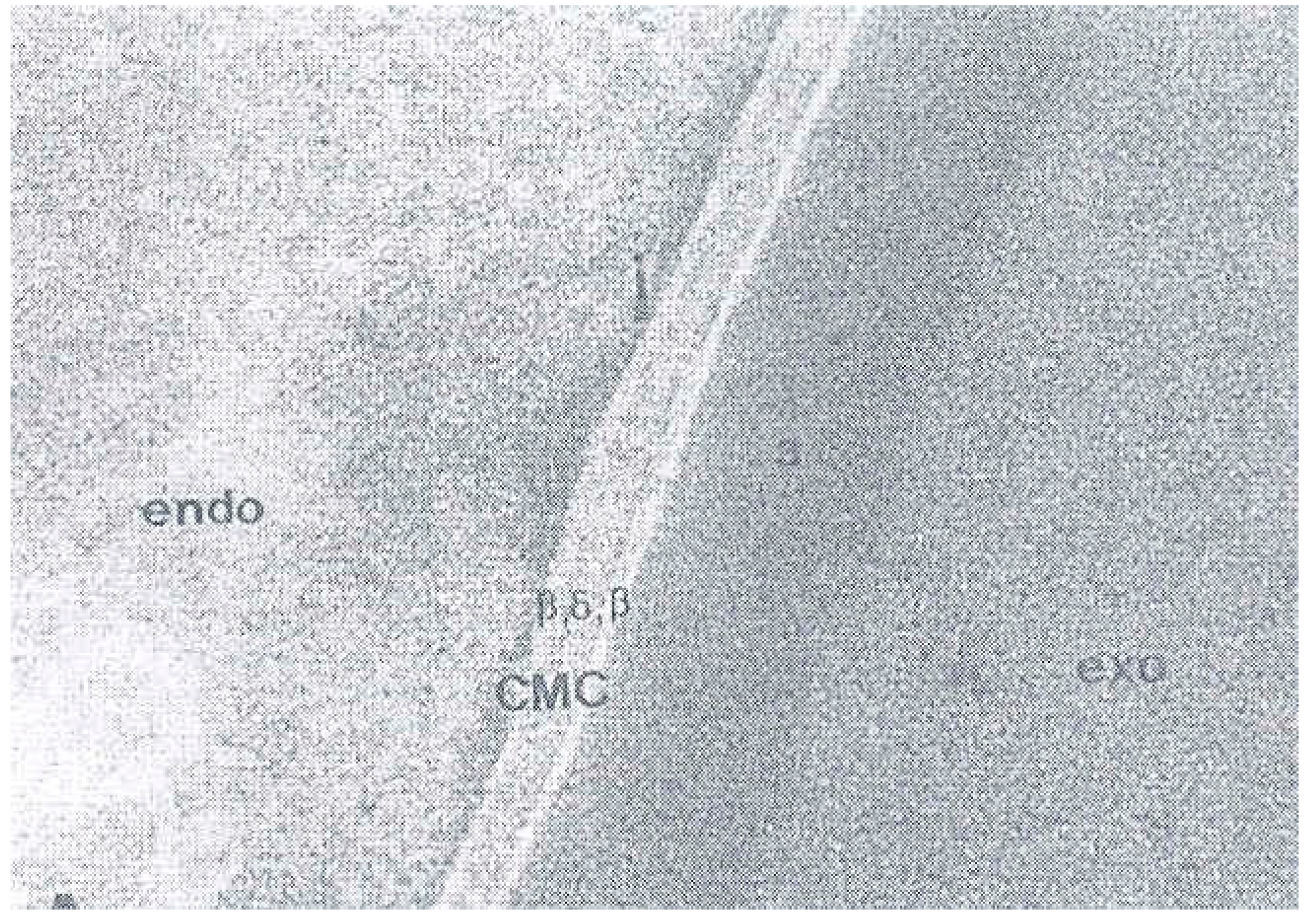
A fascinating aspect of cuticle structure is the change that the plasma membranes between cuticle cells (and cuticle-cortical) boundaries undergo in the development of the cuticle during growth of the hair. This change has been the subject of extensive reports and reviews [21,22]. Very little is known of their formation and their composition and most of our knowledge comes from TEM [22] and also AFM [23] studies of keratinised hair or wool. The early cuticle intercellular membrane is a typical trilaminar unit membrane consisting of a bilipid layer flanked by protein layers and an intercellular space between adjacent cells. However, at later stages of hair growth the separation of the nearly keratinised cuticle cells involves a very different structure, the cell membrane complex. A central denser layer, the δ-layer, presumably protein, develops between adjacent cuticle cells and is flanked by two low-density β-layers that are each interpreted to be a bilipid layer (Figure 6).
TEM studies of the membrane complex of the hair cuticle from subjects with a mutation of the 18-MEA synthetic pathway (Maple Syrup Urine disease (MSUD) [24] revealed that the likely position of the MEA is on only one face, the upper surface of each cuticle cell. It is this surface layer of 18-MEA and other fatty acids that is responsible for the hydrophobic fibre surface (Figure 7). How the CMC cleaves to expose the underlying lipid has been proposed by Jones [6]. What properties the 18-MEA provide for hair is unclear but it has been suggested that since 18-MEA has the branched chain, the melting point is much lower than that of the straight chain and as consequence the lipid layer on the cuticle surface would be more fluid. That could alter frictional properties.
5. Discussion: Some Unknown Aspects of Cuticle Structure that Need Innovative Techniques to Resolve
The exocuticle:
The formation of the droplets of exocuticle precursor, their fusion and migration to the outer surface are fascinating processes. Any study of this must also explain the distinct development of the A-layer with its significantly higher cystine content. The droplets appear to form by self-aggregation of the protein components. To study that possibility would require preparation of those proteins. Only KAP5 and KAP10 are known to be major components, so more protein analysis of the exocuticle is necessary to identify others. The gene sequences are known for KAP5 and KAP10 and could be prepared by expression in bacterial systems. Study of their interaction would then be amenable by standard proteomics methods such as those used in studying the binding of a KAP to an IF keratin [25]. The migration aspect however, is more difficult to study because it is a dynamic event. Could it be driven by a microtubule pathway? There is no report to indicate their involvement, so a thorough TEM study at many follicle levels using immunochemical tracking would be useful.
The epicuticle
Amino acid analyses have shown that there are cysteine residues to provide anchorage of fatty acids (the links are cleaved by the chlorination to elicit the epicuticle). As for the origin of the proteins, there is strong evidence indicating the involvement of the A-layer [26]. However it is possible that proteins from the cell membrane complex that compose the intercellular cement are also involved. In an effort to seek the proteins that have 18-MEA and other fatty acids attached by thioester bonds, experiments were conducted on fibres before keratinisation (G. Rogers and K. Koike, unpublished data). By using this approach, the keratin structures were not cross-linked and more amenable to studies in solution. The proteins such as the cell membrane complex were expected to be present in low abundance, so we faced a quantitative problem. Consequently, we used follicles epilated from sheepskin; thousands of follicles in anagen phase can be obtained. Their proteins were solubilised with 8M urea and the SH groups blocked with iodoacetamide. In extensive studies the authors used several different methods to identify any proteins with fatty acids covalently linked to cysteinyl groups. The most direct technique was to tag the acylated sites using acyl-biotin exchange [27]. This procedure entailed incubating the urea-soluble iodoacetamide-blocked proteins with hydroxylamine at 20 °C, which specifically breaks thioester linkages and releases the fatty acids. Any exposed SH groups of the protein were then tagged with a biotin moiety and any biotin- tagged proteins were captured with streptavidin beads. Unbound proteins were removed by washing and any bound proteins released from the beads. After concentrating the eluates, the proteins were examined by gel electrophoresis. The controls for the experiments were identical but with omission of the hydroxylamine step. After several attempts we could not obtain consistent results. We suspect that this was caused by an unexpected problem with the method. We found that the hydroxylamine in the presence of 8 M urea produced a gel that could have interrupted the hydrolysis. Gelation occurred in the absence of protein and no explanation could be found in the literature. These acyl-biotin exchange experiments need repeating but using other chaotropic agents to replace the urea, such as guanidine thiocyanate, or lithium salts, to avoid the gelation phenomenon but to keep the proteins in the solution.
6. At What Stage of Hair Growth is 18-MEA Attached to the Cuticle and How Does This Occur?
The synthesis of 18-MEA occurs in the liver, it circulates in the blood and is available for capture and enzymic transfer to thiol groups presumably by a transferase enzyme. The known location of 18-MEA in hair is on the upper surface of the flattened cuticle cells. This asymmetry as a structural feature of the β-layer and the question of how the presence of 18-MEA attached on one face only occurs is curious and remains without explanation.
We don’t know at what follicle level 18-MEA is coupled to cuticle protein but it must occur when the proteins have sulphydryl groups available for acylation with fatty acids; the region defined as the elongation zone of the follicle [4] begins proximal to the hair bulb.
One feasible method to locate these events would be to utilise TEM autoradiography. That would require injection of sufficient tritiated 18-MEA into a mouse or young guinea pig, preferably the latter because most of the follicles are in anagen. After selected times skin biopsies would be taken, processed, submitted to autoradiographic detection and examined in TEM. The problem here is that tritiated or C14-labelled 18-MEA are not commercially available. However, they could be synthesised for this purpose. Carbon-14 is usable for autoradiography at the TEM level but is less efficient than tritium.
7. Summary
We now have a comprehensive view of the structure of the hair cuticle cell as a result of the application of an array of physical and chemical techniques. The power of TEM has played a central role because of the miniscule nature of the hair follicle and the macromolecular components of the hair. Some unanswered questions have been discussed in this essay and clearly they are of fundamental interest but they will be difficult to solve and undoubtedly will require novel approaches. To justify future research in that direction will probably largely depend on the applicability of further knowledge to the usage and treatment of hair.
Funding
The preparation of this review received no external funding. The author acknowledges the provision of writing facilities by the Department of Molecular and Biomedical Science.
Conflicts of Interest
The author declares no conflict of interest.
References
- Auber, L. The anatomy of follicles producing wool fibres with special reference to keratinization. Trans. R. Soc. Edinb. 1951, 62 Pt 1, 191–254. [Google Scholar] [CrossRef]
- Rogers, G.E. Electron microscope studies of hair and wool. Ann. N. Y. Acad. Sci. 1959, 83, 378–399. [Google Scholar] [CrossRef] [PubMed]
- Rogers, G.E. Electron. microscopy of wool. J. Ultrastruct. Res. 1959, 2, 309–330. [Google Scholar] [CrossRef]
- Woods, L.J.; Orwin, D.F.G. Studies on the surface layers of the wool fibre cuticle. In Fibrous Proteins: Scientific, Medical and Industrial Aspects; Parry, D.A.D., Creamer, L.K., Eds.; Academic Press: London, UK, 1980; pp. 141–150. [Google Scholar]
- Orwin, D.F.G. The cytology and cytochemistry of the wool follicle. Int. Rev. Cytol. 1979, 60, 331–374. [Google Scholar]
- Jones, L.N. The hair fibre surface. In Skin, Hair and Nails; Forslind, B., Lindberg, M., Eds.; CRC: Boca Raton, FL, USA, 2003; pp. 285–316. [Google Scholar]
- Evans, D.J.; Leeder, J.D.; Rippon, J.A.; Rivett, D.E. Separation and analysis of surface lipids of the wool fibre. In Proceedings of the 7th International Wool Textile Research Conference, Tokyo, Japan, 28 August–3 September 1985; pp. 181–193. [Google Scholar]
- Negri, A.P.; Cornell, H.J.; Rivett, D.E. The nature of covalently bound fatty acids in wool fibres. Aust. J. Agric. Res. 1991, 42, 1285–1292. [Google Scholar] [CrossRef]
- Negri, A.P.; Cornell, H.J.; Rivett, D.E. A model for the surface of keratin fibres. Text. Res. J. 1993, 63, 109–115. [Google Scholar] [CrossRef]
- Ward, R.J.; Willis, H.A.; George, G.A.; Guise, G.B.; Denning, R.J.; Evans, D.J.; Short, R.D. Surface analysis of wool by X-ray photoelectron spectroscopy and static secondary ion mass spectrometry. Text. Res. J. 1993, 63, 362–368. [Google Scholar] [CrossRef]
- Jones, N.L.; Rivett, D.E. The role of 18-methyleicosanoic acid in the structure and formation of mammalian hair fibres. Micron 1997, 28, 469–485. [Google Scholar] [CrossRef]
- Zahn, H.; Messinger, H.; Hocker, H. Covalently-linked fatty acids at the surface of wool: Part of the “cuticle cell envelope. Text. Res. J. 1994, 64, 554–555. [Google Scholar] [CrossRef]
- Kizawa, K.; Uchiwa, H.; Murakami, U. Highly expressed S100A3, a calcium binding protein in human hair cuticle. Biochim. Biophys. Acta 1996, 13, 94–98. [Google Scholar] [CrossRef]
- Lindberg, J.; Philip, B.; Gralen, N. Occurrence of thin membranes in the structure of wool. Nature 1948, 162, 458. [Google Scholar] [CrossRef]
- vonAllworden, K. The properties of wool and a new chemical method for detecting damaged wool. Angew. Chem. 1916, 29, 77. [Google Scholar]
- Bradbury, H.J.; Leeder, J.D. Keratin fibres. IV. Structure of the cuticle. Aust. J. Biol. Sci. 1970, 23, 843–854. [Google Scholar] [CrossRef] [PubMed]
- King, R.N.L.; Bradbury, J.H. The chemical composition of wool. V: The epicuticle. Aust. J. Biol. Sci. 1968, 21, 375–384. [Google Scholar] [CrossRef]
- Rogers, G.E.; Harding, H.W.J.; Llewellyn-Smith, I.J. The origin of citrulline-containing proteins in the hair follicle and the chemical nature of trichohyalin, an intracellular precursor. Biochim. Biophys. Acta 1977, 495, 159–175. [Google Scholar] [CrossRef]
- Rogers, G.E. Some observations on the proteins of the inner root sheath cells of hair follicles. Biochim. Biophys. Acta 1958, 29, 33–43. [Google Scholar] [CrossRef]
- Rogers, G.E. Newer findings on the enzymes and proteins of hair follicles. Ann. N. Y. Acad. Sci. 1959, 83, 408–428. [Google Scholar] [CrossRef]
- Leeder, J.D. Wool-Nature’s Wonder Fibre; Australasian Textile Publishers: Ocean Grove, Australia, 1984. [Google Scholar]
- Jones, L.N.; Horr, T.J.; Kaplin, I.J. Formation of surface membranes in developing mammalian hair fibres. Micron 1994, 25, 589–595. [Google Scholar] [CrossRef]
- Swift, A.J.; Bews, B. The chemistry of human hair cuticle- II: The isolation and amino acid analysis of the cell membranes and A-layer. J. Soc. Cosmet. Chem. 1974, 25, 355–366. [Google Scholar]
- Jones, L.N.; Peet, D.J.; Danks, D.M.; Negri, A.P.; Rivett, D.E. Hairs from patients with maple syrup urine disease show a structural defect in the fiber cuticle. J. Investig. Dermatol. 1996, 106, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, H.; Fujimoto, A.; Farooq, M.; Ito, M.; Shimomura, Y. Characterization of the human hair shaft cuticle-specific keratin-associated protein 10 family. J. Investig. Dermatol. 2013, 133, 2780–2782. [Google Scholar] [CrossRef] [PubMed]
- Bringans, S.D.; Plowman, J.E.; Dyer, J.M.; Clerens, S.; Vernon, J.A.; Bryson, W.G. Characterization of the exocuticle a-layer proteins of wool. Exp. Dermatol. 2007, 16, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Drisdel, C.R.; Green, W.N. Labeling and quantifying sites of protein palmitoylation. Biotechniques 2004, 36, 276–285. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(a) SEM of wool fibre ×1500; (b) SEM of hair fibre ×400.

Figure 2.
TEM of cross-section of human hair with five overlapping cuticle cells. Each cell has a dense A-layer and a less dense exocuticle beneath. The lower layer is endocuticle. ×35,000.
Figure 2.
TEM of cross-section of human hair with five overlapping cuticle cells. Each cell has a dense A-layer and a less dense exocuticle beneath. The lower layer is endocuticle. ×35,000.

Figure 3.
Early stage f formation of A-layer and exocuticle showing type 1 droplets migrated to cell membrane and mixed with filaments (black arrow). The larger Type 2 droplets nearby are in the cytoplasm (arrowheads). From reference [4].
Figure 3.
Early stage f formation of A-layer and exocuticle showing type 1 droplets migrated to cell membrane and mixed with filaments (black arrow). The larger Type 2 droplets nearby are in the cytoplasm (arrowheads). From reference [4].

Figure 4.
Late stage of exocuticle (exo) formation in a cuticle cell (FCU) adjacent to inner root sheath cell (IRSCU) and separated by an intercellular CMC layer (IL). Note the inner dark straining in the exocuticle laminathat might be A-layer protein. This stage is markedly different from that in Figure 3. Provided by Dr. L Jones.
Figure 4.
Late stage of exocuticle (exo) formation in a cuticle cell (FCU) adjacent to inner root sheath cell (IRSCU) and separated by an intercellular CMC layer (IL). Note the inner dark straining in the exocuticle laminathat might be A-layer protein. This stage is markedly different from that in Figure 3. Provided by Dr. L Jones.

Figure 5.
A cuticle cell of a wool fibre with almost complete consolidation of the exocuticle (ex) and the denser A-layer at the surface (arrows). From reference [4].
Figure 5.
A cuticle cell of a wool fibre with almost complete consolidation of the exocuticle (ex) and the denser A-layer at the surface (arrows). From reference [4].

Figure 6.
The cell membrane complex between two cuticle cells showing the central dense δ—layer and flanked by the two low density β-layers. From reference [24].
Figure 6.
The cell membrane complex between two cuticle cells showing the central dense δ—layer and flanked by the two low density β-layers. From reference [24].

Figure 7.
The cell membrane complex in a Maple syrup disease hair showing the disruption of the β-layer on the upper surface of each cuticle cell (FCU). This demontrates that the stability of the layer is dependent upon the preence of 18-MEA. From reference [24].
Figure 7.
The cell membrane complex in a Maple syrup disease hair showing the disruption of the β-layer on the upper surface of each cuticle cell (FCU). This demontrates that the stability of the layer is dependent upon the preence of 18-MEA. From reference [24].

© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rogers, G.E. Known and Unknown Features of Hair Cuticle Structure: A Brief Review. Cosmetics 2019, 6, 32. https://doi.org/10.3390/cosmetics6020032
AMA Style
Rogers GE. Known and Unknown Features of Hair Cuticle Structure: A Brief Review. Cosmetics. 2019; 6(2):32. https://doi.org/10.3390/cosmetics6020032
Chicago/Turabian StyleRogers, George E. 2019. "Known and Unknown Features of Hair Cuticle Structure: A Brief Review" Cosmetics 6, no. 2: 32. https://doi.org/10.3390/cosmetics6020032
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.