Evaluation of the Effect of Plant Mixture Ethanol Extracts Containing Biota orientalis L. Extract on Suppression of Sebum in Cultured Sebocytes and on Stimulation of Growth of Keratinocytes Co-cultured with Hair Papilla Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Samples
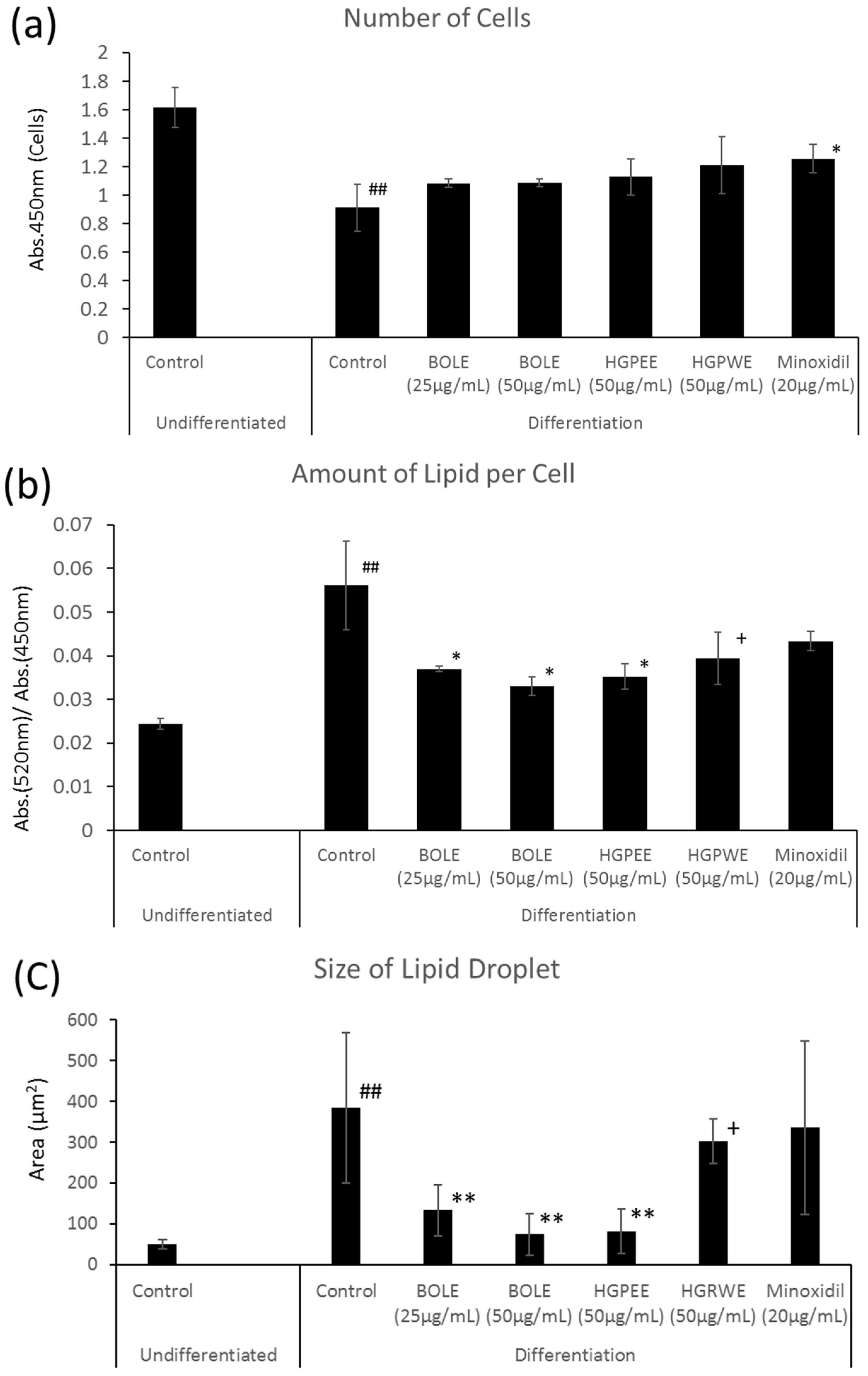
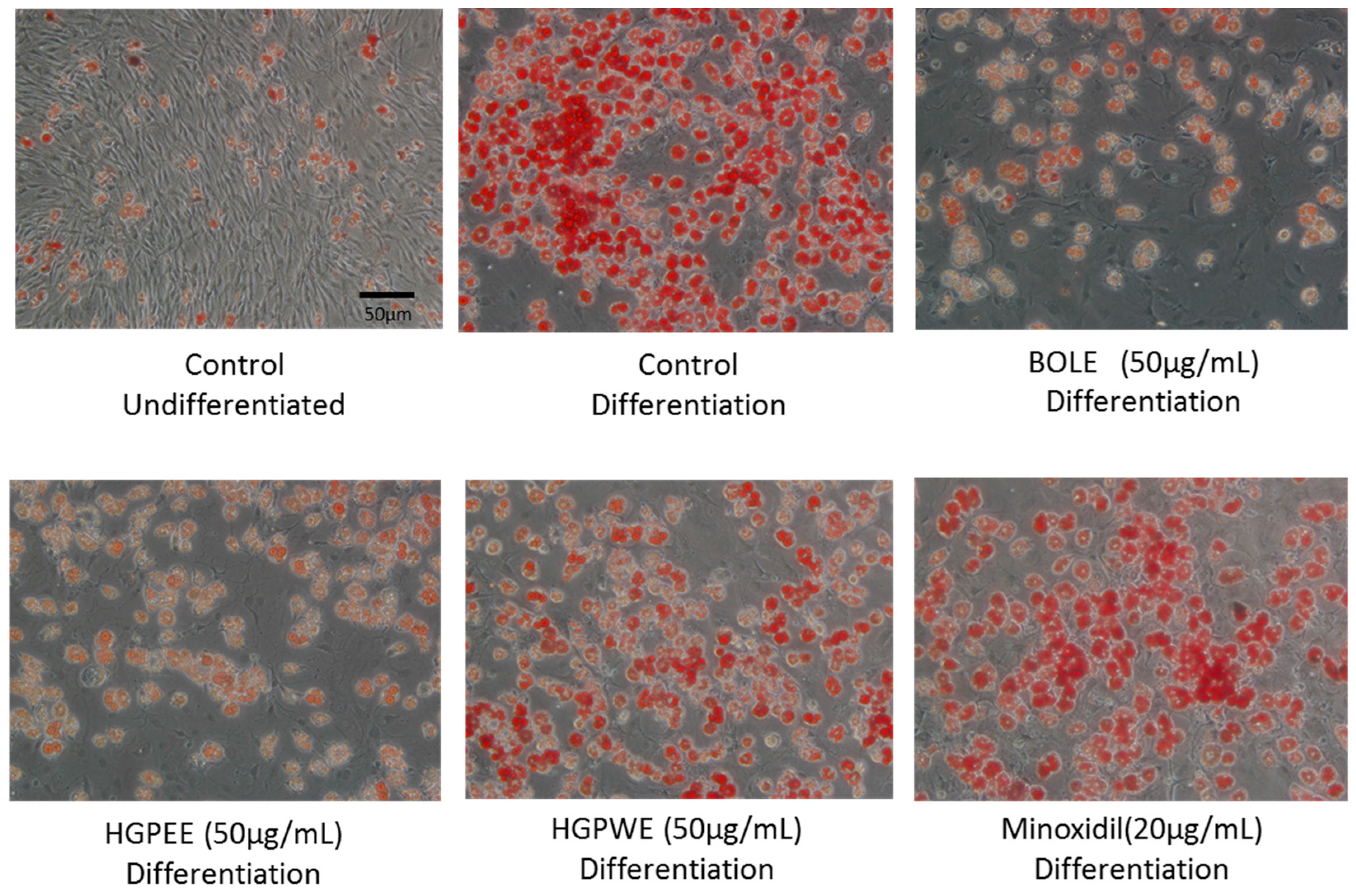
2.3. Measurement of Lipids in Sebocytes
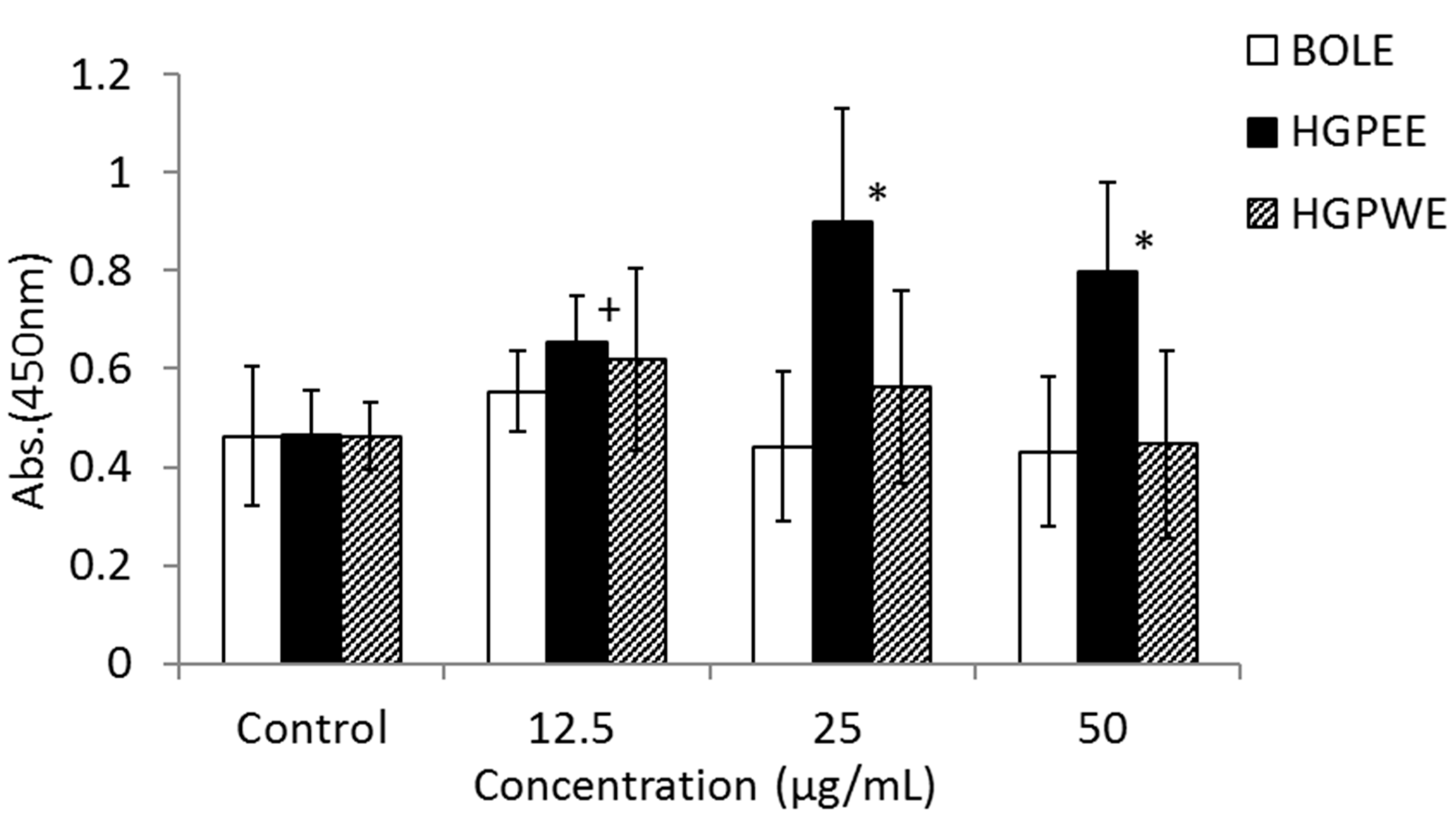
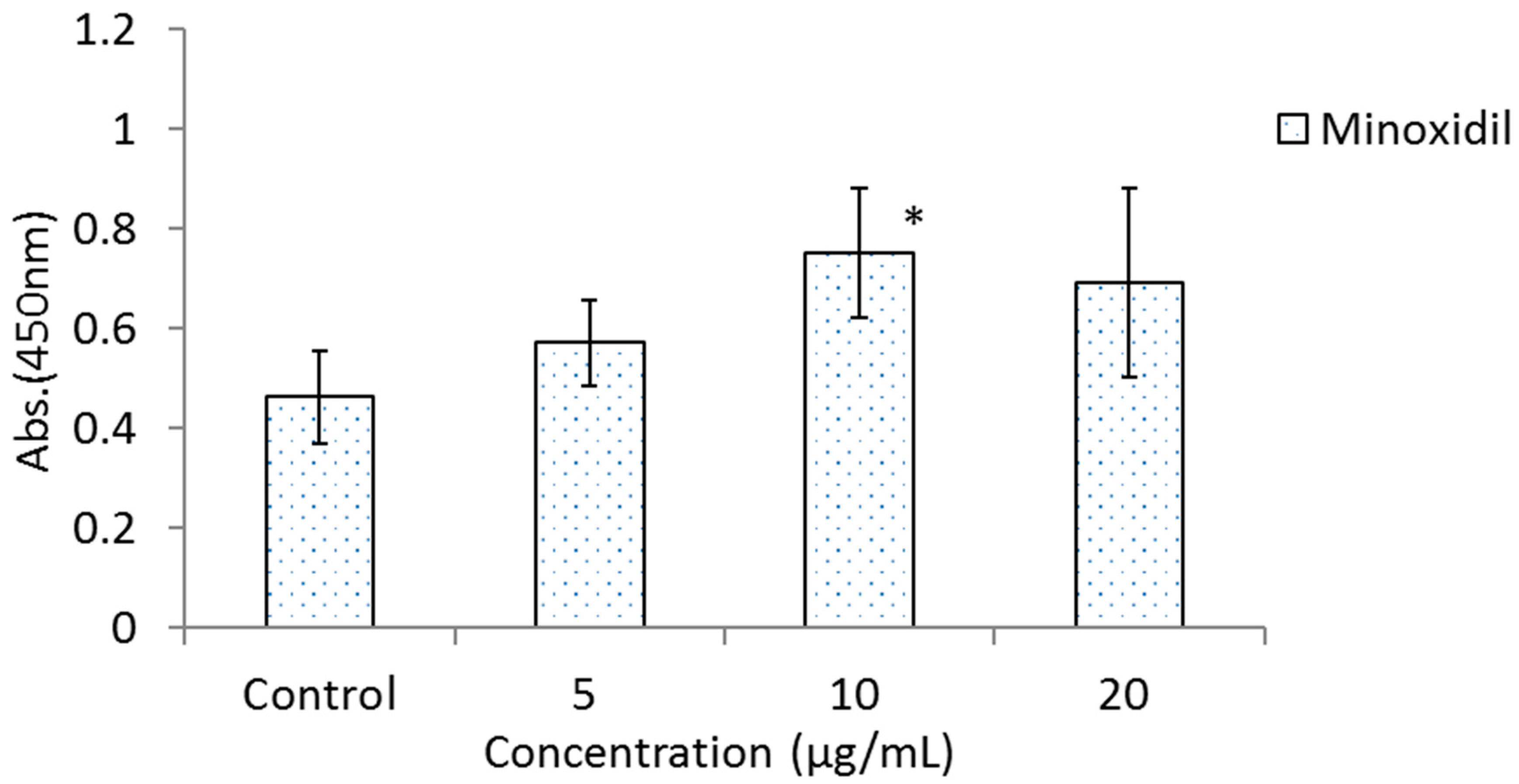
2.4. Effect on Growth of Human Fetal Epidermal Keratinocytes in a Two-Layer Culture System of Human Hair Papilla Cells and Human Epidermal Keratinocytes
2.5. Influence on Gene Expression Related to Hair Growth/Restoration
2.6. Statistical Analysis
3. Results
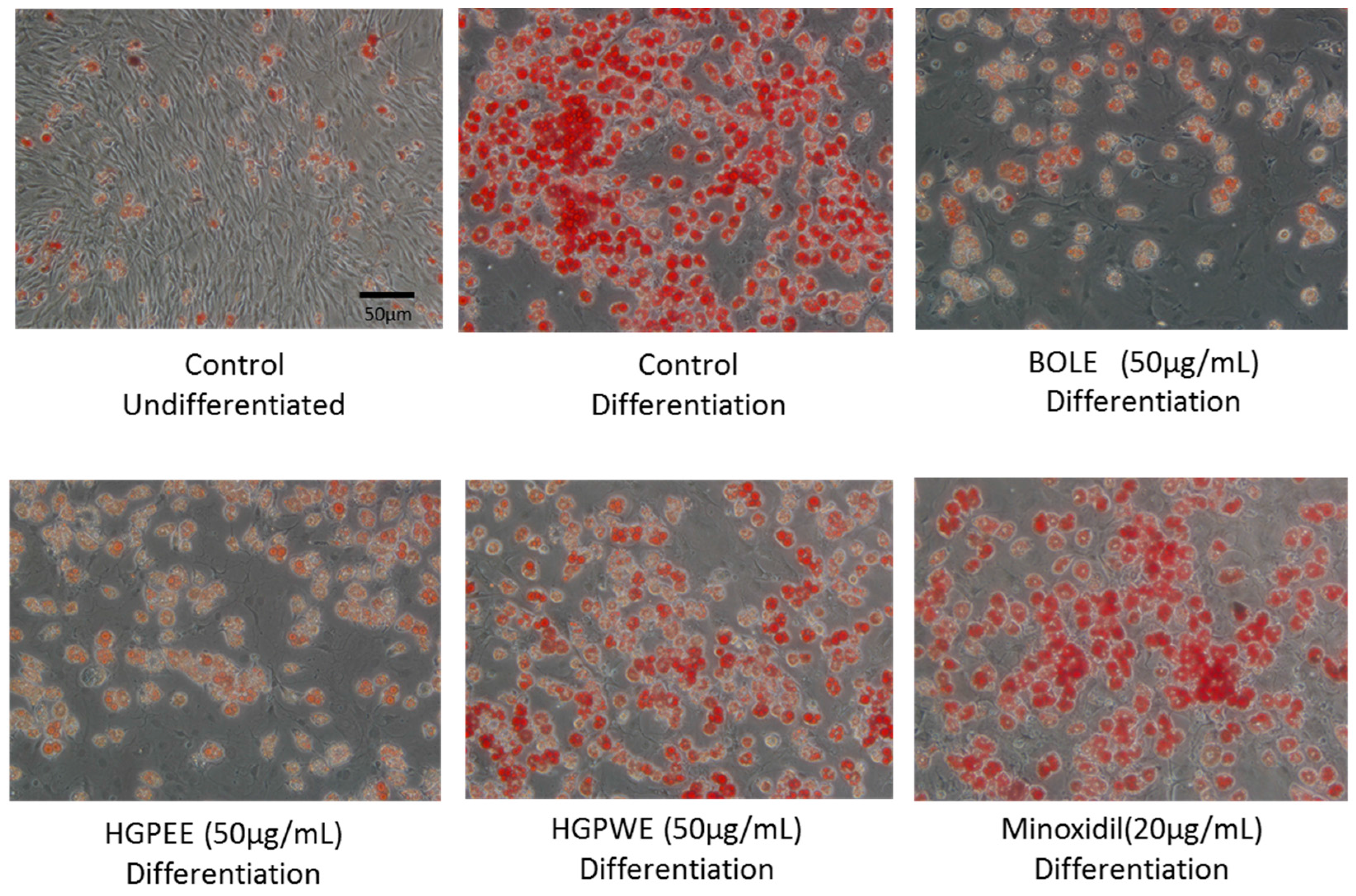
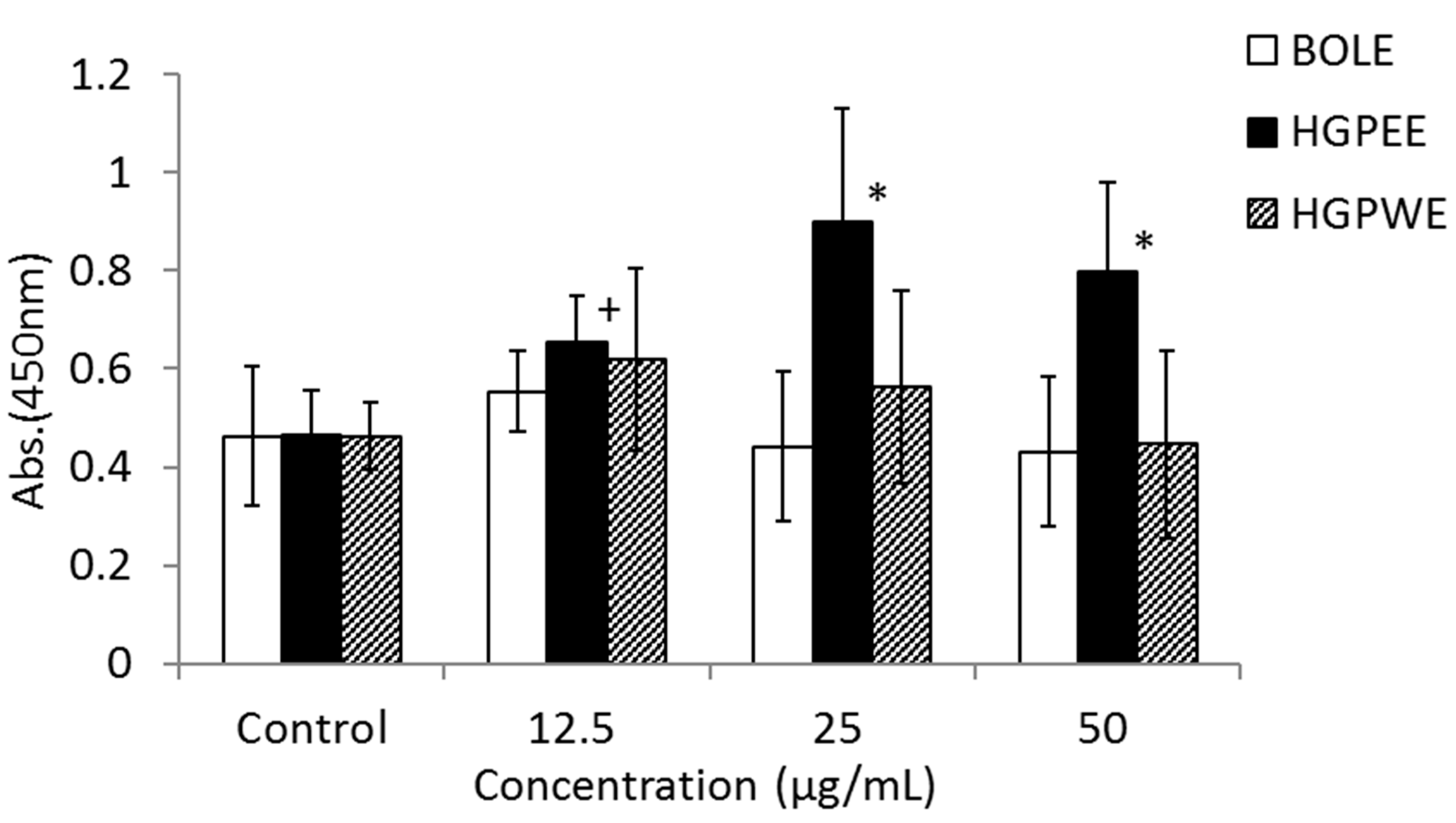
3.1. Effects of Biota Orientalis L. Leaf Extract (BOLE), Hair Growth Plant Ethanol Extract (HGPEE), Hair Growth Plant Ethanol Extract (HGPEE), and Minoxidil on the Proliferation of Sebocytes and Amount of Lipid Droplets
3.2. Effects of BOLE, HGPEE, HGPWE, and Minoxidil on the Growth of Human Fetal Epidermal Keratinocytes in a Two-Layer Culture System of Human Hair Papilla Cells and Human Epidermal Keratinocytes
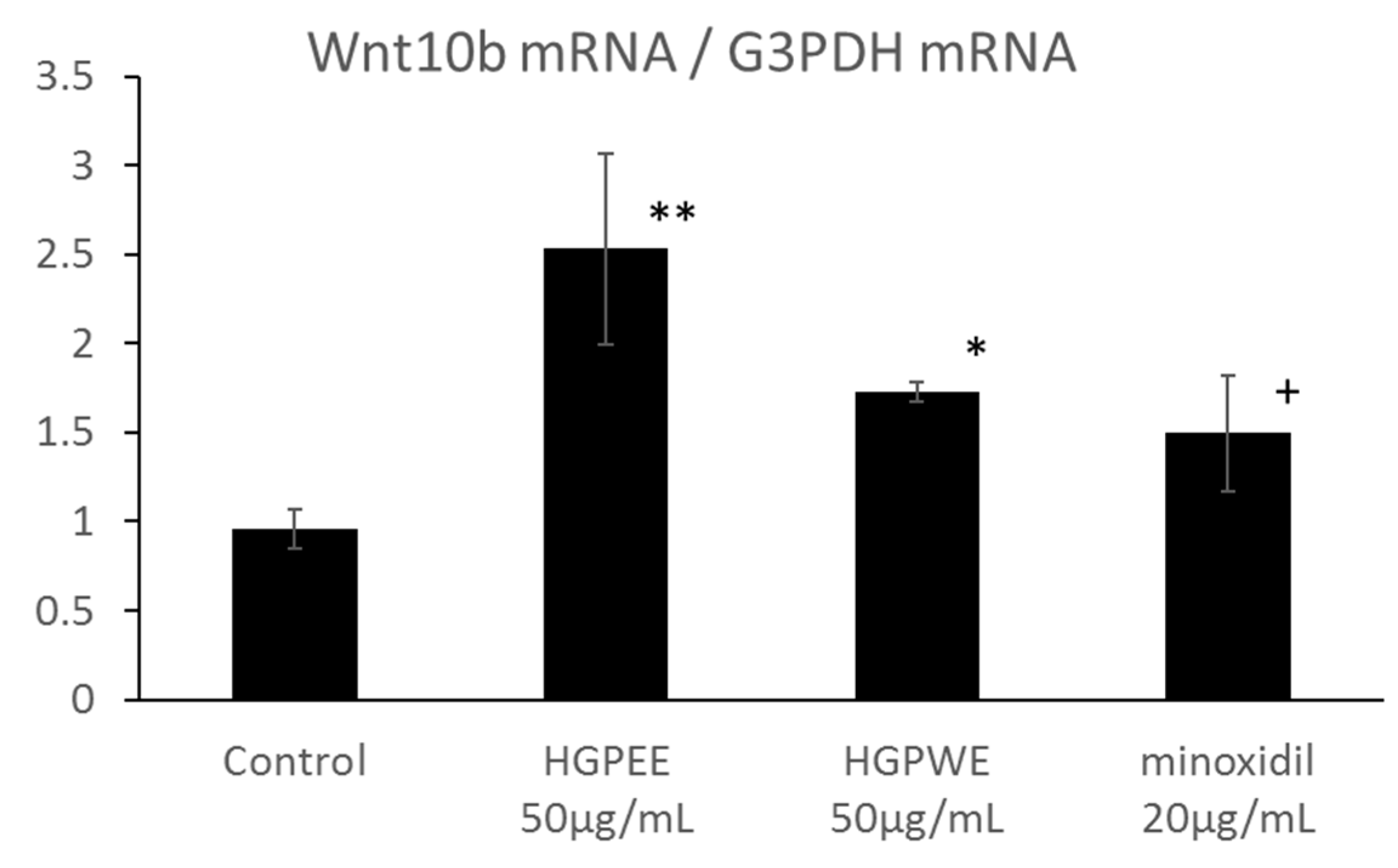
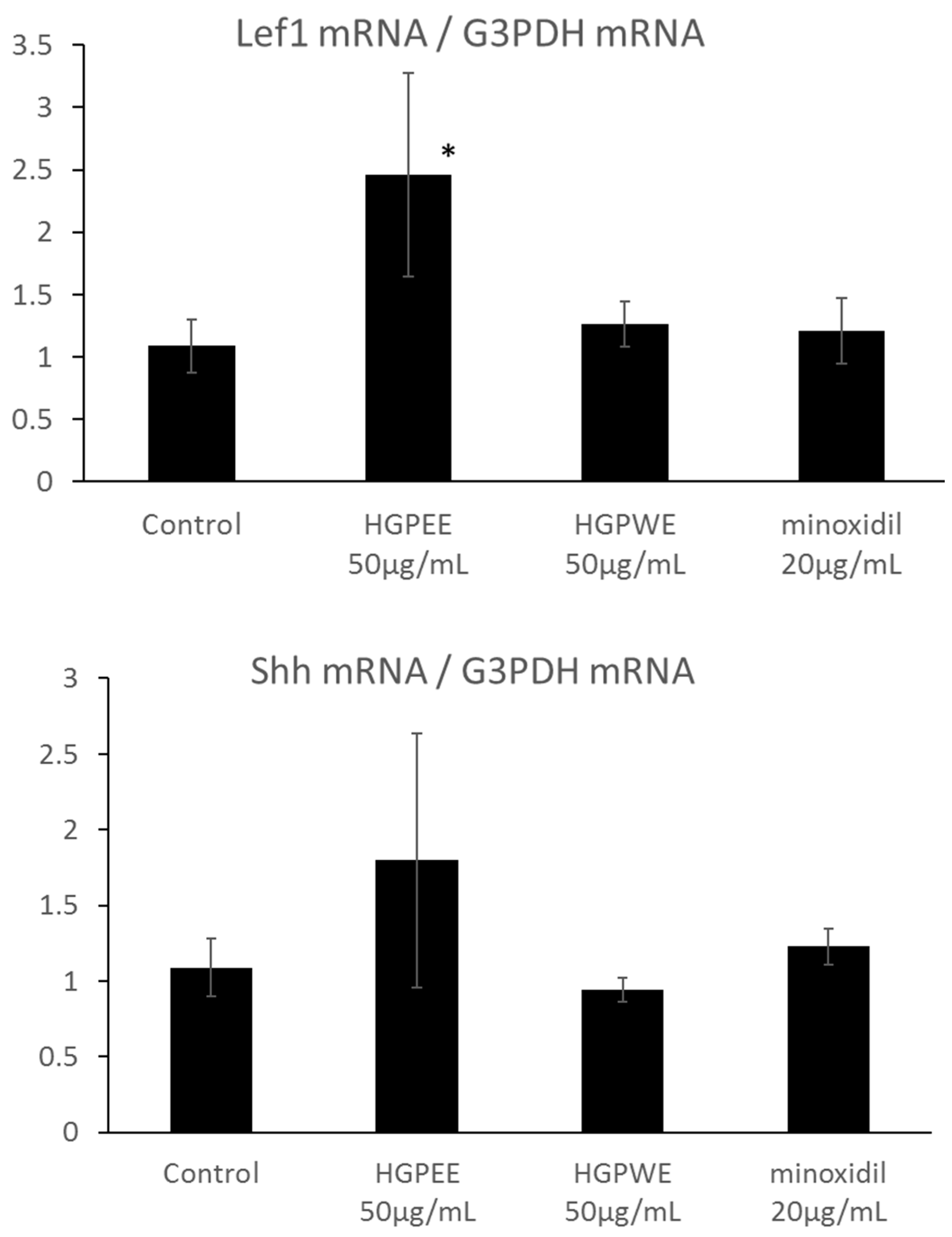
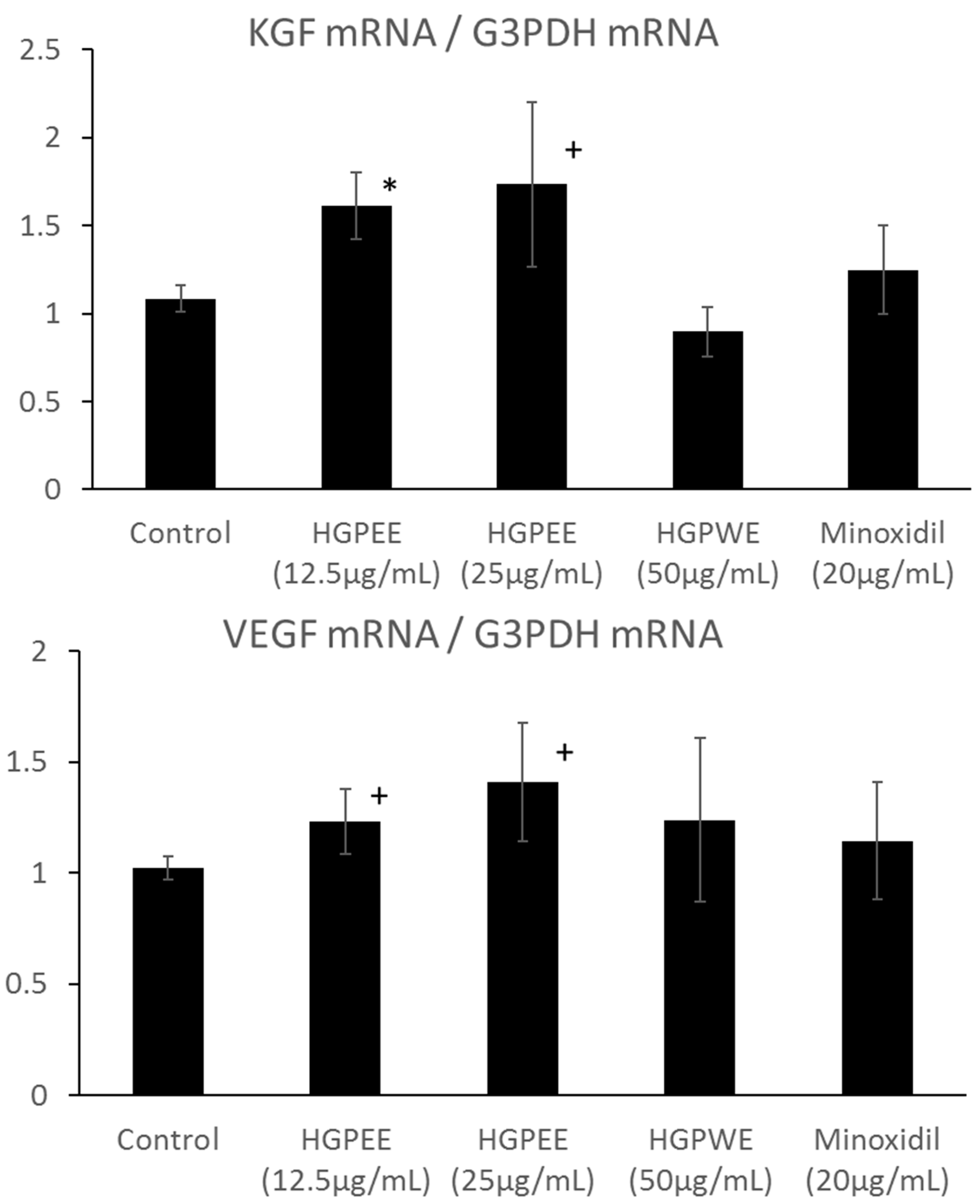
3.3. Effects of HGPEE, HGPWE, and Minoxidil on Expression of mRNA in Human Fetal Epidermal Keratinocytes Related to Hair Growth
3.4. Effects of HGPEE, HGPWE, and Minoxidil on the mRNA Expressions of Hair Growth-Related Factors in Human Hair Papilla Cells
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Ethics Approval and Consent to Participate
Funding
References
- Kellum, R.E. Human sebaceous gland lipids. Analysis by thin-layer chromatography. Arch. Dermatol. 1967, 95, 218–220. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Sakiguchi, T.; Kitamura, K.; Akamatsu, H.; Horio, T. Establishment of a tissue culture system for hamster sebaceous gland cells. Dermatology 1988, 197, 238–244. [Google Scholar] [CrossRef]
- Sato, T.; Imai, N.; Akimoto, N.; Sakiguchi, T.; Kitamura, K.; Ito, A. Epidermal growth factor and 1alpha,25-dihydroxyvitamin D3 suppress lipogenesis in hamster sebaceous gland cells in vitro. J. Investig. Dermatol. 2001, 117, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Millar, S.E. Molecular mechanisms regulating hair follicle development. J. Investig. Dermatol. 2002, 118, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.; Andl, T.; Bagasra, A.; Lu, M.M.; Epstein, D.J.; Morrisey, E.E.; Millar, S.E. Characterization of Wnt gene expression in developing and postnatal hair follicles and identification of Wnt5a as a target of Sonic hedgehog in hair follicle morphogenesis. Mech. Dev. 2001, 107, 69–82. [Google Scholar] [CrossRef]
- Chiang, C.; Swan, R.Z.; Grachtchouk, M.; Bolinger, M.; Litingtung, Y.; Robertson, E.K.; Cooper, M.K.; Gaffield, W.; Westphal, H.; Beachy, P.A.; et al. Essential role for Sonic hedgehog during hair follicle morphogenesis. Dev. Biol. 1999, 205, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ouji, Y.; Yoshikawa, M.; Moriya, K.; Nishiofuku, M.; Matsuda, R.; Ishizaka, S. Wnt-10b, uniquely among Wnts, promotes epithelial differentiation and shaft growth. Biochem. Biophys. Res. Commun. 2008, 367, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Van Genderen, C.; Okamura, R.M.; Fariñas, I.; Quo, R.G.; Parslow, T.G.; Bruhn, L.; Grosschedl, R. Development of several organs that require inductive epithelial-mesenchymal interactions is impaired in LEF-1-deficient mice. Genes Dev. 1994, 8, 2691–2703. [Google Scholar] [CrossRef] [PubMed]
- Goldman, C.K.; Tsai, J.C.; Soroceanu, L.; Gillespie, G.Y. Loss of vascular endothelial growth factor in human alopecia hair follicles. J. Investig. Dermatol. 1995, 104, 18S–20S. [Google Scholar] [CrossRef] [PubMed]
- Lachgar, S.; Moukadiri, H.; Jonca, F.; Charveron, M.; Bouhaddioui, N.; Gall, Y.; Bonafe, J.L.; Plouët, J. Vascular endothelial growth factor is an autocrine growth factor for hair dermal papilla cells. J. Investig. Dermatol. 1996, 106, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Lachgar, S.; Charveron, M.; Ceruti, I.; Lagarde, J.M.; Gall, Y.; Bonafe, J.L. VEGF mRNA expression in different stages of the human hair cycle, analysis by confocal laser microscopy. In Hair Research for the Next Millennium; Van Neste, D., Randall, V.A., Eds.; Elsevier Science: Amsterdam, The Netherlands, 1996; p. 407. [Google Scholar]
- Guo, L.; Degenstein, L.; Fuchs, E. Keratinocyte growth factor is required for hair development but not for wound healing. Genes Dev. 1996, 10, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Danilenko, D.M.; Ring, B.D.; Yanagihara, D.; Benson, W.; Wiemann, B.; Starnes, C.O.; Pierce, G.F. Keratinocyte growth factor is an important endogenous mediator of hair follicle growth, development, and differentiation. Normalization of the nu/nu follicular differentiation defect and amelioration of chemotherapy-induced alopecia. Am. J. Pathol. 1995, 147, 145–154. [Google Scholar] [PubMed]
- Yeung, H.C. Handbook of Chinese Herbs and Formulas; Institute of Chinese Medicine: Los Angeles, CA, USA, 1985. [Google Scholar]
- Zhang, N.N.; Park, D.K.; Park, H.J. Hair growth-promoting activity of hot water extract of Thuja orientalis. BMC Complement Altern. Med. 2013, 13, 9. [Google Scholar] [CrossRef] [PubMed]
- Park, W.S.; Lee, C.H.; Lee, B.G.; Chang, I.S. The extract of Thujae occidentalis semen inhibited 5alpha-reductase and androchronogenetic alopecia of B6CBAF1/j hybrid mouse. J. Dermatol. Sci. 2003, 31, 91–98. [Google Scholar] [CrossRef]
- Jain, R.K.; Garg, S.C. Antimicrobial activity of the essential oil of Thuja orientalis L. Ancient. Sci. Life 1997, 16, 186–189. [Google Scholar]
- Hassanzadeh, M.K.; Rahimizadeh, M.; Fazly Bazzaz, B.S.; Emami, S.A.; Asili, J. Chemical and antimicrobial studies of Platycladus orientalis essential oils. Pharm. Biol. 2001, 5, 388–390. [Google Scholar] [CrossRef]
- Duhan, J.S.; Saharan, P.; Surekha; Kumar, A. Antimicrobial potential of various fractions of Thuja orientalis. Afr. J. Microbiol. Res. 2013, 7, 3179–3186. [Google Scholar]
- Jasuja, N.D.; Sharma, S.K.; Saxena, R.; Choudhary, J.; Sharma, R.; Joshi, S.C. Antibacterial, antioxidant and phytochemical investigation of Thuja orientalis leaves. J. Med. Plants Res. 2013, 7, 1886–1893. [Google Scholar]
- Choi, Y.; Moon, A.; Kim, Y.C. A pinusolide derivative, 15-methoxypinusolidic acid from Biota orientalis inhibits inducible nitric oxide synthase in microglial cells, implication for a potential anti-inflammatory effect. Int. Immunopharmacol. 2008, 8, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.X.; Wang, Y.; Kong, L.D.; Yang, C.; Zhang, X. Effects of Biota orientalis extract and its flavonoid constituents, quercetin and rutin on serum uric acid levels in oxonate-induced mice and xanthine dehydrogenase and xanthine oxidase activities in mouse liver. J. Ethnopharmacol. 2004, 93, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liu, Z.; Wang, Z.; Wei, D. Quality evaluation of Platycladus orientalis (L.) Franco through simultaneous determination of four bioactive flavonoids by high-performance liquid chromatography. J. Pharm. Biomed. Anal. 2006, 4, 1186–1190. [Google Scholar] [CrossRef] [PubMed]
- Duhan, J.S.; Saharan, P.; Gahlawat, S.K. Antioxidant potential of various extracts of stem of Thuja orientalis, in vitro study. Intern. J. App. Bio Pharma. Technol. 2013, 3, 264–271. [Google Scholar]
- Zhang, Y.; Xing, Y.; Guo, H.; Ma, X.; Li, Y. Immunohistochemical study of hair follicle stem cells in regenerated hair follicles induced by Wnt10b. Int. J. Med. Sci. 2016, 13, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Takebe, K.; Oka, Y.; Radisky, D.; Tsuda, H.; Tochigui, K.; Koshida, S.; Kogo, K.; Hirai, Y. Epimorphin acts to induce hair follicle anagen in C57BL/6 mice. FASEB J. 2003, 17, 2037–2047. [Google Scholar] [CrossRef] [PubMed]
- Lo Celso, C.; Prowse, D.M.; Watt, F.M. Transient activation of beta-catenin signalling in adult mouse epidermis is sufficient to induce new hair follicles but continuous activation is required to maintain hair follicle tumours. Development 2004, 131, 1787–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Marubayashi, A.; Nakaya, Y.; Fukui, K.; Arase, S. Dermal papilla cells possess sulfonylurea receptor 2B and adenosine receptors which are the possible mediators of minoxidil-induced VEGF productions. J. Investig. Dermatol. 2001, 117, 1594–1600. [Google Scholar] [PubMed]
- Yano, K.; Brown, L.F.; Detmar, M. Control of hair growth and follicle size by VEGF-mediated angiogenesis. J. Clin. Investig. 2001, 107, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Sheets, L. Excessive activation of ionotropic glutamate receptors induces apoptotic hair-cell death independent of afferent and efferent innervation. Sci. Rep. 2017, 7, 41102. [Google Scholar] [CrossRef] [PubMed]
- Koo, K.A.; Kim, S.H.; Lee, M.K.; Kim, Y.C. 15-Methoxypinusolidic acid from Biota orientalis attenuates glutamate-induced neurotoxicity in primary cultured rat cortical cells. Toxicol. In Vitro 2006, 20, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Bown, D. Encyclopaedia of Herbs and Their Uses; Dorling Kindersley: London, UK, 1995. [Google Scholar]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, H.; Gu, L.; Maeda, K. Evaluation of the Effect of Plant Mixture Ethanol Extracts Containing Biota orientalis L. Extract on Suppression of Sebum in Cultured Sebocytes and on Stimulation of Growth of Keratinocytes Co-cultured with Hair Papilla Cells. Cosmetics 2017, 4, 29. https://doi.org/10.3390/cosmetics4030029
Zeng H, Gu L, Maeda K. Evaluation of the Effect of Plant Mixture Ethanol Extracts Containing Biota orientalis L. Extract on Suppression of Sebum in Cultured Sebocytes and on Stimulation of Growth of Keratinocytes Co-cultured with Hair Papilla Cells. Cosmetics. 2017; 4(3):29. https://doi.org/10.3390/cosmetics4030029
Chicago/Turabian StyleZeng, Haifeng, Lihao Gu, and Kazuhisa Maeda. 2017. "Evaluation of the Effect of Plant Mixture Ethanol Extracts Containing Biota orientalis L. Extract on Suppression of Sebum in Cultured Sebocytes and on Stimulation of Growth of Keratinocytes Co-cultured with Hair Papilla Cells" Cosmetics 4, no. 3: 29. https://doi.org/10.3390/cosmetics4030029