
Nanoscale Electric Characteristics and Oriented Assembly of Halobacterium salinarum Membrane Revealed by Electric Force Microscopy

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
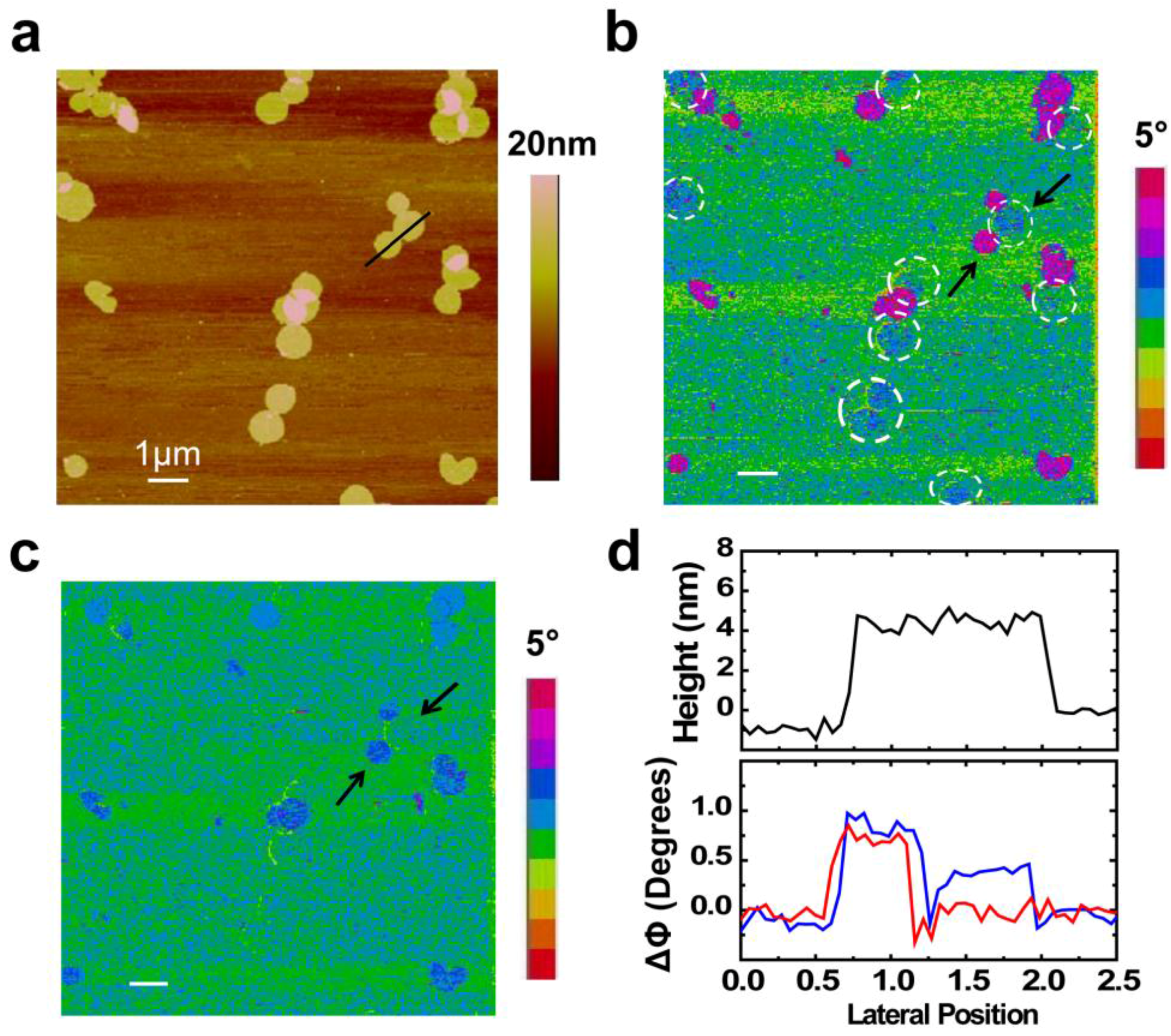
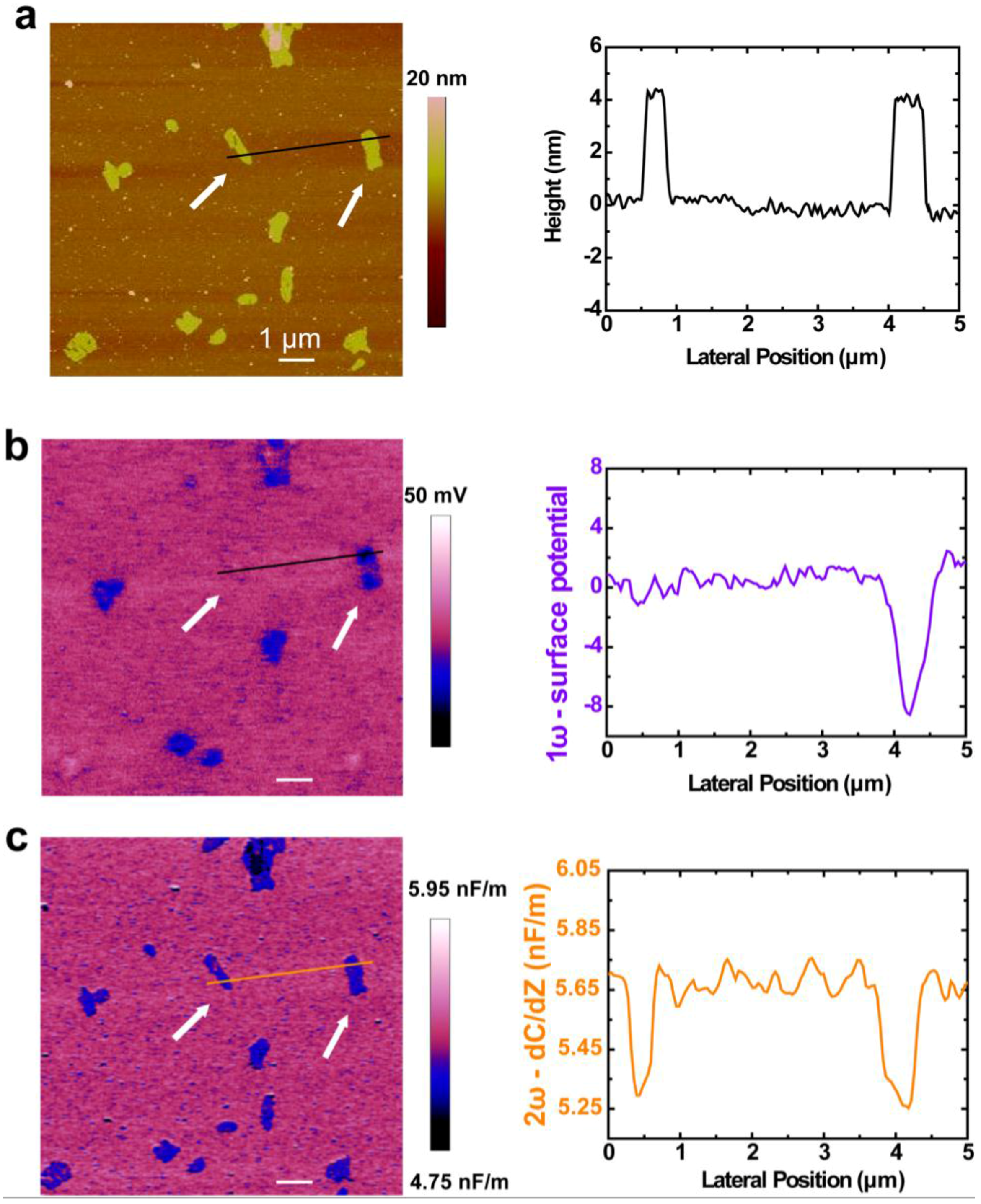
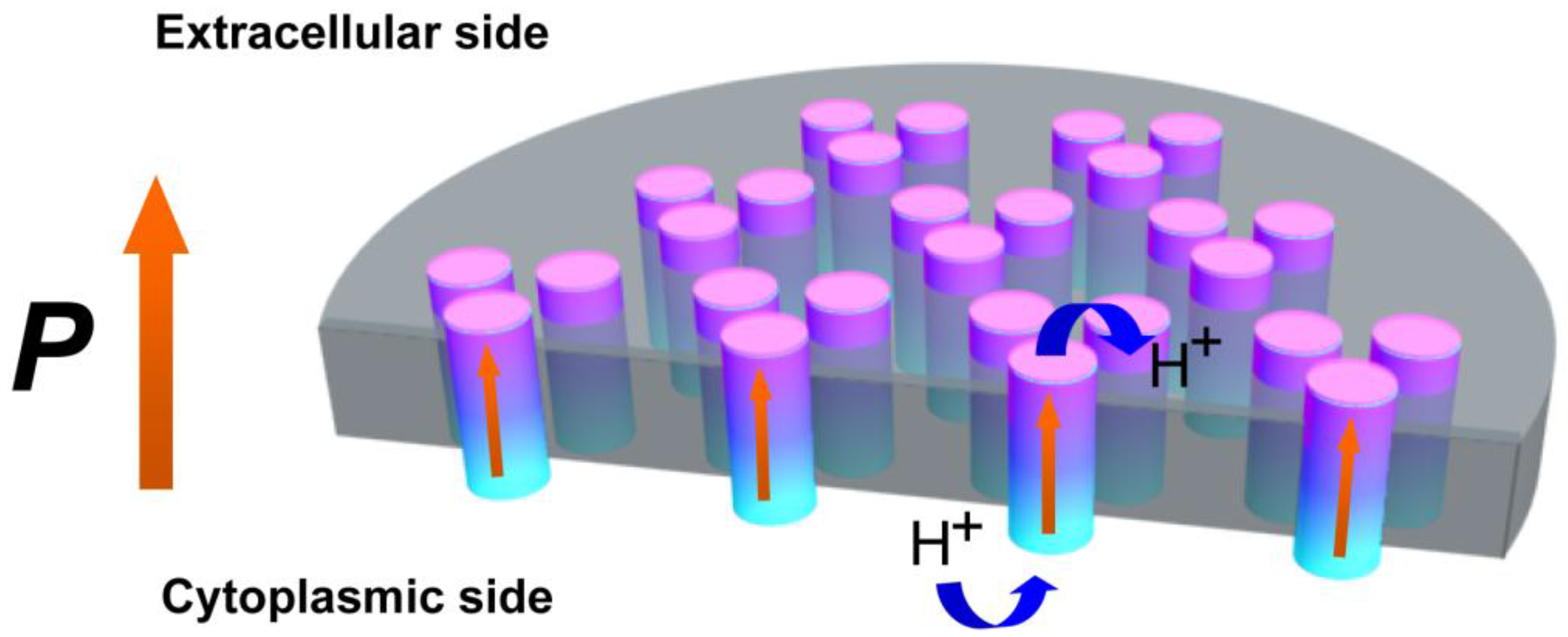
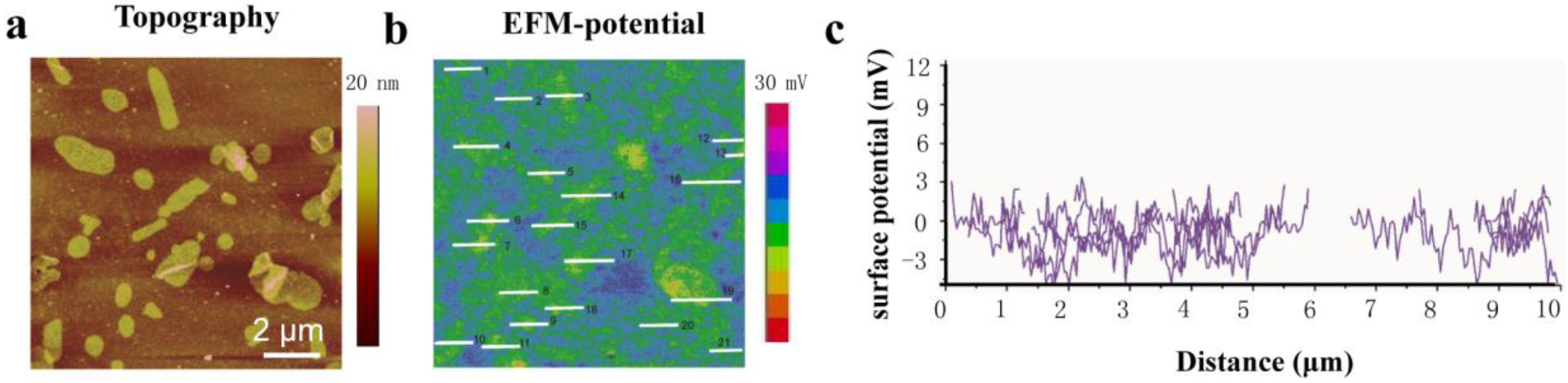
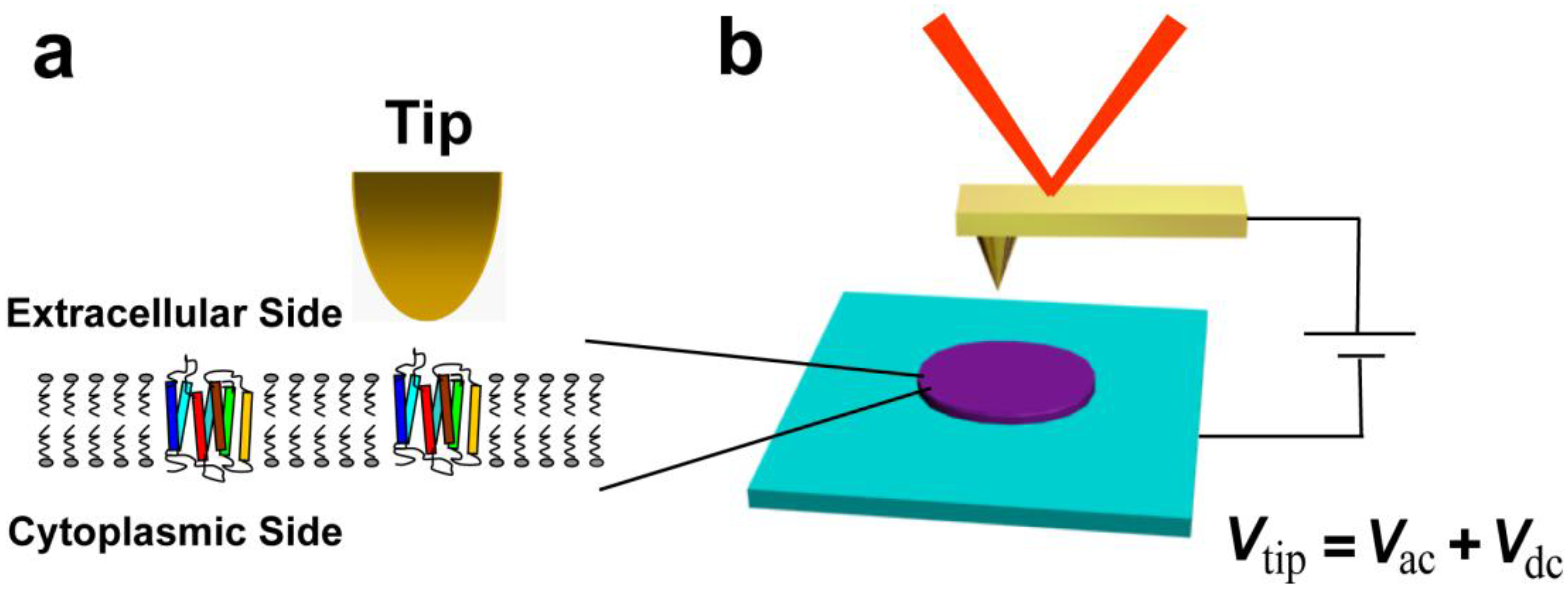
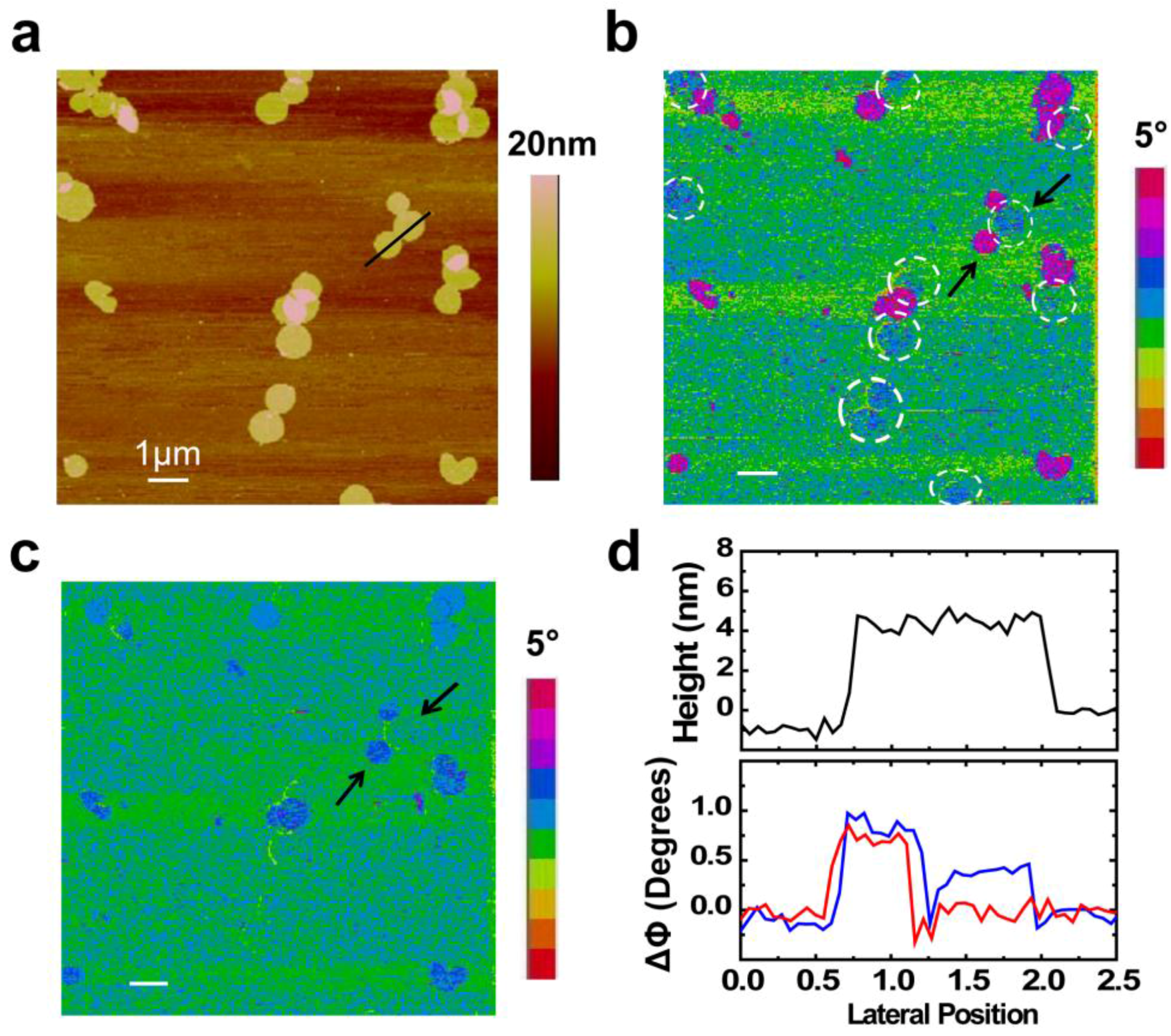
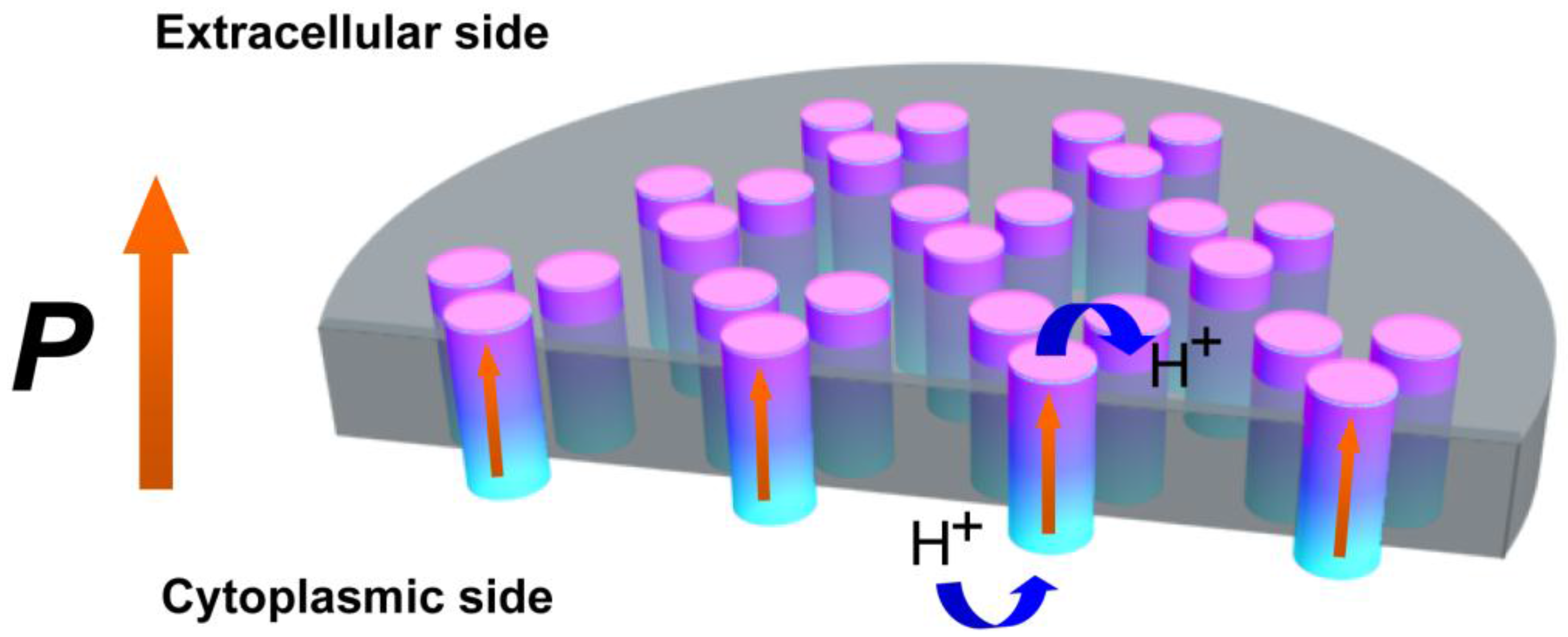
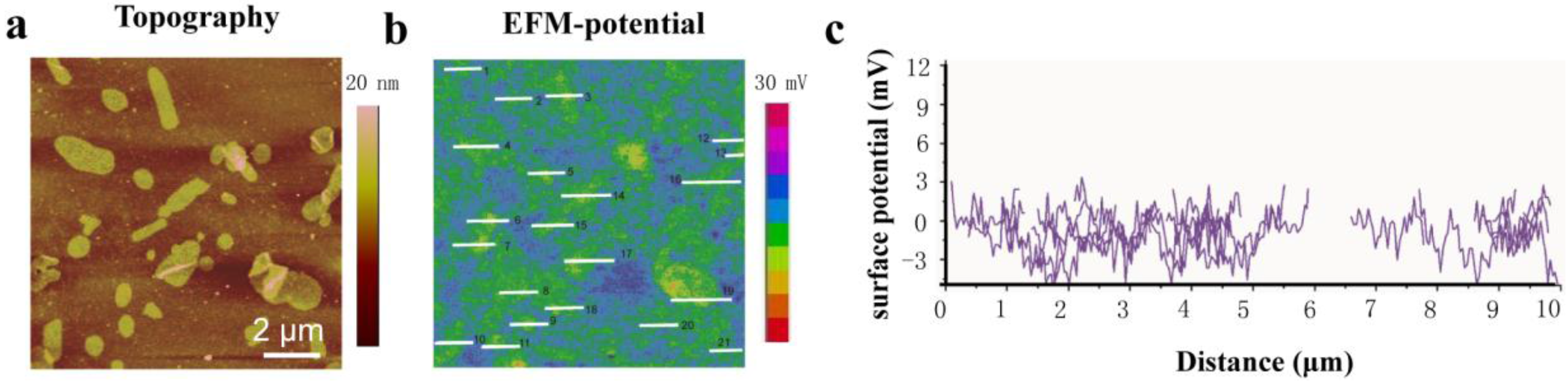
2. Results and Discussion
3. Materials and Methods
3.1. Materials
3.2. Nano-Characterization of Purple Membranes with Electric Force Microscopy Measurements
3.3. Electric Potential and Dielectric Properties Measurements
3.4. Surface Plasma Resonance (SPR) Experiments
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hampp, N. Bacteriorhodopsin as a photochromic retinal protein for optical memories. Chem. Rev. 2000, 100, 1755–1776. [Google Scholar] [CrossRef] [PubMed]
- Haupts, U.; Tittor, J.; Oesterhelt, D. Closing in on bacteriorhodopsin: Progress in understanding the molecule. Annu. Rev. Biophys. Biomol. Struct. 1999, 28, 367–399. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, S.; Henderson, R. Molecular mechanism of vectorial proton translocation by bacteriorhodopsin. Nature 2000, 406, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Lanyi, J.K. Bacteriorhodopsin. Annu. Rev. Physiol. 2004, 66, 665–688. [Google Scholar] [CrossRef] [PubMed]
- Oesterhelt, D. The structure and mechanism of the family of retinal proteins from halophilic archaea. Curr. Opin. Struc. Boil. 1998, 8, 489–500. [Google Scholar] [CrossRef]
- Korenstein, R.; Hess, B. Hydration effects on photocycle of bacteriorhodopsin in thin layers of purple membrane. Nature 1977, 270, 184–186. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Vassylyev, D.G.; Miyazawa, A.; Kidera, A.; Matsushima, M.; Mitsuoka, K. Surface of bacteriorhodopsin revealed by high-resolution electron crystallography. Nature 1997, 389, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Alexiev, U.; Mollaaghababa, R.; Scherrer, P.; Khoranat, H.G.; Heyn, M.P. Rapid long-range proton diffusion along the surface of the purple membrane and delayed proton transfer into the bulk. Proc. Natl. Acad. Sci. USA 1995, 92, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Butt, H.J. Measuring local surface charge densities in electrolyte solutions with a scanning force microscope. Biophys. J. 1992, 63, 578–582. [Google Scholar] [CrossRef]
- Müller, D.J.; Engel, A. The height of biomolecules measured with the atomic force microscope depends on electrostatic interactions. Biophys. J. 1997, 73, 1633–1644. [Google Scholar] [CrossRef]
- Contera, S.A.; Voïtchovskyx, K.; Ryan, J.F. Controlled ionic condensation at the surface of a native extremophile membrane. Nanoscale 2010, 2, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Mogi, T.; Stern, L.J.; Marti, T.; Chao, B.H.; Khorana, H.G. Aspartic acid substitutions affect protontranslocation by bacteriorhodopsin. Proc. Natl. Acad. Sci. USA 1988, 85, 4148–4152. [Google Scholar] [CrossRef] [PubMed]
- Hsu, K.C.; Rayfield, G.W.; Needleman, R. Reversal of the surface charge asymmetry in purple membrane due to single amino acid substitutions. Biophys. J. 1996, 70, 2358–2365. [Google Scholar] [CrossRef]
- Qi, G.C.; Yang, Y.L.; Yan, H.; Guan, L.; Li, Y.B.; Qiu, X.H.; Wang, C. Quantifying surface charge density by using an electric force microscope with a referential structure. J. Phys. Chem. C 2009, 113, 204–207. [Google Scholar] [CrossRef]
- Girard, P. Electrostatic force microscopy: Principles and some applications to semiconductors. Nanotechnology 2001, 12, 485–490. [Google Scholar] [CrossRef]
- Yan, L.; Punckt, C.; Aksay, I.A.; Mertin, W.; Bacher, G. Local voltage drop in a single functionalized graphene sheet characterized by Kelvin probe force microscopy. Nano Lett. 2011, 11, 3543–3549. [Google Scholar] [CrossRef] [PubMed]
- Liam, S.C.; Obadiah, G.R.; David, S.G. Electrical scanning probe microscopy on active organic electronic devices. Adv. Mater. 2009, 21, 19–28. [Google Scholar]
- Li, D.; Yan, H.; Li, C.; Yang, Y.; Wei, Z.; Wang, C. Nanoscale structural and electronic evolution for increased efficiency in polymer solar cells monitored by electric scanning probe microscopy. Chin. Sci. Bull. 2014, 59, 360–368. [Google Scholar] [CrossRef]
- Yan, H.; Li, D.; Li, C.; Lu, K.; Zhang, Y.; Wei, Z.; Yang, Y.; Wang, C. Bridging mesoscopic blend structure and property to macroscopic device performance via in situ optoelectronic characterization. J. Mater. Chem. 2012, 22, 4349–4355. [Google Scholar] [CrossRef]
- Ron, I.; Pecht, I.; Sheves, M.; Cahen, D. Proteins as solid-state electronic conductors. Acc. Chem. Res. 2010, 43, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Frolov, L.; Rosenwaks, Y.; Carmeli, C.; Carmeli, I. Fabrication of a photoelectronic device by direct chemical binding of the photosynthetic reaction center protein to metal surfaces. Adv. Mater. 2005, 17, 2434–2437. [Google Scholar] [CrossRef]
- Du, H.; Li, D.; Wang, Y.; Wang, C.; Zhang, D.; Yang, Y.; Wang, C. Determination of the surface charge density and temperature dependence of purple membrane by electric force microscopy. J. Phys. Chem. B 2013, 117, 9895–9899. [Google Scholar] [CrossRef] [PubMed]
- Knapp, H.F.; Mesquida, P.; Stemmer, A. Imaging the surface potential of active purple membrane. Surf. Interface Anal. 2002, 33, 108–112. [Google Scholar] [CrossRef]
- Engelman, D.M. Membranes are more mosaic than fluid. Nature 2005, 438, 578–580. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, K.; Sheets, E.D.; Simson, R. Revisiting the fluid mosaic model of membranes. Science 1995, 268, 1441–1442. [Google Scholar] [CrossRef] [PubMed]
- Philippsen, A.; Im, W.; Engel, A.; Schirmer, T.; Roux, B.; Müller, D.J. Imaging the electrostatic potential of transmembrane channels: Atomic probe microscopy of OmpF porin. Biophys. J. 2002, 82, 1667–1676. [Google Scholar] [CrossRef]
- Pfreundschuh, M.; Hensen, U.; Müller, D.J. Quantitative imaging of the electrostatic field and potential generated by a transmembrane protein pore at subnanometer resolution. Nano Lett. 2013, 13, 5585–5593. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Honig, T.; Ron, I.; Friedman, N.; Sheves, M.; Cahen, D. Bacteriorhodopsin as an electronic conduction medium for biomolecular electronics. Chem. Soc. Rev. 2008, 37, 2422–2432. [Google Scholar] [CrossRef] [PubMed]
- Tunuguntla, R.H.; Bangar, M.A.; Kim, K.; Stroeve, P.; Grigoropoulos, C.; Ajo-Franklin, C.M.; Noy, A. Bioelectronic light-gated transistors with biologically tunable performance. Adv. Mater. 2015, 27, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, G.; Magliulo, M.; Mallardi, A.; Angione, M.D.; Gobeljic, D.; Scamarcio, G.; Fratini, E.; Ridi, F.; Torsi, L. Electronic transduction of proton translocations in nanoassembled lamellae of bacteriorhodopsin. ACS Nano 2014, 8, 7834–7845. [Google Scholar] [CrossRef] [PubMed]
- Staii, C.; Johnson, A.T.; Pinto, N.J. Quantitative analysis of scanning conductance microscopy. Nano Lett. 2004, 4, 859–862. [Google Scholar] [CrossRef]
- Bockrath, M.; Markovic, N.; Shepard, A.; Tinkham, M. Scanned conductance microscopy of carbon nanotubes and lambda-DNA. Nano Lett. 2002, 2, 187–190. [Google Scholar] [CrossRef]
- Wang, S.; Wang, R.; Wang, X.; Zhang, D.; Qiu, X. Nanoscale charge distribution and energy band modification in defect-patterned grapheme. Nanoscale 2012, 4, 2651–2657. [Google Scholar] [CrossRef] [PubMed]
- Liscio, A.; Palermo, V.; Samorì, P. Nanoscale quantitative measurement of the potential of charged nanostructures by electrostatic and Kelvin probe force microscopy: Unraveling electronic processes in complex materials. Acc. Chem. Res. 2010, 43, 541–550. [Google Scholar] [CrossRef] [PubMed]
- García, R.; Pérez, R. Dynamic atomic force microscopy methods. Surf. Sci. Rep. 2002, 47, 197–301. [Google Scholar] [CrossRef]
- Baumann, R.P.; Busch, A.P.; Heidel, B.; Hampp, N. A new class of purple membrane variants for the construction of highly oriented membrane assemblies on the basis of noncovalent interactions. J. Phys. Chem. B 2012, 116, 4134–4140. [Google Scholar] [CrossRef] [PubMed]
- Ermolina, I.; Lewis, A.; Feldman, Y. Dielectric properties of the bR membrane. J. Phys. Chem. B 2003, 107, 14537–14544. [Google Scholar] [CrossRef]
- Gemici, Z.; Schwachulla, P.I.; Williamson, E.H.; Rubner, M.F.; Cohen, R.E. Targeted functionalization of nanoparticle thin films via capillary condensation. Nano Lett. 2009, 9, 1064–1068. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, L.; Ferrari, G.; Sampietro, M.; Gomila, G. Quantitative nanoscale dielectric microscopy of single-layer supported biomembranes. Nano Lett. 2009, 9, 1604–1608. [Google Scholar] [CrossRef] [PubMed]
- Koyama, K.; Yamaguchi, N.; Miyasaka, T. Antibody-mediated bacteriorhodopsin orientation for molecular device architec-tures. Science 1994, 265, 762–765. [Google Scholar] [CrossRef] [PubMed]
- Panzer, M.J.; Aidala, K.E.; Anikeeva, P.O.; Halpert, J.E.; Bawendi, M.G.; Bulović, V. Nanoscale morphology revealed at the interface between colloidal quantum dots and organic semiconductor films. Nano Lett. 2010, 10, 2421–2426. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Majima, T.; Miyake, J.; Ajiki, S.; Sugino, H.; Toyotama, H.; Kawamura, S. Oriented immobilization of bacterial photosynthetic membrane. Appl. Microbiol. Biotechnot. 1990, 32, 544–549. [Google Scholar] [CrossRef]
- Fisher, K.A.; Yanagimoto, K.; Stoeckentus, W. Oriented adsorption of purple membrane to cationic surfaces. J. Cell Biol. 1978, 77, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Re, F.; Airoldi, C.; Zona, C. Beta amyloid aggregation inhibitors: Small molecules as candidate drugs for therapy of alzheimers disease. Curr. Med. Chem. 2010, 17, 2990–3006. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Li, D.; Wang, Y.; Wang, C.; Li, P.; Yang, Y.; Wang, C. Peptide-assisted directional adsorption of purple membrane at the liquid-solid interface. J. Phys. Chem. C 2014, 118, 29770–29776. [Google Scholar] [CrossRef]
- Adamczyk, M.; Moore, J.A.; Yu, Z. Application of surface plasmon resonance toward studies of low-molecular-weight an-tigen-antibody binding interactions. Methods 2000, 20, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Malesevic, M.; Jahreis, G. The architecture of protein-ligand binding sites revealed through template-assisted in-tramolecular peptide-peptide interactions. Angew. Chem. Int. Ed. 2005, 44, 1408–1412. [Google Scholar] [CrossRef] [PubMed]
- Culha, M.; Lavrik, N.V.; Schell, F.M. Characterization of volatile, hydrophobic cyclodextrin derivatives as thin films for sensor applications. Sens. Actuators B Chem. 2003, 92, 171–180. [Google Scholar] [CrossRef]
- Oesterhelt, D.; Stoeckenius, W. Isolation of the cell membrane of Halobacterium halobium and its fractionation into red and purple membrane. Methods Enzymol. 1974, 31, 667–678. [Google Scholar] [PubMed]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Wang, Y.; Du, H.; Xu, S.; Li, Z.; Yang, Y.; Wang, C. Nanoscale Electric Characteristics and Oriented Assembly of Halobacterium salinarum Membrane Revealed by Electric Force Microscopy. Nanomaterials 2016, 6, 197. https://doi.org/10.3390/nano6110197
Li D, Wang Y, Du H, Xu S, Li Z, Yang Y, Wang C. Nanoscale Electric Characteristics and Oriented Assembly of Halobacterium salinarum Membrane Revealed by Electric Force Microscopy. Nanomaterials. 2016; 6(11):197. https://doi.org/10.3390/nano6110197
Chicago/Turabian StyleLi, Denghua, Yibing Wang, Huiwen Du, Shiwei Xu, Zhemin Li, Yanlian Yang, and Chen Wang. 2016. "Nanoscale Electric Characteristics and Oriented Assembly of Halobacterium salinarum Membrane Revealed by Electric Force Microscopy" Nanomaterials 6, no. 11: 197. https://doi.org/10.3390/nano6110197