
Chemokine-Releasing Nanoparticles for Manipulation of the Lymph Node Microenvironment

Abstract
:1. Introduction
2. Results
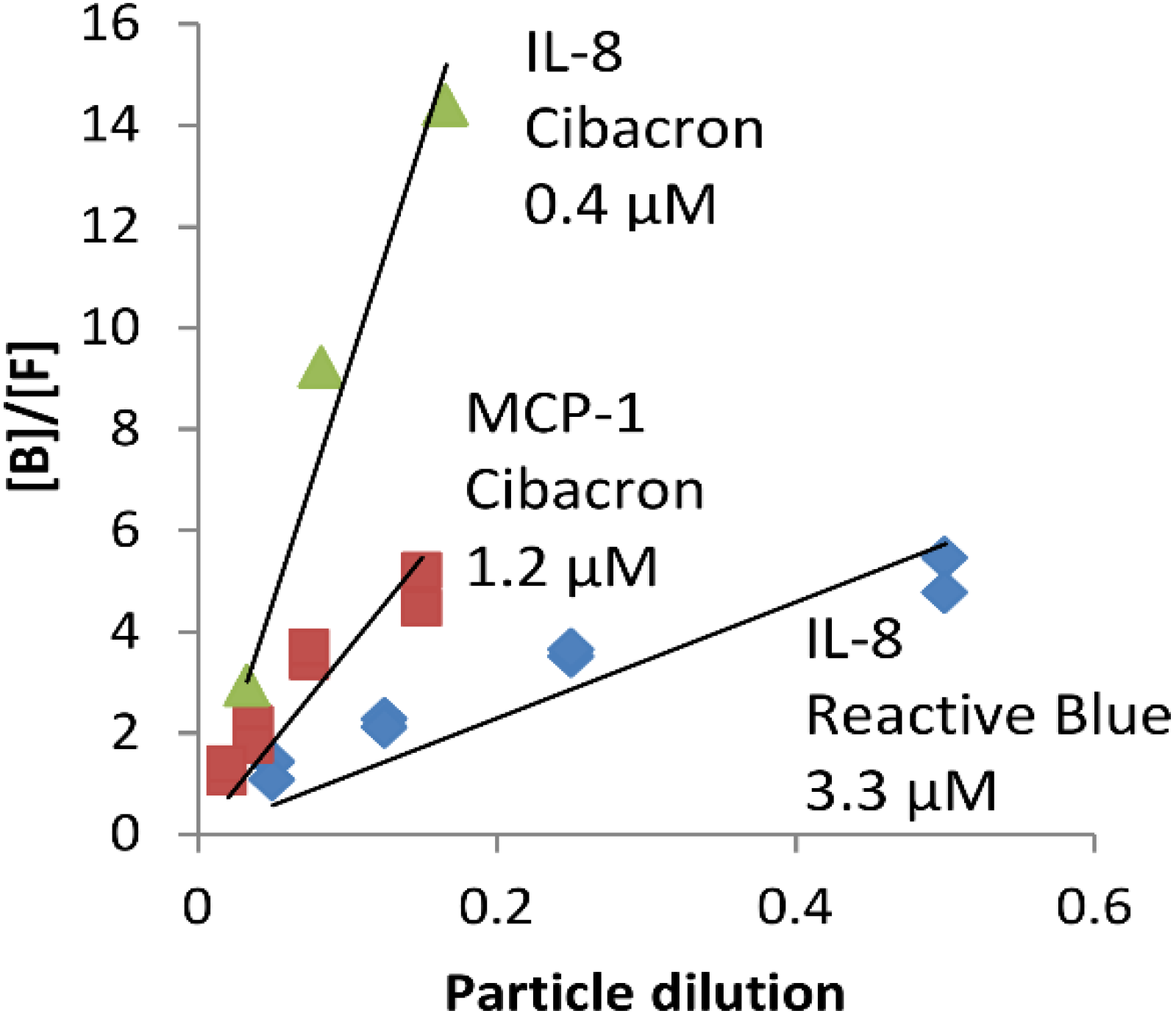
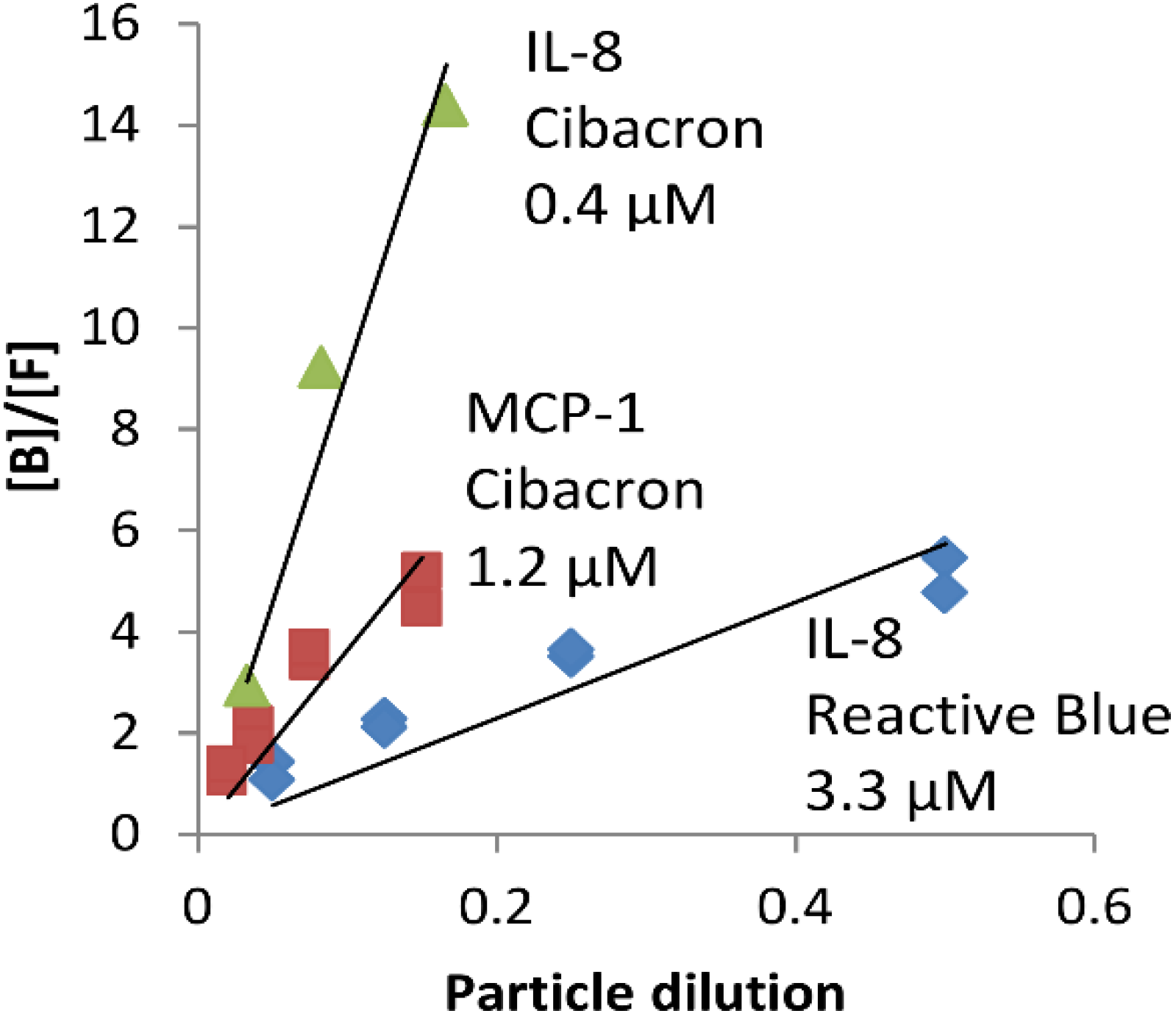
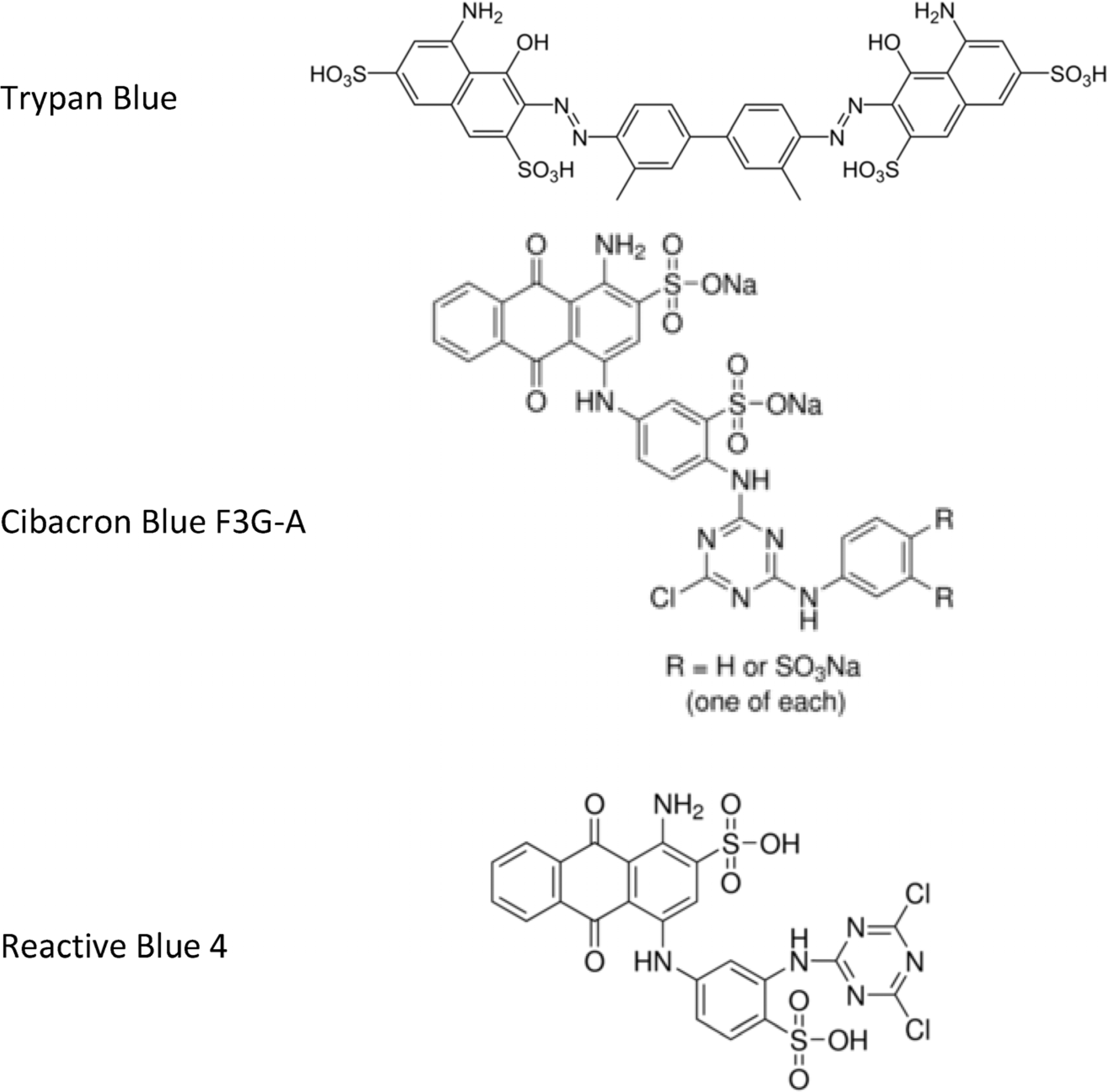
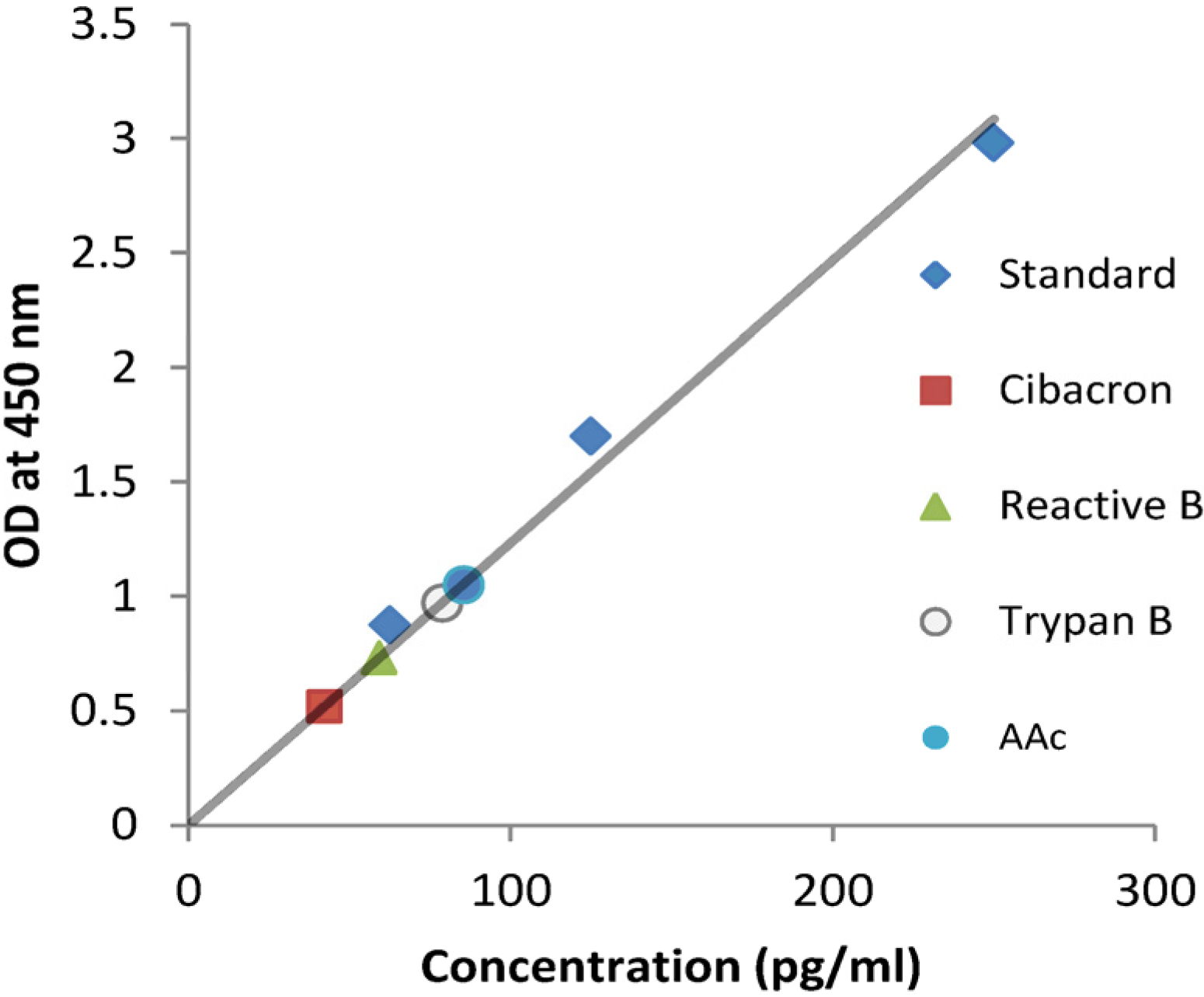
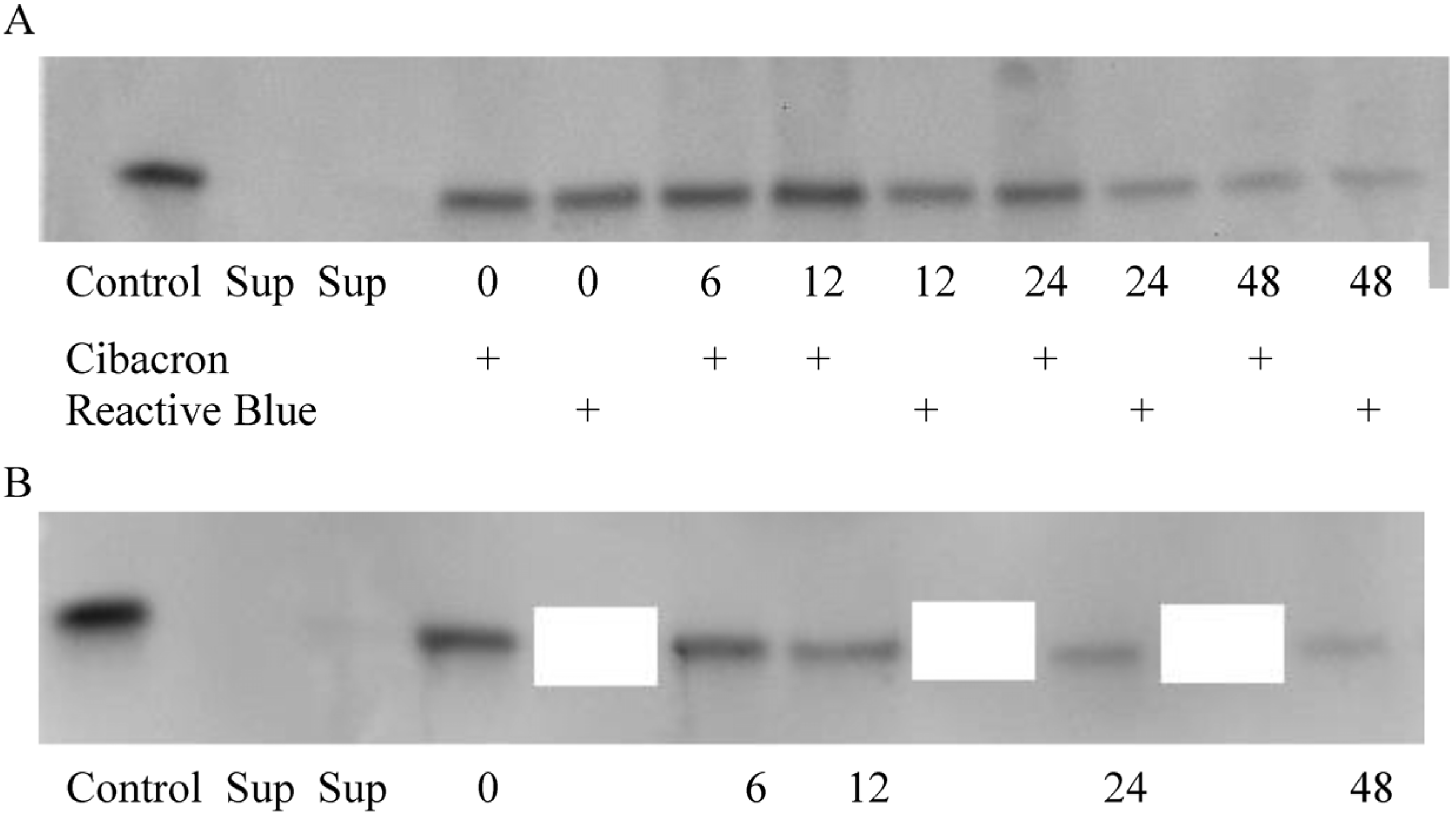
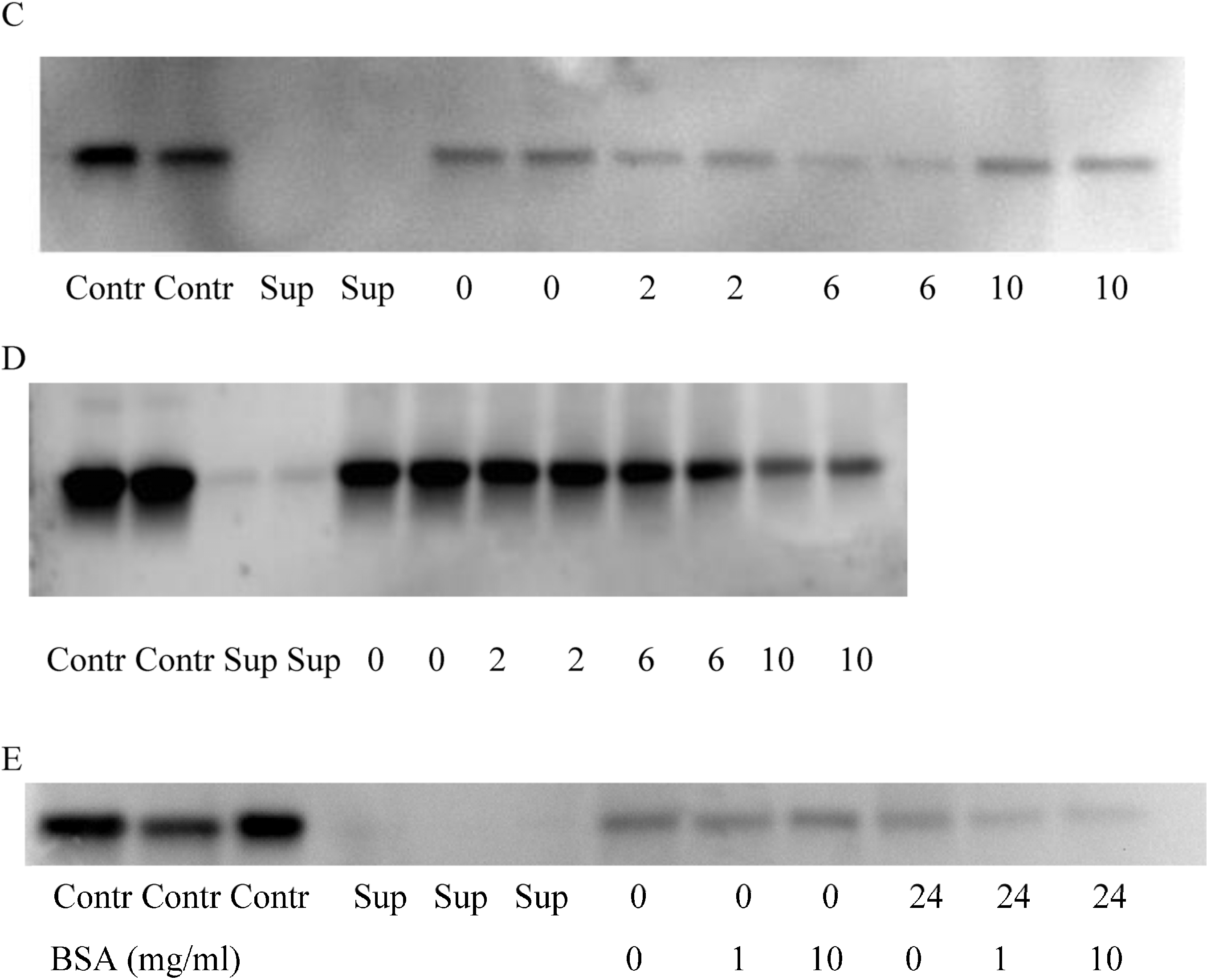
2.1. NPs Containing Different Chemical Baits Can Be Loaded with CKs
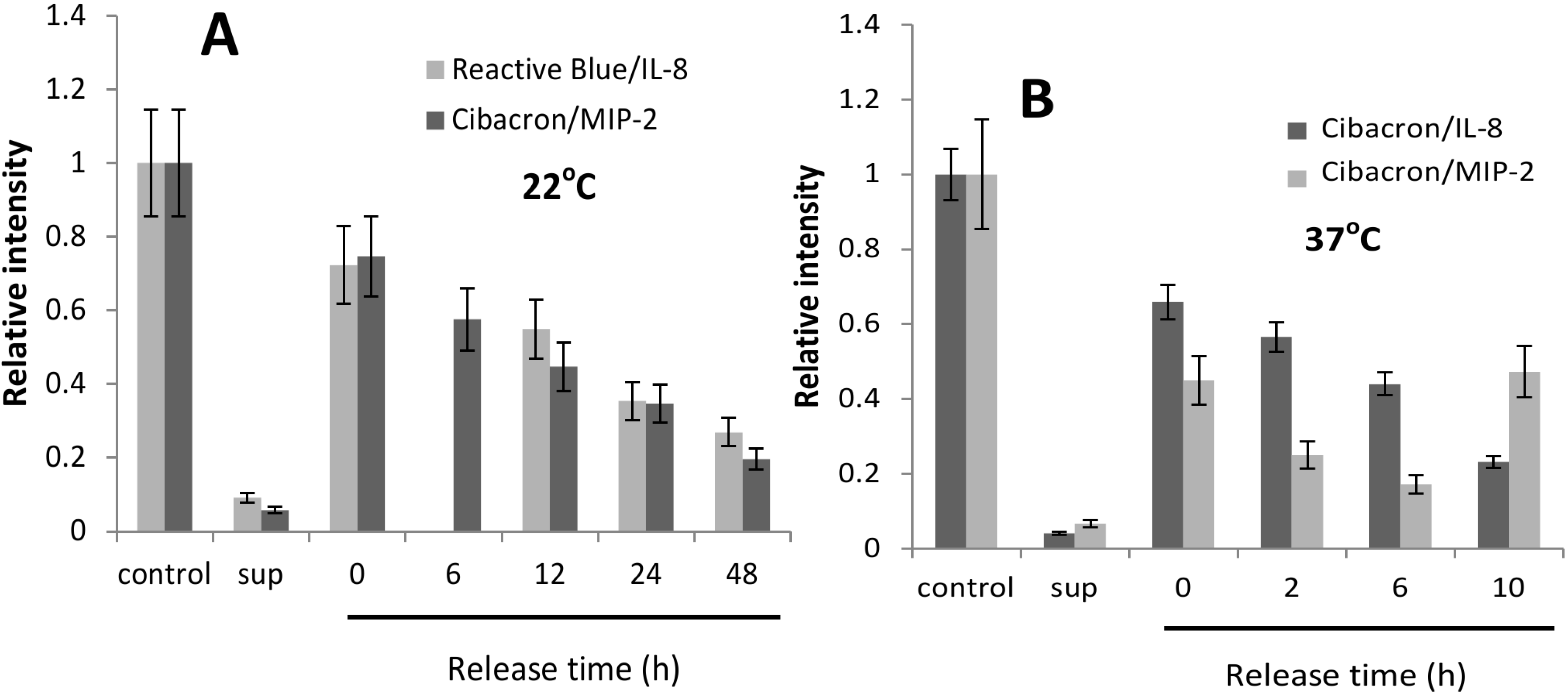
2.2. CK-Loaded NPs Provide a Sustained Release of Their Cargo
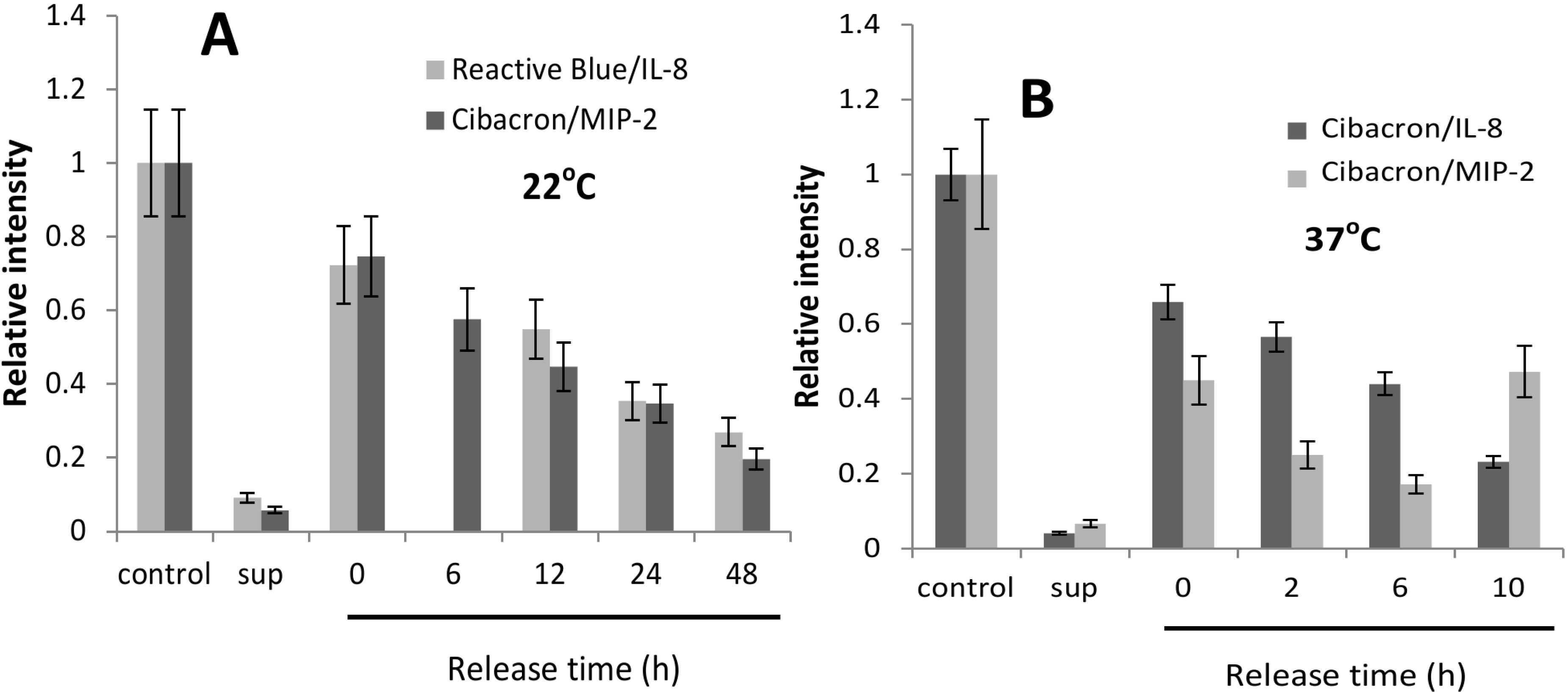

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bait | 22 °C | 37 °C | ||
|---|---|---|---|---|
| IL-8 | MIP-2 | IL-8 | MIP-2 | |
| CB | kd 0.016 ± 0.005 * h−1 t1/2 43.5 h | kd 0.030 ± 0.002 * h−1 t1/2 23.7 h | kd 0.094 ± 0.017 * h−1 t1/2 7.44 h (BSA 1 mg/mL) *** | kd 0.17 ± 0.05 * h−1 t1/2 4.1 h (BSA 1 mg/mL) *** |
| RB | kd 0.022 ± 0.004 * h−1 t1/2 31.3 h | ND ** | ND ** | ND ** |
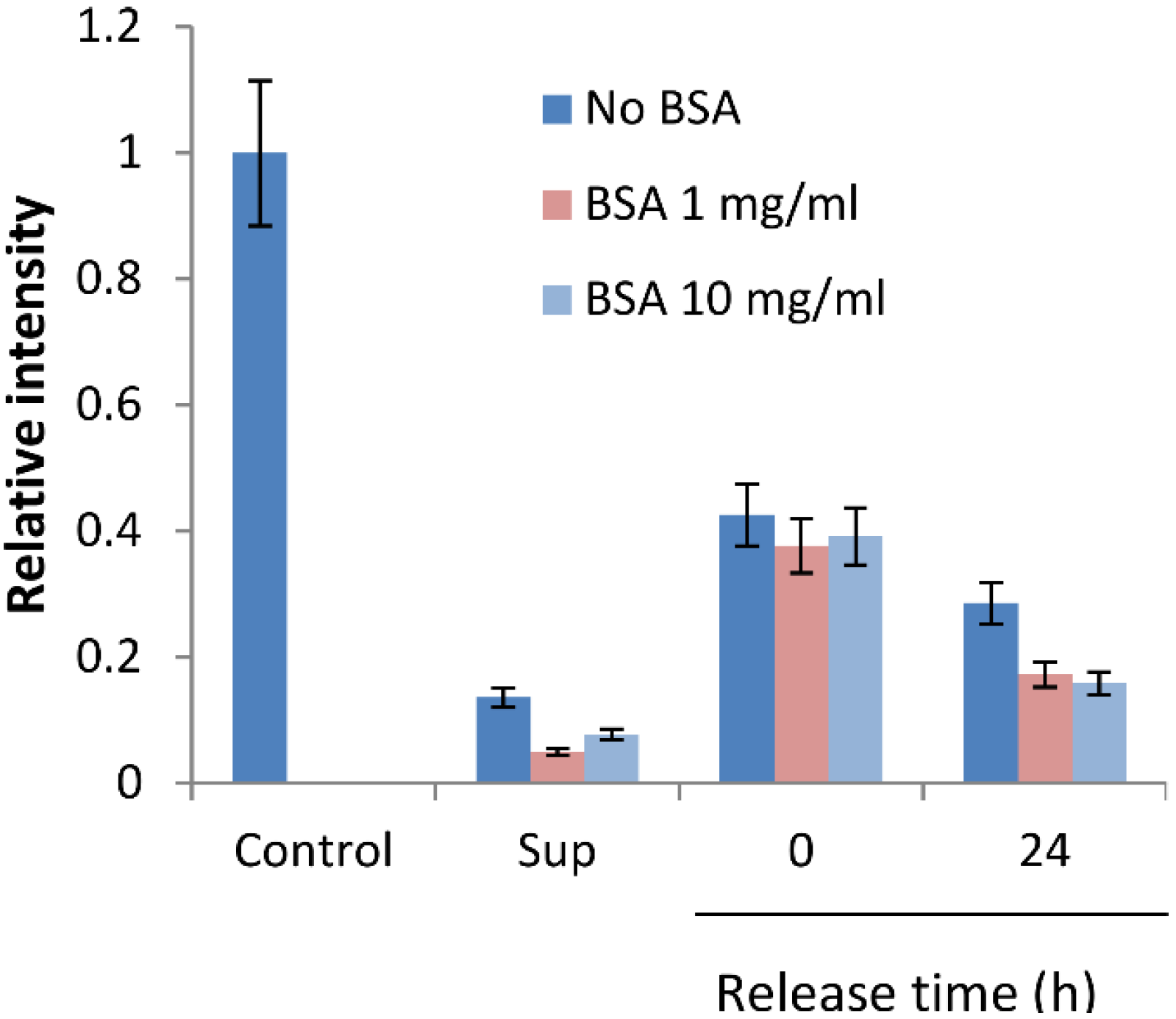
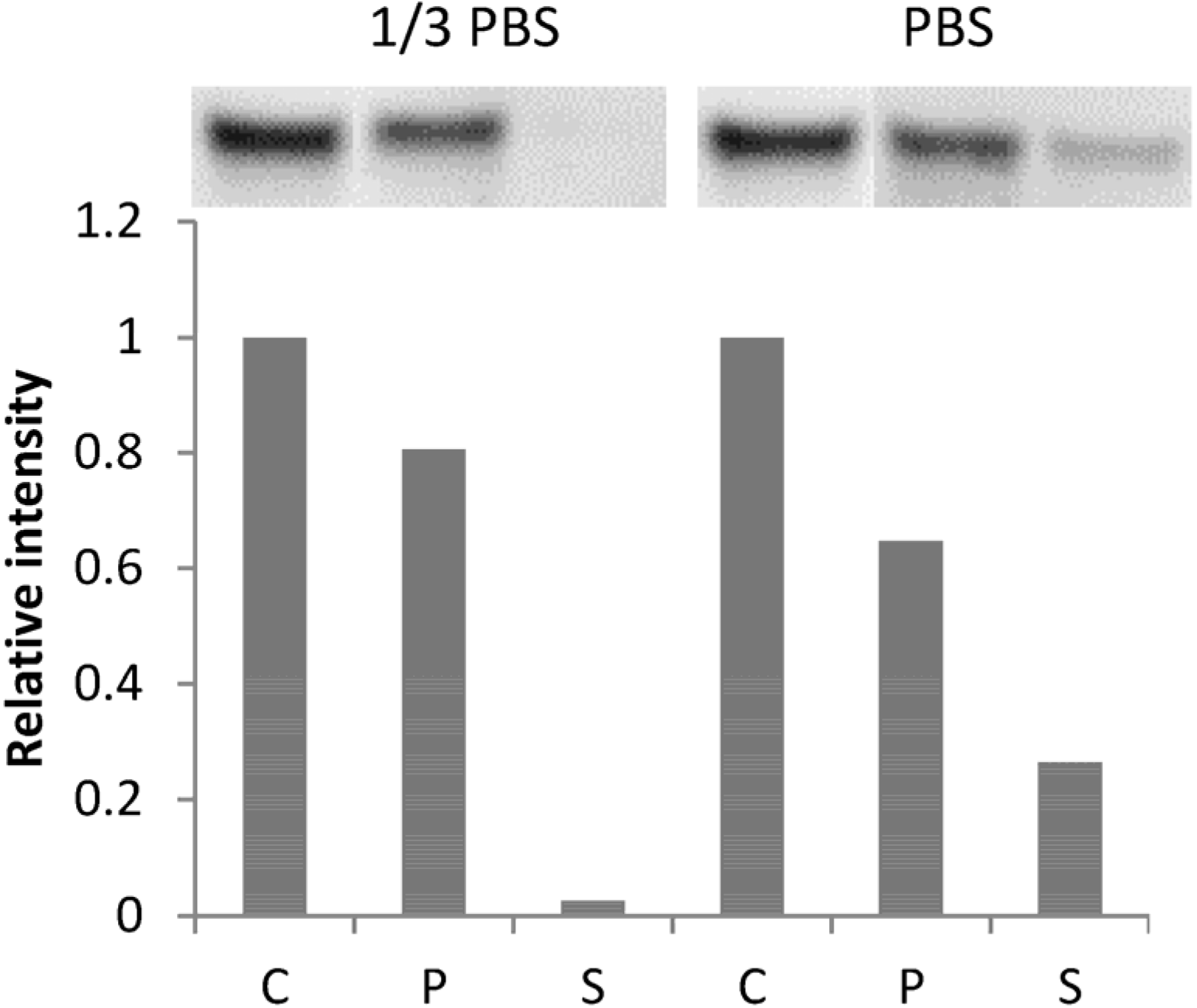
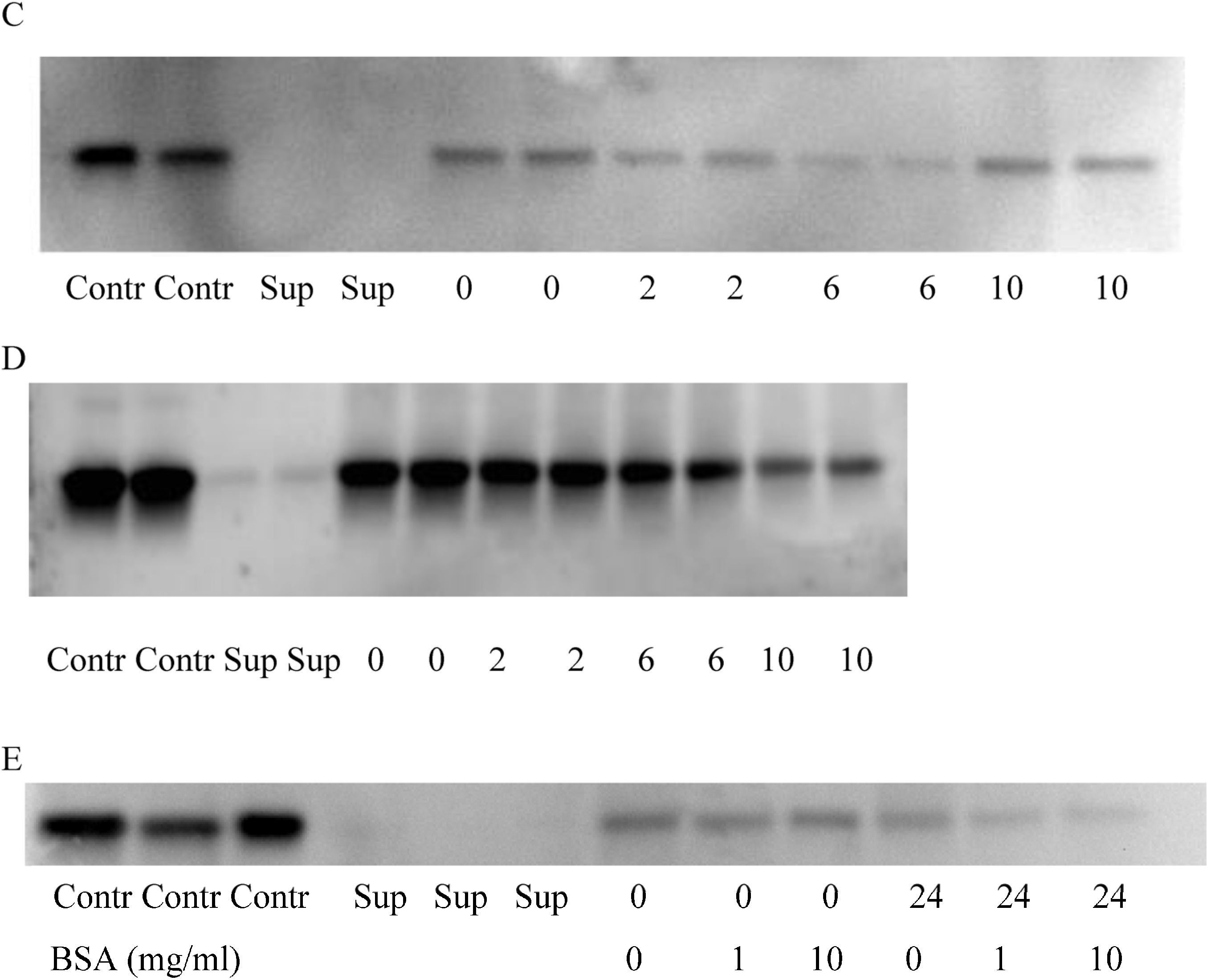
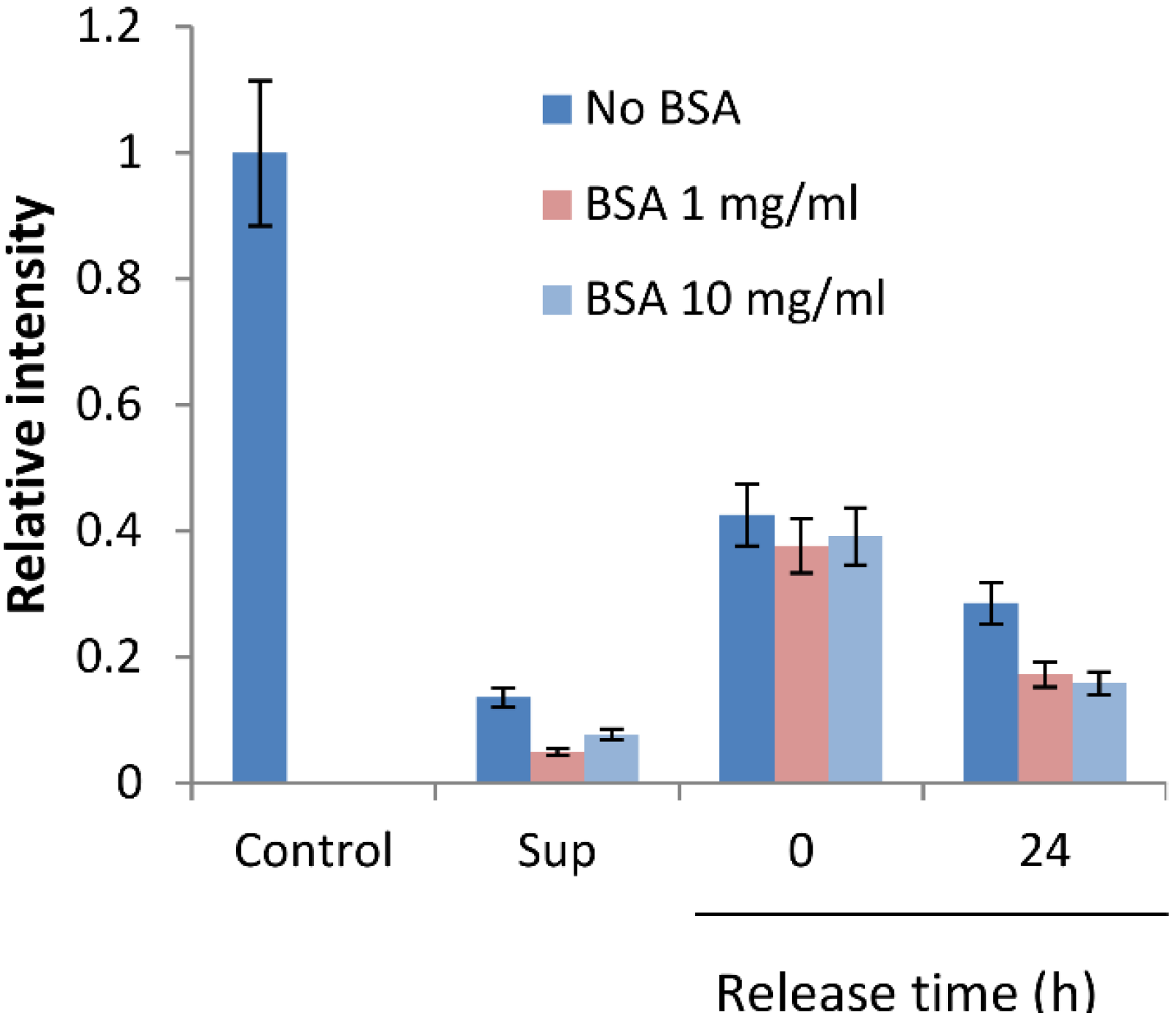
2.3. BSA Does Not Interfere with CK Loading and Release
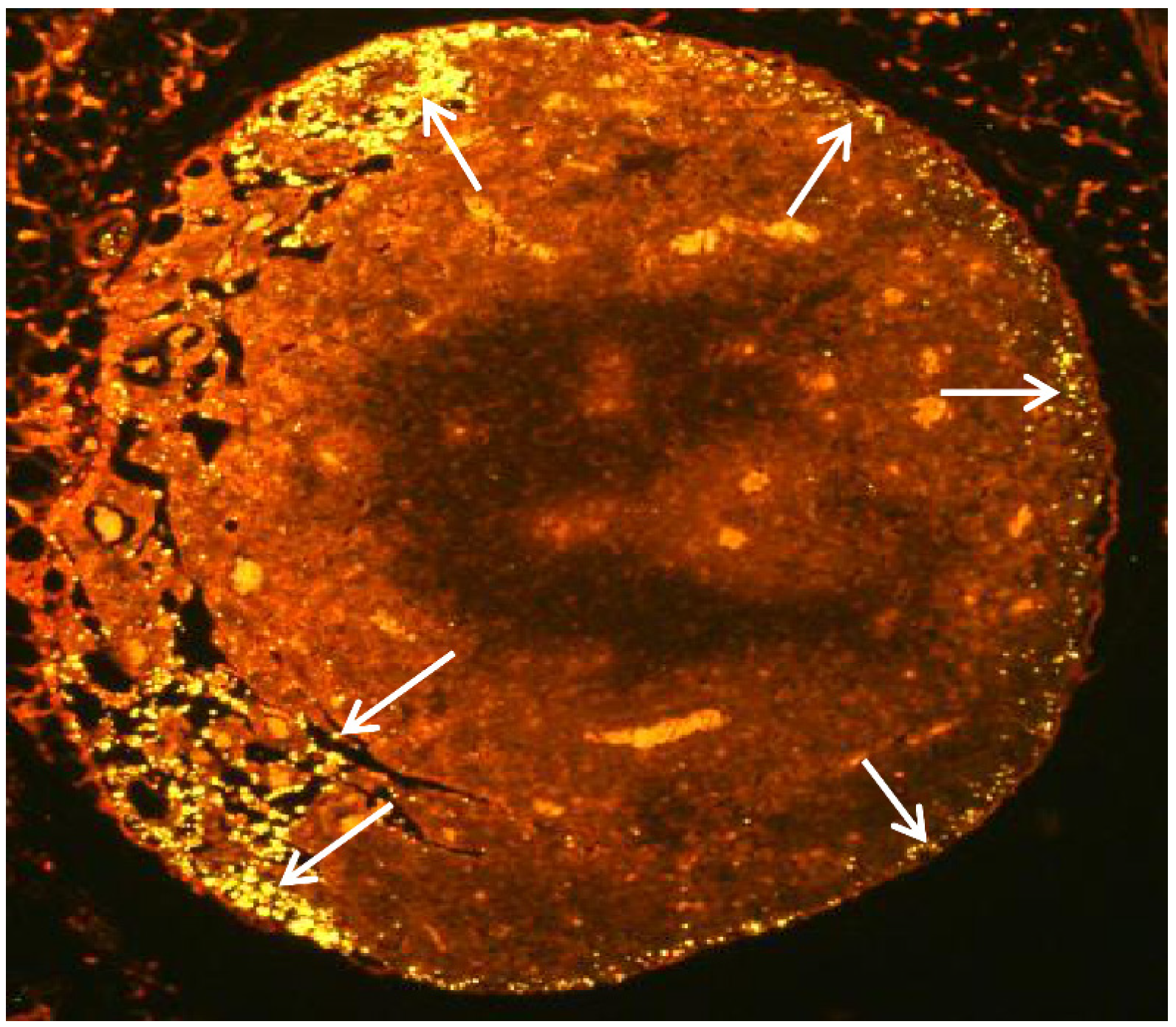
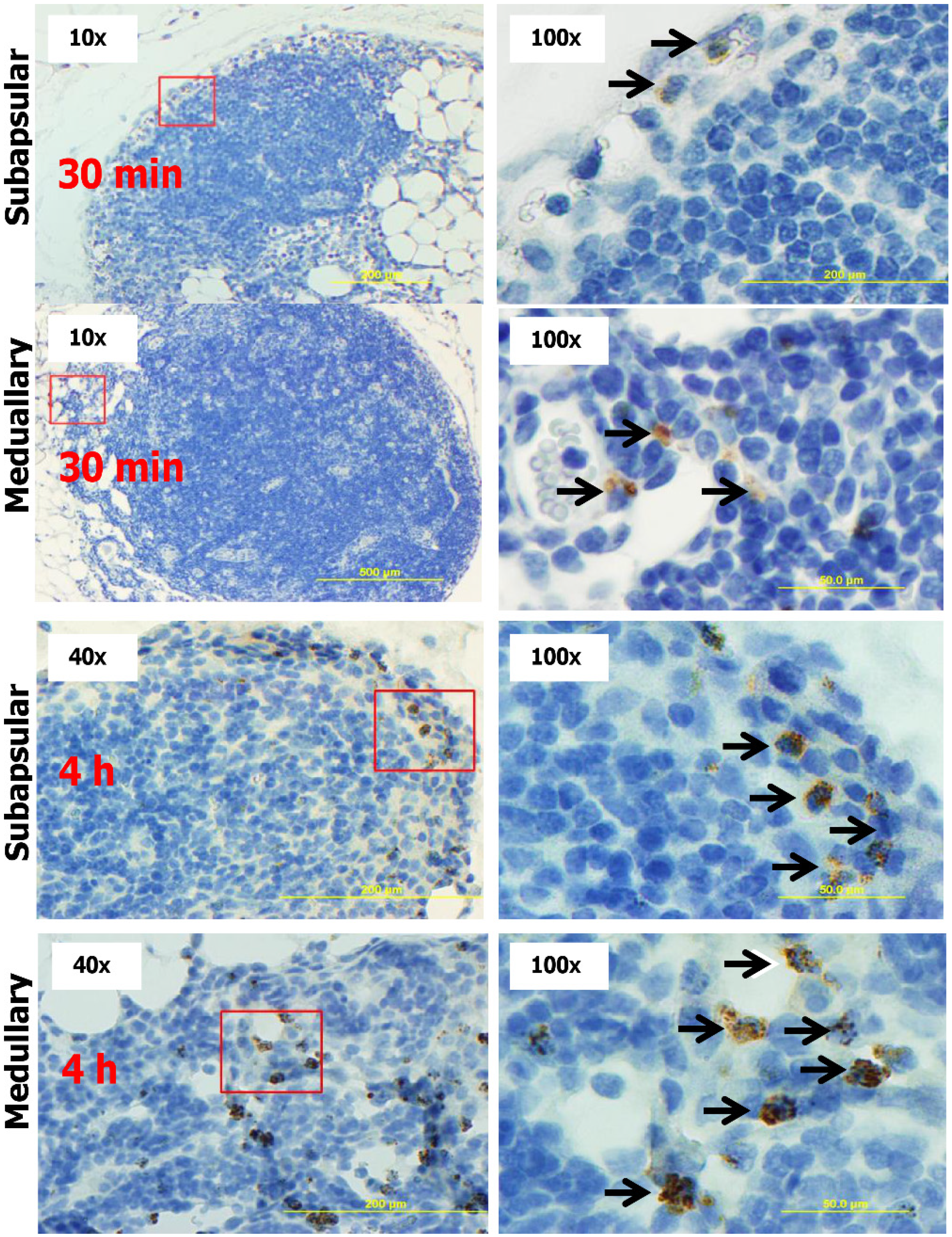
2.4. Subcutaneous Injection Quickly Delivers NPs to Regional LNs
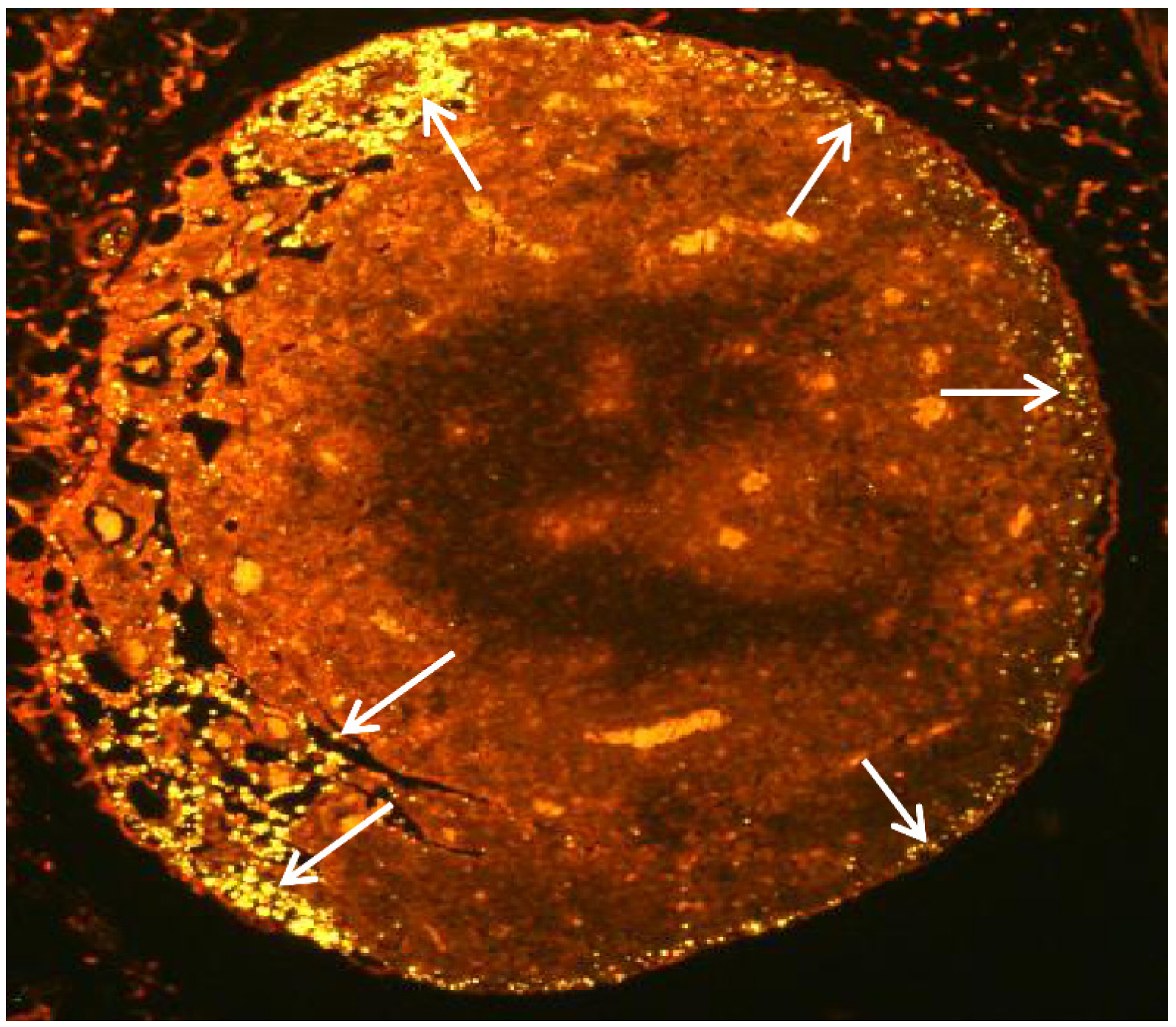
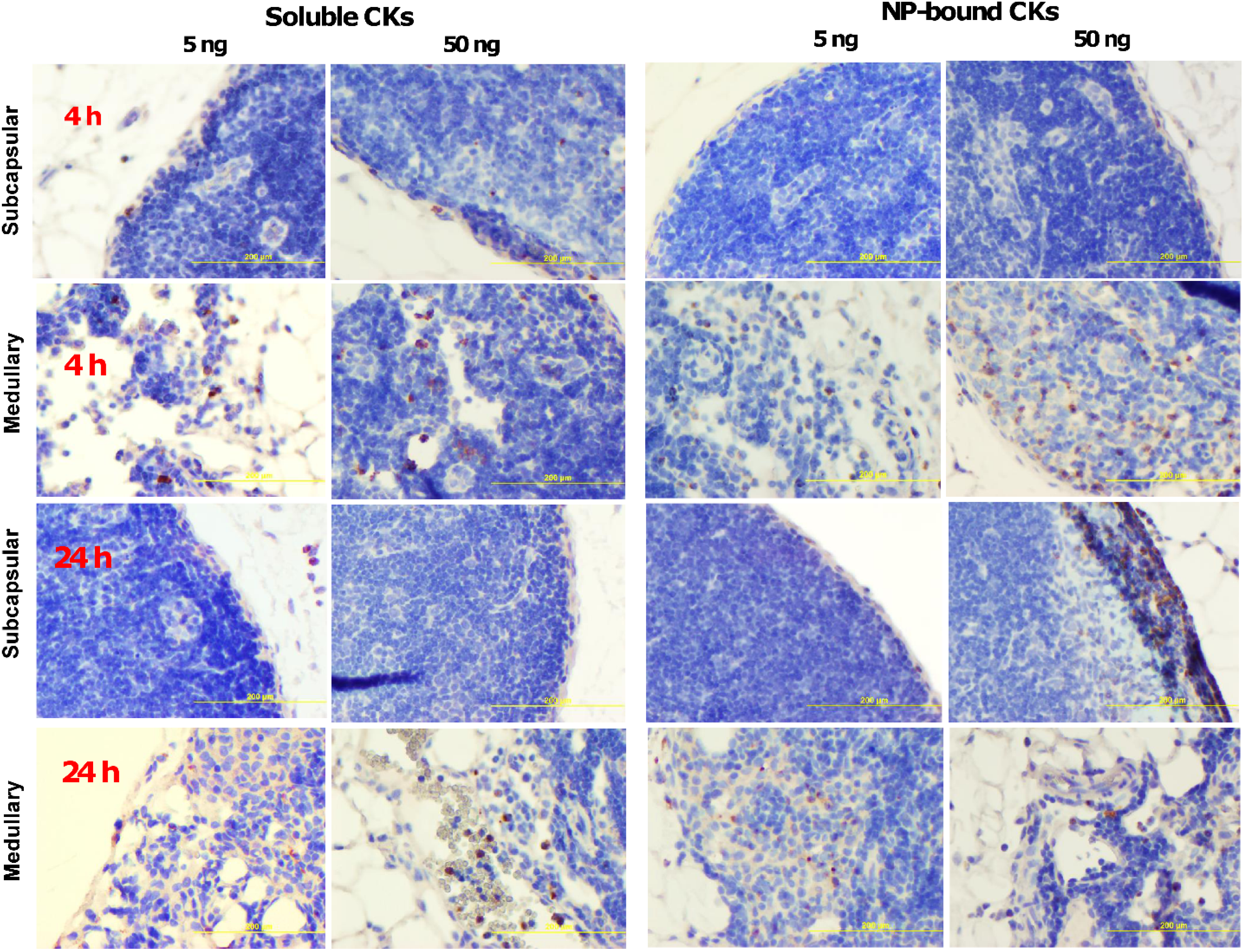
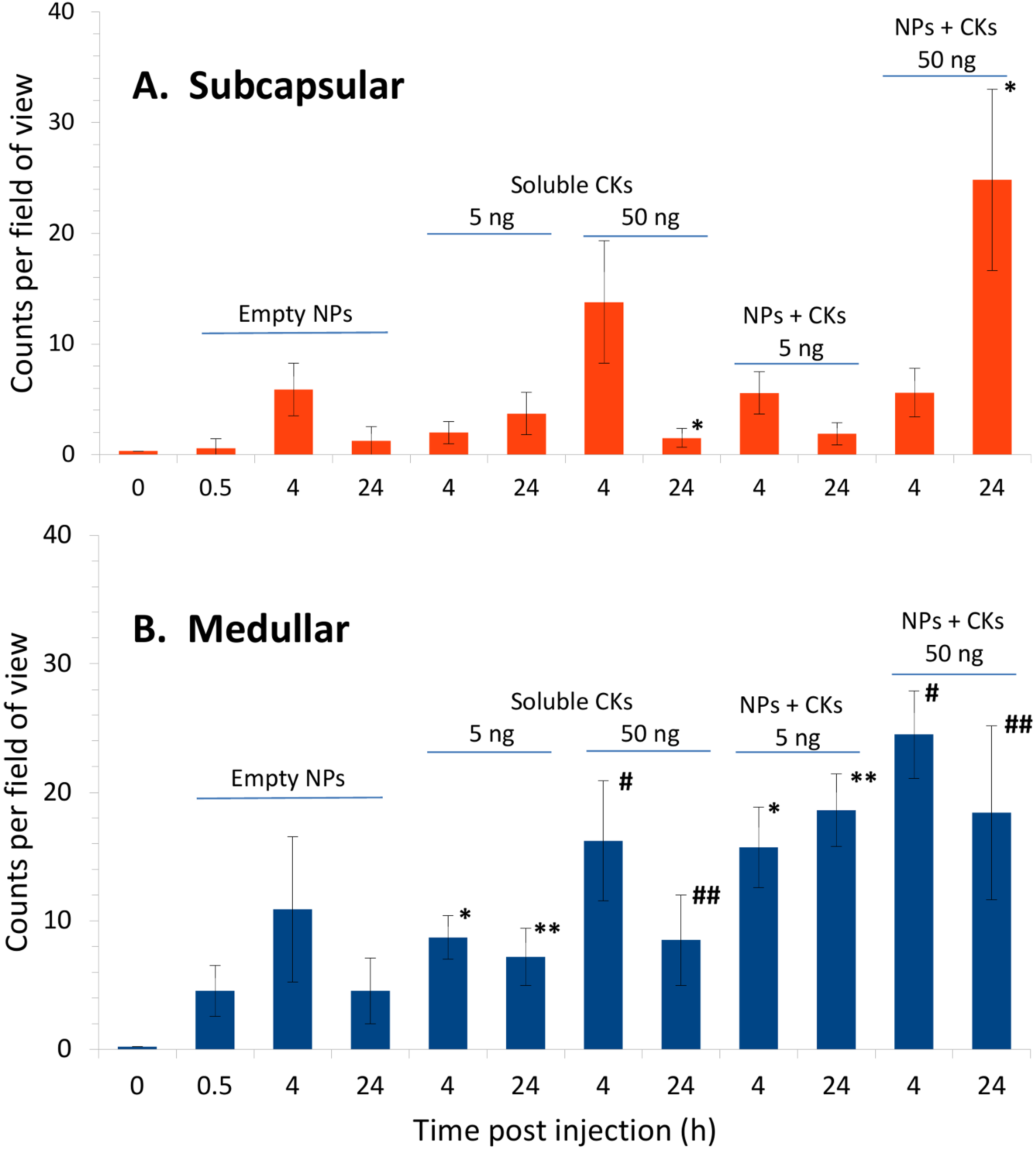
2.5. CK-Loaded NPs Mobilize Immune Cells to the LNs upon Administration to Mice
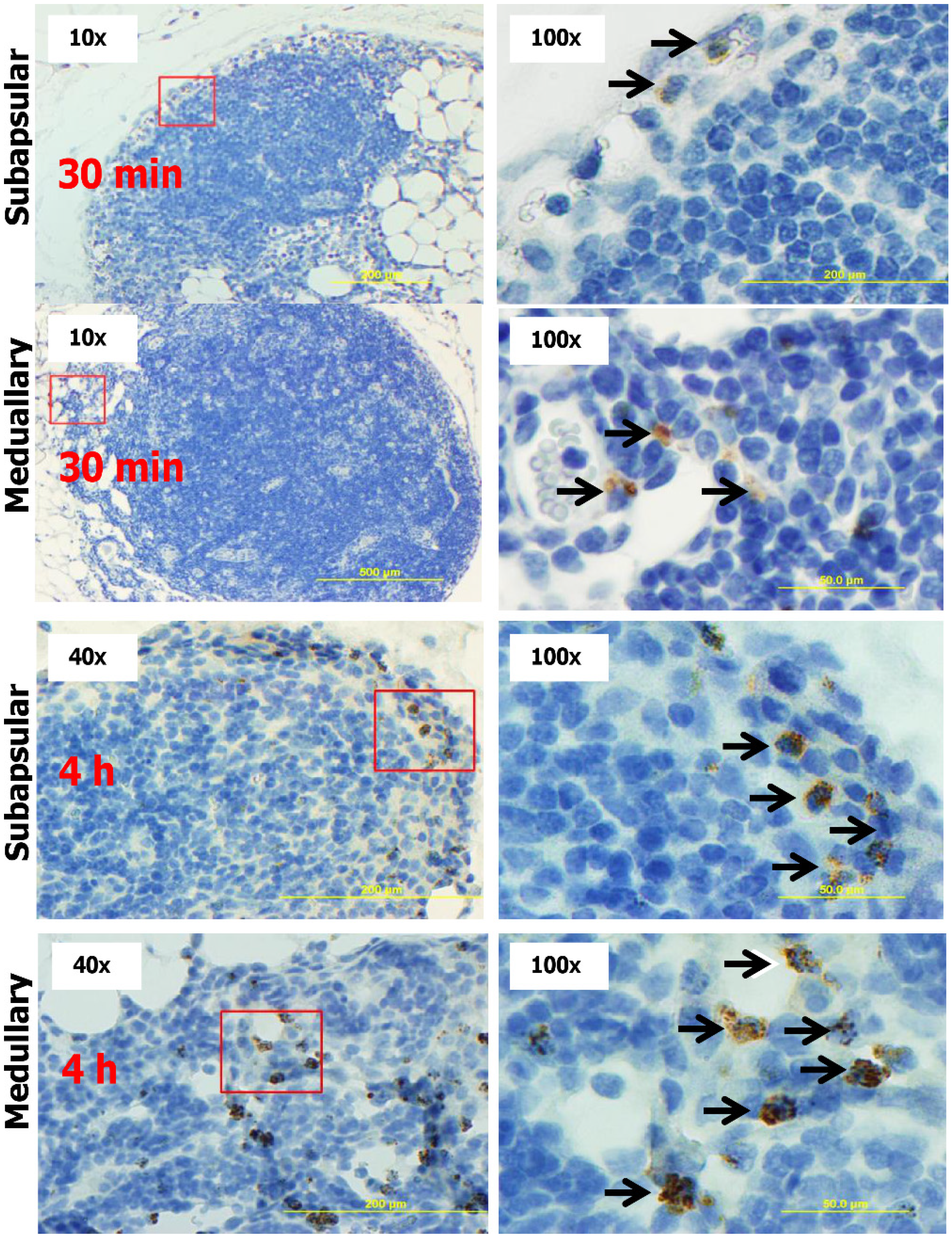
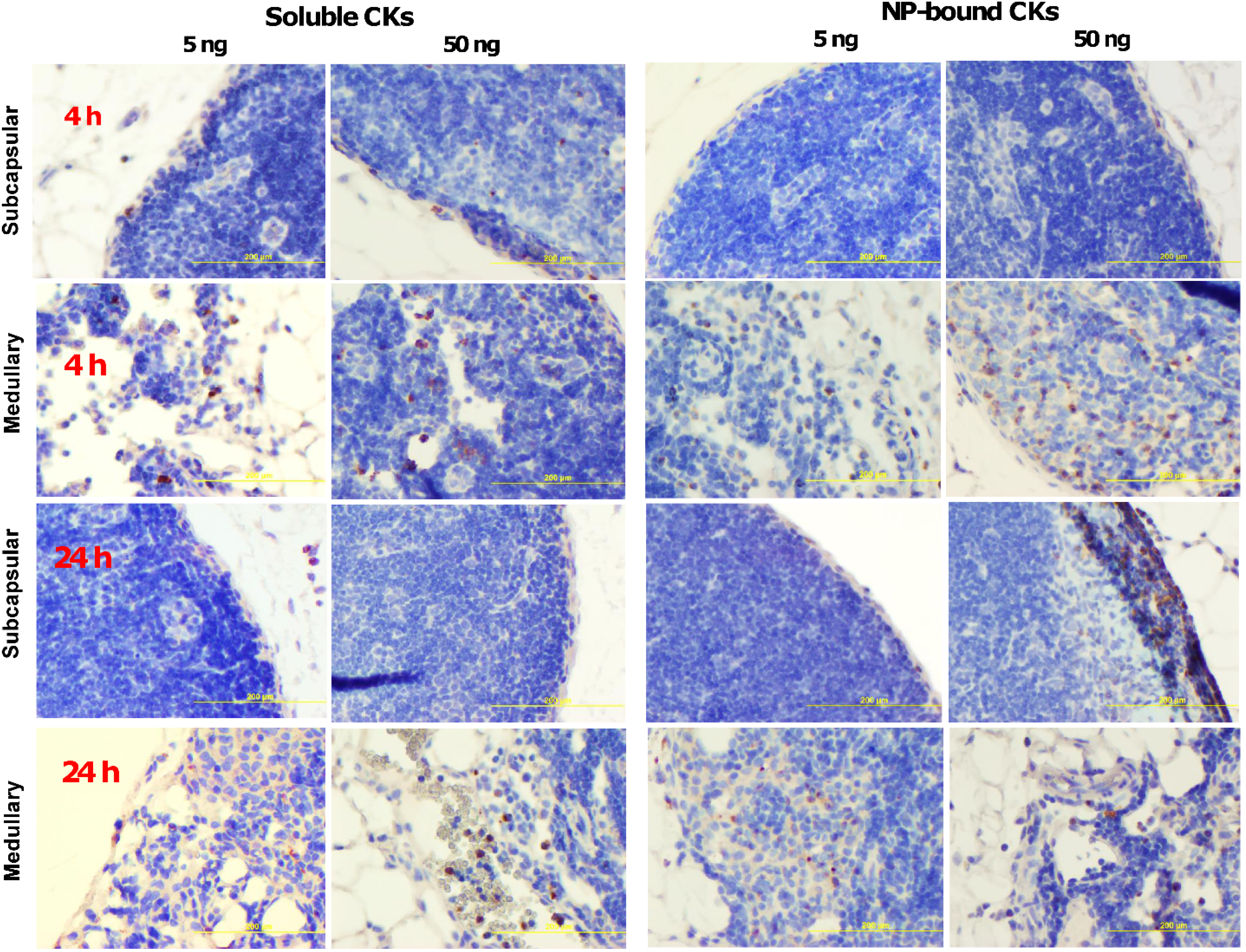
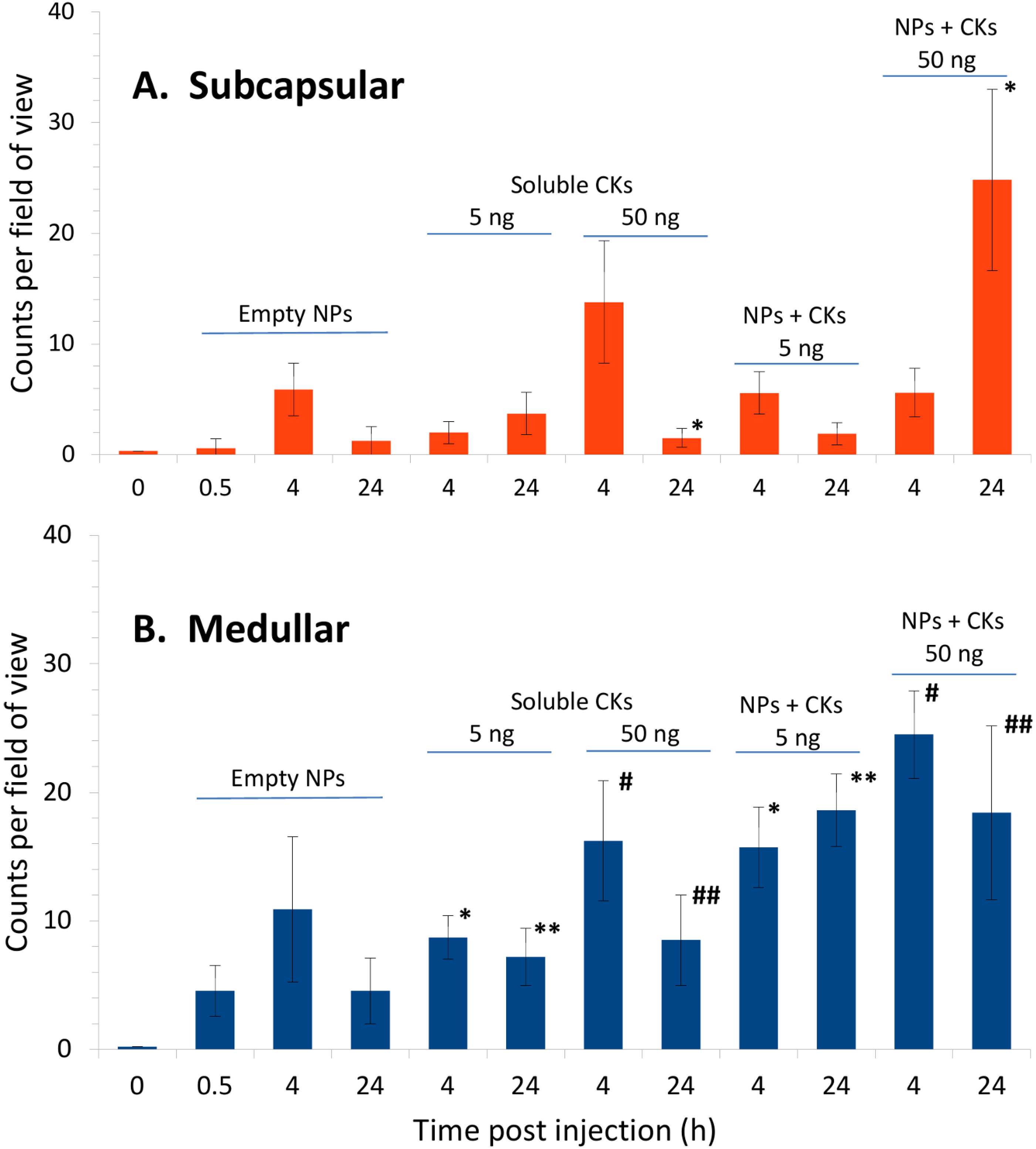
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Synthesis of NPs
4.3. Analysis of CK Binding Using ELISA and Western Blot
4.4. Labeling of pNIPAm-co-AA Particles with Alexa Fluor 555
4.5. Animal Challenge and LN Analysis
5. Conclusions
Acknowledgments
Author Contributions
Appendix


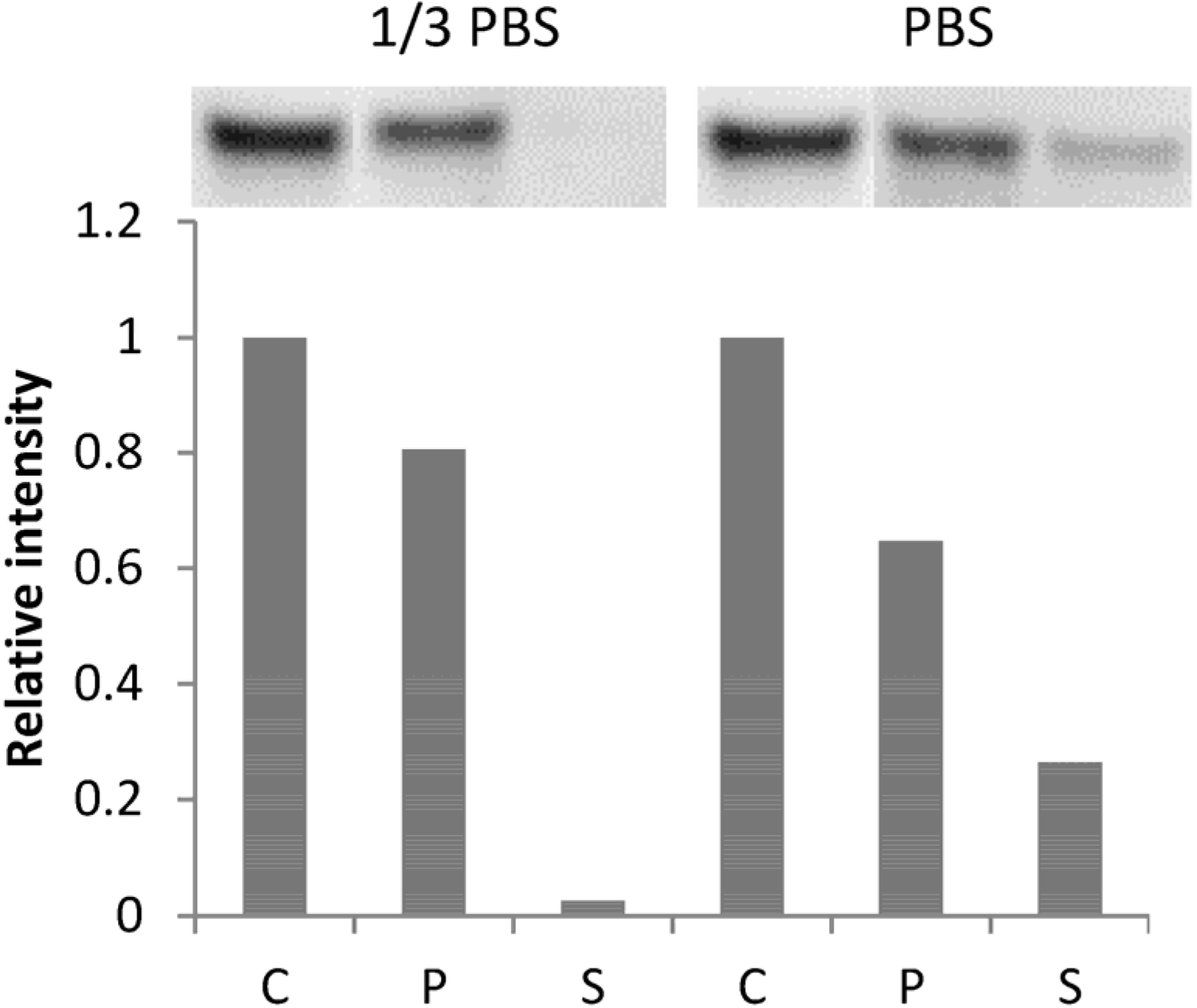

Conflicts of Interest
References
- Sadik, C.D.; Luster, A.D. Lipid-cytokine-chemokine cascades orchestrate leukocyte recruitment in inflammation. J. Leukoc. Biol. 2012, 91, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.; Hwang, J.M.; Kubes, P. Modulating leukocyte recruitment in inflammation. J. Allergy Clin. Immunol. 2007, 120, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Godaly, G.; Bergsten, G.; Hang, L.; Fischer, H.; Frendéus, B.; Lundstedt, A.C.; Samuelsson, M.; Samuelsson, P.; Svanborg, C. Neutrophil recruitment, chemokine receptors, and resistance to mucosal infection. J. Leukoc. Biol. 2001, 69, 899–906. [Google Scholar] [PubMed]
- Liston, A.; McColl, S. Subversion of the chemokine world by microbial pathogens. BioEssays 2003, 25, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Hiroshi Kohara, Y.T. Review: Tissue engineering technology to enhance cell recruitment for regeneration therapy. J. Med. Biol. Eng. 2010, 30, 267–276. [Google Scholar]
- Houshmand, P.; Zlotnik, A. Therapeutic applications in the chemokine superfamily. Curr. Opin. Chem. Biol. 2003, 7, 457–460. [Google Scholar] [CrossRef]
- Balmert, S.C.; Little, S.R. Biomimetic delivery with micro- and nanoparticles. Adv. Mater. 2012, 24, 3757–3778. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Irvine, D.J. Engineering chemoattractant gradients using chemokine-releasing polysaccharide microspheres. Biomaterials 2011, 32, 4903–4913. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Jain, S.; Benjamin Larman, H.; Gonzalez, S.; Irvine, D.J. Directed cell migration via chemoattractants released from degradable microspheres. Biomaterials 2005, 26, 5048–5063. [Google Scholar] [CrossRef] [PubMed]
- Kumamoto, T.; Huang, E.K.; Paek, H.J.; Morita, A.; Matsue, H.; Valentini, R.F.; Takashima, A. Induction of tumor-specific protective immunity by in situ Langerhans cell vaccine. Nat. Biotechnol. 2002, 20, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Suri, S.; Roy, K. In-situ crosslinking hydrogels for combinatorial delivery of chemokines and siRNA-DNA carrying microparticles to dendritic cells. Biomaterials 2009, 30, 5187–5200. [Google Scholar] [CrossRef] [PubMed]
- Kress, H.; Park, J.-G.; Mejean, C.O.; Forster, J.D.; Park, J.; Walse, S.S.; Zhang, Y.; Wu, D.; Weiner, O.D.; Fahmy, T.M.; et al. Cell stimulation with optically manipulated microsources. Nat. Methods 2009, 6, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Cross, D.P.; Wang, C. Stromal-derived factor-1 alpha-loaded PLGA microspheres for stem cell recruitment. Pharm. Res. 2011, 28, 2477–2489. [Google Scholar] [CrossRef] [PubMed]
- Kuraitis, D.; Zhang, P.; Zhang, Y.; Padavan, D.T.; McEwan, K.; Sofrenovic, T.; McKee, D.; Zhang, J.; Griffith, M.; Cao, X.; et al. A stromal cell-derived factor-1 releasing matrix enhances the progenitor cell response and blood vessel growth in ischaemic skeletal muscle. Eur. Cell. Mater. 2011, 22, 109–123. [Google Scholar]
- Zhang, G.; Nakamura, Y.; Wang, X.; Hu, Q.; Suggs, L.J.; Zhang, J. Controlled release of stromal cell-derived factor-1 alpha in situ increases c-kit+ cell homing to the infarcted heart. Tissue Eng. 2007, 13, 2063–2071. [Google Scholar] [CrossRef] [PubMed]
- Van de Weert, M.; Hennink, W.E.; Jiskoot, W. Protein instability in poly(lactic-co-glycolic acid) microparticles. Pharm. Res. 2000, 17, 1159–1167. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Mallery, S.R.; Schwendeman, S.P. Stabilization of proteins encapsulated in injectable poly (lactide-co-glycolide). Nat. Biotechnol. 2000, 18, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Luchini, A.; Geho, D.H.; Bishops, B.; Tran, D.; Xia, C.; Dufour, R.L.; Jones, C.D.; Espina, V.; Patanarut, A.; Zhou, W.; et al. Smart hydrogel particles: Biomarker harvesting: One-step affinity purification, size exclusion, and protection against degradation. Nano Lett. 2008, 8, 350–361. [Google Scholar] [CrossRef]
- Longo, C.; Patanarut, A.; George, T.; Bishop, B.; Zhou, W.; Fredolini, C.; Ross, M.M.; Espina, V.; Pellacani, G.; Petricoin, E.F.; et al. Core-shell hydrogel particles harvest, concentrate and preserve labile low abundance biomarkers. PLoS One 2009, 4, e4763. [Google Scholar] [CrossRef]
- Luchini, A.; Fredolini, C.; Espina, B.H.; Meani, F.; Reeder, A.; Rucker, S.; Petricoin, E.F.; Liotta, L.A. Nanoparticle technology: Addressing the fundamental roadblocks to protein biomarker discovery. Curr. Mol. Med. 2010, 10, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Tamburro, D.; Fredolini, C.; Espina, V.; Douglas, T.A.; Ranganathan, A.; Ilag, L.; Zhou, W.; Russo, P.; Espina, B.H.; Muto, G.; et al. Multifunctional core-shell nanoparticles: Discovery of previously invisible biomarkers. J. Am. Chem. Soc. 2011, 133, 19178–19188. [Google Scholar] [CrossRef]
- Longo, C.; Gambara, G.; Espina, V.; Luchini, A.; Bishop, B.; Patanarut, A.S.; Petricoin, E.F.; Beretti, F.; Ferrari, B.; Garaci, E.; et al. A novel biomarker harvesting nanotechnology identifies Bak as a candidate melanoma biomarker in serum. Exp. Dermatol. 2011, 20, 29–34. [Google Scholar] [CrossRef]
- Douglas, T.A.; Tamburro, D.; Fredolini, C.; Espina, B.H.; Lepene, B.S.; Ilag, L.; Espina, V.; Petricoin, E.F.; Liotta, L.A.; Luchini, A. The use of hydrogel microparticles to sequester and concentrate bacterial antigens in a urine test for Lyme disease. Biomaterials 2011, 32, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Luchini, A.; Tamburro, D.; Magni, R.; Fredolini, C.; Espina, V.; Bosch, J.; Garaci, E.; Petricoin, E.F.; Liotta, L. Application of analyte harvesting nanoparticle technology to the measurement of urinary HGH in healthy individuals. J. Sports Med. Doping Stud. 2012, 2, 2–4. [Google Scholar] [CrossRef]
- Bosch, J.; Luchini, A.; Pichini, S.; Tamburro, D.; Fredolini, C.; Liotta, L.; Petricoin, E.; Pacifici, R.; Facchiano, F.; Segura, J.; et al. Analysis of urinary human growth hormone (HGH) using hydrogel nanoparticles and isoform differential immunoassays after short recombinant HGH treatment: Preliminary results. J. Pharm. Biomed. Anal. 2013, 85, 194–197. [Google Scholar] [CrossRef]
- Magni, R.; Espina, B.H.; Liotta, L.A.; Luchini, A.; Espina, V. Hydrogel nanoparticle harvesting of plasma or urine for detecting low abundance proteins. J. Vis. Exp. 2014, e51789. [Google Scholar]
- Castro-Sesquen, Y.E.; Gilman, R.H.; Galdos-Cardenas, G.; Ferrufino, L.; Sánchez, G.; Valencia Ayala, E.; Liotta, L.; Bern, C.; Luchini, A. Use of a novel chagas urine nanoparticle test (Chunap) for diagnosis of congenital chagas disease. PLoS Negl. Trop. Dis. 2014, 8, e3211. [Google Scholar] [CrossRef] [PubMed]
- Shafagati, N.; Patanarut, A.; Luchini, A.; Lundberg, L.; Bailey, C.; Petricoin, E.; Liotta, L.; Narayanan, A.; Lepene, B.; Kehn-Hall, K. The use of nanotrap particles for biodefense and emerging infectious disease diagnostics. Pathog. Dis. 2014, 71, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Wenger, Y.; Schneider, R.J.; Reddy, G.R.; Kopelman, R.; Jolliet, O.; Philbert, M.A. Tissue distribution and pharmacokinetics of stable polyacrylamide nanoparticles following intravenous injection in the rat. Toxicol. Appl. Pharmacol. 2011, 251, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Lemperle, G.; Morhenn, V.; Charrier, U. Human histology and persistence of various injectable filler substances for soft tissue augmentation. Aesthet. Plast. Surg. 2003, 27, 354–366. [Google Scholar] [CrossRef]
- Gao, D.; Xu, H.; Philbert, M.A.; Kopelman, R. Bioeliminable nanohydrogels for drug delivery. Nano Lett. 2008, 8, 3320–3324. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Punekar, N.S. High-throughput screening of dye-ligands for chromatography. Methods Mol. Biol. 2014, 1129, 53–65. [Google Scholar] [PubMed]
- Denizli, A.; Pişkin, E. Dye-ligand affinity systems. J. Biochem. Biophys. Methods 2001, 49, 391–416. [Google Scholar] [CrossRef] [PubMed]
- Sereikaite, J.; Bumelis, V.A. Examination of dye-protein interaction by gel-permeation chromatography. Biomed. Chromatogr. 2006, 20, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Proudfoot, A.E.I.; Handel, T.M.; Johnson, Z.; Lau, E.K.; LiWang, P.; Clark-Lewis, I.; Borlat, F.; Wells, T.N.C.; Kosco-Vilbois, M.H. Glycosaminoglycan binding and oligomerization are essential for the in vivo activity of certain chemokines. Proc. Natl. Acad. Sci. USA 2003, 100, 1885–1890. [Google Scholar] [CrossRef] [PubMed]
- Al-Alwan, L.A.; Chang, Y.; Mogas, A.; Halayko, A.J.; Baglole, C.J.; Martin, J.G.; Rousseau, S.; Eidelman, D.H.; Hamid, Q. Differential roles of CXCL2 and CXCL3 and their receptors in regulating normal and asthmatic airway smooth muscle cell migration. J. Immunol. 2013, 191, 2731–2741. [Google Scholar] [CrossRef] [PubMed]
- Dennis, M.S.; Zhang, M.; Meng, Y.G.; Kadkhodayan, M.; Kirchhofer, D.; Combs, D.; Damico, L.A. Albumin binding as a general strategy for improving the pharmacokinetics of proteins. J. Biol. Chem. 2002, 277, 35035–35043. [Google Scholar] [CrossRef] [PubMed]
- Böhme, U.; Scheler, U. Effective charge of bovine serum albumin determined by electrophoresis NMR. Chem. Phys. Lett. 2007, 435, 342–345. [Google Scholar] [CrossRef]
- Manolova, V.; Flace, A.; Bauer, M.; Schwarz, K.; Saudan, P.; Bachmann, M.F. Nanoparticles target distinct dendritic cell populations according to their size. Eur. J. Immunol. 2008, 38, 1404–1413. [Google Scholar] [CrossRef] [PubMed]
- Das, S.T.; Rajagopalan, L.; Guerrero-Plata, A.; Sai, J.; Richmond, A.; Garofalo, R.P.; Rajarathnam, K. Monomeric and dimeric CXCL8 are both essential for in vivo neutrophil recruitment. PLoS One 2010, 5, e11754. [Google Scholar] [CrossRef] [PubMed]
- Baggiolini, M.; Walz, A.; Kunkel, S.L. Neutrophil-activating peptide-1/interleukin 8, a novel cytokine that activates neutrophils. J. Clin. Investig. 1989, 84, 1045–1049. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.D.L.; Canetti, C.; Souto, J.T.; Silva, J.S.; Hogaboam, C.M.; Ferreira, S.H.; Cunha, F.Q. MIP-1alpha[CCL3] acting on the CCR1 receptor mediates neutrophil migration in immune inflammation via sequential release of TNF-alpha and LTB4. J. Leukoc. Biol. 2005, 78, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-W.; Strong, B.S.I.; Miller, M.J.; Unanue, E.R. Neutrophils influence the level of antigen presentation during the immune response to protein antigens in adjuvants. J. Immunol. 2010, 185, 2927–2934. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-W.; Unanue, E.R. Neutrophils control the magnitude and spread of the immune response in a thromboxane A2-mediated process. J. Exp. Med. 2013, 210, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Massena, S.; Christoffersson, G.; Hjertström, E.; Zcharia, E.; Vlodavsky, I.; Ausmees, N.; Rolny, C.; Li, J.P.; Phillipson, M. A chemotactic gradient sequestered on endothelial heparan sulfate induces directional intraluminal crawling of neutrophils. Blood 2010, 116, 1924–1931. [Google Scholar] [CrossRef] [PubMed]
- Kuschert, G.S.; Coulin, F.; Power, C.A.; Proudfoot, A.E.; Hubbard, R.E.; Hoogewerf, A.J.; Wells, T.N. Glycosaminoglycans interact selectively with chemokines and modulate receptor binding and cellular responses. Biochemistry 1999, 38, 12959–12968. [Google Scholar] [CrossRef] [PubMed]
- Hoogewerf, A.J.; Kuschert, G.S.; Proudfoot, A.E.; Borlat, F.; Clark-Lewis, I.; Power, C.A.; Wells, T.N. Glycosaminoglycans mediate cell surface oligomerization of chemokines. Biochemistry 1997, 36, 13570–13578. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Kao, W.J. Drug release kinetics and transport mechanisms of non-degradable and degradable polymeric delivery systems. Expert Opin. Drug Deliv. 2010, 7, 429–444. [Google Scholar] [CrossRef] [PubMed]
- Colditz, I.; Zwahlen, R.; Dewald, B.; Baggiolini, M. In vivo inflammatory activity of neutrophil-activating factor, a novel chemotactic peptide derived from human monocytes. Am. J. Pathol. 1989, 134, 755–760. [Google Scholar] [PubMed]
- Zwahlen, R.; Walz, A.; Rot, A. In vitro and in vivo activity and pathophysiology of human interleukin-8 and related peptides. Int. Rev. Exp. Pathol. 1993, 34B, 27–42. [Google Scholar] [PubMed]
- Taub, D.D.; Anver, M.; Oppenheim, J.J.; Longo, D.L.; Murphy, W.J. T lymphocyte recruitment by interleukin-8 (IL-8). IL-8-induced degranulation of neutrophils releases potent chemoattractants for human T lymphocytes both in vitro and in vivo. J. Clin. Investig. 1996, 97, 1931–1941. [Google Scholar] [CrossRef] [PubMed]
- Menten, P.; Wuyts, A.; van Damme, J. Macrophage inflammatory protein-1. Cytokine Growth Factor Rev. 2002, 13, 455–481. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Park, K. Environment-sensitive hydrogels for drug delivery. Adv. Drug Deliv. Rev. 2001, 53, 321–339. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, S.K.; Lee, J.C.; Murphy, P.M. CXC chemokines bind to unique sets of selectivity determinants that can function independently and are broadly distributed on multiple domains of human interleukin-8 receptor B. Determinants of high affinity binding and receptor activation are distinct. J. Biol. Chem. 1996, 271, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Holmes, W.E.; Lee, J.; Kuang, W.J.; Rice, G.C.; Wood, W.I. Structure and functional expression of a human interleukin-8 receptor. Science 1991, 253, 1278–1280. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.M.; Tiffany, H.L. Cloning of complementary DNA encoding a functional human interleukin-8 receptor. Science 1991, 253, 1280–1283. [Google Scholar] [CrossRef] [PubMed]
- Konrad, F.M.; Reutershan, J. CXCR2 in acute lung injury. Mediat. Inflamm. 2012, 2012, 740987. [Google Scholar]
- Ren, M.; Guo, Q.; Guo, L.; Lenz, M.; Qian, F.; Koenen, R.R.; Xu, H.; Schilling, A.B.; Weber, C.; Ye, R.D.; et al. Polymerization of MIP-1 chemokine (CCL3 and CCL4) and clearance of MIP-1 by insulin-degrading enzyme. EMBO J. 2010, 29, 3952–3966. [Google Scholar] [CrossRef] [PubMed]
- Abadie, V.; Badell, E.; Douillard, P.; Ensergueix, D.; Leenen, P.J.M.; Tanguy, M.; Fiette, L.; Saeland, S.; Gicquel, B.; Winter, N. Neutrophils rapidly migrate via lymphatics after Mycobacterium bovis BCG intradermal vaccination and shuttle live bacilli to the draining lymph nodes. Blood 2005, 106, 1843–1850. [Google Scholar] [CrossRef] [PubMed]
- Maletto, B.A.; Ropolo, A.S.; Alignani, D.O.; Liscovsky, M.V.; Ranocchia, R.P.; Moron, V.G.; Pistoresi-Palencia, M.C. Presence of neutrophil-bearing antigen in lymphoid organs of immune mice. Blood 2006, 108, 3094–3102. [Google Scholar] [CrossRef] [PubMed]
- Chtanova, T.; Schaeffer, M.; Han, S.J.; van Dooren, G.G.; Nollmann, M.; Herzmark, P.; Chan, S.W.; Satija, H.; Camfield, K.; Aaron, H.; et al. Dynamics of neutrophil migration in lymph nodes during infection. Immunity 2008, 29, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Brackett, C.M.; Muhitch, J.B.; Evans, S.S.; Gollnick, S.O. IL-17 promotes neutrophil entry into tumor-draining lymph nodes following induction of sterile inflammation. J. Immunol. 2013, 191, 4348–4357. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popova, T.G.; Teunis, A.; Magni, R.; Luchini, A.; Espina, V.; Liotta, L.A.; Popov, S.G. Chemokine-Releasing Nanoparticles for Manipulation of the Lymph Node Microenvironment. Nanomaterials 2015, 5, 298-320. https://doi.org/10.3390/nano5010298
Popova TG, Teunis A, Magni R, Luchini A, Espina V, Liotta LA, Popov SG. Chemokine-Releasing Nanoparticles for Manipulation of the Lymph Node Microenvironment. Nanomaterials. 2015; 5(1):298-320. https://doi.org/10.3390/nano5010298
Chicago/Turabian StylePopova, Taissia G., Allison Teunis, Ruben Magni, Alessandra Luchini, Virginia Espina, Lance A. Liotta, and Serguei G. Popov. 2015. "Chemokine-Releasing Nanoparticles for Manipulation of the Lymph Node Microenvironment" Nanomaterials 5, no. 1: 298-320. https://doi.org/10.3390/nano5010298