An In Vitro Model for Assessing Corneal Keratocyte Spreading and Migration on Aligned Fibrillar Collagen

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
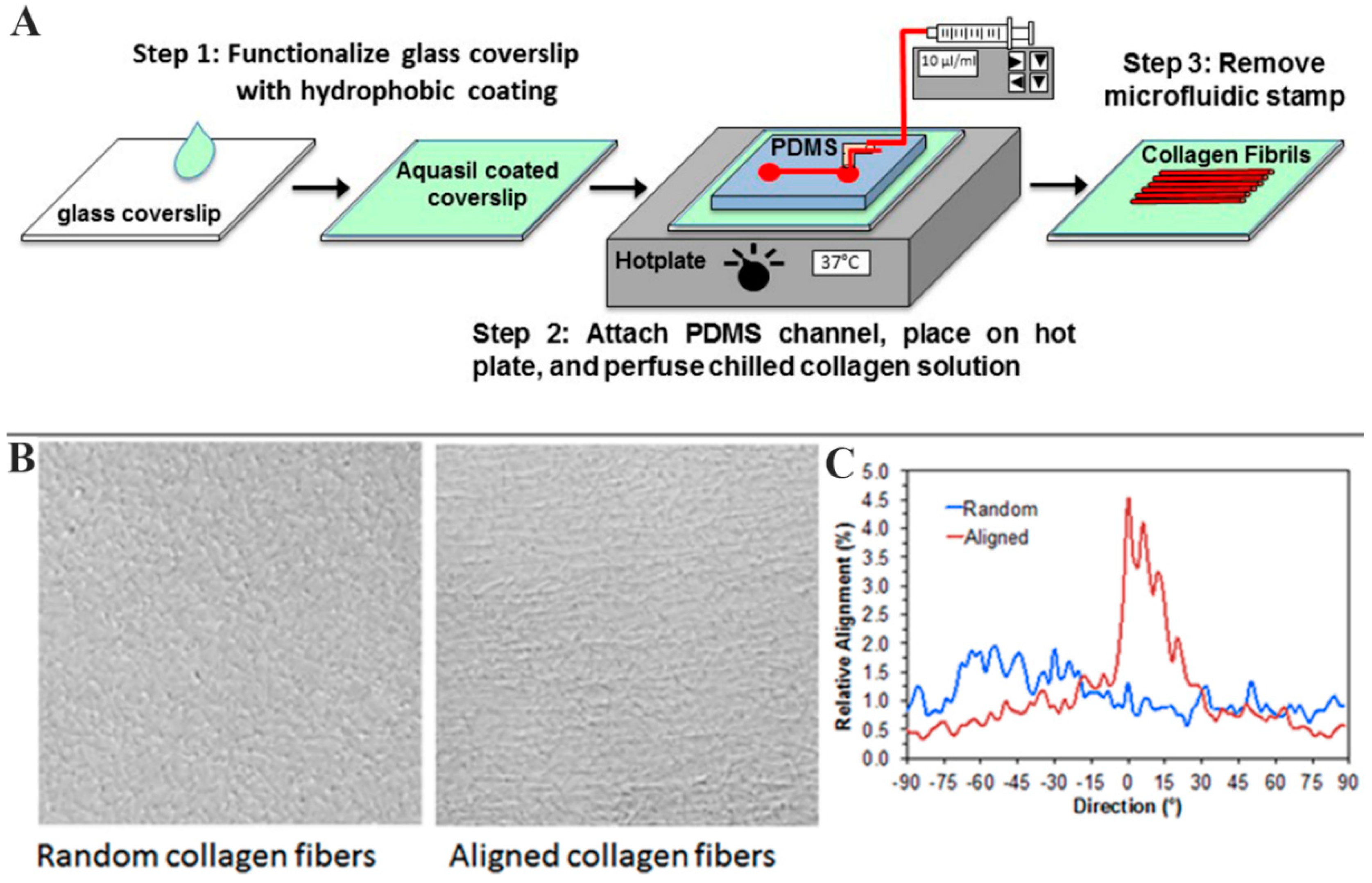
2.1. Experimental Models
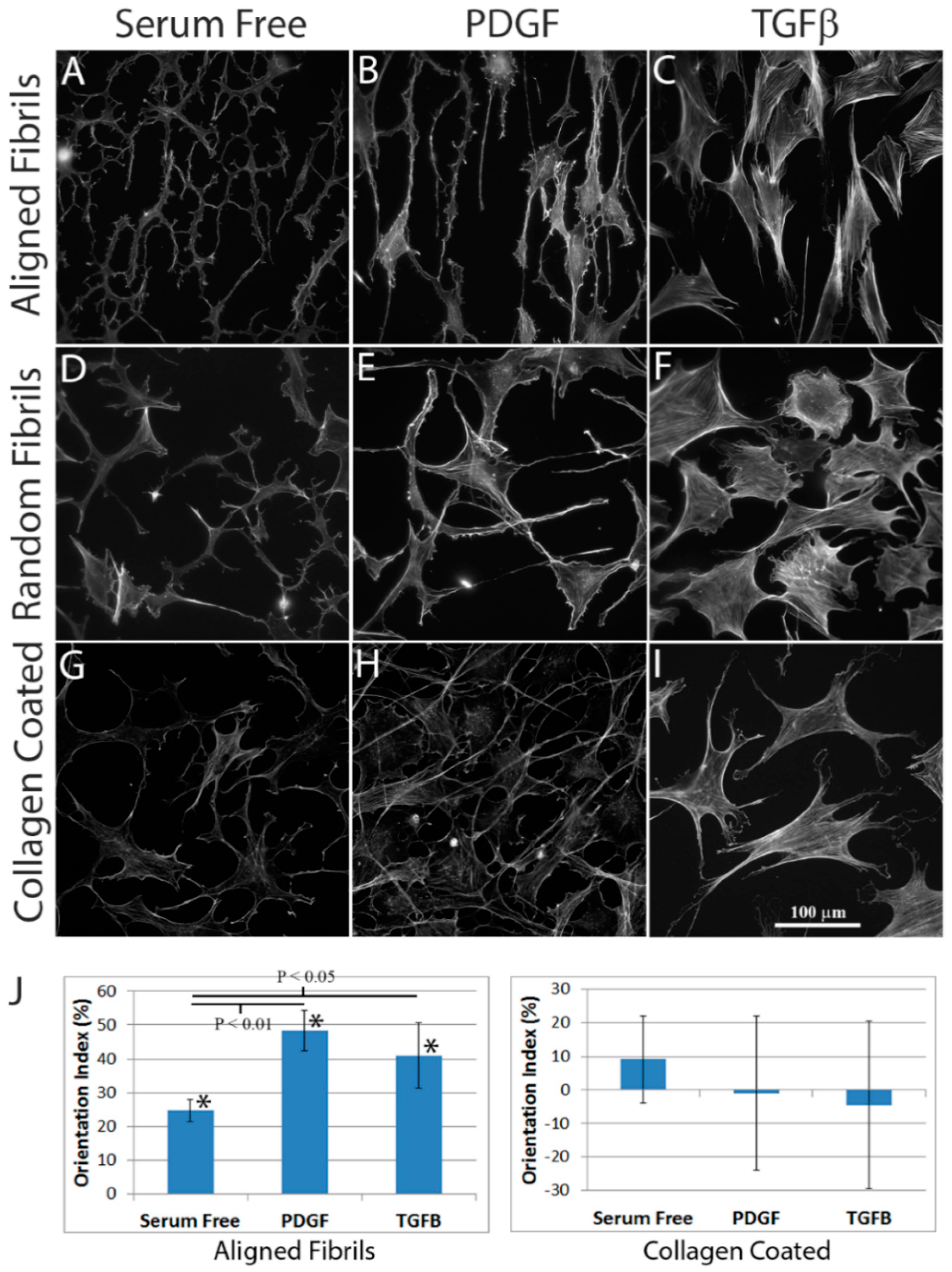
2.2. Effect of Substrate Alignment on Corneal Keratocyte Patterning
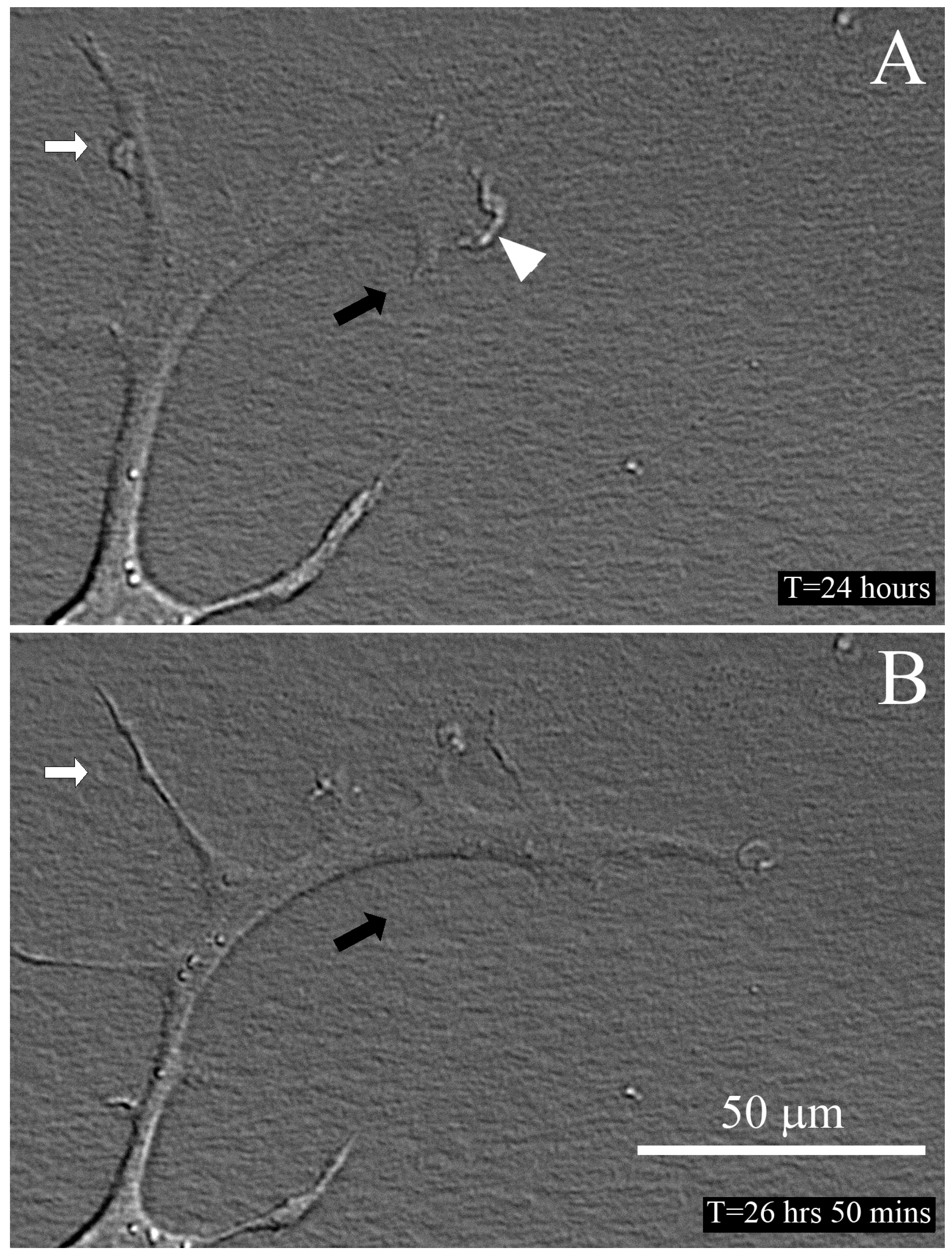
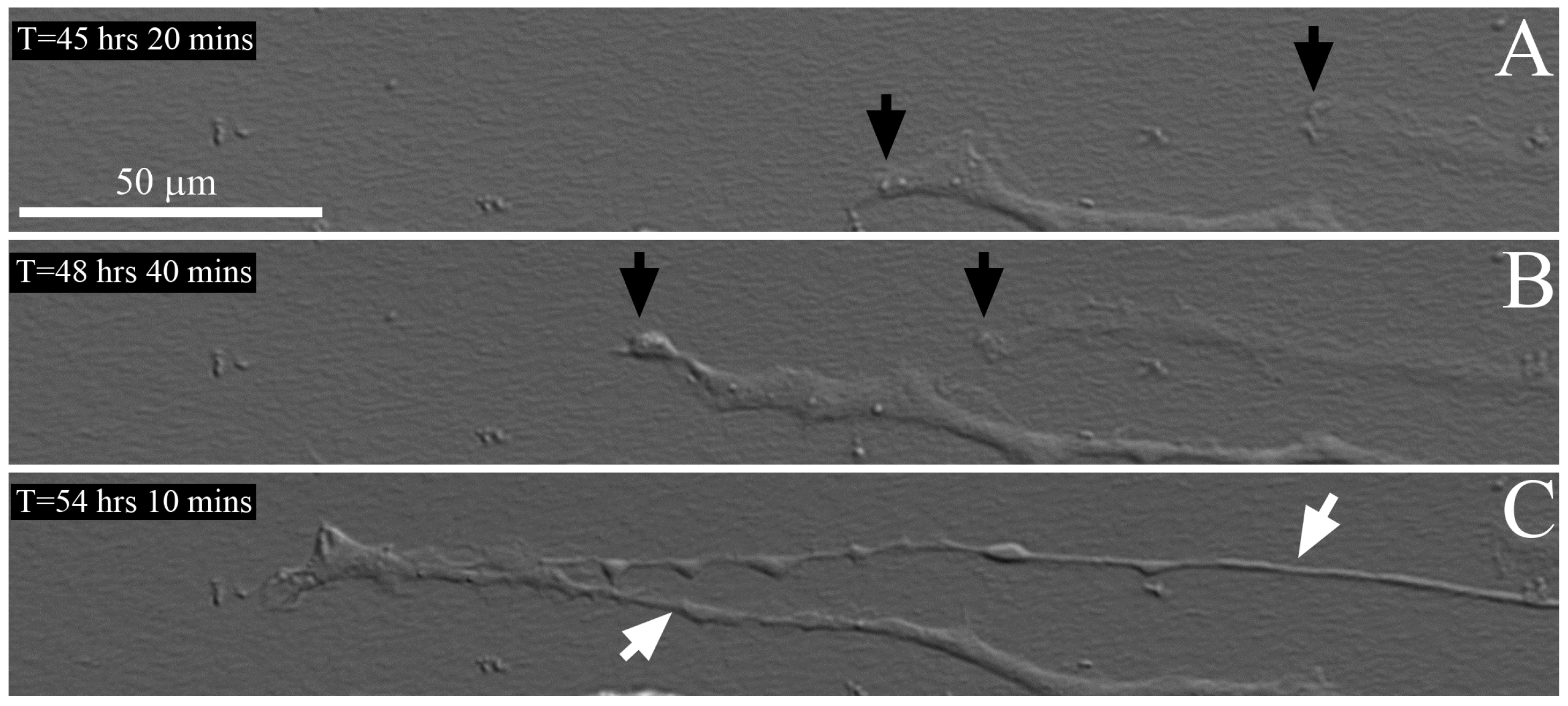
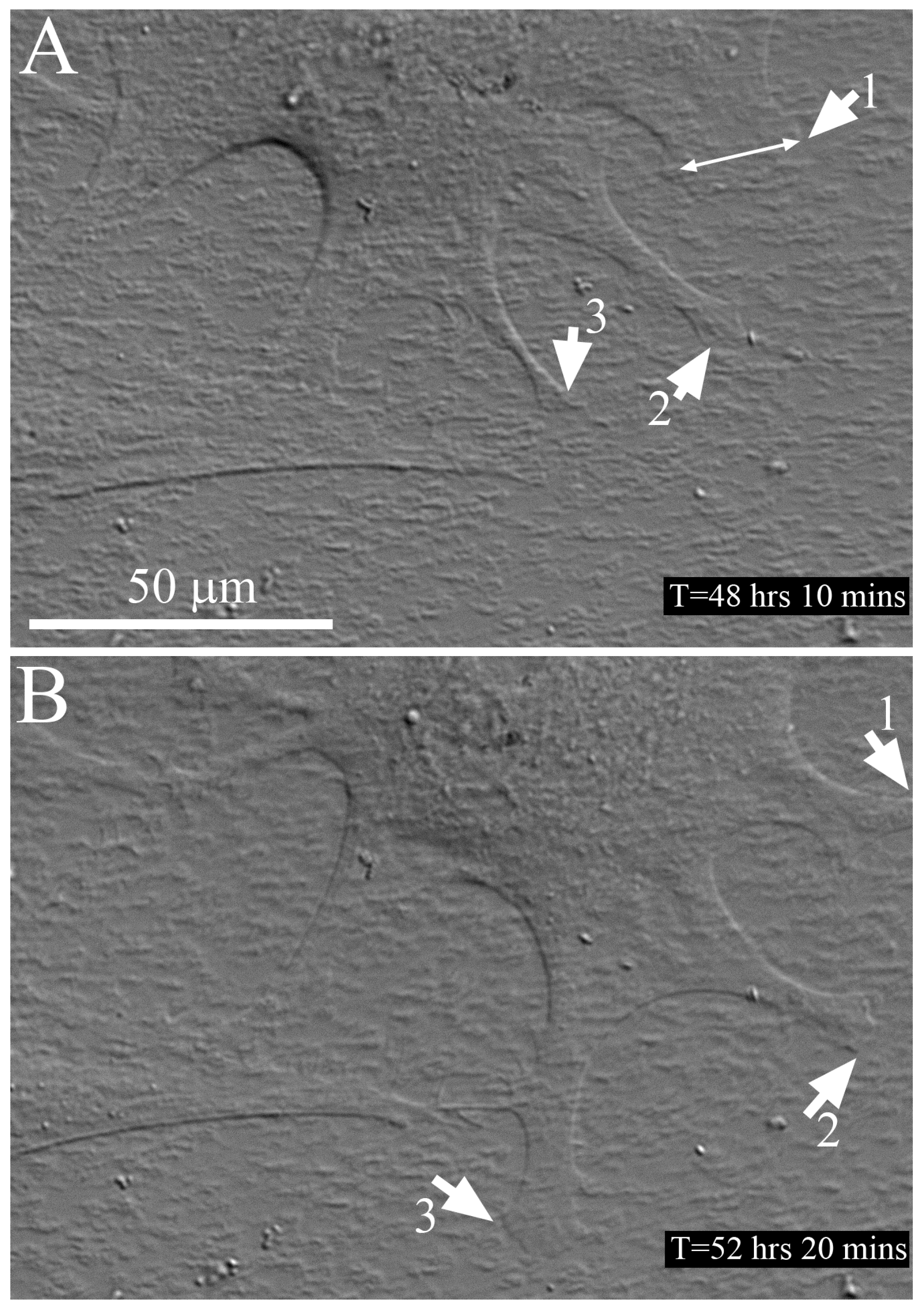
2.3. Time-Lapse Imaging of Cell-Collagen Interactions
3. Materials and Methods
3.1. Fabrication of Microfluidics Device
3.2. Fabrication of Collagen Substrates
3.3. Cell Culture
3.4. Freeze Injury (FI)
3.5. Immunocytochemistry
3.6. Scanning Electron Microscopy (SEM)
3.7. Orientation Index
3.8. Statistical Analysis
3.9. Time-Lapse Imaging
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Perez-Gomez, I.; Efron, N. Change to corneal morphology after refractive surgery (myopic laser in situ keratomileusis) as viewed with a confocal microscope. Optom. Vis. Sci. 2003, 80, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Chakravarti, S.; Petroll, W.M.; Hassell, J.; Jester, J.V.; Lass, J.H.; Paul, J.; Birk, D.E. Corneal opacity in lumican-null mice: Defects in collagen fibril structure and packing in the posterior stroma. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3365–3373. [Google Scholar]
- Funderburgh, J.L.; Mann, M.M.; Funderburgh, M.L. Keratocyte phenotype mediates proteoglycan structure: A role for fibroblasts in corneal fibrosis. J. Biol. Chem. 2003, 278, 45629–45637. [Google Scholar] [CrossRef] [PubMed]
- Hassell, J.R.; Birk, D.E. The molecular basis of corneal transparency. Exp. Eye Res. 2010, 91, 326–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Krieg, M.; Arboleda-Estudillo, Y.; Puech, P.-H.; Kafer, J.; Graner, F.; Muller, D.J.; Heisenberg, C.-P. Tensile forces govern germ-layer organization in zebrafish. Nat. Cell Biol. 2008, 10, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Stopak, D.; Wessels, N.K.; Harris, A.K. Morphogenetic rearrangement of injected collagen in developing chicken limb buds. Proc. Natl. Acad. Sci. USA 1985, 82, 2804–2808. [Google Scholar] [CrossRef] [PubMed]
- Bard, J.B.L.; Higginson, K. Fibroblast-collagen interactions in the formation of the secondary stroma of the chick cornea. J. Cell Biol. 1977, 74, 816–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garana, R.M.; Petroll, W.M.; Chen, W.T.; Herman, I.M.; Barry, P.; Andrews, H.D.; Cavanagh, H.D.; Jester, J.V. Radial keratotomy II: The role of the myofibroblast in corneal wound contraction. Investig. Ophthalmol. Vis. Sci. 1992, 33, 3271–3282. [Google Scholar]
- Moller-Pedersen, T.; Li, H.; Petroll, W.M.; Cavanagh, H.D.; Jester, J.V. Confocal microscopic characterization of wound repair after photorefractive keratectomy using in vivo confocal microscopy. Investig. Ophthalmol. Vis. Sci. 1998, 39, 487–501. [Google Scholar]
- Stramer, B.M.; Zieske, J.D.; Jung, J.-C.; Austin, J.S.; Fini, M.E. Molecular mechanisms controlling the fibrotic repair phenotype in cornea: Implications for surgical outcomes. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4237–4246. [Google Scholar] [CrossRef]
- Varner, V.D.; Nelson, C.M. Toward the directed self-assembly of engineered tissues. Annu. Rev. Chem. Biomol. Eng. 2014, 5, 507–526. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, R.M.; Gonzalez-Andrades, E.; Cardona, J.C.; Gonzalez-Gallardo, C.; Ionescu, A.M.; Garzon, I.; Alaminos, M.; Gonzalez-Andrades, M.; Connon, C.J. Controlling the 3D architecture of self-lifting auto-generated tissue equivalents (slates) for optimized corneal graft composition and stability. Biomaterials 2017, 121, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Kivanany, P.B.; Grose, K.C.; Tippani, M.; Su, S.; Petroll, W.M. Assessment of corneal stromal remodeling and regeneration after photorefractive keratectomy. Sci Rep. 2018, 8, 12580. [Google Scholar] [CrossRef] [PubMed]
- Cintron, C.; Kublin, C.L. Regeneration of corneal tissue. Dev. Biol. 1977, 61, 346–357. [Google Scholar] [CrossRef]
- Etheredge, L.; Kane, B.P.; Hassell, J.R. The effect of growth factor signaling on keratocytes in vitro and its relationship to the phases of stromal wound repair. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3128–3136. [Google Scholar] [CrossRef] [PubMed]
- Ghibaudo, M.; Trichet, L.; Le Digabel, J.; Richert, A.; Hersen, P.; Ladoux, B. Substrate topography induces a crossover from 2d to 3d behavior in fibroblast migration. Biophys. J. 2009, 97, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.I.; Nealey, P.F.; Murphy, C.J. Responses of human keratocytes to micro- and nanostructured substrates. J. Biomed. Mater. Res. A 2004, 71, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.I.; Abrams, G.A.; Bertics, P.J.; Murphy, C.J.; Nealey, P.F. Epithelial contact guidance on well-defined micro- and nanostructured substrates. J. Cell Sci. 2003, 116, 1881–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myrna, K.E.; Mendonsa, R.; Russell, P.; Pot, S.A.; Liliensiek, S.J.; Jester, J.V.; Nealey, P.F.; Brown, D.; Murphy, C.J. Substratum topography modulates corneal fibroblast to myofibroblast transformation. Investig. Ophthalmol. Vis. Sci. 2012, 53, 811–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillemette, M.D.; Cui, B.; Roy, E.; Gauvin, R.; Giasson, C.J.; Esch, M.B.; Carrier, P.; Deschambeault, A.; Dumoulin, M.; Toner, M.; et al. Surface topography induces 3D self-orientation of cells and extracellular matrix resulting in improved tissue function. Integr. Biol. 2009, 1, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Saeidi, N.; Guo, X.; Hutcheon, A.E.; Sander, E.A.; Bale, S.S.; Melotti, S.A.; Zieske, J.D.; Trinkaus-Randall, V.; Ruberti, J.W. Disorganized collagen scaffold interferes with fibroblast mediated deposition of organized extracellular matrix in vitro. Biotechnol. Bioeng. 2012, 109, 2683–2698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.Q.; Hutcheon, A.E.; Melotti, S.A.; Zieske, J.D.; Trinkaus-Randall, V.; Ruberti, J.W. Morphologic characterization of organized extracellular matrix deposition by ascorbic acid-stimulated human corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4056–4060. [Google Scholar] [CrossRef] [PubMed]
- Karamichos, D.; Guo, X.Q.; Hutcheon, A.E.; Zieske, J.D. Human corneal fibrosis: An in vitro model. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1382–1388. [Google Scholar] [CrossRef] [PubMed]
- Ren, R.; Hutcheon, A.E.K.; Guo, X.Q.; Saeidi, N.; Melotti, S.A.; Ruberti, J.W.; Zieske, J.D.; Trinkaus-Randall, V. Human primary corneal fibroblasts synthesize and deposit proteoglycans in long-term cultures. Dev. Dyn. 2008, 237, 2705–2715. [Google Scholar] [CrossRef] [PubMed]
- Karamichos, D.; Funderburgh, M.L.; Hutcheon, A.E.; Zieske, J.D.; Du, Y.; Wu, J.; Funderburgh, J.L. A role for topographic cues in the organization of collagenous matrix by corneal fibroblasts and stem cells. PLoS ONE 2014, 9, e86260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petroll, W.M.; Kivanany, P.B.; Hagenasr, D.; Graham, E.K. Corneal fibroblast migration patterns during intrastromal wound healing correlate with ecm structure and alignment. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7352–7361. [Google Scholar] [CrossRef] [PubMed]
- Kivanany, P.B.; Grose, K.C.; Petroll, W.M. Temporal and spatial analysis of stromal cell and extracellular matrix patterning following lamellar keratectomy. Exp. Eye Res. 2016, 153, 56–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynn, B.P.; Bhole, A.P.; Saeidi, N.; Liles, M.; Dimarzio, C.A.; Ruberti, J.W. Mechanical strain stabilizes reconstituted collagen fibrils against enzymatic degradation by mammalian collagenase matrix metalloproteinase 8 (mmp-8). PLoS ONE 2010, 5, e12337. [Google Scholar] [CrossRef] [PubMed]
- Bhole, A.P.; Flynn, B.P.; Liles, M.; Saeidi, N.; Dimarzio, C.A.; Ruberti, J.W. Mechanical strain enhances survivability of collagen micronetworks in the presence of collagenase: Implications for load-bearing matrix growth and stability. Philos. Trans. A Math. Phys. Eng. Sci. 2009, 367, 3339–3362. [Google Scholar] [CrossRef] [PubMed]
- Grinnell, F.; Petroll, W.M. Cell motility and mechanics in three-dimensional collagen matrices. Annu. Rev. Cell Dev. Biol. 2010, 26, 335–361. [Google Scholar] [CrossRef] [PubMed]
- Karamichos, D.; Brown, R.A.; Mudera, V. Collagen stiffness regulates cellular contraction and matrix remodeling gene expresssion. J. Biomed. Mater. Res. A 2007, 83, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Lanfer, B.; Freudenberg, U.; Zimmermann, R.; Stamov, D.; Korber, V.; Werner, C. Aligned fibrillar collagen matrices obtained by shear flow deposition. Biomaterials 2008, 29, 3888–3895. [Google Scholar] [CrossRef] [PubMed]
- Saeidi, N.; Sander, E.A.; Ruberti, J.W. Dynamic shear-influenced collagen self-assembly. Biomaterials 2009, 30, 6581–6592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pot, S.A.; Liliensiek, S.J.; Myrna, K.E.; Bentley, E.; Jester, J.V.; Nealey, P.F.; Murphy, C.J. Nanoscale topography–induced modulation of fundamental cell behaviors of rabbit corneal keratocytes, fibroblasts, and myofibroblasts. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1373–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phu, D.; Wray, L.S.; Warren, R.V.; Haskell, R.C.; Orwin, E.J. Effect of substrate composition and alignment on corneal cell phenotype. Tissue Eng. Part. A 2011, 17, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Dreier, B.; Thomasy, S.M.; Mendonsa, R.; Raghunathan, V.K.; Russell, P.; Murphy, C.J. Substratum compliance modulates corneal fibroblast to myofibroblast transformation. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5901–5907. [Google Scholar] [CrossRef] [PubMed]
- Petroll, W.M.; Ma, L. Direct, dynamic assessment of cell-matrix interactions inside fibrillar collagen lattices. Cell Motil. Cytoskel. 2003, 55, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Petroll, W.M.; Ma, L.; Kim, A.; Ly, L.; Vishwanath, M. Dynamic assessment of fibroblast mechanical activity during rac-induced cell spreading in 3-D culture. J. Cell. Physiol. 2008, 217, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Miron-Mendoza, M.; Graham, E.; Manohar, S.; Petroll, W.M. Fibroblast-fibronectin patterning and network formation in 3D fibrin matrices. Matrix Biol. 2017, 64, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Petroll, W.M.; Ma, L.; Jester, J.V. Direct correlation of collagen matrix deformation with focal adhesion dynamics in living corneal fibroblasts. J. Cell Sci. 2003, 116, 1481–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimp, E.A.; Alsmadi, N.Z.; Cheng, T.; Lam, K.H.; Lewis, C.S.; Schmidtke, D.W. Effects of shear on p-selectin deposition in microfluidic channels. Biomicrofluidics 2016, 10, 024128. [Google Scholar] [CrossRef] [PubMed]
- Miron-Mendoza, M.; Graham, E.; Kivanany, P.; Quiring, J.; Petroll, W.M. The role of thrombin and cell contractility in regulating clustering and collective migration of corneal fibroblasts in different ecm environments. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2079–2090. [Google Scholar] [CrossRef] [PubMed]
- Lakshman, N.; Kim, A.; Petroll, W.M. Characterization of corneal keratocyte morphology and mechanical activity within 3-D collagen matrices. Exp. Eye Res. 2010, 90, 350–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jester, J.V.; Barry, P.A.; Lind, G.J.; Petroll, W.M.; Garana, R.; Cavanagh, H.D. Corneal keratocytes: In situ and in vitro organization of cytoskeletal contractile proteins. Investig. Ophthalmol. Vis. Sci. 1994, 35, 730–743. [Google Scholar]
- Lakshman, N.; Petroll, W.M. Growth factor regulation of corneal keratocyte mechanical phenotypes in 3-D collagen matrices. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Lakshman, N.; Petroll, W.M. Quantitative assessment of local collagen matrix remodeling in 3-D culture: The role of Rho kinase. Exp. Cell Res. 2006, 312, 3683–3692. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Petroll, W.M. Rho kinase regulation of fibroblast migratory mechanics in fibrillar collagen matrices. Cell. Mol. Bioeng. 2010, 3, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Petroll, W.M. Mmp regulation of corneal keratocyte motility and mechanics in 3-D collagen matrices. Exp. Eye Res. 2014, 121, 147–160. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kivanany, P.B.; Grose, K.C.; Yonet-Tanyeri, N.; Manohar, S.; Sunkara, Y.; Lam, K.H.; Schmidtke, D.W.; Varner, V.D.; Petroll, W.M. An In Vitro Model for Assessing Corneal Keratocyte Spreading and Migration on Aligned Fibrillar Collagen. J. Funct. Biomater. 2018, 9, 54. https://doi.org/10.3390/jfb9040054
Kivanany PB, Grose KC, Yonet-Tanyeri N, Manohar S, Sunkara Y, Lam KH, Schmidtke DW, Varner VD, Petroll WM. An In Vitro Model for Assessing Corneal Keratocyte Spreading and Migration on Aligned Fibrillar Collagen. Journal of Functional Biomaterials. 2018; 9(4):54. https://doi.org/10.3390/jfb9040054
Chicago/Turabian StyleKivanany, Pouriska B., Kyle C. Grose, Nihan Yonet-Tanyeri, Sujal Manohar, Yukta Sunkara, Kevin H. Lam, David W. Schmidtke, Victor D. Varner, and W. Matthew Petroll. 2018. "An In Vitro Model for Assessing Corneal Keratocyte Spreading and Migration on Aligned Fibrillar Collagen" Journal of Functional Biomaterials 9, no. 4: 54. https://doi.org/10.3390/jfb9040054