Protein Degradation and Protection Observed in the Presence of Novel Wound Dressing Components

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Sample Preparation
2.2. Sample Testing
2.3. Statistical Methods
3. Results and Discussion
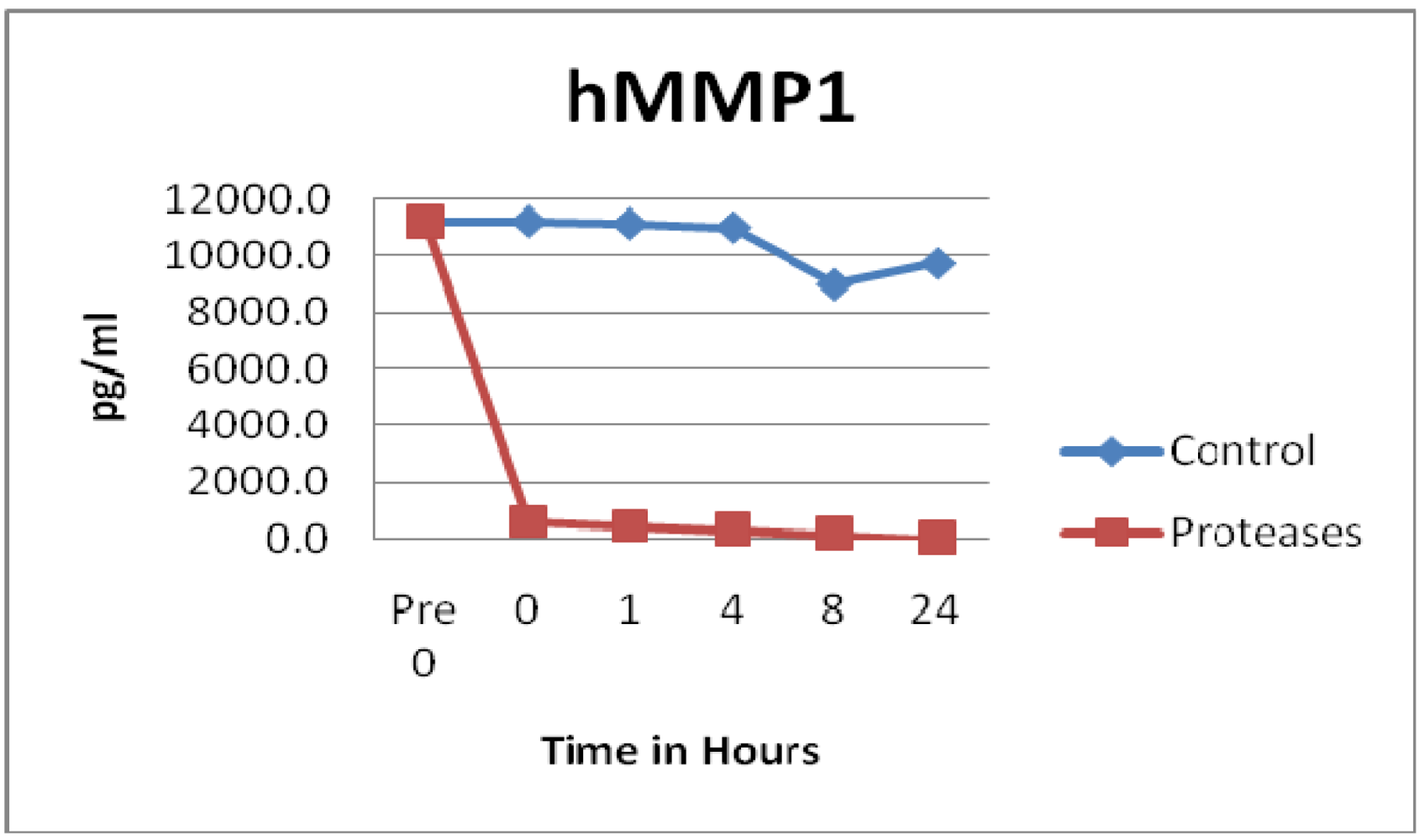
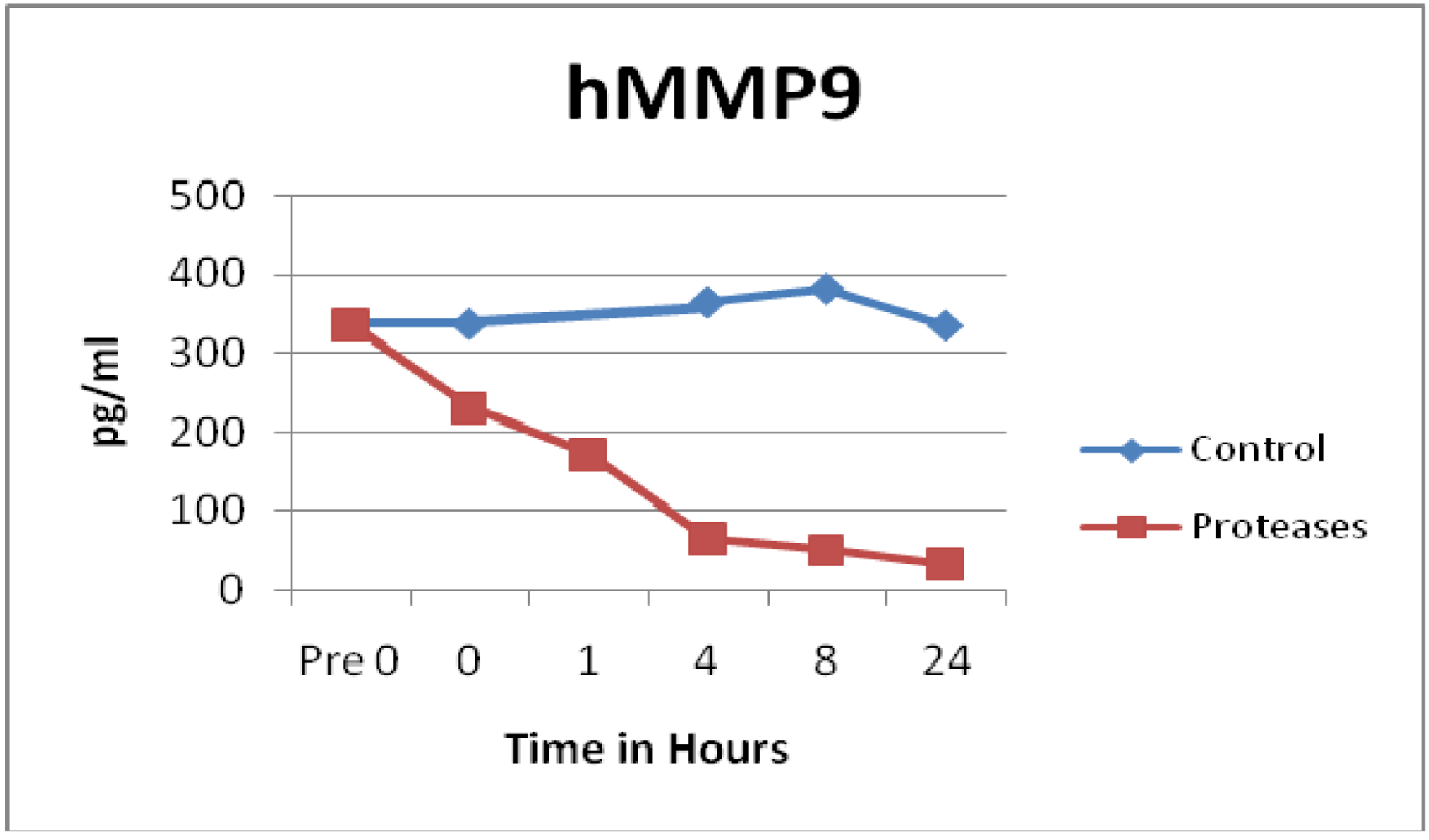
3.1. Matrix Metalloproteinases (MMPs) and Tissue Inhibitor of Matrix Metalloproteinases (TIMPs)
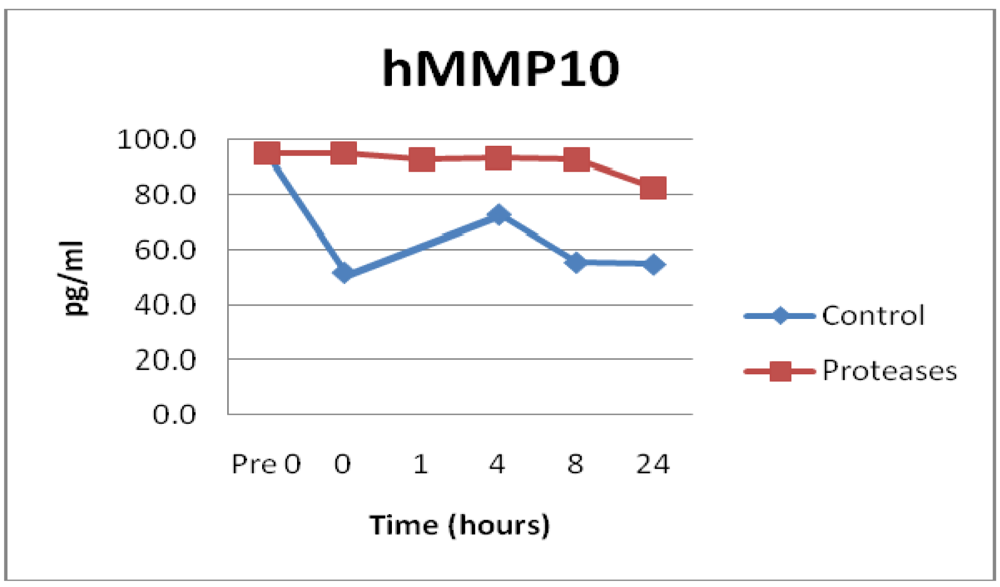

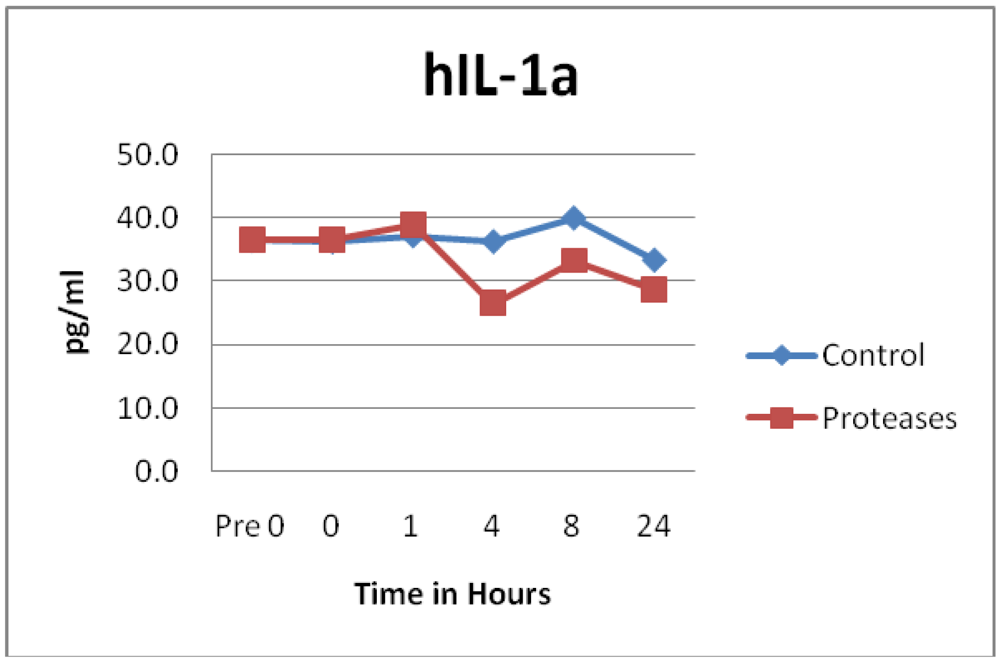
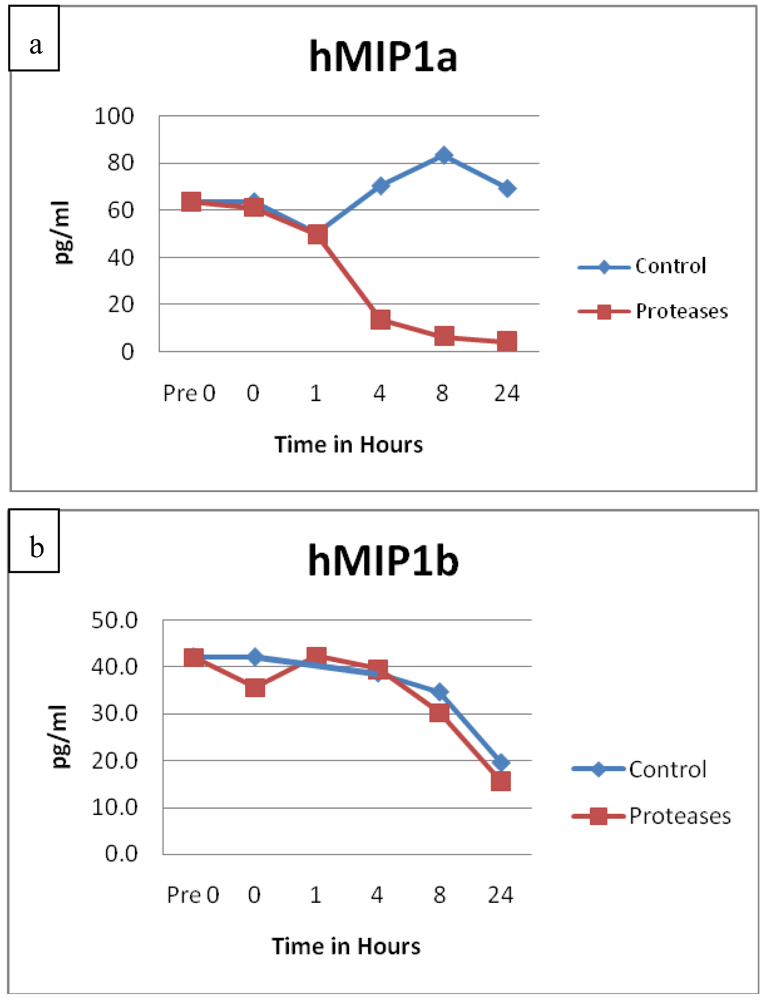
3.2. Cytokines and Chemokines


3.3. Growth Factors
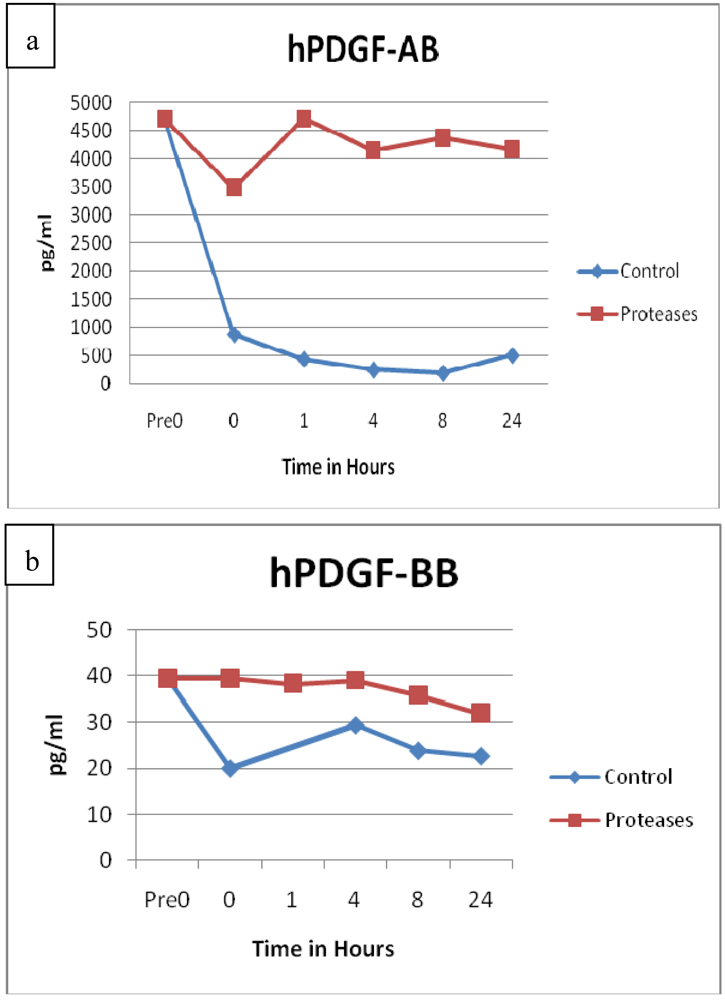
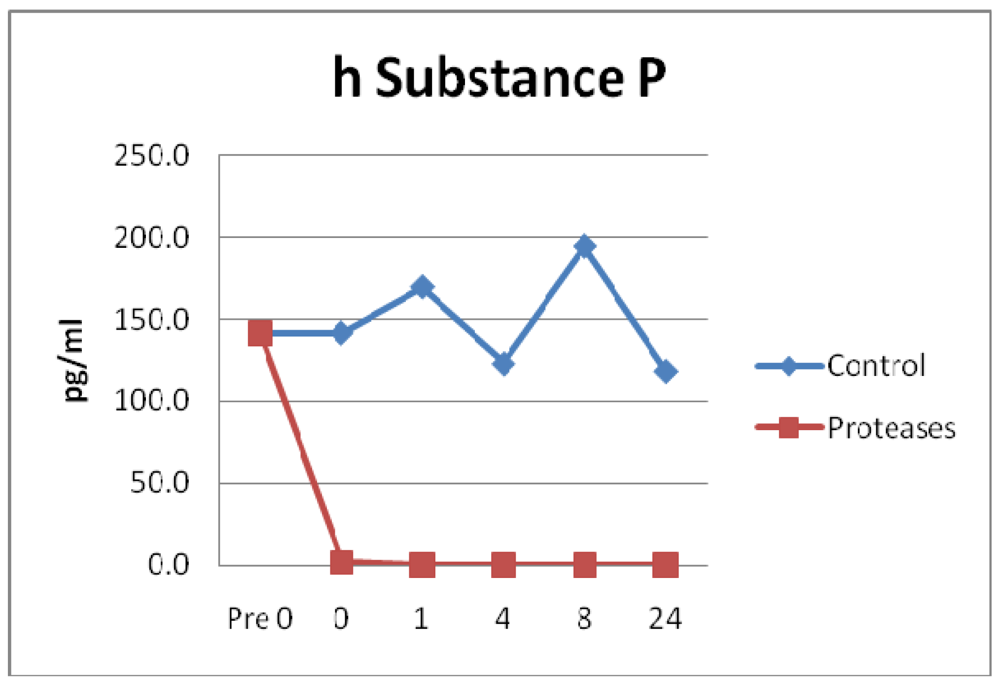
3.4. Neuropeptides
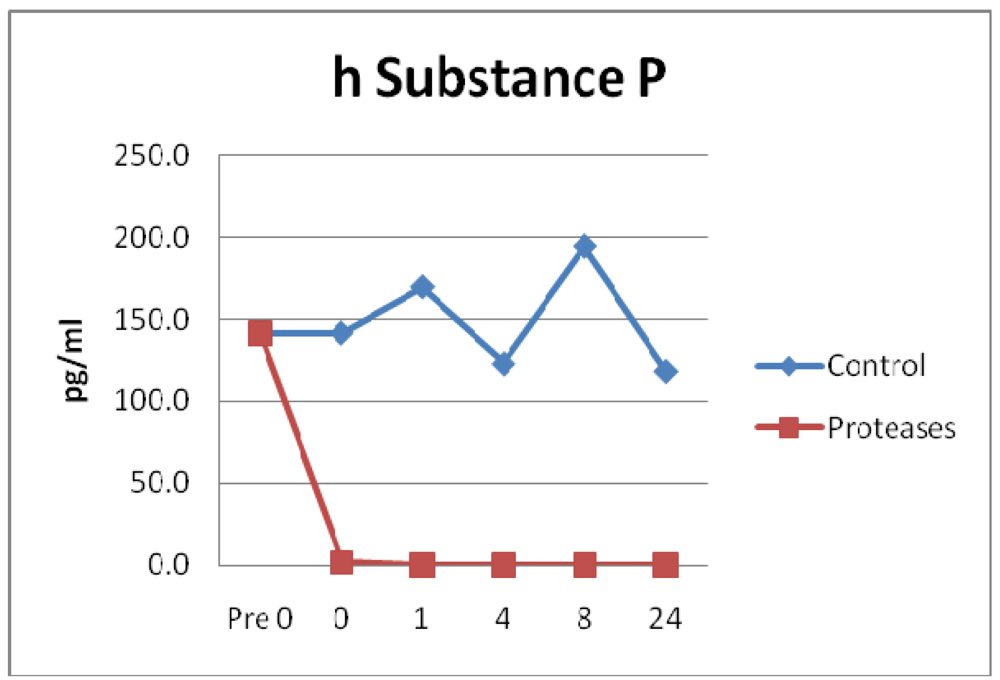
3.5. Results Summary
3.6. Discussion

4. Conclusions
Conflict of Interest
Acknowledgments
References
- Winter, G.D. Formation of scab and the rate of epithelialization of superficial wounds in the skin of the domestic pig. Nature 1962, 193, 293–294. [Google Scholar]
- Mahendale, F.; Martin, P. The cellular and molecular events of wound healing. In Cutaneous Wound Healing, 1st ed.; Falanga, V., Ed.; Martin Dunitz Ltd.: Florence, KY, USA, 2001; pp. 15–37. [Google Scholar]
- Mast, B.A. The skin. In Wound Healing-Biochemical and Clinical Aspects, 1st ed.; Cohen, I.K., Diegelmann, R.F., Lindblad, W.J., Eds.; W.B. Saunders Company, Harcourt Brace Jovanovich Inc.: Philadelphia, PA, USA, 1992; pp. 344–355. [Google Scholar]
- Efron, P.A.; Moldawer, L.L. Cytokines and wound healing: The role of cytokines and anticytokine therapy in the repair response. J. Burn Care Rehabil. 2004, 25, 149–160. [Google Scholar]
- Grotendorst, G.R. Chemoattractants and growth factors. In Wound Healing-Biochemical and Clinical Aspects, 1st ed.; Cohen, I.K., Diegelmann, R.F., Lindblad, W.J., Eds.; W.B. Saunders Company, Harcourt Brace Jovanovich Inc.: Philadelphia, PA, USA, 1992; pp. 237–247. [Google Scholar]
- Barbul, A. Role of the immune system. In Wound Healing-Biochemical and Clinical Aspects, 1st ed.; Cohen, I.K., Diegelmann, R.F., Lindblad, W.J., Eds.; W.B. Saunders Company, Harcourt Brace Jovanovich Inc.: Philadelphia, PA, USA, 1992; pp. 282–291. [Google Scholar]
- Robson, M.C.; Heggers, J.P. Eicosanoids, cytokines, and free radicals. In Wound Healing-Biochemical and Clinical Aspects, 1st ed.; Cohen, I.K., Diegelmann, R.F., Lindblad, W.J., Eds.; W.B. Saunders Company, Harcourt Brace Jovanovich Inc.: Philadelphia, PA, USA, 1992; pp. 2292–2304. [Google Scholar]
- Naldini, A.; Carraro, F. Role of inflammatory mediators in angiogenesis. Curr. Drug Targets Inflamm. Allergy 2005, 4, 3–8. [Google Scholar]
- Falanga, V.; Shen, J. Growth factors, signal transduction and cellular responses. In Cutaneous Wound Healing, 1st ed.; Falanga, V., Ed.; Martin Dunitz Ltd.: Florence, KY, USA, 2001; pp. 81–93. [Google Scholar]
- Grose, R.; Werner, S. Wound-healing studies in transgenic and knockout mice. Mol. Biotechnol. 2004, 28, 147–166. [Google Scholar]
- Harding, K.G.; Morris, H.L.; Patel, G.K. Science, medicine and the future: Healing chronic wounds. BMJ 2002, 324, 160–163. [Google Scholar]
- Plowden, J.; Renshaw-Hoelscher, M.; Engleman, C.; Katz, J; Sambhara, S. Innate immunity in aging: Impact on macrophage function. Aging Cell 2004, 3, 161–167. [Google Scholar]
- Di, C.T.; Wang, L.; Wille, J. Epidermal expression of collagenase delays wound-healing in transgenic mice. J. Invest. Dermatol. 1998, 111, 1029–1033. [Google Scholar]
- Romer, J.; Bugge, T.H.; Pyke, C. Impaired wound healing in mice with a disrupted plasminogen gene. Cell 1996, 2, 287–292. [Google Scholar]
- Bugge, T.H.; Flick, M.J.; Danton, M.J.; Daugherty, C.C.; Romer, J.; Dano, K.; Carmeliet, P.; Collen, D.; Degen, J.L. Urokinase-type plasminogen activator is effective in fibrin clearance in the absence of its receptor or tissue-type plasminogen activator. Proc. Natl. Acad. Sci. USA 1996, 93, 5899–5904. [Google Scholar]
- Mast, B.A.; Schultz, G.S. Interactions of cytokines, growth factors, and proteases in acute and chronic wound fluid. Wound Repair Regen. 1996, 4, 411–420. [Google Scholar]
- Tarnuzzer, R.W.; Schultz, G.S. Biochemical analysis of acute and chronic wound environments. Wound Repair Regen. 1996, 4, 321–325. [Google Scholar]
- Trengove, N.J.; Stacey, M.C.; MacAuley, S.; Bennett, N.; Gibson, J.; Burslem, F.; Murphy, G.; Schultz, G. Analysis of the acute and chronic wound environments: The role of proteases and their inhibitors. Wound Repair Regen. 1999, 7, 442–452. [Google Scholar]
- Trengove, N.J.; Langton, S.R.; Stacey, M.C. Biochemical analysis of wound fluid from nonhealing and healing chronic leg ulcers. Wound Repair Regen. 1996, 4, 234–239. [Google Scholar]
- Itoh, Y.; Nagase, H. Preferential inactivation of tissue inhibitor of metalloproteinases-1 that is bound to the precursor of matrix metalloproteinase 9 (progelatinase B) by human neutrophil elastase. J. Biol. Chem. 1995, 270, 16618–16621. [Google Scholar]
- Nwomeh, B.C.; Yager, D.R.; Cohen, I.K. Physiology of the chronic wound. Clin. Plast. Surg. 1998, 25, 341–356. [Google Scholar]
- European Wound Management Association (EWMA). Position Document: Wound Bed Preparation in Practice; MEP Ltd.: London, UK, 2004. [Google Scholar]
- Steed, D.L. The diabetic ulcer study group. Clinical evaluation of recombinant human platelet derived growth factor for the treatment of lower extremity diabetic ulcers. J. Vasc. Surg. 1995, 21, 71–81. [Google Scholar]
- Cullen, B.; Smith, R.; McCulloch, E.; Silcock, D.; Morrison, L. Mechanism of action of PROMOGRAN, a protease modulating matrix, for the treatment of diabetic foot ulcers. Wound Repair Regen. 2002, 10, 16–25. [Google Scholar]
- Fuller, G.C.; Cutroneo, K.R. Pharmacological interventions. In Wound Healing-Biochemical and Clinical Aspects, 1st ed.; Cohen, I.K., Diegelmann, R.F., Lindblad, W.J., Eds.; W.B. Saunders Company, Harcourt Brace Jovanovich Inc: Philadelphia, PA, USA, 1992; pp. 305–315. [Google Scholar]
- Gosain, A.; Gamelli, R.L. A primer in cytokines. J. Burn Care Rehabil. 2005, 26, 7–12. [Google Scholar]
- Baskovich, B.; Sampson, E.M.; Schultz, G.S.; Parnell, L.K.S. Wound dressing components degrade proteins detrimental to wound healing. Int. Wound J. 2008, 5, 543–551. [Google Scholar]
- Metze, D. Skin nerve anatomy: Neuropeptide distribution and its relationship to itch. In Itch Basic Mechanisms and Therapy, 1st ed.; Yosipovitch, G., Greaves, M.W., Fleischer, A.B., Jr., McGlone, F., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 2004; pp. 71–86. [Google Scholar]
- Andoh, T.; Kuraishi, Y. Substance P and itch. In Itch Basic Mechanisms and Therapy, 1st ed.; Yosipovitch, G., Greaves, M.W., Fleischer, A.B., Jr., McGlone, F., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 2004; pp. 87–95. [Google Scholar]
- Yager, D.R.; Chen, S.M.; Ward, S.; Olutoye, O.O.; Diegelmann, R.F.; Cohen, I.K. The ability of chronic wound fluids to degrade peptide growth factors is associated with increased levels of elastase activity and diminished levels of proteinase inhibitors. Wound Repair Regen. 1997, 5, 23–32. [Google Scholar]
- Barnett, L.; Parnell, L.K.S. Contact dermatitis treated with new topical products: A case study. Ostomy Wound Manag. 2001, 47, 47–53. [Google Scholar]
- Brinton, A.M.; Parnell, L.K.S. Use of a protease containing antifungal to treat damaged and contaminated perineal skin. Presented at the Symposium on Advances in Wound Care, Orlando, FL, USA, 17–20 April 2007.
- Parnell, L.K.S.; Ciufi, B.; Gokoo, C.F. Preliminary use of a hydrogel containing enzymes in the treatment of stage II and stage III pressure ulcers. Ostomy Wound Manag. 2005, 51, 50–60. [Google Scholar]
- Nedelec, B.; Rachelska, G.; Parnell, L.K.S.; LaSalle, L. Double-blind, randomized, pilot study assessing the resolution of post-burn pruritus. J. Burn Care Res. 2011, in press. [Google Scholar]
- Wright, J.B.; Shi, L. Accuzyme papain-urea debriding ointment: A historical review. Wounds 2002, 15, 2S–12S. [Google Scholar]
- Smith, R.G. Enzymatic debriding agents: An evaluation of the medical literature. Ostomy Wound Manag. 2008, 54, 16–34. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Parnell, L.K.S. Protein Degradation and Protection Observed in the Presence of Novel Wound Dressing Components. J. Funct. Biomater. 2011, 2, 338-354. https://doi.org/10.3390/jfb2040338
Parnell LKS. Protein Degradation and Protection Observed in the Presence of Novel Wound Dressing Components. Journal of Functional Biomaterials. 2011; 2(4):338-354. https://doi.org/10.3390/jfb2040338
Chicago/Turabian StyleParnell, Laura K.S. 2011. "Protein Degradation and Protection Observed in the Presence of Novel Wound Dressing Components" Journal of Functional Biomaterials 2, no. 4: 338-354. https://doi.org/10.3390/jfb2040338