Effects of Contingency versus Constraints on the Body-Mass Scaling of Metabolic Rate


Department of Biology, Juniata College, Huntingdon, PA 16652, USA
Challenges 2018, 9(1), 4; https://doi.org/10.3390/challe9010004
Submission received: 16 December 2017
/
Revised: 16 January 2018
/
Accepted: 17 January 2018
/
Published: 29 January 2018
Abstract
:I illustrate the effects of both contingency and constraints on the body-mass scaling of metabolic rate by analyzing the significantly different influences of ambient temperature (Ta) on metabolic scaling in ectothermic versus endothermic animals. Interspecific comparisons show that increasing Ta results in decreasing metabolic scaling slopes in ectotherms, but increasing slopes in endotherms, a pattern uniquely predicted by the metabolic-level boundaries hypothesis, as amended to include effects of the scaling of thermal conductance in endotherms outside their thermoneutral zone. No other published theoretical model explicitly predicts this striking variation in metabolic scaling, which I explain in terms of contingent effects of Ta and thermoregulatory strategy in the context of physical and geometric constraints related to the scaling of surface area, volume, and heat flow across surfaces. My analysis shows that theoretical models focused on an ideal 3/4-power law, as explained by a single universally applicable mechanism, are clearly inadequate for explaining the diversity and environmental sensitivity of metabolic scaling. An important challenge is to develop a theory of metabolic scaling that recognizes the contingent effects of multiple mechanisms that are modulated by several extrinsic and intrinsic factors within specified constraints.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
The sizes of living things vary enormously, which has major consequences for how they look, act, and function. How biological traits vary with body size is the subject of an exciting and controversial field called “allometry” or “biological scaling” [1,2,3,4,5,6,7,8]. This field of study is important because it has great potential for contributing to a unifying theory of biological systems [9,10]. All living systems have size and most of their structural and functional features scale to size. Moreover, these scaling relationships are often not simple proportional (1:1 or isometric) relationships, but may involve diverse kinds of disproportionate (allometric) relationships. Although these various kinds of scaling relationships have been the subject of decades of research, they remain little understood.
A classic example is the scaling of the rate of metabolism (i.e., the transformation of energy and materials in support of various biological processes). Early theoretical and empirical studies in the 1800s suggested that the scaling of metabolic rate in endothermic (warm-blooded) birds and mammals obeys a 2/3-power law [11,12]. Maintenance of a constant, high body temperature requires metabolic heat production to balance exactly the loss of heat through the body surface, which scales to the 2/3 power in relation to body mass or volume (assuming isomorphic body shapes). Therefore, metabolic rate should also scale to the 2/3 power, as observed within many species of birds and mammals ([3,12,13,14,15], but see [16]). However, in the 1930s, animal scientists such as Kleiber and Brody discovered that the interspecific scaling of basal (resting) metabolic rate in mammals more closely matched a 3/4-power relationship than a 2/3-power relationship [17,18]. In 1960, Hemmingsen [19] reported 3/4-power metabolic scaling relationships for not only endothermic birds and mammals, but also various ectothermic (cold-blooded) animals and even unicellular organisms. This finding led to the proclamation of a universal 3/4-power scaling law for metabolic rate (also called Kleiber’s Law) that most biologists accepted [4,5,6,20,21] until quite recently [10,16,22]. A belief in the 3/4-power law also spurred a continuing search for intrinsic physical causes of metabolic scaling. If such a universal law existed, contingent biological and ecological factors would not be important in explaining it. As a universal law, it by definition must operate in all organisms and in all environments. Taxonomic and ecological variation should be irrelevant, or at most secondary in importance. In addition, the physical explanation could not be a simple function of heat loss through body surfaces because the power of the scaling relationship is allegedly 3/4 and not 2/3. Moreover, this law reputedly applied to organisms that did not maintain constant body temperatures, and thus need not balance their metabolic heat production with heat loss. In short, the once conventional thermoregulatory model of metabolic scaling appeared to be inadequate, at least as a general explanation, though it may still be applicable to endotherms, at least in part, as argued here and in [23,24,25,26,27,28].
Many investigators have attempted to provide a universal, intrinsic, physical explanation of 3/4-power metabolic scaling, which has been called the “Holy Grail in comparative biology” (Carol Beuchat cited in [29]), but none of these explanations enjoy wide acceptance (reviewed in [10,16,22,30,31]). The lack of a consensus view results from two major problems. First, recent work has made it clear that no universal 3/4-power scaling law for metabolic rate actually exists. The scaling slopes of metabolic rate vary from ~0 to >1 (but mostly between 2/3 and 1) in various kinds of organisms under various kinds of conditions [10,16,22]. Second, all of the proposed models are limited in scope, and thus not universally applicable, and (or) have other problems with their assumptions, internal logic and (or) predictions [10,16,22,30,31,32].
Consider, for example, the most influential of the metabolic scaling explanations based on intrinsic physical constraints published to date. According to this resource-transport-network (RTN) model [33], and others like it [34,35,36,37], the classical 3/4- or 2/3-power scaling of metabolic rate results from increased physical and geometric constraints on the supply of resources to cells in large organisms with longer transport distances, on average, than those of smaller organisms. However, RTN models have received extensive criticism [10,16,22,30,31,38]. First, they are not universally applicable because they do not apply to organisms without closed resource-transport (e.g., circulatory) systems. Second, several lines of evidence contradict the fundamental assumption of RTN models that resource supply limits resting metabolic rates in organisms [16,27,30,38]. For example, in various kinds of animals, cells of large species are not more oxygen limited than are cells of smaller species [38]. In addition, avian and mammalian cells grown in nutrient- and oxygen rich culture media often exhibit lower rather than higher metabolic rates compared to cells in vivo, contrary to predictions of RTN models [27]. Third, scaling slopes of metabolic rate in pelagic invertebrates that grow mainly in two dimensions more closely match those predicted by surface-area models (slope ~ 1) than those predicted by RTN models (slopes ~ 1/2, 5/8 or 2/3, depending on the type of network) [39,40]. Fourth, intrinsic physical explanations, such as those of RTN models, cannot explain the extensive variation of metabolic scaling systematically associated with various biological and ecological factors.
Several recent studies have shown that many kinds of extrinsic environmental factors affect the scaling of metabolic rate [10,16,22,32]. These include abiotic factors, such as temperature, pH and oxygen availability [10,16,26,41], and biotic factors, such as predation, parasitism and food quantity and quality [10,16,42,43,44,45]. Metabolic scaling relationships also vary with lifestyle, developmental stage and physiological status [10,16,23,26,46,47,48]. As a result, it has become increasingly clear that biological and ecological contingency plays an important role in metabolic scaling. A major challenge now is to determine the relative influences that contingency versus constraints have on metabolic scaling. Some investigators still promote the view that internal constraints predominate, especially the proponents of RTN models [8,9,21,33,34,35,36,37] and the influential metabolic theory of ecology [49]. Others claim that heterogeneous contingent factors are importantly involved in determining both the slopes and elevations of metabolic scaling relationships [10,16,23,32,50,51,52].
As a point of departure for my discussion about the contingency versus constraints debate, I focus on published data showing that an extrinsic factor, namely ambient temperature, not only significantly affects the scaling of metabolic rate, but also does so in fundamentally different ways in ectothermic and endothermic animals. This comparison allows for not only a fresh assessment of the relative roles of contingency and constraints in metabolic scaling, but also a useful comparative evaluation of several prominent theoretical models. I conclude with a brief discussion about what future research is required to advance our understanding of metabolic scaling.
2. Results
3. Discussion
3.1. Implications of Results for Theory
The results just presented are important and challenging, because they cannot be explained completely by any published model of metabolic scaling. RTN models are clearly inadequate. These models either ignore the possibility of temperature effects [33,34,35,36,37], or assume that body size and temperature act independently on metabolic rate [49,60]. According to the metabolic theory of ecology (MTE) [8,49], which includes the RTN model of [33], temperature should affect the elevation, but not the slope (exponent) of metabolic scaling relationships. The present results clearly contradict this prediction.
Increasing temperature tends to be associated with smaller cell sizes in a variety of organisms (reviewed in [61,62]; also see [63,64,65,66,67,68]). Small cells (and organisms containing them) tend to have higher mass-specific metabolic rates than large cells (and organisms containing them) [26,69,70,71,72,73,74,75,76,77]. Therefore, like the MTE, cell-size theory predicts that changing temperature should affect the elevation of metabolic scaling relationships, but not their slopes (but see Section 3.2.2), which again is contradicted by the results described in this study. Most other models focused on the differential scaling of various resource-demanding processes and tissues or organs with different metabolic rates (see e.g., [10,38,52]) have yet to consider how ambient temperature may affect metabolic scaling slopes (but see discussion of dynamic energy budget theory below). Furthermore, a recent study has shown that temperature acclimation does not affect the size and metabolic rate of various organs of the prawn Macrobrachium tolmerum [78].
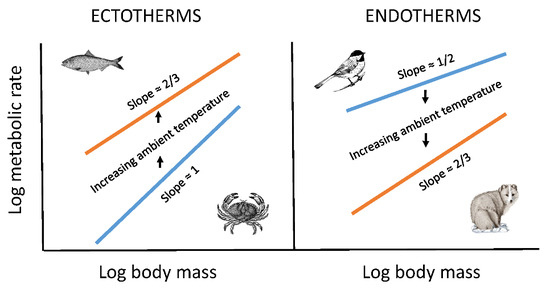
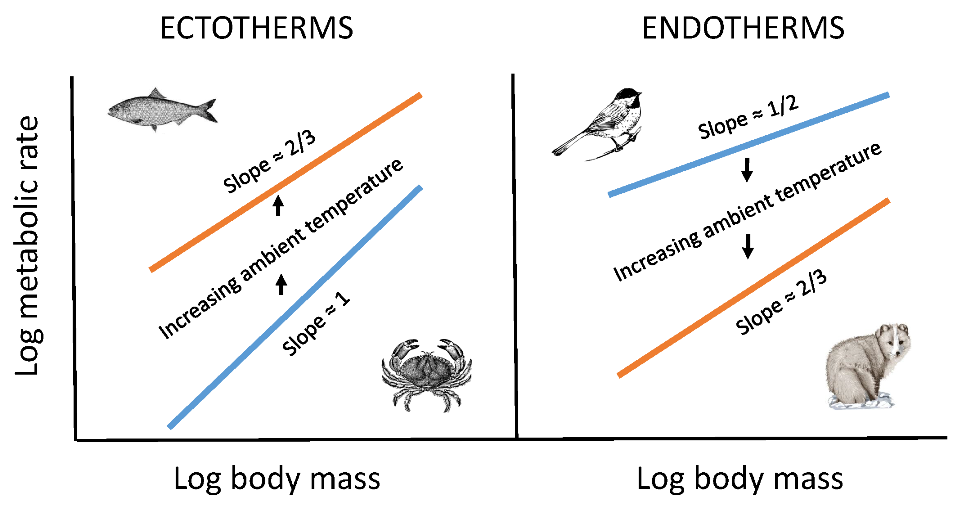
The model that best explains the observed results is the metabolic–level boundaries hypothesis (MLBH) [23,26]. It predicts that changes in temperature should affect both the elevation and slope of metabolic scaling relationships [26], as observed in this study and several other studies (e.g., [16,26,48,53]; also see Section 3.2.2). Moreover, it uniquely predicts that increasing temperature should result in decreases in the metabolic scaling exponent in ectotherms (Figure 1 and Figure 2), but increases in the exponent in endotherms (Figure 3 and Figure 4), again as observed. This is because it predicts inverse relationships between the slopes and elevations of scaling relationships for resting metabolic rate. Therefore, since decreasing temperature decreases metabolic level (scaling elevation) in ectotherms, but increases it in endotherms because of increased metabolic heat production required to maintain a constant body temperature, the opposite responses of scaling slope to temperature in ectotherms and endotherms are as expected. According to the MLBH, at low metabolic levels, volume-related tissue maintenance chiefly influences metabolic scaling (scaling slope approaching 1 in organisms with uniform scaling of metabolic rate in different tissues), whereas at high metabolic levels, surface-area-related processes (such as resource uptake, metabolic waste removal, and heat dissipation) predominate (scaling slope approaching 2/3 in isomorphic organisms). In short, the scaling exponent for resting metabolic rate should vary between 2/3 and 1, depending on metabolic level [23,26].
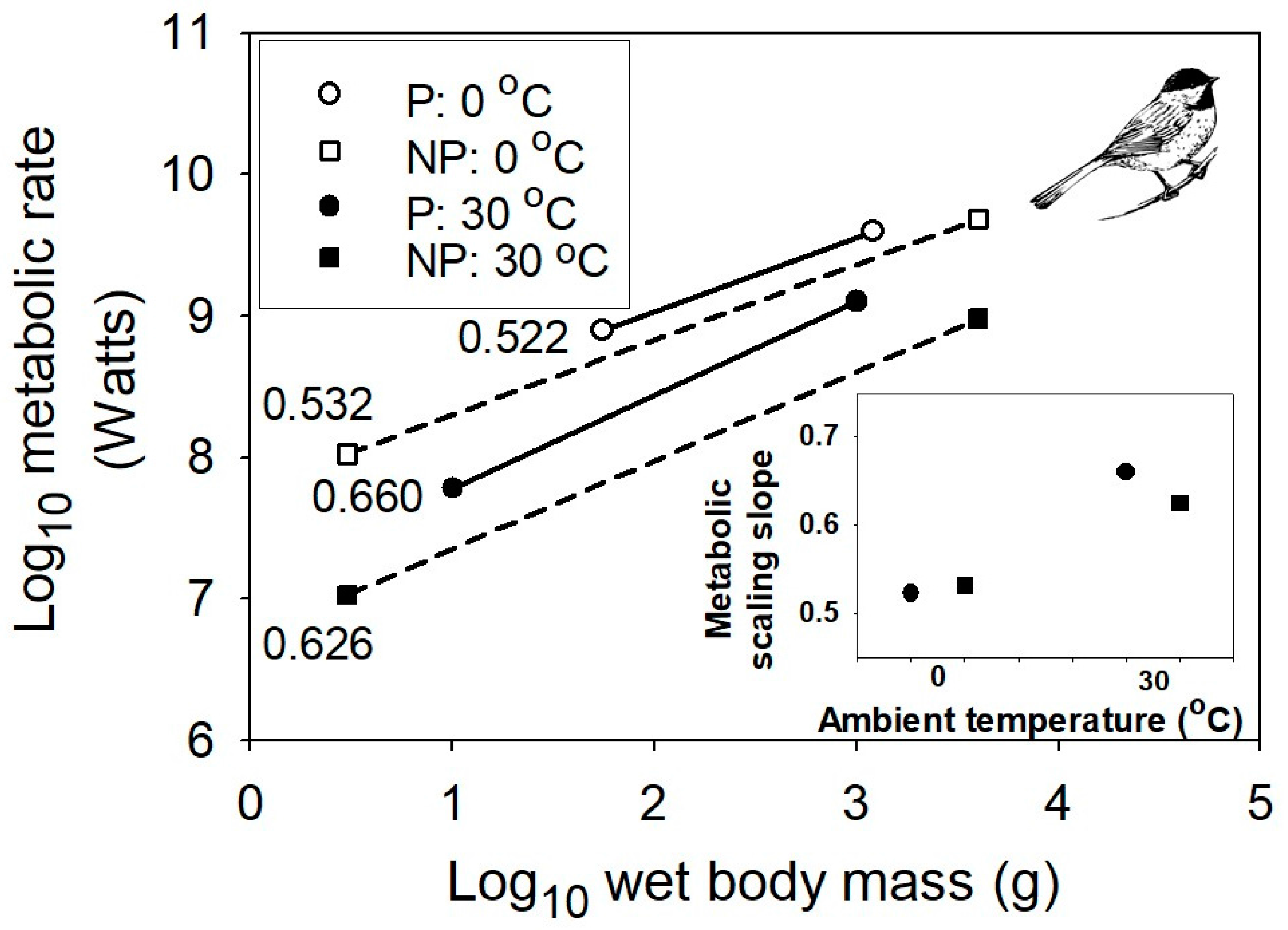
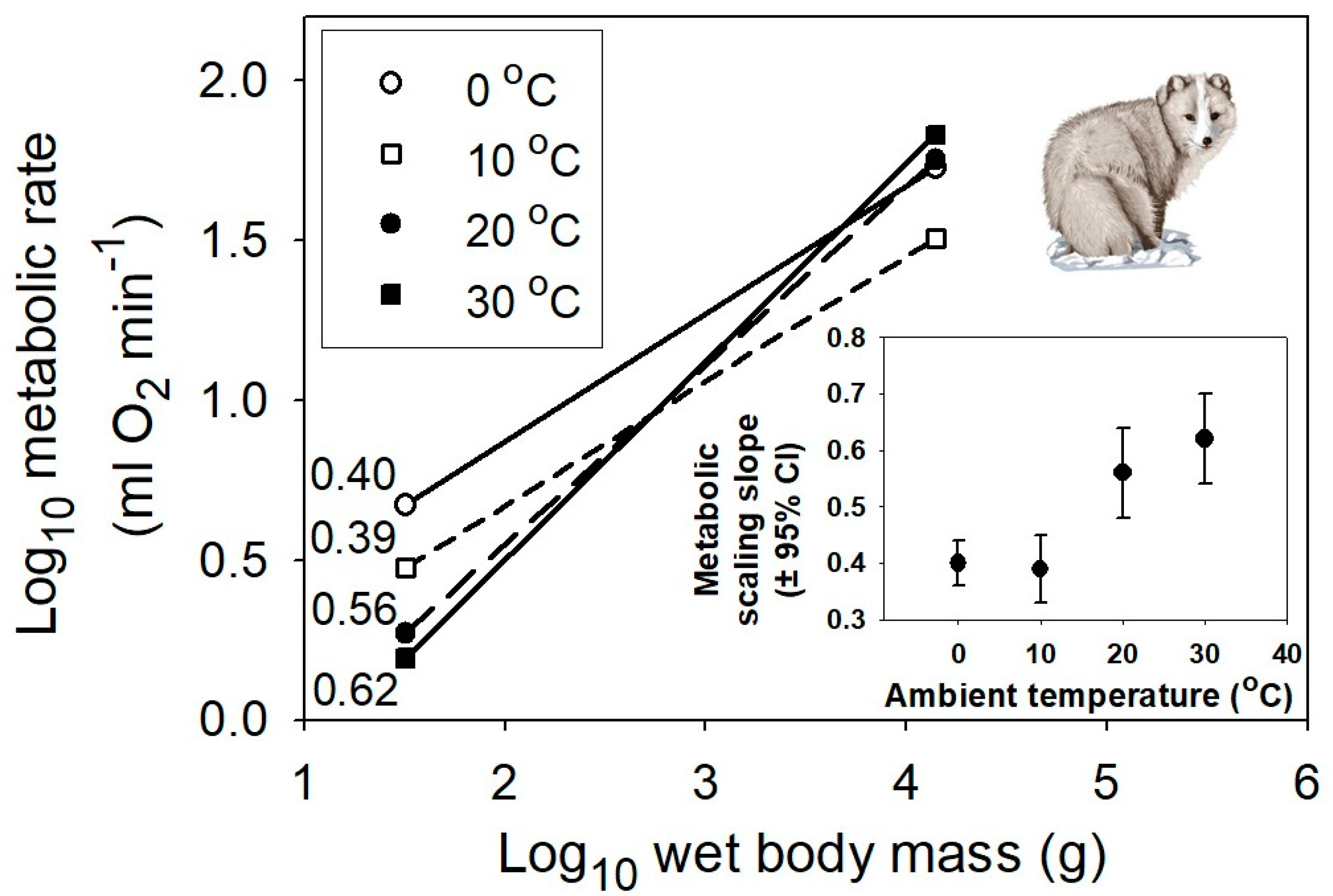
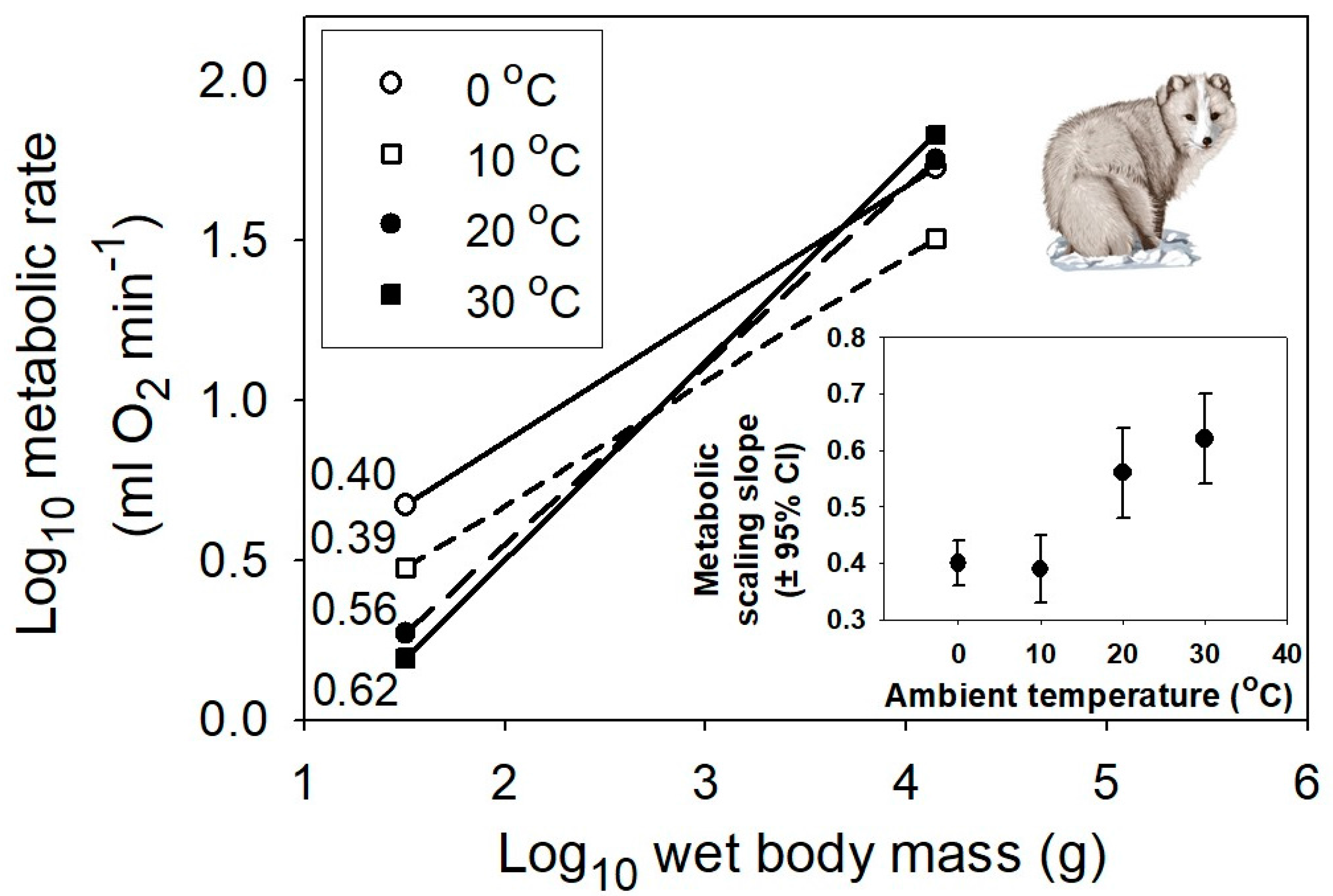
However, even the MLBH, as currently formulated, falls short of completely explaining the effects of temperature on the metabolic scaling of endotherms. In both birds and mammals, as temperature decreases, the metabolic scaling exponent falls below the lower limit of 2/3 predicted by the MLBH (see Figure 3 and Figure 4). Why? A likely answer involves an increasing influence of the body-mass scaling of thermal conductance on the scaling of metabolic rate, as ambient temperature (Ta) declines, and thus becomes increasingly different from the relatively high body temperature (Tb) maintained by endotherms. Following Fourier’s Law (which is related to Newton’s law of cooling), rate of heat flow across an organism’s surface is a function of four major factors: surface area, thickness and thermal conductivity of the surface layer of insulation, and the temperature differential (Tb – Ta) between the inside and outside of an organism [25,56]. At thermoneutrality, which is near 30 °C in most endothermic vertebrates, the temperature differential is relatively small, and the cost of thermoregulation is minimal, thus causing the metabolic scaling exponent to approximate 2/3, which follows the classic surface law [11,12], as pointed out by [56]. Within the thermoneutral zone, metabolic heat production exactly balances the heat dissipated, which is chiefly a function of surface area. However, as the cross-surface temperature differential increases, the metabolic scaling slope should approach ~0.45–0.55, as typically observed for the scaling of thermal conductance in birds and mammals [6,79,80,81,82,83,84,85,86]. As predicted, when Ta equals 0 °C (which is greater than 30 °C below Tb), the metabolic scaling exponents for mammals (0.40), passerine birds (0.52) and nonpasserine birds (0.53) ([56,58]; Figure 3 and Figure 4) all approach that observed for the scaling exponent of thermal conductance.
Therefore, the MLBH requires amendment to explain the scaling of resting metabolic rate of endotherms outside their thermoneutral zone. Heat dissipation importantly affects the metabolic scaling of endotherms at all ambient temperatures (also see [23,24,25,26,28,56,58,87]), but in the thermoneutral zone, surface area is chiefly important, whereas at increasingly lower ambient temperatures outside the thermoneutral zone, insulation and the cross-surface temperature differential become increasingly important, as well. As Kendeigh and colleagues argued [56], metabolic heat production that exactly compensates for heat dissipation in the cold should scale approximately to the 0.5 power, or as (M0.167)(M0.667)/(W0.333), which are the hypothesized power relations for the thermal conductivity of the surface layer of insulation (h), surface area (A), and insulation thickness (I), respectively, for endotherms. This dimensional analysis uses a formula for the rate of heat flow across a surface (k) that is based on Fourier’s Law: i.e., k = hA/I(Tb − Ta) (cf. [6,25,28,58]). It also assumes that Tb and body shape are constant and pelage mass increases isometrically with body mass (scaling slope = 1), and ignores the effects of other factors such as radiation and air convection [6,25]. Notably, the hypothesized power relations for h, A and I approximate empirical estimates quite closely [56,83]. Similarly, Schmidt-Nielsen [6] noted that the insulation of mammals scales as M0.17, and thus thermal conductance should scale as (M0.67)/(W0.17) or as M0.50, as approximately observed (cf. [88]). Therefore, the greater insulation of larger mammals causes the scaling slope for heat dissipation at low Ta to approximate 0.5, which in turn requires the scaling slope for metabolic heat production to approximate 0.5, in order to maintain a constant body temperature (cf. [25,89]). In support, maximal non-shivering thermogenesis (induced by noradrenaline injection) scales similarly in mammals (exponent = 0.546 [90]). In addition, if one minimizes the size-related effect of insulation by exposing birds or mammals of different size to low temperatures in a He-O2 atmosphere with high thermal conductivity, surface-area related heat dissipation chiefly influences metabolic scaling, which thus has a scaling slope approximating 2/3 (see [91,92,93]).
I know of only two other theoretical models that predict that the metabolic scaling slope should vary with Ta. According to dynamic energy budget theory, as recently modified by [94,95], negative associations between Ta and the metabolic scaling exponent should occur in some, but not all colonial animals. However, the ability of this model to explain Ta effects on the metabolic scaling of unitary organisms is problematic. It invokes effects of Ta on growth rate, which cannot explain negative associations between Ta and the metabolic scaling exponent commonly observed in ectothermic organisms. Decreasing Ta inhibits growth rate, and decreased growth rates are associated with lower, not higher metabolic scaling exponents (see [10,32,44,96]).
According to the viscosity hypothesis [97], small aquatic animals have greater difficulty engaging in respiratory ventilation and thus oxygen uptake in colder, more viscous water than do larger animals. Consequently, as water temperature decreases, and viscosity increases, smaller animals should exhibit a greater depression of metabolic rate than that of larger animals, thus resulting in a negative association between temperature and the metabolic scaling slope. However, this model is not general, as it applies only to aquatic, actively ventilating, ectothermic animals, and not to plants, terrestrial ectothermic animals, or endothermic animals. Nevertheless, inverse relationships between the metabolic scaling slope and temperature also occur in plants [26,98] and various terrestrial ectothermic animals (e.g., [99,100,101]), as predicted by the MLBH [26]. In addition, the viscosity hypothesis cannot explain why the metabolic scaling exponent decreases with decreasing temperature in endotherms (Figure 3 and Figure 4). Nor can it explain responses of the metabolic scaling exponent to metabolic level related to other factors besides temperature (e.g., taxonomic variation [23,102], lifestyle [23,26,48], and activity state [16,23,26,103,104]), as predicted by the MLBH. Therefore, the MLBH has broader predictive power than the viscosity hypothesis.
3.2. Challenges for Future Research
3.2.1. General Perspective
The results of this study clearly show that both contingency and constraints affect metabolic scaling. The contingencies considered here include Ta and thermoregulatory lifestyle (ectothermy versus endothermy). The metabolic scaling slope varies with Ta, and the nature of this variation differs markedly between ectotherms and endotherms (Figure 1, Figure 2, Figure 3 and Figure 4). One cannot understand this variation in metabolic scaling without recognizing these and other contingencies. In birds and mammals, photoperiod and time of day of metabolic measurements may also affect thermal conductance and its scaling with body mass [6,82], and by association, metabolic rate and its scaling with body mass in the cold [56]. These contingent effects operate within the boundaries of geometric and physical constraints, including surface-area-to-volume relationships, and physical laws of heat flow.
However, one might argue that fish and crustaceans show different effects of Ta on metabolic scaling than do birds and mammals, because of differences in ecological habitat (aquatic vs. terrestrial), rather than thermoregulatory strategy (ectothermy vs. endothermy). Increasing temperature not only increases metabolic demand for oxygen both in water and on land, but in water, it also decreases oxygen concentration and thus its availability. The double jeopardy of higher oxygen demand and lower oxygen supply at high Ta in water, especially as body size increases, may therefore cause inverse relationships between Ta and the metabolic scaling exponent to be more prevalent in aquatic vs. terrestrial animals. However, as pointed out in Section 3.1, negative associations between Ta and the metabolic scaling exponent also occur in many terrestrial ectotherms. In addition, the habitat hypothesis cannot explain why associations between Ta and the metabolic scaling exponent are positive in endotherms. Therefore, I believe that it is more likely that the taxonomic differences in how Ta affects metabolic scaling observed in this study relate more to differences in thermoregulatory strategy than to habitat.
The MLBH succeeds in explaining temperature effects on metabolic scaling because it recognizes both contingent effects and physical constraints. Other prominent theoretical models based strictly on internal physical constraints cannot explain these patterns (see Section 3.1). The MLBH is part of a more general theory, the contextual multimodal theory, which includes multiple mechanisms that act in contingent ways ([10]; also see Section 5).
3.2.2. Suggestions for Future Research
I recommend future research that attempts to explain the diversity of metabolic scaling and its sensitivity to a variety of intrinsic and extrinsic factors, rather than a non-existent universal metabolic scaling law (e.g., 3/4-power law) (see [10,16,23,32,105]). Potentially productive approaches include comparative and experimental studies exploring how various intrinsic and extrinsic factors interact to produce a variety of metabolic scaling relationships, and how biological regulation mediates these interactions.
Further studies of how Ta affects metabolic scaling are required for comparisons both within and among species. As predicted by the MLBH (as amended in Section 3.1), in the laboratory mouse (Mus musculus domesticus), metabolic level is higher at 21 versus 29 °C, but the metabolic scaling slope is lower [106]. Varied patterns occur for other rodents, but these analyses suffer from very small body-mass ranges [107]. In addition, as predicted by the MLBH, the metabolic rate of marine zooplankton (including diverse taxa) increases in the order of boreal, temperate and tropical species, whereas the interspecific scaling exponent (based on dry masses) decreases (0.830, 0.691, and 0.538, respectively) [108], though this may not be true for planktonic crustaceans [109]. Although inverse relationships between Ta and the exponent (slope) for ontogenetic metabolic scaling are common in ectotherms, other patterns are also possible, including non-significant, positive and nonlinear relationships (reviewed in [16,26]). In this study, I suggest that thermoregulatory lifestyle (ectothermy versus endothermy) can affect the temperature-sensitivity of interspecific metabolic scaling. Other differences in lifestyle may also help explain the varied responses of intraspecific metabolic scaling to Ta observed in ectotherms. One key lifestyle feature may be activity level. According to the MLBH, for resting (inactive) metabolism, increasing metabolic level should be associated with decreasing scaling exponents, whereas for active metabolism, increasing metabolic level should be associated with increasing scaling exponents (because of an increase in the relative influence of volume-related locomotor power production) ([16,23,26]; cf. [92]). Therefore, in sedentery (immobile) organisms, a thermally increased metabolic level should result in a lowered metabolic scaling exponent, whereas, in actively mobile animals, a thermally increased metabolic level may result in a variety of effects on the metabolic scaling exponent, depending on the relative size-specific effects of Ta on activity level (also see [26]). Consistent with this hypothesis, sedentery or mostly stationary organisms (including plants, oysters, mussels, chitons, and ascideans) usually show strong negative associations between Ta and the resting metabolic scaling exponent (e.g., [110,111,112,113,114]), whereas actively mobile animals show a variety of responses (as reviewed in [16,26]; and as shown in an unpublished data set). As further evidence, when the effects of activity are removed in an actively mobile species, such as the fish Coregonus albula, Ta and the resting metabolic scaling exponent are strongly negatively correlated [115], as predicted by the MLBH [16,26]. Further studies of the effects of various abiotic and biotic ecological factors on metabolic scaling would also be worthwhile (see also [10,16,22,26,32,41,44,45,46,48]).
My study suggests that not only Ta, but also the mode of regulation of Tb, affect metabolic scaling. My interpretation of these contingent thermal effects contributes to a growing revival of the old, controversial view that thermoregulation is importantly involved in the metabolic scaling of endotherms ([10,11,12,16,23,24,25,26,27,28,32,56,58,87,89,93], but see [9,21,22,33,92,116,117]). This thermoregulatory view is testable. For example, since thermal insulation scales allometrically (slope ~ 0.17) in mammals with a body mass <10 kg, but remains constant in larger mammals [6], the thermoregulatory view predicts that at low Ta below the thermoneutral zone, the scaling exponent for metabolic rate should approach 0.50 in small mammals, but be near 0.67 in large mammals (cf. Section 3.1). The data provided in Figure 4 are only for mammals ≤14 kg, which show metabolic scaling exponents of 0.39–0.56 at Ta below the thermoneutral zone, which encompass the predicted slope of 0.50. As a further test, we now need data for larger mammals exposed to the cold. A thermoregulatory view may also help explain the curvilinearity of the scaling of mammalian basal metabolic rate in the thermoneutral zone, showing a slope near 2/3 in small mammals, and a higher slope (≥3/4) in larger mammals (e.g., [93,118,119]). Is it a coincidence that the breakpoint in the scaling of mammalian basal metabolic rate occurs near a body mass of 10 kg (4.25 or 20 kg, according to the two-segmented linear models of Kozłowski and Konarzewski [120] and Makarieva and colleagues [121], respectively)?
Furthermore, I suggest that contingent effects on metabolic scaling are often mediated by various biological regulatory systems. Although the importance of regulation in metabolic scaling was discussed over 50 years ago (e.g., [20,106,122]), it has been a neglected focus of research in recent years (but see [10,27,96,123,124]). The effects of regulation can be dramatic, as seen when mammals engage in regulated depression of their Tb and metabolic rate during torpor and hibernation [125]: as a result, their metabolic scaling slope shifts markedly from ~0.67–0.75 to ~1, as predicted by the MLBH [10,23,26,93]. This observation also reinforces the view that thermoregulation plays a major role in the metabolic scaling of endotherms. Therefore, I recommend that increased attention should be given to how various regulatory systems at various levels of biological organization (from biochemical signaling pathways to neuroendocrine systems) affect metabolic scaling. Metabolic scaling is not merely the result of physical constraints, but is also mediated by various regulated processes involving resource supply and demand, metabolic waste removal, and heat dissipation [10,27,96].
I also encourage investigators to employ well-controlled experiments and incisive comparative analyses to test further the relative merits of multiple theoretical models. All too often investigators have tested single models without considering the relative merits of alternative models. Testing “multiple working hypotheses” [126] simultaneously in a variety of taxa under diverse physiological and ecological conditions is a promising approach for resolving heated debates about the relative importance and range of applicability of various proposed mechanisms determining metabolic scaling (see e.g., [10,39,40,44,94,95,127,128]). In addition, attempts should be made to synthesize multiple models and mechanisms [10,32]. For example, synthesizing the MLBH with cell-size theory may expand the predictive power of both [26,129]. Although cell-size theory by itself cannot explain effects of Ta on the metabolic scaling exponent (see Section 3.1), it can, if combined with the MLBH. Thermally increased metabolic level may cause decreases in the scaling exponent for resting metabolic rate, not only because of the increased influence of surface-area related metabolic processes at the whole body level, but also at the cellular level, a hypothesis requiring testing [10,26,129]. Other examples of multi-mechanistic models of metabolic scaling are reviewed in [10,32].
4. Materials and Methods
The data sets used in this study were the best that I could find that showed the effects of ambient temperature on the scaling of metabolic rate, based on large samples of species with broad body-mass ranges. The ectotherm data sets [48,53] were the only ones available that used typical, native habitat temperatures (thus minimizing acute thermal stress responses). The endotherm data sets [56,58] were the only ones available that examined effects of ambient temperature on interspecific metabolic scaling in birds and mammals under normal atmospheric conditions. Resting/routine metabolic rate was estimated as the rate of oxygen consumption during fasting for crustaceans [53] fish [48] and mammals [58], and as rate of metabolized food energy (ingested–excreted energy) over several days for caged birds with stable body masses and essentially no extra energy demand beyond mere existence, i.e., “existence metabolism” [56]. Existence metabolism included energy used for maintenance, temperature regulation, food-processing and minimal locomotor activity, but excluded energy required for growth, reproduction, molting, migratory activity and fat deposition. Estimates of metabolized energy intake required determining the caloric content of ingested food and excreta. This method is comparable to that based on oxygen consumption during fasting because Ta affects the scaling slopes of existence metabolism and oxygen consumption rates in similar ways. As shown for existence metabolism (Figure 3), the interspecific scaling slope for oxygen consumption rate of birds decreases at lower Ta (30 °C: summer and winter scaling slopes = 0.658, 0.688 for passerines, and 0.701, 0.728 for non-passerines; 0 °C: summer and winter scaling slopes = 0.528, 0.531 for passerines, and 0.571, 0.594 for non-passerines [56]). Figure 3 depicts scaling relationships for existence metabolism, because they had larger sample sizes (and thus were more reliable) than those for oxygen consumption.
5. Conclusions
In my opinion, it is no longer a question of whether contingency or constraints affect metabolic scaling, but what are the relative roles of each. Continuing efforts to develop metabolic scaling theory based only on a single, universally applicable, physical mechanism are misguided. A comprehensive theory of metabolic scaling should be “meta-mechanistic”, i.e., it should recognize multiple, interactive mechanisms whose relative effects depend on various contingent intrinsic (biological) and extrinsic (ecological) conditions, within specified constraints [10]. The recently proposed contextual multimodal theory [10] offers one possible conceptual framework requiring further development and testing [32]. The challenge is to explain the actually observed diversity of metabolic scaling, not an idealized, non-existent 3/4-power law.
Acknowledgments
I thank the anonymous reviewers for their useful comments.
Conflicts of Interest
The author declares no conflict of interest.
References
- Huxley, J.S. Problems of Relative Growth; Dover Publications: New York, NY, USA, 1932. [Google Scholar]
- Gould, S.J. Allometry and size in ontogeny and phylogeny. Biol. Rev. 1966, 41, 587–638. [Google Scholar] [CrossRef] [PubMed]
- McMahon, T.A.; Bonner, J.T. On Size and Life; Scientific American Library: New York, NY, USA, 1983. [Google Scholar]
- Peters, R.H. The Ecological Implications of Body Size; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar]
- Calder, W.A. Size, Function and Life History; Harvard University Press: Cambridge, MA, USA, 1984. [Google Scholar]
- Schmidt-Nielsen, K. Scaling: Why Is Animal Size So Important? Cambridge University Press: New York, NY, USA, 1984. [Google Scholar]
- Brown, J.H.; West, G.B. (Eds.) Scaling in Biology; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Sibly, R.M.; Brown, J.H.; Kodric-Brown, A. (Eds.) Metabolic Ecology: A Scaling Approach; Wiley-Blackwell: Oxford, UK, 2012. [Google Scholar]
- West, G.B.; Brown, J.H. The origin of allometric scaling laws in biology from genomes to ecosystems: Towards a quantitative unifying theory of biological structure and organization. J. Exp. Biol. 2005, 208, 1575–1592. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. Metabolic scaling in complex systems. Systems 2014, 2, 451–540. [Google Scholar] [CrossRef]
- Sarrus, F.; Rameaux, J.F. Application des sciences accessoires et principalement des mathématiques à la physiologie générale (Rapport sur une mémoire adressé à l’Académic royale de Médecine, séance du 23 juillet 1839). Bull. Acad. R. Méd. 1839, 3, 1094–1100. [Google Scholar]
- Rubner, M. Über den Einfluss der Körpergrösse auf Stoff- und Kraftwechsel. Z. Biol. 1883, 19, 535–562. [Google Scholar]
- Richet, C.R. La Chaleur Animale; Bibliothèque Scientifique Internationale, Felix Alcan: Paris, France, 1889. [Google Scholar]
- Voit, E. Über die Grösse des Energiebedarfs der Tiere in Hungerzustande. Z. Biol. 1901, 41, 113–154. [Google Scholar]
- Heusner, A.A. Energy metabolism and body size I. Is the 0.75 mass exponent of Kleiber’s equation a statistical artifact? Respir. Physiol. 1982, 48, 1–12. [Google Scholar] [CrossRef]
- Glazier, D.S. Beyond the “3/4-power law”: Variation in the intra- and interspecific scaling of metabolic rate in animals. Biol. Rev. 2005, 80, 611–662. [Google Scholar] [CrossRef] [PubMed]
- Kleiber, M. Body size and metabolism. Hilgardia 1932, 6, 315–353. [Google Scholar] [CrossRef]
- Brody, S.; Procter, R.C. Relation between basal metabolism and mature body weight in different species of mammals and birds. Relat. Basal Metab. Bodyweight Man 1932, 116, 89–101. [Google Scholar]
- Hemmingsen, A.M. Energy metabolism as related to body size and respiratory surfaces, and its evolution. Rep. Steno Mem. Hosp. Nord. Insulin Lab. 1960, 9, 1–110. [Google Scholar]
- Kleiber, M. The Fire of Life; Wiley: New York, NY, USA, 1961. [Google Scholar]
- Savage, V.M.; Gillooly, J.F.; Woodruff, W.H.; West, G.B.; Allen, A.P.; Enquist, B.J.; Brown, J.H. The predominance of quarter-power scaling in biology. Funct. Ecol. 2004, 18, 257–282. [Google Scholar] [CrossRef]
- White, C.R.; Kearney, M.R. Metabolic scaling in animals: Methods, empirical results, and theoretical explanations. Compr. Physiol. 2014, 4, 231–256. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. A unifying explanation for diverse metabolic scaling in animals and plants. Biol. Rev. 2010, 85, 111–138. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.F.; Lightfoot, E.N.; Porter, W.P. A new model for the body size–metabolism relationship. Physiol. Biochem. Zool. 2010, 83, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Speakman, J.R.; Król, E. Maximal heat dissipation capacity and hyperthermia risk: Neglected key factors in the ecology of endotherms. J. Anim. Ecol. 2010, 79, 726–746. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. The scaling of metabolic scaling within physical limits. Systems 2014, 2, 425–450. [Google Scholar] [CrossRef]
- Glazier, D.S. Body-mass scaling of metabolic rate: What are the relative roles of cellular versus systemic effects? Biology 2015, 4, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Kwak, H.S.; Im, H.G.; Shim, E.B. A model for allometric scaling of mammalian metabolism with ambient heat loss. Integr. Med. Res. 2016, 5, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Norris, S. Of mice and mammoths: New approaches to understanding the biological implications of body size. BioScience 1998, 48, 887–892. [Google Scholar] [CrossRef]
- Agutter, P.S.; Wheatley, D.N. Metabolic scaling: Consensus or controversy? Theor. Biol. Med. Model. 2004, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agutter, P.S.; Tuszynski, J.A. Analytic theories of metabolic scaling. J. Exp. Biol. 2011, 214, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. Rediscovering and reviving old observations and explanations of metabolic scaling in living systems. Systems 2018, in press. [Google Scholar]
- West, G.B.; Brown, J.H.; Enquist, B.J. A general model for the origin of allometric scaling laws in biology. Science 1997, 276, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Banavar, J.R.; Maritan, A.; Rinaldo, A. Size and form in efficient transportation networks. Nature 1999, 399, 130–132. [Google Scholar] [CrossRef] [PubMed]
- Banavar, J.R.; Moses, M.E.; Brown, J.H.; Damuth, J.; Rinaldo, A.; Sibly, R.M.; Maritan, A. A general basis for quarter-power scaling in animals. Proc. Natl. Acad. Sci. USA 2010, 107, 15816–15820. [Google Scholar] [CrossRef] [PubMed]
- Dodds, P.S. On the optimal form of branching supply and collection networks. Phys. Rev. Lett. 2010, 104. [Google Scholar] [CrossRef] [PubMed]
- Banavar, J.R.; Cooke, T.J.; Rinaldo, A.; Maritan, A. Form, function, and evolution of living organisms. Proc. Natl. Acad. Sci. USA 2014, 111, 3332–3337. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.F. Do performance–safety tradeoffs cause hypometric metabolic scaling in animals? Trends Ecol. Evol. 2017, 32, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Hirst, A.G.; Glazier, D.S.; Atkinson, D. Body shape-shifting during growth permits tests that distinguish between competing geometric theories of metabolic scaling. Ecol. Lett. 2014, 17, 1274–1281. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S.; Hirst, A.G.; Atkinson, D. Shape shifting predicts ontogenetic changes in metabolic scaling in diverse aquatic invertebrates. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142302. [Google Scholar] [CrossRef] [PubMed]
- Vaca, H.F.; White, C.R. Environmental modulation of metabolic allometry in ornate rainbowfish Rhadinocentrus ornatus. Biol. Lett. 2010, 6, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Teissier, G. Recherches Morphologiques et Physiologiques sur la Croissance des Insectes: Thèses Presentées a la Faculté des Sciences de L’Université de Paris Pour Obtenir le Grade de Docteur ès Sciences Naturelles; Les Presses Universitaires de France: Paris, France, 1931. [Google Scholar]
- Duerr, F.G. Changes in the size-metabolic rate relationship of Lymnaea stagnalis appressa Say produced by digenetic trematode parasitism. Comp. Biochem. Physiol. 1967, 20, 391–398. [Google Scholar] [CrossRef]
- Glazier, D.S.; Butler, E.M.; Lombardi, S.A.; Deptola, T.J.; Reese, A.J.; Satterthwaite, E.V. Ecological effects on metabolic scaling: Amphipod responses to fish predators in freshwater springs. Ecol. Monogr. 2011, 81, 599–618. [Google Scholar] [CrossRef]
- McFeeters, B.J.; Xenopoulos, M.A.; Spooner, D.E.; Wagner, N.D.; Frost, P.C. Intraspecific mass-scaling of field metabolic rates of a freshwater crayfish varies with stream land cover. Ecosphere 2011, 2, 1–10. [Google Scholar] [CrossRef]
- Glazier, D.S. The 3/4-power law is not universal: Evolution of isometric, ontogenetic metabolic scaling in pelagic animals. BioScience 2006, 56, 325–332. [Google Scholar] [CrossRef]
- Killen, S.S.; Costa, I.; Brown, J.A.; Gamperl, A.K. Little left in the tank: Metabolic scaling in marine teleosts and its implications for aerobic scope. Proc. R. Soc. Lond. B 2007, 274, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Killen, S.S.; Atkinson, D.; Glazier, D.S. The intraspecific scaling of metabolic rate with body mass in fishes depends on lifestyle and temperature. Ecol. Lett. 2010, 13, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- McNab, B.K. An analysis of the factors that influence the level and scaling of mammalian BMR. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 151, 5–28. [Google Scholar] [CrossRef] [PubMed]
- McNab, B.K. Ecological factors affect the level and scaling of avian BMR. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2009, 152, 22–45. [Google Scholar] [CrossRef] [PubMed]
- Hulbert, A.J. A sceptics view: “Kleiber’s Law” or the “3/4 Rule” is neither a law nor a rule but rather an empirical approximation. Systems 2014, 2, 186–202. [Google Scholar] [CrossRef]
- Ivleva, I.V. The dependence of crustacean respiration rate on body mass and habitat temperature. Int. Rev. Gesamten Hydrobiol. Hydrogr. 1980, 65, 1–47. [Google Scholar] [CrossRef]
- Crab Cliparts Black #2812105. Available online: http://clipart-library.com/clipart/947704.htm (accessed on 24 January 2018).
- Drawing Fish #1416367. Available online: http://clipart-library.com/clipart/piode7j6T.htm (accessed on 24 January 2018).
- Kendeigh, S.C.; Dol’nik, V.R.; Gavrilov, V.M. Avian energetics. In Granivorous Birds in Ecosystems; Pinowski, J., Kendeigh, S.C., Eds.; Cambridge University Press: Cambridge, UK, 1977; pp. 127–204. [Google Scholar]
- Chickadee. Available online: https://pixabay.com/en/chickadee-bird-animal-wildlife-48019/ (accessed on 24 January 2018).
- Withers, P.C.; Casey, T.M.; Casey, K.K. Allometry of respiratory and haematological parameters of arctic mammals. Comp. Biochem. Physiol. A Physiol. 1979, 64, 343–350. [Google Scholar] [CrossRef]
- Fox Clipart Arctic Fox #1906. Available online: http://moziru.com/explore/Fox%20clipart%20arctic%20fox/#go_post_2942_polar-fox-clipart-19.jpg (accessed on 24 January 2018).
- Gillooly, J.F.; Brown, J.H.; West, G.B.; Savage, V.M.; Charnov, E.L. Effects of size and temperature on metabolic rate. Science 2001, 293, 2248–2251. [Google Scholar] [CrossRef] [PubMed]
- Arendt, J. Ecological correlates of body size in relation to cell size and cell number: Patterns in flies, fish, fruits and foliage. Biol. Rev. 2007, 82, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Hessen, D.O.; Daufresne, M.; Leinaas, H.P. Temperature-size relations from the cellular-genomic perspective. Biol. Rev. 2013, 88, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Czarnoleski, M.; Cooper, B.S.; Kierat, J.; Angilletta, M.J. Flies developed small bodies and small cells in warm and in thermally fluctuating environments. J. Exp. Biol. 2013, 216, 2896–2901. [Google Scholar] [CrossRef] [PubMed]
- Jalal, M.; Andersen, T.; Hessen, D.O. Temperature and developmental responses of body and cell size in Drosophila; effects of polyploidy and genome configuration. J. Therm. Biol. 2015, 51, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Adrian, G.J.; Czarnoleski, M.; Angilletta, M.J. Flies evolved small bodies and cells at high or fluctuating temperatures. Ecol. Evol. 2016, 6, 7991–7996. [Google Scholar] [CrossRef] [PubMed]
- Hermaniuk, A.; Rybacki, M.; Taylor, J.R. Low temperature and polyploidy result in larger cell and body size in an ectothermic vertebrate. Physiol. Biochem. Zool. 2016, 89, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Czarnoleski, M.; Labecka, A.M.; Starostová, Z.; Sikorska, A.; Bonda-Ostaszewska, E.; Woch, K.; Kubička, L.; Kratochvíl, L.; Kozlowski, J. Not all cells are equal: Effects of temperature and sex on the size of different cell types in the Madagascar ground gecko Paroedura picta. Biol. Open 2017, 6, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Kierat, J.; Szentgyörgyi, H.; Czarnoleski, M.; Woyciechowski, M. The thermal environment of the nest affects body and cell size in the solitary red mason bee (Osmia bicornis L.). J. Therm. Biol. 2017, 68, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Davison, J. An analysis of cell growth and metabolism in the crayfish (Procambarus alleni). Biol. Bull. 1956, 110, 264–273. [Google Scholar] [CrossRef]
- Szarski, H. Cell size and the concept of wasteful and frugal evolutionary strategies. J. Theor. Biol. 1983, 105, 201–209. [Google Scholar] [CrossRef]
- Gregory, T.R. A bird’s-eye view of the C-value enigma: Genome size, cell size, and metabolic rate in the class Aves. Evolution 2002, 56, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Starostová, Z.; Kubička, L.; Kozłowski, J.; Konarzewski, M.; Kratochvíl, L. Cell size but not genome size affects scaling of metabolic rate in eyelid geckos. Am. Nat. 2009, 174, E100–E105. [Google Scholar] [CrossRef] [PubMed]
- Maciak, S.; Janko, K.; Kotusz, J.; Choleva, L.; Boroń, A.; Juchno, D.; Kujawa, R.; Kozlowski, J.; Konarzewski, M. Standard metabolic rate (SMR) is inversely related to erythrocyte and genome size in allopolyploid fish of the Cobitis taenia hybrid complex. Funct. Ecol. 2011, 25, 1072–1078. [Google Scholar] [CrossRef]
- Huang, Q.; Zhang, Y.; Liu, S.; Wang, W.; Luo, Y. Intraspecific scaling of the resting and maximum metabolic rates of the crucian carp (Carassius auratus). PLoS ONE 2013, 8, e82837. [Google Scholar] [CrossRef] [PubMed]
- Maciak, S.; Bonda-Ostaszewska, E.; Czarnoleski, M.; Konarzewski, M.; Kozlowski, J. Mice divergently selected for high and low basal metabolic rates evolved different cell size and organ mass. J. Evol. Biol. 2014, 27, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, Q.; Liu, S.; He, D.; Wei, G.; Luo, Y. Intraspecific mass scaling of metabolic rates in grass carp (Ctenopharyngodon idellus). J. Comp. Physiol. B 2014, 184, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Hermaniuk, A.; Rybacki, M.; Taylor, J.R. Metabolic rate of diploid and triploid Edible Frog Pelophylax esculentus correlates inversely with cell size in tadpoles but not in frogs. Physiol. Biochem. Zool. 2017, 90, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Crispin, T.S.; White, C.R. Effect of thermal acclimation on organ mass, tissue respiration, and allometry in Leichhardtian River prawns Macrobrachium tolmerum (Riek, 1951). Physiol. Biochem. Zool. 2013, 86, 470–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herreid, C.F.; Kessel, B. Thermal conductance in birds and mammals. Comp. Biochem. Physiol. 1967, 21, 405–414. [Google Scholar] [CrossRef]
- Lasiewski, R.C.; Weathers, W.W.; Bernstein, M.H. Physiological responses of the giant hummingbird, Patagona gigas. Comp. Biochem. Physiol. 1967, 23, 797–813. [Google Scholar] [CrossRef]
- Bradley, S.R.; Deavers, D.R. A re-examination of the relationship between thermal conductance and body weight in mammals. Comp. Biochem. Physiol. A Physiol. 1980, 65, 465–476. [Google Scholar] [CrossRef]
- Aschoff, J. Thermal conductance in mammals and birds: Its dependence on body size and circadian phase. Comp. Biochem. Physiol. A Physiol. 1981, 69, 611–619. [Google Scholar] [CrossRef]
- Schleucher, E.; Withers, P.C. Re-evaluation of the allometry of wet thermal conductance for birds. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 129, 821–827. [Google Scholar] [CrossRef]
- Withers, P.C.; Cooper, C.E.; Larcombe, A.N. Environmental correlates of physiological variables in marsupials. Physiol. Biochem. Zool. 2006, 79, 437–453. [Google Scholar] [CrossRef] [PubMed]
- Fristoe, T.S.; Burger, J.R.; Balk, M.A.; Khaliq, I.; Hof, C.; Brown, J.H. Metabolic heat production and thermal conductance are mass-independent adaptations to thermal environment in birds and mammals. Proc. Nat. Acad. Sci. USA 2015, 112, 15934–15939. [Google Scholar] [CrossRef] [PubMed]
- Rezende, E.L.; Bacigalupe, L.D. Thermoregulation in endotherms: Physiological principles and ecological consequences. J. Comp. Physiol. B 2015, 185, 709–727. [Google Scholar] [CrossRef] [PubMed]
- Kooijman, S.A.L.M. Dynamic Energy and Mass Budgets in Biological Systems; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Kleiber, M. Body size, conductance for animal heat flow and Newton’s law of cooling. J. Theor. Biol. 1972, 37, 139–150. [Google Scholar] [CrossRef]
- Porter, W.P.; Kearney, M. Size, shape, and the thermal niche of endotherms. Proc. Nat. Acad. Sci. USA 2009, 106 (Suppl. 2), 19666–19672. [Google Scholar] [CrossRef] [PubMed]
- Heldmaier, G. Cold-adaptive changes of heat production in mammals. In Proceedings of the International Symposium on Environmental Physiology: Bioenergetics; Smith, R.E., Hannon, J.P., Shields, J.L., Horwitz, B.A., Eds.; Federation of American Societies for Experimental Biology: Bathesda, MD, USA, 1972; pp. 79–82. [Google Scholar]
- Rezende, E.L.; Swanson, D.L.; Novoa, F.F.; Bozinovic, F. Passerines versus nonpasserines: So far, no statistical differences in the scaling of avian energetics. J. Exp. Biol. 2002, 205, 101–107. [Google Scholar] [PubMed]
- White, C.R.; Seymour, R.S. Allometric scaling of mammalian metabolism. J. Exp. Biol. 2005, 208, 1611–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazier, D.S. Effects of metabolic level on the body size scaling of metabolic rate in birds and mammals. Proc. R. Soc. B 2008, 275, 1405–1410. [Google Scholar] [CrossRef] [PubMed]
- White, C.R.; Kearney, M.R.; Matthews, P.G.; Kooijman, S.A.; Marshall, D.J. A manipulative test of competing theories for metabolic scaling. Am. Nat. 2011, 178, 746–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearney, M.R.; White, C.R. Testing metabolic theories. Am. Nat. 2012, 180, 546–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazier, D.S. Is metabolic rate a universal “pacemaker” for biological processes? Biol. Rev. 2015, 90, 377–407. [Google Scholar] [CrossRef] [PubMed]
- Verberk, W.C.; Atkinson, D. Why polar gigantism and Palaeozoic gigantism are not equivalent: Effects of oxygen and temperature on the body size of ectotherms. Funct. Ecol. 2013, 27, 1275–1285. [Google Scholar] [CrossRef] [Green Version]
- Hoque, A.T.M.R.; Sharma, S.; Suwa, R.; Mori, S.; Hagihara, A. Seasonal variation in the size-dependent respiration of mangroves Kandelia obovata. Mar. Ecol. Prog. Ser. 2010, 404, 31–37. [Google Scholar] [CrossRef]
- Roy, A. Analyse des facteurs du taux de metabolism chez la limace Arion circumscriptus. Rev. Can. Biol. 1969, 28, 33–43. [Google Scholar] [PubMed]
- Al-Sadoon, M.K.; Abdo, N.M. Temperature and body mass effects on the metabolic rate of Acanthodactylus schmidti Weigmann (Reptilia: Lacertidae). J. Arid Environ. 1991, 21, 351–361. [Google Scholar]
- Weldon, C.W.; Daniels, S.R.; Clusella-Trullas, S.; Chown, S.L. Metabolic and water loss rates of two cryptic species in the African velvet worm genus Opisthopatus (Onychophora). J. Comp. Physiol. B 2013, 183, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. Ontogenetic body-mass scaling of resting metabolic rate covaries with species-specific metabolic level and body size in spiders and snakes. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2009, 153, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. Activity affects intraspecific body-size scaling of metabolic rate in ectothermic animals. J. Comp. Physiol. B 2009, 179, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Carey, N.; Sigwart, J.D.; Richards, J.G. Economies of scaling: More evidence that allometry of metabolism is linked to activity, metabolic rate and habitat. J. Exp. Mar. Biol. Ecol. 2013, 439, 7–14. [Google Scholar] [CrossRef]
- Patterson, M.R. A mass transfer explanation of metabolic scaling relationships in some aquatic invertebrates and algae. Science 1992, 255, 1421–1423. [Google Scholar] [CrossRef] [PubMed]
- Von Bertalanffy, L. Basic concepts in quantitative biology of metabolism. Helgol. Wiss. Meeresunters. 1964, 9, 5–37. [Google Scholar] [CrossRef]
- Hart, J.S. Rodents. In Comparative Physiology of Thermoregulation: Volume II Mammals; Whittow, G.C., Ed.; Academic Press: New York, NY, USA, 1971; pp. 1–149. [Google Scholar]
- Ikeda, T. Relationship between respiration rate and body size in marine plankton animals as a function of the temperature of habitat. Bull. Fac. Fish. Hokkaido Univ. 1970, 21, 91–112. [Google Scholar]
- Uye, S.I.; Yashiro, M. Respiration rates of planktonic crustaceans from the Inland Sea of Japan with special reference to the effects of body weight and temperature. J. Oceanogr. 1988, 44, 47–51. [Google Scholar] [CrossRef]
- Carey, N.; Sigwart, J.D. Size matters: Plasticity in metabolic scaling shows body-size may modulate responses to climate change. Biol. Lett. 2014, 10, 20140408. [Google Scholar] [CrossRef] [PubMed]
- Dame, R.F. The ecological energies of growth, respiration and assimilation in the intertidal American oyster Crassostrea virginica. Mar. Biol. 1972, 17, 243–250. [Google Scholar] [CrossRef]
- Read, K.R.H. Respiration of the bivalved molluscs Mytilus edulis L. and Brachidontes demissus plicatulus Lamarck as a function of size and temperature. Comp. Biochem. Physiol. 1962, 7, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Haure, J.; Penisson, C.; Bougrier, S.; Baud, J.P. Influence of temperature on clearance and oxygen consumption rates of the flat oyster Ostrea edulis: Determination of allometric coefficients. Aquaculture 1998, 169, 211–224. [Google Scholar] [CrossRef]
- Jiang, A.; Guo, J.; Cai, W.; Wang, C. Oxygen consumption of the ascidian Styela clava in relation to body mass, temperature and salinity. Aquac. Res. 2008, 39, 1562–1568. [Google Scholar] [CrossRef]
- Ohlberger, J.; Staaks, G.; Hölker, F. Effects of temperature, swimming speed and body mass on standard and active metabolic rate in vendace (Coregonus albula). J. Comp. Physiol. B 2007, 177, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Heusner, A.A. Energy metabolism and body size II. Dimensional analysis and energetic non-similarity. Respir. Physiol. 1982, 48, 13–25. [Google Scholar] [CrossRef]
- Da Silva, J.K.L.; Garcia, G.J.; Barbosa, L.A. Allometric scaling laws of metabolism. Phys. Life Rev. 2006, 3, 229–261. [Google Scholar] [CrossRef]
- Lovegrove, B.G. The zoogeography of mammalian basal metabolic rate. Am. Nat. 2000, 156, 201–219. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.; Rothery, P.; Isaac, N.J.B. Scaling of basal metabolic rate with body mass and temperature in mammals. J. Anim. Ecol. 2010, 79, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Kozłowski, J.; Konarzewski, M. West, Brown and Enquist’s model of allometric scaling again: The same questions remain. Funct. Ecol. 2005, 19, 739–743. [Google Scholar] [CrossRef]
- Makarieva, A.M.; Gorshkov, V.G.; Li, B.-L. A note on metabolic rate dependence on body size in plants and animals. J. Theor. Biol. 2003, 221, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Von Bertalanffy, L.; Pirozynski, W.J. Tissue respiration, growth, and basal metabolism. Biol. Bull. 1953, 105, 240–256. [Google Scholar] [CrossRef]
- Chaui-Berlinck, J.G.; Navas, C.A.; Monteiro, L.H.A.; Bicudo, J.E.P.W. Control of metabolic rate is a hidden variable in the allometric scaling of homeotherms. J. Exp. Biol. 2005, 208, 1709–1716. [Google Scholar] [CrossRef] [PubMed]
- Suarez, R.K. Energy and metabolism. Compr. Physiol. 2012, 2, 2527–2539. [Google Scholar] [CrossRef] [PubMed]
- Jastroch, M.; Giroud, S.; Barrett, P.; Geiser, F.; Heldmaier, G.; Herwig, A. Seasonal control of mammalian energy balance: Recent advances in the understanding of daily torpor and hibernation. J. Neuroendocrinol. 2016, 28. [Google Scholar] [CrossRef] [PubMed]
- Chamberlin, T.C. The method of multiple working hypotheses. Science 1965, 148, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Van der Meer, J. Metabolic theories in ecology. Trends Ecol. Evol. 2006, 21, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Isaac, N.J.; Carbone, C. Why are metabolic scaling exponents so controversial? Quantifying variance and testing hypotheses. Ecol. Lett. 2010, 13, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; He, D.; Li, G.; Xie, H.; Zhang, Y.; Huang, Q. Intraspecific metabolic scaling exponent depends on red blood cell size in fishes. J. Exp. Biol. 2015, 218, 1496–1503. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
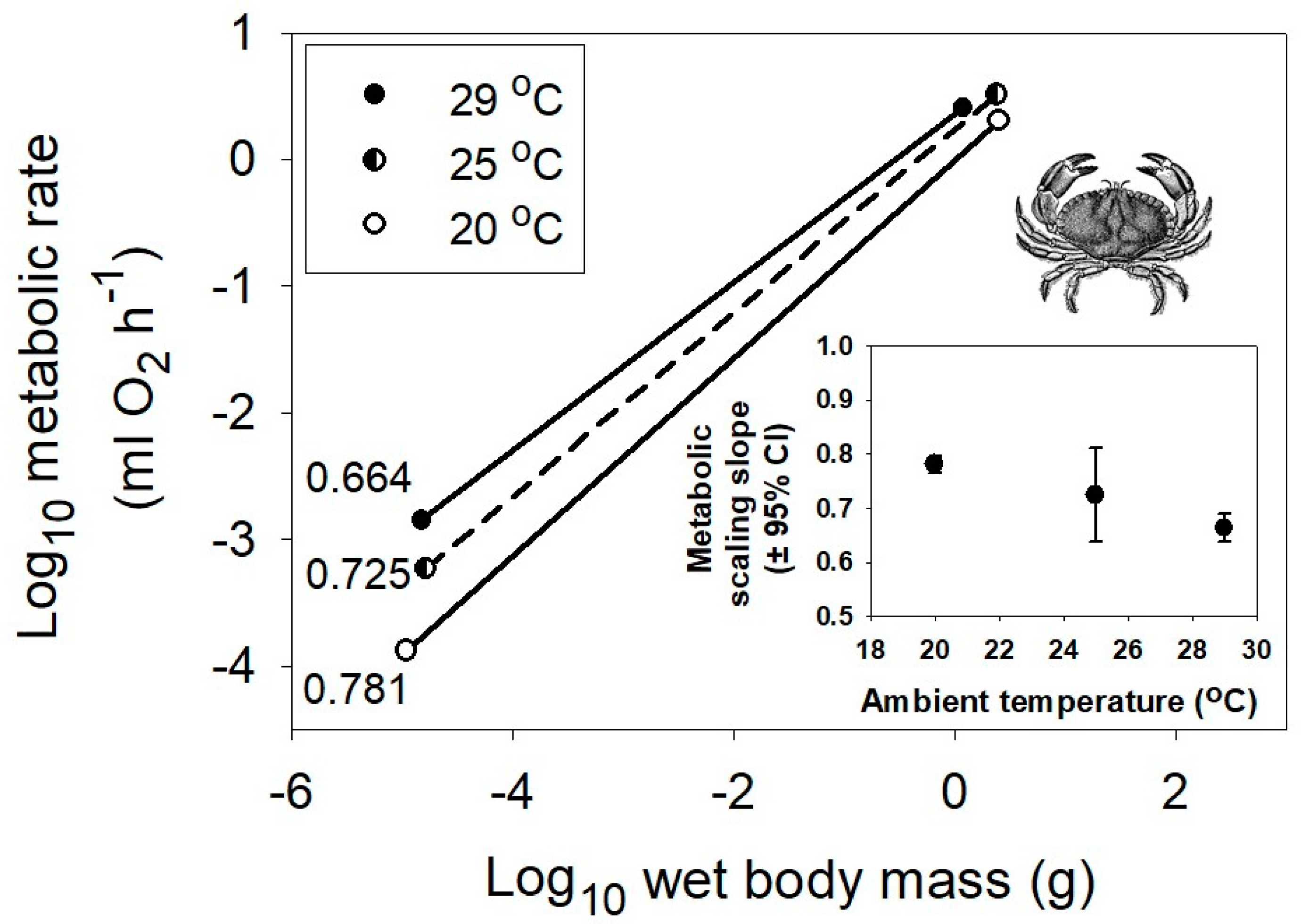
Interspecific scaling of metabolic rate in relation to body mass of marine crustaceans at three different habitat temperatures, 29, 25 and 20 °C (data from [53]). The correlation coefficients (r) and sample sizes (n) are 0.963 and 212, 0.727 and 249, and 0.977 and 247, respectively. All probability values are <0.001. The inset graph shows the negative relationship between the metabolic scaling slope (±95% confidence intervals) and habitat temperature. The crab picture is from [54].
Figure 1.
Interspecific scaling of metabolic rate in relation to body mass of marine crustaceans at three different habitat temperatures, 29, 25 and 20 °C (data from [53]). The correlation coefficients (r) and sample sizes (n) are 0.963 and 212, 0.727 and 249, and 0.977 and 247, respectively. All probability values are <0.001. The inset graph shows the negative relationship between the metabolic scaling slope (±95% confidence intervals) and habitat temperature. The crab picture is from [54].
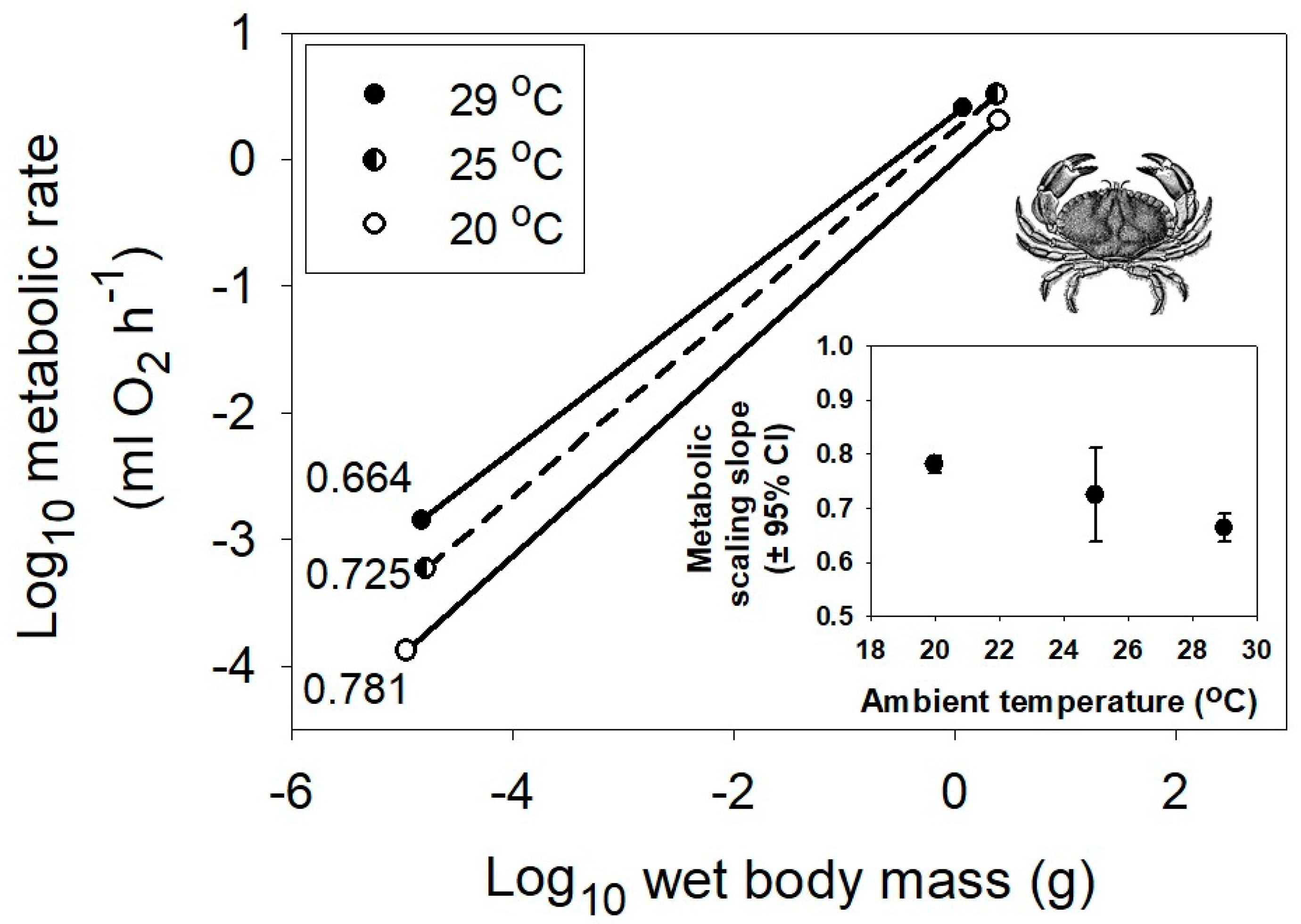
Figure 2.
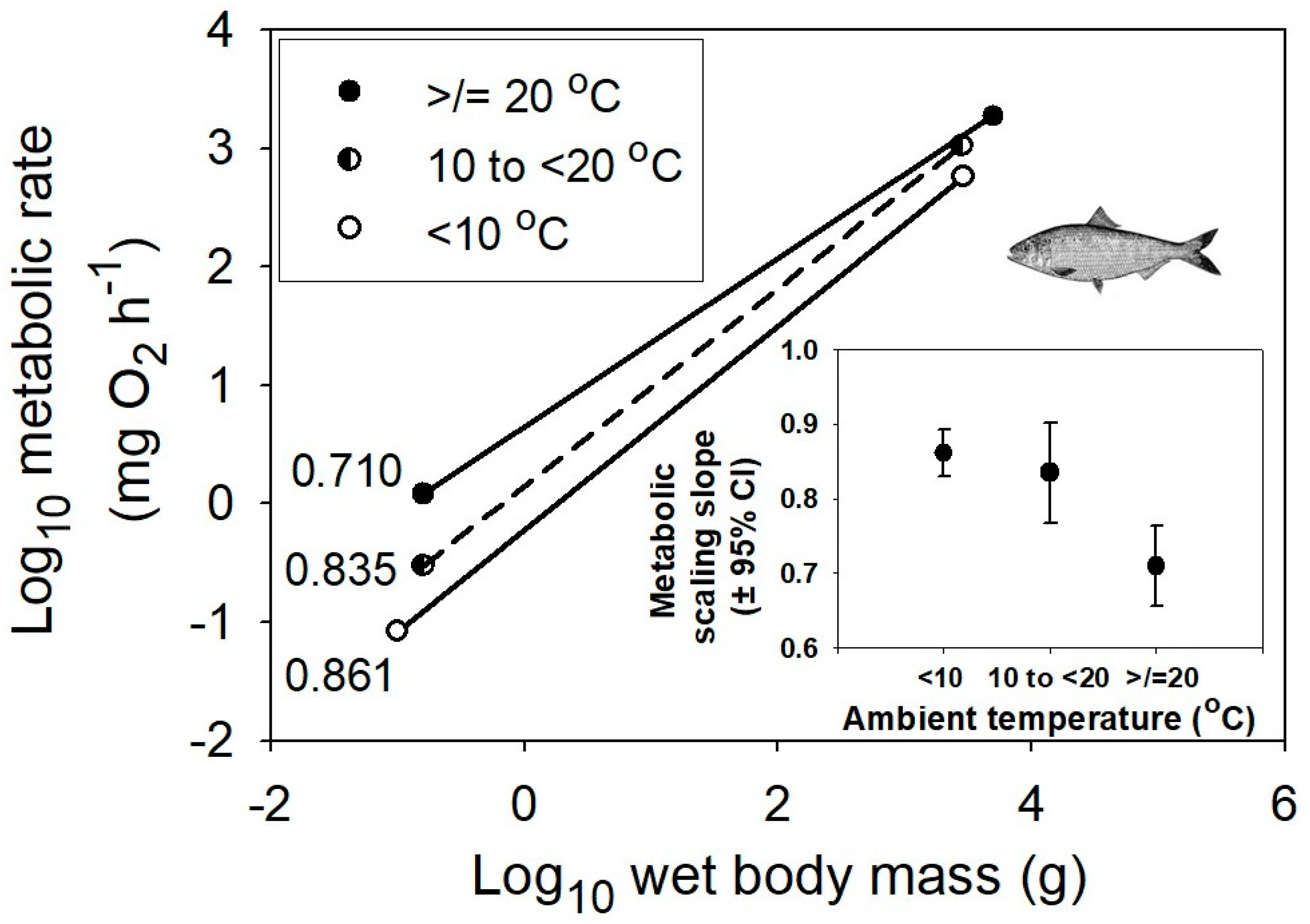
Ontogenetic scaling of metabolic rate in relation to body mass averaged for multiple species of marine teleost fishes at three different habitat temperature ranges, ≥20, 10 to <20, and <10 °C (data from [48]). The sample sizes (n) are 30, 19 and 40, respectively. The inset graph shows the negative relationship between the mean metabolic scaling slope (±95% confidence intervals) and habitat temperature. The fish picture is from [55].
Figure 2.
Ontogenetic scaling of metabolic rate in relation to body mass averaged for multiple species of marine teleost fishes at three different habitat temperature ranges, ≥20, 10 to <20, and <10 °C (data from [48]). The sample sizes (n) are 30, 19 and 40, respectively. The inset graph shows the negative relationship between the mean metabolic scaling slope (±95% confidence intervals) and habitat temperature. The fish picture is from [55].
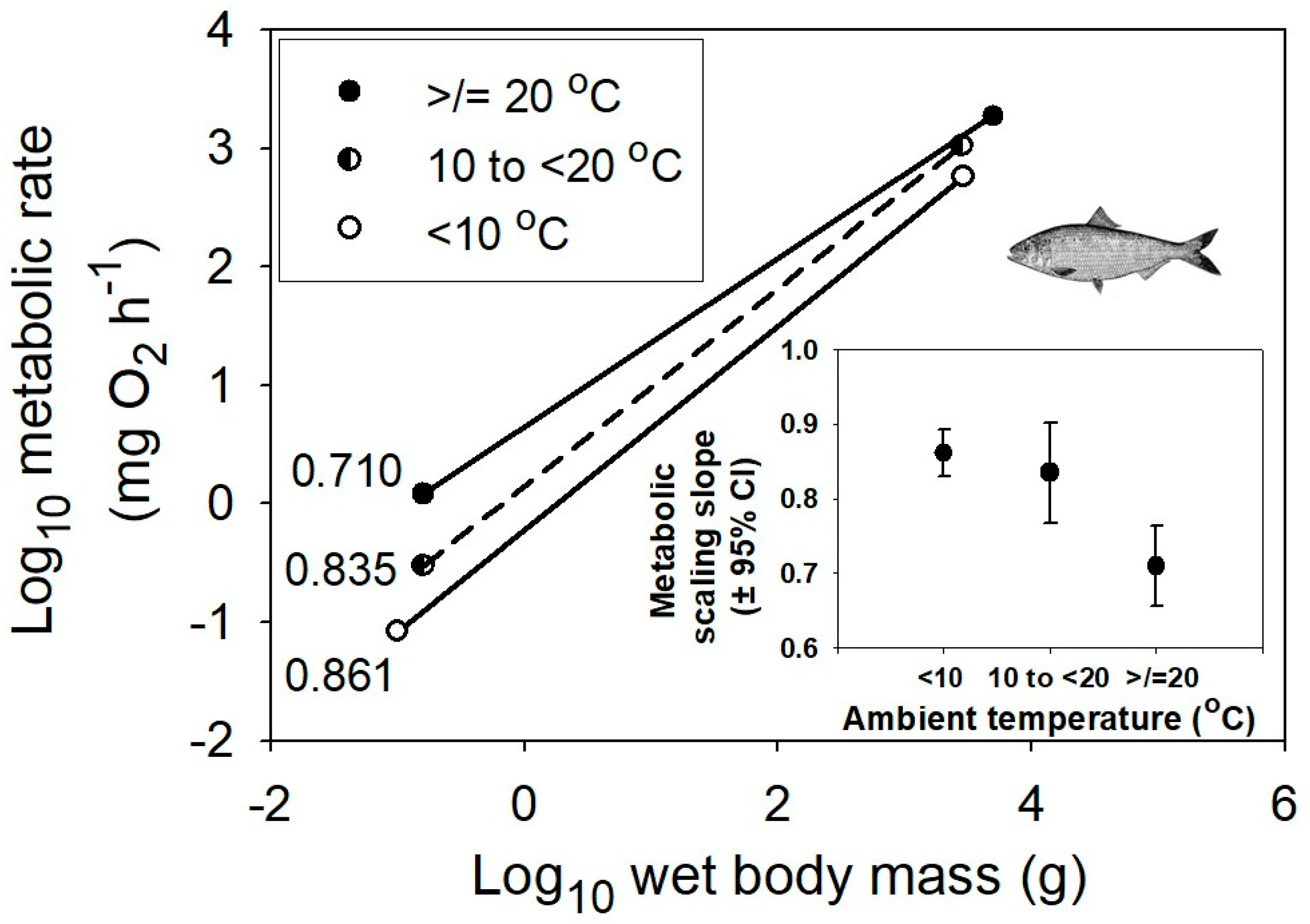
Figure 3.
Interspecific scaling of existence metabolism in relation to body mass for multiple species of passerine and non-passerine birds at two different air temperatures, 0 and 30 °C (data from [56]). The sample sizes (n) are 71 for passerine birds, and 40 for non-passerine birds. The inset graph shows the positive relationships between the metabolic scaling slopes and temperature for both passerine and non-passerine birds. The bird picture is from [57].
Figure 3.
Interspecific scaling of existence metabolism in relation to body mass for multiple species of passerine and non-passerine birds at two different air temperatures, 0 and 30 °C (data from [56]). The sample sizes (n) are 71 for passerine birds, and 40 for non-passerine birds. The inset graph shows the positive relationships between the metabolic scaling slopes and temperature for both passerine and non-passerine birds. The bird picture is from [57].
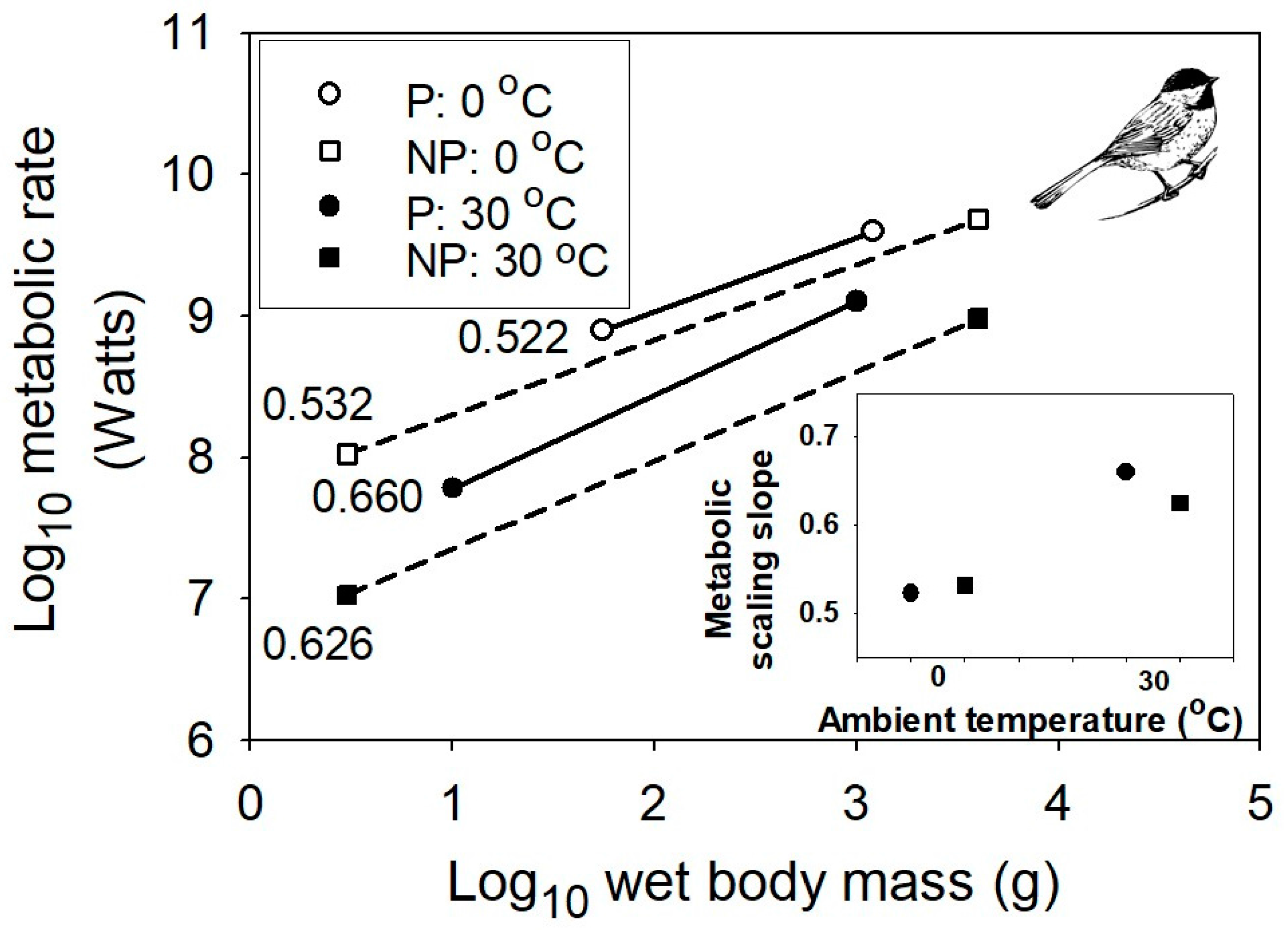
Figure 4.
Interspecific scaling of metabolic rate in relation to body mass for multiple species of eutherian mammals at four different air temperatures, 0, 10, 20 and 30 °C (data from [58]). The correlation coefficients (r) and sample sizes (n) are 0.91 and 61, 0.84 and 69, 0.88 and 73, and 0.89 and 81, respectively. All probability values are <0.001. The inset graph shows the positive relationship between the metabolic scaling slope (±95% confidence intervals) and temperature. The mammal picture is from [59].
Figure 4.
Interspecific scaling of metabolic rate in relation to body mass for multiple species of eutherian mammals at four different air temperatures, 0, 10, 20 and 30 °C (data from [58]). The correlation coefficients (r) and sample sizes (n) are 0.91 and 61, 0.84 and 69, 0.88 and 73, and 0.89 and 81, respectively. All probability values are <0.001. The inset graph shows the positive relationship between the metabolic scaling slope (±95% confidence intervals) and temperature. The mammal picture is from [59].
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Glazier, D.S. Effects of Contingency versus Constraints on the Body-Mass Scaling of Metabolic Rate. Challenges 2018, 9, 4. https://doi.org/10.3390/challe9010004
AMA Style
Glazier DS. Effects of Contingency versus Constraints on the Body-Mass Scaling of Metabolic Rate. Challenges. 2018; 9(1):4. https://doi.org/10.3390/challe9010004
Chicago/Turabian StyleGlazier, Douglas S. 2018. "Effects of Contingency versus Constraints on the Body-Mass Scaling of Metabolic Rate" Challenges 9, no. 1: 4. https://doi.org/10.3390/challe9010004
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.