Roles of Nicotinamide Adenine Dinucleotide (NAD+) in Biological Systems



Institute of Sciences of Food Productions, CNR-ISPA, Ecotekne, via prov.le Lecce-Monteroni km 7, 73100 Lecce, Italy
*
Author to whom correspondence should be addressed.
Challenges 2018, 9(1), 3; https://doi.org/10.3390/challe9010003
Submission received: 12 December 2017
/
Revised: 12 January 2018
/
Accepted: 16 January 2018
/
Published: 19 January 2018
(This article belongs to the Special Issue Poly ADP Ribose Polymerases (PARP) and Post-Translational Modifications)
Abstract
:NAD+ has emerged as a crucial element in both bioenergetic and signaling pathways since it acts as a key regulator of cellular and organism homeostasis. NAD+ is a coenzyme in redox reactions, a donor of adenosine diphosphate-ribose (ADPr) moieties in ADP-ribosylation reactions, a substrate for sirtuins, a group of histone deacetylase enzymes that use NAD+ to remove acetyl groups from proteins; NAD+ is also a precursor of cyclic ADP-ribose, a second messenger in Ca++ release and signaling, and of diadenosine tetraphosphate (Ap4A) and oligoadenylates (oligo2′-5′A), two immune response activating compounds. In the biological systems considered in this review, NAD+ is mostly consumed in ADP-ribose (ADPr) transfer reactions. In this review the roles of these chemical products are discussed in biological systems, such as in animals, plants, fungi and bacteria. In the review, two types of ADP-ribosylating enzymes are introduced as well as the pathways to restore the NAD+ pools in these systems.
Keywords:
Nicotinamide adenine dinucleotide (NAD+); ADP ribose (ADPr); ADP Ribosyl Transferase (ADPRT); nicotinamide (Nam); nicotinamide mononucleotide (NMN); nicotinamide riboside (NR); nicotinic acid (NA); nicotinic acid adenine dinucleotide phosphate (NAADP); cyclic ADP ribose (cADPr); post-translational modification (PTM); deacetylation; poly ADP ribose glycohydrolase (PARG); Nicotinamide mononucleotide adenylyl transferease (NMNAT-1); nicotinamide phosphoribosyl transferase (NAMPT)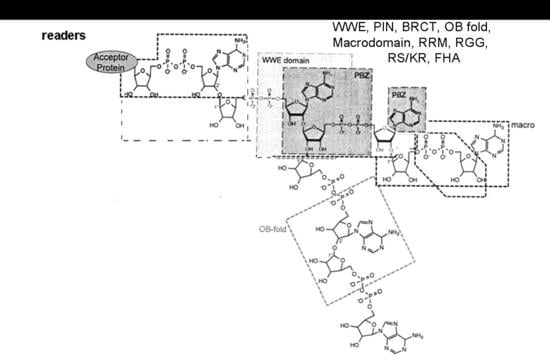
{kind=link}
{kind=link}
{kind=link}
1. Introduction
1.1. Nicotinamide Adenine Dinucleotide (NAD+)
NAD+ has emerged as a dual faced compound, present in all the kingdoms of life, being a cofactor in metabolic pathways and redox reactions, and also a substrate used by enzymes involved in post-translational modifications, in particular in deacetylation reactions, and in ADP ribosylation reactions, producing mono ADP ribose (MAR) and poly ADP ribose (PAR) moieties bound to proteins and nucleic acids [1,2,3,4,5,6]. In the majority of organisms, ADP ribosylation refers to NAD+-dependent, MAR/PAR modification of proteins, catalyzed by enzymes of the ADP-ribosyl transferase (ART diphtheria toxin like, ARTD) family, as well as some members of the Sirtuin family [7,8,9].
In deacetylation reactions, sirtuins, belonging to the histone deacetylase (HDAC) family, use NAD+ to remove acetyl groups from proteins: the deacetylation reaction couples lysine deacetylation to NAD hydrolysis, yielding O-acetyl-ADP-ribose, and nicotinamide. In specific compartments, a certain number of sirtuins behave as ADPr modifying enzymes.
Finally, NAD+ is a precursor of the second messengers cyclic ADP-ribose and nicotinic acid adenine dinucleotide phosphate (NAADP). NAD+ is mostly consumed in ADP-ribose (ADPr) transfer reactions, and NAD+ is also used to produce diadenosine tetraphosphate (Ap4A) and oligoadenylates (oligo2’-5’A). Finally, in plants, extracellular NAD+ is recognized by a membrane receptor and sensed as a damage activated molecular pattern, and the increased availability of NAD+ increases the immune response to plant pathogens.
1.2. NAD+ Dependent ADP Ribosylation and Deacetylation Reactions
Among the post-translational modifications of proteins, ADP ribosylation plays a large part. When the reaction involves a single monomer attached to proteins, it is named mono ADP ribosylation (MARylation), while the attachment of a complex, branched polymer to proteins is named poly ADP ribosylation (PARylation). ADP-ribosyl transferases (ARTs) are described as writers [5], either MAR- or PAR- writers. ADP-ribosylation exerts allosteric effects on enzymes, thereby controlling their catalytic activity, as well as taking together proteins that need to be in tight proximity to form protein complexes activating specific pathways, such as immune response, DNA damage response, and transcription activation. Post-translational modification of macromolecules requires that the reaction be reversible, that coupling enzymes catalyze the modification (ADPRTs, writers), and that enzymes reverse the modification (ADPR- glycohydrolases, erasers). Principal erasers are poly ADP ribose glycohydrolases (PARG), which depolymerize the PAR branches in a cycle of PAR synthesis by PARP1/2, and rapid degradation by PARG. In addition, there are two ADP ribosyl-acceptor hydrolases (ARH), ARH3 and ARH1, that are able to cut the bond between the proximal ADP-ribose and the amino acid [1].
The amino acids being modified during protein ADP-ribosylation are various, the attachment may occur on carboxylic groups, such as on Aspartate (D) and Glutamate (E), on amino or guanidino groups, such as in Lysine (K) and Arginine (R), thiol group in Cysteine (C) (non-enzymatic attachment), and on hydroxyl groups as in Serine (S). Therefore, various and different post-translational modifications may modify the same acceptor amino acid precluding its modification by another type of enzyme, or may impede the access to a neighbor site by hindrance, or by repulsing charges.
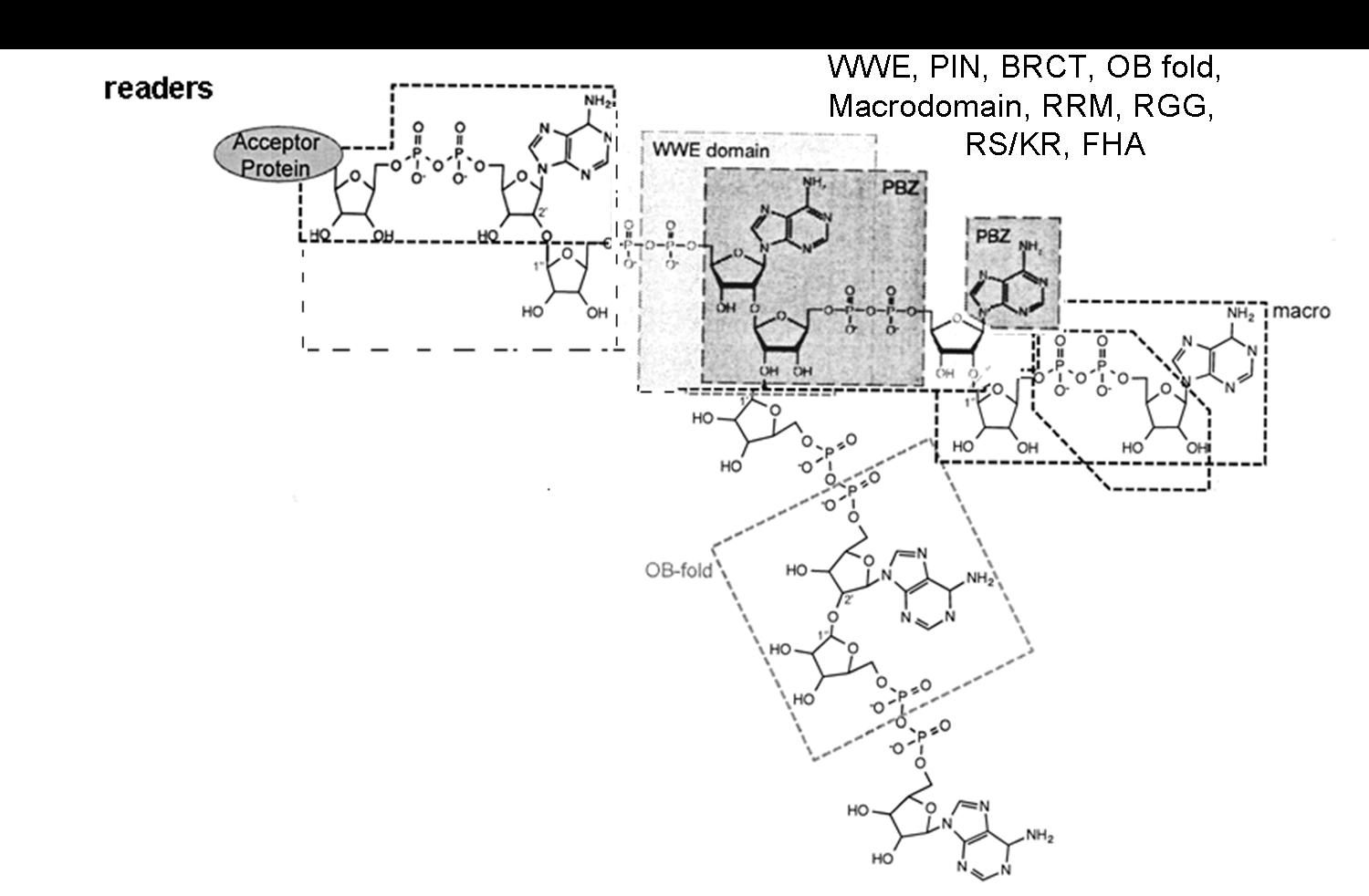
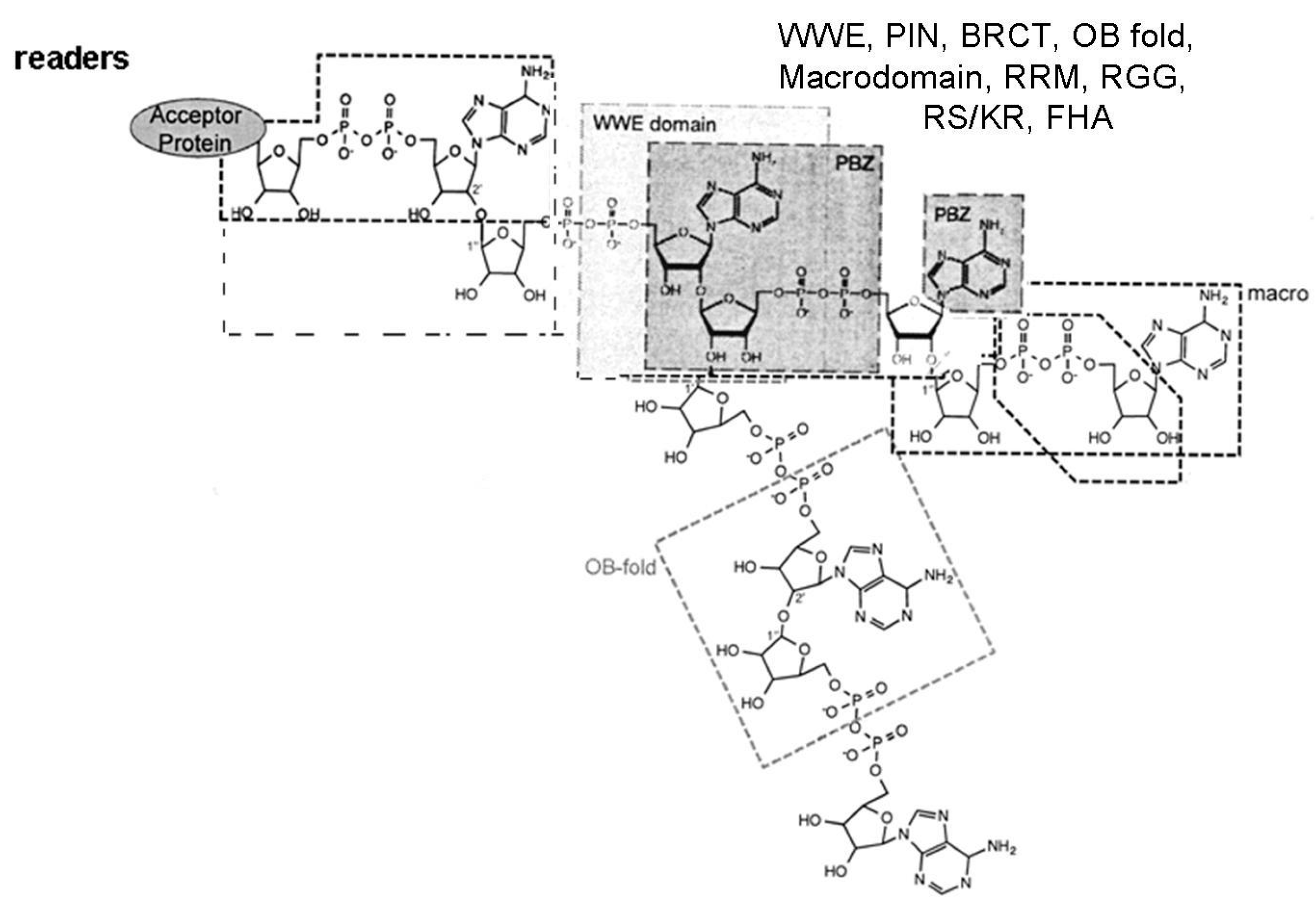
The complex of modification enzymes, in addition to writers and erasers, includes also the readers and proteins that bind to MAR or to PAR segments, interacting with specific domains [5] (Figure 1). The MAR and PAR regions are recognized (read) by protein motifs such as Macrodomains, PAR Binding Motif (PBM), PAR Zinc finger (PBZ) domains (binding to ADP ribose-ADP ribose junctions), WWE domains (tryptophan/glutamate) and Forkhead-associated (FHA) domain (binding to iso ADP ribose), oligonucleotide binding fold (OB fold), RNA recognition Motif (RRM), the serine arginine repeats and lysine arginine repeats (SR/KR) motives in RNA binding proteins, PIN (PilT N-terminal) domain in PIN and Exo1 nucleases, and BRCA1 C-Terminal region (BRCT) in the Breast Cancer 1 (BRCA1) antioncogene, binding to (ADP ribose)n units: these recognition events enable the proteins to aggregate other proteins and form protein-protein interaction complexes [5].
Several Macrodomain-containing proteins have been identified; some of them possess only PAR or MAR binding (readers), while other proteins have eraser activity: macrodomains carrying glycohydrolase activity can release the MAR/PAR from the modified amino acid of the protein target.
Macrodomain containing histones are activated by PAR formation [10]. Parp9, a mono-ADP-ribosyl transferase reported to be enzymatically inactive, forms heterodimers with Dtx3L, a histone ubiquitin (Ub) E3 ligase: Dtx3L/Parp9 ADP-ribosylates the carboxyl group of Ub Gly76. Parp9 ADP-ribosylation activity therefore restrains the E3 function of Dtx3L. Mutation of the NAD+ binding site in Parp9 increases the DNA repair activity of the heterodimer. Moreover, poly(ADP-ribose) binding to the Parp9 macrodomains increases E3 activity [11].
Very few organisms are devoid of the enzymes involved in PAR synthesis and degradation. Almost all fungi contain either PARP and/or PARG genes. In Trichoderma species, T. reesei has only a PARP protein (TR_22115). Similar to A. nidulans and N. crassa, T. atroviride and T. virens have two genes encoding PARP (TA_295780 and TV_89857) and PARG (TA_219648 and TV_4413) proteins [12]. PARG could be dispensable or a different poly(ADP-ribose) hydrolysis pathway operates in T. reesei.
Budding yeast does not possess proteins with significant sequence similarity to the human PARP family of proteins. It has been shown that the expression of human PARP in Saccharomyces cerevisiae yeasts led to cell growth arrest. Yeast growth was restored by coexpression of PARG ensuring cycling of PAR polymer. PARP-1 reduced UV-induced homologous recombination in S. cerevisiae [13,14,15].
1.3. Bacterial Toxins
Bacteria infecting eukaryotic cells, as well as plant pathogens, produce toxins that promote bacterial infections. In plants the toxins, or effectors, are injected into the apoplast by bacterial type III secretion (T3S) systems. Among the toxins known, that are ADPRT enzymes that modify cellular proteins in order to derail from their activity. The majority of ADP-ribosylating toxins belong to the poly ADP-ribose polymerase (PARP)/diphteria toxin-type ADP-ribosyl transferase (ARTD) family [9], while other toxins belong to the cholera toxin-type, ARTC family. Plant pathogens produce several effectors with ART activity, able to disrupt the signaling pathway leading to plant immunity. For instance, Pseudomonas syringae HopU1, a Cholera toxin (C-type) ART with mono-ART activity, HopF2, a D-type ART, and HopM1 target plant proteins regulating plant signaling and immunity, such as MAPKKs, GTP Exchange Factors (GEF) and RNA binding proteins [16,17].
1.4. ADPRTs Modifying Nucleic Acids
ADP-ribosyl transferase reactions are also exploited by few classes of enzymes to modify DNA, such as pierisins in insects and CARP in shellfish, and DaRT, a ssDNA PARylating enzyme involved in SOS stress responses, that together with DarG, the macrodomain glycohydrolase, have a role in reversible DNA ADP-ribosylation, a toxin/antitoxin system in bacteria [18].
1.5. Sirtuins
The yeast sirtuin silent information regulator 2 (Sir2) originally described as a regulator of transcriptional silencing of mating-type loci, telomeres and ribosomal DNA, extends yeast lifespan, in a pathway different from calorie restriction. Sir2 was found as a NAD-dependent histone deacetylase (HDAC), acting as a negative regulator of chronological lifespan during yeast ageing. Nicotinamide supplementation showed a phenotype similar to SIR2 inactivation, modulating carbon metabolism and increasing respiration rate and respiratory reserve capacity [19].
It is recognised that sirtuins serve both as energy sensors and as transcriptional effectors by controlling the acetylation state of histones. The fungal sirtuin SirA uses NAD(+) as a co-substrate to deacetylate lysine 16 in histone H4, an epigenetic control that leads to target gene expression repression. Furthermore, sirtuins deacetylate also a wide range of transcriptional regulators (i.e., p53 protein), thereby controlling their activity. Sirtuins are classified according to their cellular localization (nuclear, intraorganellar and cytoplasmic), on the basis of their enzymatic activities and their protein targets. The sirtuin substrate, NAD+, is cleaved into nicotinamide and ADP-ribose, that is acetylated, removing the acetyl residue form the target protein.
Some sirtuin perform different enzymatic activities using NAD+ as a substrate: it is the case of sirtuins with ADP-ribosylating activity, producing MARylated proteins. This activity can be found in various organisms, from mammals to parasites. In Leishmania donovani, SIR2RP2 is an ADP-ribosylating sirtuin localised to mitochondria of promastigotes [20]. LdSIR2RP2null mutants had restrictive growth phenotype and were sensitised to sirtuin inhibitors.
In bacteria, in addition to ART toxins, sirtuin enzymes (SirTs) with ART activity are involved in the control of lypoate, an important cofactor. In Lactobacillales and Staphylococcaceae, the SirTM operon (coding for a sirtuin and for a macrodomain) is flanked by two ORFs, the glycine cleavage system H-like (GcvH-L), coding for proteins involved in glycine detoxification and one-carbon metabolism, and lipoate-protein ligase homolog (LplA2), coding for an enzyme scavenging the lipoate cofactor. GCVH-L is ADP ribosylated when in the lipoylated form [21]. The team led by Dr. Ahel showed that lipoylation of GcvH-L is a prerequisite for its MARylation. ADP-ribosylation of GcvH-L inhibits GcvH-L interaction with the oxidoreductase when it is not required. In order to reverse the post-translational modification, SirM macrodomain allows the recognition of the MARylated structure and the release of ADPr from the protein.
1.6. Viruses and Virulence
Viral infection in vertebrates involves the derangement of cellular functions such as the formation of the stress granule, where viruses exploit cellular machinery for their translation and replication. Several plus-strand RNA (+ssRNA) viruses, including alphaviruses and coronaviruses contain proteins with the macrodomain fold. Viral Macrodomain proteins bind to ADP-ribose and counteract ADP-ribosylation signals in host defence against viruses, altering the stress granule formation, and metabolism of ADP-ribose derivatives [2]. The non-structural protein 3 (nsp3) Macrodomain of Sindbis virus (SINV) is a virulence factor important for viral replication [2]. The nsp3 Macrodomain of murine coronavirus, Severe Acute Respiratory Syndrome (SARS) coronavirus and Chikungunya virus (CHIKV), are critical for virus replication and virulence [22].
1.7. NAD+ Recycling, Localized NAD+ Consumption, and Effects of Defective NAD+ Metabolism
Poly ADP ribose glycohydrolases (PARG) depolymerize the PAR branches in a cycle of PAR synthesis by ADPRT1/2, and rapid degradation by PARG, in most biological systems. Considering the rapid activation of ADPRT1 by DNA damage, and the high rate of polymerization of ADP-ribose branches, it is deduced that in few minutes the consecutive action of ADPRT and PARG reduces the local concentration of NAD+, and increases the ratio of nicotinamide/NAD. However, in the context of the subcellular compartment, NAM can behave as stimulator of SIRT1, thanks to the ability to activate the NAD+ salvage pathway [26].
When activated by DNA damage, PARP/ARTD1 catalyses the transfer of ADP-ribose units from NAD+ to substrate proteins to form branched polymers of ADP-ribose, leading to the decline of intracellular NAD+ levels. SIRT1 and PARPs compete for the intracellular NAD+ pool, thus a functional link between PARPs and SIRT1 has been proposed and verified.
ADPr and O-acetyl-ADP-ribose are a source of nicotinamide to be recycled into NAD+. Reactions dependent on NAD binding enzymes are affected by compounds with competitive binding, such as nicotinamide (Nam). It has been proposed that in cells stressed by excessive activation of NAD+ consuming enzymes or being involved in cell death pathways, or in necrosis, the consumption of NAD+ may be the trigger of ATP consumption and finally of cell death. Instead, it may be likely that localized storage of NAD+ may be shifted toward an excess of nicotinamide, that is a well known inhibitor of either ADPRT enzymes as well as of sirtuins, leading the cells to a block of their enzymatic activities, that may bring to a halt of their activity in a compartmentally localized way; local increase in nicotinamide. being a competitive inhibitor of ADPRTs and sirtuins, may block these two classes of enzymes.
After cleavage of ADPr moieties from the polymer, PAR degradation is a process that supports the restoration of NAD+ pool. Among the enzymes involved in nicotinamide/NAD+ cycling, nicotinamide mononucleotide (NMN) deamidase is a key player in bacterial pyridine nucleotide cycle, where it catalyzes the conversion of NMN into nicotinic acid mononucleotide (NaMN), which is converted to NAD+ in the Preiss-Handler pathway. Quinolinic acid is converted to nicotinic acid mononucleotide (NaMN) by transfer of a phosphoribose moiety. An adenylate moiety is then transferred to form nicotinic acid adenine dinucleotide (NaAD). Finally, the nicotinic acid moiety in NaAD is amidated to nicotinamide (Nam).
In eukaryotic cells, the NAD+ biosynthetic enzymes Nicotinamide mononucleotide adenylyl transferease (NMNAT-1) and nicotinamide/nicotinic acid phosphoribosyl transferase (NAMPT) form the nuclear NAD+ salvage pathway; the restoration of the NAD+ pool impacts and sustains SIRT1 activity [27,28,29]. Niacin (nicotinic acid), nicotinamide (Nam), nicotinamide riboside, NMN nicotinamide mononucleotide (NMN) and nicotinic acid mononucleotide (NaMN) are NAD+ precursors (see Figure 2).
PAR homeostasis involves nucleoside diphosphates linked to some moiety X (NUDIX, or NUDT) hydrolases, that cleave the substrate ADP-ribose to produce adenosine monophosphate (AMP) and ribose-5-phosphate (R5P). In Aspergillus nidulans, NdxA increases acetylation levels of histone H4. In a NdxA-deficient A. nidulans strain, impaired acetylation of histone and chromatin structure was found. Thus, NdxA controls total levels of NAD+, regulating sirtuin function [30].
Streptomyces as well as many bacteria utilize protein ADP-ribosylation, yet their enzymes that synthesize and remove this modification are not well known. Dr. Ahel’s colleagues have determined the crystal structure and characterized, both biochemically and functionally, the macrodomain protein SCO6735 from Streptomyces coelicolor, showing that it hydrolyzes PARP-dependent protein bound ADP-ribose polymers, is induced by DNA damage, and its deletion increases antibiotic production [31].
Nudix homology domains (NHD) proteins depend on NAD+ for activity: a conserved NAD+ binding pocket of DBC1 (deleted in breast cancer 1) prevents it from inhibiting PARP1 and DNA repair. Thus, NHD containing proteins regulate protein-protein interactions during aging, and the process can be slowed down by replenishing the NAD+ pool. When NAD+ concentrations decline, DBC1 is increasingly bound to PARP1, causing DNA damage to accumulate, a process rapidly reversed by restoring the abundance of NAD+ [32].
Bacterial Nudix hydrolase EcRppH and human HsNudT16 have been used to purify the ADP-ribose moiety from ADP ribosylated proteins in proteomic studies, leaving only a phosphoribose (pR) tag, even in presence of labile bonds such as in MARylated serine residues [33,34]. Bacterial Nudix hydrolase EcRppH and human HsNudT16 process the ADP-ribose chain through an alternative mechanism, converting it into protein-conjugated ribose-5′-phosphate [35]. The ectonucleotide pyrophosphatase/phosphodiesterase 1 (ENPP1) is a mammalian enzyme that generates ribose-5′-phosphate from ADP-ribose bound to proteins, either as protein-conjugated mono(ADP-ribose) and poly(ADP-ribose) [35]. These works showed that at least two enzyme families, Nudix and ENPP/NPP, are able to metabolize protein-conjugated ADP-ribose to pR in vitro, suggesting that pR exists and may be conserved from bacteria to mammals.
1.8. Non Enzymatic ADP-Ribose Attachment
ADP-ribose, the intermediate produced during the metabolism of mono- or poly-ADP ribosylated proteins, and cyclic-ADP ribose hydrolysis, is a protein-glycating agent, and excess levels of ADP-ribose in the cell can cause non-enzymatic ADP-ribosylation. Non-enzymatic ADP-ribosylation can inactivate protein targets that contain nucleotide-binding sites when the adenylate moiety of ADP-ribose binds to them, and it can also interfere with metabolic regulation that occurs via enzymatic ADP-ribosylation.
1.9. Ap4A and Oligoadenylates
Diadenosine tetraphosphate (Ap4A) is a second messenger and a putative ubiquitous alarmone, being present in all kingdoms of life, from bacteria to humans, capable of inducing various physiological effects in most organisms [36,37,38,39,40,41].
NUDIX homology domains (NHD) in enzymes and the Nudix type 2 gene product, Ap4A hydrolase, have been shown responsible for Ap4A degradation. The human fragile histidine triad (FHIT) is a tumor suppressor, whose Ap4A hydrolase activity is not required for antioncogene activity, while Ap4A binding has a positive effect [42,43,44,45,46,47]. The Microphthalmia transcription factor (MITF) is a basic helix-loop-helix leucine zipper (bHLH-Zip) DNA-binding protein. MITF transcriptional activity is inhibited by the histidine triad nucleotide-binding protein 1 (HINT1) through direct binding. The association is disrupted by the binding of Ap4A to HINT1 [44].
A pharmacological use of Ap4A in various disorders has been envisaged, such as in ocular tissue pathologies and retinal detachment, as well as in inhibition of ATP-induced excitotoxicity, as neuroprotection for traumatic spinal cord injury treatment [45,46,47]. A phosphodiesterase, NUDT16, can hydrolyze protein bound ADP-ribose, in vertebrate systems [35].
The 5′-triphosphorylated, 2′-5′-linked oligoadenylates polyribonucleotides (2-5As) are central to the interferon-induced antiviral 2-5A system, in vertebrate systems. Oligoadenylate synthases (OAS) are enzymes with a broad range of substrates, being thus described as 2′-5′ nucleotidyl transferases. An assay for synthesis of oligoadenylates (oligo2-5A) has been set up. The assays used dATP and the A(pA)3 tetramer core as substrates. In alternative, a synthetase can use dNTPs together with NAD+ as substrates [48]. Therefore, the Ap4A alarmone can be converted also into oligo2-5A.
1.10. Nicotinic Acid Adenine Dinucleotide Phosphate (NAADP) and Cyclic ADP Ribose (cADPr)
Extracellular NAD+ is used as substrate by a series of receptors on cell membranes. For instance, TRPM2 (melastatin-like transient receptor potential 2 channel), a ligand-gated Ca2+-permeable nonselective cation channel, possesses a NUDT9 Homology Domain that is activated by ADP-Ribose, 2′-deoxy-ADPR and by ADP-ribose-2′-phosphate [49,50,51,52].
The cluster of differentiation 38 (CD38), extracellular receptor for NAD+, known as cyclic ADP ribose (cADPR) hydrolase or NADase, is a glycoprotein of many immune cells. CD38 is a multifunctional enzyme that catalyzes the synthesis from NAD+ of cyclic ADP-ribose (cADPr) and its hydrolysis to ADP-ribose. The reaction products are essential for the regulation of intracellular Ca2+ [52,53]. CD38 activity is necessary for functions such as memory, amnesia, and autism: single nucleotide polymorphisms of CD38 alter the availability of cADPR, regulating brain oxytocin secretion. CD38 levels increase during aging, exerting a high NADase activity [52,53,54]. Mitochondrial depletion of NAD+ has been linked to ageing and various human diseases [8,55,56,57,58,59,60,61]. There are relatively independent pools of NAD+ in the nucleus, cytosol, and mitochondria [60] in eukaryotes. Camacho-Pereira, Chini and colleagues addressed the gerontological issue of the NAD+ levels fall in ageing. They showed that the major culprit is the rise in levels of the extracellular NADase CD38, that converts NAD+ into cyclic ADP ribose, a Ca2+ mobilising factor. CD38 affects NAD+ levels and this decreases SIRT3 activity in mitochondria, causing age-related metabolic decline [53]. Given that NADase activity has been detected on the plasma membrane and in mitochondria, it is not known which compartments affect the change in NAD+ levels during aging.
NAD+ depletion has been also linked to the activation of PARP1 and excessive PARylation following dioxin binding to the AhR nuclear receptor [56], followed by decreased SIRT6 activity: providing the cells with additional NAD+ or even the precursor, NAM, was sufficient to restore cell function and vitality [57]. Various roles have been assigned to O-acetyl-ADP-ribose, in particular in the assembly of the SIR complex [59,60]. NAD+ uptake in mammalian cells may be lead to important mechanisms of regulation, production of new secondary metabolites, and replenishment of this important cofactor and substrate [61].
Nicotinic acid adenine dinucleotide phosphate (NAADP) is a ubiquitous trigger for initiating and co-ordinating calcium signals and cyclic ADP-ribose regulates calcium-induced calcium release and transduction of Ca2+ signals [62,63].
In conclusion, ADPRT1/PARP1 is the main enzyme consuming NAD+ in the nuclei, in most biological systems examined, performing post-translational modifications on proteins, with a role in protein-protein interaction, assemblage of protein complexes, and cellular stress leading to cell death [64,65,66].
2. Plants: Role of NAD+ and PARP Domain Enzymes in Immune Response
In plants, three PARP proteins and several PARP domain proteins have been described, with a role in biotic stress response [69,70,71,72,73]. PARP-domain proteins, such as radical-induced cell death 1 (RCD1) and Similar to RCD One (SRO) proteins, have been shown to possess antiviral activity or to be involved in plant response to virus infection. In particular, plants’ response to ozone stress or reactive oxygen species (ROS) is dependent on radical cell death 1 (RCD1) and on Similar to RCD One (SRO) proteins to protect the plant from radicals [69,70,71], and their mutations lead to stress-induced morphogenetic response (SIMR), and to developmental defects. It is important to discriminate the PARP proteins necessary in plants to protect them from abiotic and biotic stress [72]: to this aim a triple PARP knockout Arabidopsis mutant was established [73]. Studying Arabidopsis, Rissel and colleagues showed that PARP domain proteins (RCD1, SRO1, and its paralogues) still contribute to protect the plant from biotic stress through PAR modification, while levels of protein PARylation even increased in the parp-/- triple mutant. Candidates for PARP-inhibitor targets may be found in the RCD1/SRO proteins family [73].
In plants, a mechanistic understanding of the role of ADP-ribosylation in stress response is still lacking, as well as in plant interaction with bacteria [74]. In 2016, Feng et al. identified the first set of non-histone plant PARylated proteins, showing that in vivo PARylation of DAWDLE (DDL) is important in plant immunity [75,76]. DDL, a Forkhead associated (FHA) domain protein, domain that may bind to PAR polymer and to phosphothreonine, is involved in plant immunity, in a pathway downstream to PARP-domain activity and upstream to activation of gene expression of defense-related proteins [76].
In plants there are also various sirtuins, shown to exert various functions necessary for survival, such as inhibition of glycolysis, and regulation of the heterofunctional GAPDH [77].
2.1. Extracellular NAD+ as DAMP Signal in the Apoplast: Plant NAD+ Receptors on Cell Membrane
Extracellular NAD+ is sensed in the apoplast as a DAMP, and is recognized by a NAD+ specific lectin-type receptor, LecRK-I.8, localized at the plasma membrane, possessing an active cytoplasmic kinase domain sensing NAD+ as extracellular ligand. The saturation curve for NAD+ binding showed a dissociation constant (Kd) of 436.5 ± 104.8 nM, which falls well below the extracellular NAD(H) concentration (~0.4 mM) in pathogen-infected leaf tissues, and thus indicates a relatively high affinity [78]. Binding assays with 32P-labeled NAD+ detected NAD+ binding activity for the immunoprecipitated eLecRK-I.8-GFP protein, showing NAD+ binding activity specific to eLecRK-I.8. Given the structural similarity between NAD+ and NADP+ and the fact that NADP+ can similarly induce immune responses in plants [79], various unlabeled nucleotides were tested in binding competition assay together with 32P-labeled NAD+: unlabeled NAD+ exhibited strong competition for binding while NADP+, ATP, ADP, and AMP showed no competition. LecRK-I.8 binds NAD+, but not NADP+, ATP, ADP, or AMP. Thus, LecRK-I.8 is very likely the eNAD+ receptor in Arabidopsis.
It has been reported that exogenous application of NAD+ in Arabidopsis enhanced disease resistance against Fusarium graminearum and the pathogen Pseudomonas syringae pv. maculicola Psm ES43269. Thus, pretreatment with NMN enhanced disease resistance against F. graminearum in Arabidopsis leaves and flowers. It was shown that Pseudomonas syringae pv. tomato PstDC3000, harboring the avirulence factor AvrRpt2, induce almost all the genes induced by NAD+ [80,81,82,83,84,85].
Sirtuins have also a great role in epigenetic regulation, in metabolism and in energy production [86].
In addition to NAD+ based nucleotides, in plants, such as in seeds, there are other condensation products, that may have NAD+ storage function: trigonelline, an alkaloid formed by the methylation of the nitrogen atom of nicotinamide, i.e., methyl betaine of nicotinic acid, present in fenugreek, in seeds of several plants, and in coffee [87]. Furthermore, minor mononucleotides, such as adenosine-5′-tetraphosphate (p4A) and adenosine-5′-pentaphosphate (p5A), and dinucleotides, such as diadenosine-5′,5′′′-P1, P3-triphosphate (Ap3A) and diadenosine-5′,5′′′-P1, P4-tetraphosphate (Ap4A), are present in plants, as well as the enzymes synthesizing and those involved in their degradation (NUDT hydrolases) [39].
The Nudix hydrolase NUDX (or NUDT) family proteins also play an important role in fine-tuning plant immune responses. In Arabidopsis, several NUDX genes affect plant response to stress. AtNUDX7 negatively regulates plant Pattern Triggered Immunity (PTI) and effector triggered immunity (ETI). AtNUDX6 and AtNUDX7 have distinct roles in stress responses and accumulation of plant defense hormone salicylic acid (SA) and its induced genes [88]. AtNUDX7 is involved in the response to oxidative stress [88], while AtNUDX6 positively regulates salicylic acid signaling and activity of the Transcription Factor NPR1 [89], a global regulator of hormone signaling and defense responses. Depending on the NUDX considered, the effective enzymatic activity can either be NAD+-hydrolysing or PAR-hydrolysing and might influence NADH/NAD+ ratio or PAR content. AtNUDX2 protein has pyrophosphatase activities towards both ADP-ribose and NADH.
In support of the importance of NAD+ in plant immunity, it was shown that the effector AvrRxo1 phosphorylates NAD deactivating the immune response in plants [90].
2.2. Bacterial Effectors, PARP Domain Proteins, NAD+ and Plant Immunity
The first layer of plant innate immunity consists in the recognition of microbial fingerprints, called pathogen/microbe associated molecular patterns (PAMPs/MAMPs), by a set of inherited receptors, Pattern recognition receptors (PRR). In case fingerprints responsible of elicitation comes from the host, the molecules are referred as Damage Associated Molecular Patterns (DAMPs).
It was shown that upregulating NAD+ levels by expression of a bacterial nadC quinolinate phosphoribosyl transferase catalysing conversion of quinolinate to NaMN, enabled the plants to resist their pathogens [84]. Pétriacq and colleagues used the nadC gene derived from E. coli expressed in Arabidopsis; they reported that higher NAD+ contents in nadC-overexpressing plants with the addition of quinolinate activated the plant immune response and resulted in disease resistance against a virulent bacterial pathogen, P, syringae pv. tomato, harboring the Pst-AvrRpm1 avirulence factor [85]. Therefore, the accumulation of NAD+ and related metabolites induces disease resistance against bacterial phytopathogens. NMN was found to be enriched in the uninoculated barley spikes of the two resistant cultivars compared with the susceptible cultivars. The amount of NAD+ significantly increased in barley leaves inoculated with the biotrophic fungal pathogen Erysiphe graminis, the causal organism of powdery mildew [84].
High NAD+ pools are beneficial for plant immunity by stimulating SA-mediated signaling and pathogen resistance. NAD+ promoted the induction of various pathogen-related genes typified by the salicylic acid (SA)-responsive defense marker PR1. Extensive comparison with publicly available transcriptomic databases further showed that gene expression under high NAD+ content was similar to that obtained under biotic stress, elicitor challenge or SA treatment.
2.3. Hypothesis for the Role of NAD+: Convergence of Signaling Pathways in Plant Systems
Recently, the priming of plants to increase the immune response and basal defense against biotic stress has progressed quickly, showing that the response of plants to ß-aminobutyric acid (BABA) is mediated by the BABA receptor, BABA-induced Immunity 1 (IBI1) gene, coding for an aspartate tRNA synthase, AspRS [91,92,93,94,95]. The signaling downstream of BABA sensing involves the induction of SIMR and plant growth inhibition, with expression of defense related genes. R-BABA, by blocking L-asp-IBI1 binding, thus prevents a stress response pathway that require the PARP domain proteins SRO and RCD-1, and their downstream signaling. RCD-1 and SRO mutant plants showed growth inhibition and SIMR phenotype [91,92,93,94,95]. Is this just a casual intersection of two differently regulated pathways? Or, is this an interaction of signals that exists between BABA signaling translated into a SIMR phenotype, and the SIMR-antagonistic, protective phenotype caused by PARP domain RCD-1 and SRO proteins? It could be possible that the role of RCD-1 and SRO in protection from SIMR requires NAD+ as substrate, or on the contrary the activity of PARP domain proteins could be independent from enzymatic transformation of NAD+ in nicotinamide and ADP-ribose.
3. Conclusions
NAD+ is a cofactor for enzymes involved in a series of reactions in all kingdoms of life. The cycling form MARylation and PARylation (of proteins and nucleic acids), leading to their catabolism into O-acetyl-ADP-ribose and ribose-5′-phosphate (R5P), and the restoration of NAD pool through different biosynthesis pathway, is conserved in most of the organisms examined, with the exception of S. cerevisiae. It is envisaged that other important functions will be brought to light in the coming years.
Author Contributions
Palmiro Poltronieri conceived the study of ADPRT sources in various kingdoms of life, Nataša Čerekovic reviewed the manuscript and elaborated on plant proteins and role in immunity. We thank Masanao Miwa for this precious assistance and enlightening insights.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Poltronieri, P.; Miwa, M. Overview on ADP ribosylation and PARP superfamily of proteins. Curr. Protein Pept. Sci. 2016, 17, 630–632. [Google Scholar] [CrossRef] [PubMed]
- Poltronieri, P. ADP ribosylation reactions in animals, plants and bacteria. Challenges 2017, 8, 14. [Google Scholar] [CrossRef]
- Bürkle, A.; Virág, L. Poly(ADP-ribose): PARadigms and PARadoxes. Mol. Asp. Med. 2013, 34, 1046–1065. [Google Scholar] [CrossRef] [PubMed]
- Miwa, M.; Ida, C.; Yamashita, S.; Tanaka, M.; Fujisawa, J. Poly(ADP-ribose): Structure, physicochemical properties and quantification in vivo, with special reference to Poly(ADP-ribose) binding protein modules. Curr. Protein Pept. Sci. 2016, 17, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Verheugd, P.; Bütepage, M.; Eckei, L.; Lüscher, B. Players in ADP-ribosylation: Readers and Erasers. Curr. Protein Pept. Sci. 2016, 17, 654–667. [Google Scholar] [CrossRef] [PubMed]
- Fouquerel, E.; Sobol, R.W. ARTD1 (PARP1) activation and NAD(+) in DNA repair and cell death. DNA Repair 2014, 23, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Bock, F.J.; Chang, P. Stress response. PARP1 911. Nat. Chem. Biol. 2015, 11, 179–180. [Google Scholar] [CrossRef] [PubMed]
- Faraone-Mennella, M.R. A new facet of ADP-ribosylation reactions: SIRTs and PARPs interplay. Front. Biosci. 2015, 20, 458–473. [Google Scholar] [CrossRef]
- Aravind, L.; Zhang, D.; de Souza, R.F.; Anand, S.; Iyer, L.M. The natural history of ADP-ribosyltransferases and the ADP-ribosylation system. Curr. Top. Microbiol. Immunol. 2015, 384, 3–32. [Google Scholar] [PubMed]
- Timinszky, G.; Till, S.; Hassa, P.O.; Hothorn, M.; Kustatscher, G.; Nijmeijer, B.; Colombelli, J.; Altmeyer, M.; Stelzer, E.H.; Scheffzek, K.; et al. A macrodomain-containing histone rearranges chromatin upon sensing PARP1 activation. Nat. Struct. Mol. Biol. 2009, 16, 923–929. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Jividen, K.; Spencer, A.; Dworak, N.; Ni, L.; Oostdyk, L.T.; Chatterjee, M.; Kusmider, B.; Reon, B.; Parlak, M.; Gorbunova, V.; Abbas, T.; Jeffery, E.; Sherman, N.E.; Paschal, B.M. Ubiquitin modification by the E3 Ligase/ADP-Ribosyltransferase Dtx3L/Parp9. Mol. Cell 2017, 66, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Schmoll, M.; Dattenbock, C.; Carreras-Villasenor, N.; Mendoza-Mendoza, A.; Tisch, D.; Alemán, M.I.; Baker, S.E.; Brown, C.; Cervantes-Badillo, M.G.; Cetz-Chel, J.; et al. The genomes of three uneven siblings: Footprints of the lifestyles of three Trichoderma species. Microbiol. Mol. Biol. Rev. 2016, 80, 205–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rissel, D.; Heym, P.P.; Peiter, E. A yeast growth assay to characterize plant poly(ADP-ribose) polymerase (PARP) proteins and inhibitors. Anal. Biochem. 2017, 527, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Gao, P.; Liu, H.W. Studies of the expression of human poly(ADP-ribose) polymerase-1 in Saccharomyces cerevisiae and identification of PARP-1 substrates by yeast proteome microarray screening. Biochemistry 2009, 48, 11745–11754. [Google Scholar] [CrossRef] [PubMed]
- La Ferla, M.; Mercatanti, A.; Rocchi, G.; Lodovichi, S.; Cervelli, T.; Pignata, L.; Caligo, M.A.; Galli, A. Expression of human poly (ADP-ribose) polymerase 1 in Saccharomyces cerevisiae: Effect on survival, homologous recombination and identification of genes involved in intracellular localization. Mutat. Res. 2015, 774, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Lu, D.; Kabbage, M.; Wei, H.L.; Swingle, B.; Records, A.R.; Dickman, M.; He, P.; Shan, L. Bacterial effector HopF2 suppresses Arabidopsis innate immunity at the plasma membrane. Mol. Plant Microbe Interact. 2011, 24, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Durán, R.; Bourdais, G.; He, S.Y.; Robatzek, S. The bacterial effector HopM1 suppresses PAMP-triggered oxidative burst and stomatal immunity. New Phytol. 2014, 202, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Jankevicius, G.; Ariza, A.; Ahel, M.; Ahel, I. The Toxin-Antitoxin system DarTG catalyzes reversible ADP-Ribosylation of DNA. Mol. Cell 2016, 64, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Orlandi, I.; Pellegrino Coppola, D.; Strippoli, M.; Ronzulli, R.; Vai, M. Nicotinamide supplementation phenocopies SIR2 inactivation by modulating carbon metabolism and respiration during yeast chronological aging. Mech. Ageing Dev. 2017, 161, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Mittal, N.; Muthuswami, R.; Madhubala, R. The mitochondrial SIR2 related protein 2 (SIR2RP2) impacts Leishmania donovani growth and infectivity. PLoS Negl. Trop. Dis. 2017, 11, e0005590. [Google Scholar] [CrossRef] [PubMed]
- Rack, J.G.M.; Morra, R.; Barkauskaite, E.; Kraehenbuehl, R.; Ariza, A.; Qu, Y.; Ortmayer, M.; Leidecker, O.; Cameron, D.R.; Matic, I.; et al. Identification of a class of protein ADP-ribosylating sirtuins in microbial pathogens. Mol. Cell 2015, 59, 309–320. [Google Scholar] [CrossRef] [PubMed]
- McPherson, R.L.; Abraham, R.; Sreekumar, E.; Ong, S.E.; Cheng, S.J.; Baxter, V.K.; Kistemaker, H.A.; Filippov, D.V.; Griffin, D.E.; Leung, A.K. ADP-ribosylhydrolase activity of Chikungunya virus macrodomain is critical for virus replication and virulence. Proc. Natl. Acad. Sci. USA 2017, 114, 1666–1671. [Google Scholar] [CrossRef] [PubMed]
- Steffen, J.D.; Pascal, J.M. New players to the field of ADP-ribosylation make the final cut. EMBO J. 2013, 32, 1225–1237. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, C.; Song, Y.; Shao, C.; Zhang, X.; Zang, J. Structural insights into the mechanism of Escherichia coli YmdB: A 2’-O-acetyl-ADP-ribose deacetylase. J. Struct. Biol. 2015, 192, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Haikarainen, T.; Lehtiö, L. Proximal ADP-ribose hydrolysis in Trypanosomatids is catalyzed by a macrodomain. Sci. Rep. 2016, 6, 24213. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S.; Song, S.B. Nicotinamide is an inhibitor of SIRT1 in vitro, but can be a stimulator in cells. Cell Mol. Life Sci. 2017, 74, 3347–3362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Berrocal, J.G.; Frizzell, K.M.; Gamble, M.J.; DuMond, M.E.; Krishnakumar, R.; Yang, T.; Sauve, A.A.; Kraus, W.L. Enzymes in the NAD+ salvage pathway regulate SIRT1 activity at target gene promoters. J. Biol. Chem. 2009, 284, 20408–20417. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Berrocal, J.G.; Yao, J.; DuMond, M.E.; Krishnakumar, R.; Ruhl, D.D.; Ryu, K.W.; Gamble, M.J.; Kraus, W.L. Regulation of poly(ADP-ribose) polymerase-1-dependent gene expression through promoter-directed recruitment of a nuclear NAD+ synthase. J. Biol. Chem. 2012, 287, 12405–12416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Kraus, W.L. SIRT1-dependent regulation of chromatin and transcription: Linking NAD(+) metabolism and signaling to the control of cellular functions. Biochim. Biophys. Acta 2010, 1804, 1666–1675. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Masuo, S.; Fujita, T.; Doi, Y.; Kamimura, Y.; Takaya, N. Hydrolase controls cellular NAD, sirtuin, and secondary metabolites. Mol. Cell Biol. 2012, 32, 3743–3755. [Google Scholar] [CrossRef] [PubMed]
- Lalić, J.; PosavecMarjanović, M.; Palazzo, L.; Perina, D.; Sabljić, I.; Žaja, R.; Colby, T.; Pleše, B.; Halasz, M.; Jankevicius, G.; et al. Disruption of Macrodomain protein SCO6735 increases antibiotic production in Streptomyces coelicolor. J. Biol. Chem. 2016, 291, 23175–23187. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Bonkowski, M.S.; Moniot, S.; Zhang, D.; Hubbard, B.P.; Ling, A.J.; Rajman, L.A.; Qin, B.; Lou, Z.; Gorbunova, V.; et al. A conserved NAD+ binding pocket that regulates protein-protein interactions during aging. Science 2017, 355, 1312–1317. [Google Scholar] [CrossRef] [PubMed]
- Daniels, C.M.; Thirawatananond, P.; Ong, S.E.; Gabelli, S.B.; Leung, A.K. Nudix hydrolases degrade protein-conjugated ADP-ribose. Sci. Rep. 2015, 5, 18271. [Google Scholar] [CrossRef] [PubMed]
- Bonfiglio, J.J.; Colby, T.; Matic, I. Mass spectrometry for serine ADP-ribosylation? Think o-glycosylation! Nucleic Acids Res. 2017, 45, 6259–6264. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, L.; Daniels, C.M.; Nettleship, J.E.; Rahman, N.; McPherson, R.L.; Ong, S.E.; Kato, K.; Nureki, O.; Leung, A.K.; Ahel, I. ENPP1 processes protein ADP-ribosylation in vitro. FEBS J. 2016, 283, 3371–3388. [Google Scholar] [CrossRef] [PubMed]
- Alderson, T. Ribonucleotide metabolism: Fresh approaches to viral and cancer chemotherapy. Biol. Rev. 1989, 64, 159–196. [Google Scholar] [CrossRef] [PubMed]
- Carmi-Levy, I.; Yannay-Cohen, N.; Kay, G.; Razin, E.; Nechushtan, H. Diadenosine tetraphosphate hydrolase is part of the transcriptional regulation network in immunologically activated mast cells. Mol. Cell. Biol. 2008, 28, 5777–5784. [Google Scholar] [CrossRef] [PubMed]
- Monds, R.D.; Newell, P.D.; Wagner, J.C.; Schwartzman, J.A.; Lu, W.; Rabinowitz, J.D.; O’Toole, G.A. Di-adenosine tetraphosphate (Ap4A) metabolism impacts biofilm formation by Pseudomonas fluorescens via modulation of c-di-GMP-dependent pathways. J. Bacteriol. 2010, 192, 3011–3023. [Google Scholar] [CrossRef] [PubMed]
- Guranowski, A. Metabolism of diadenosine tetraphosphate (Ap4A) and related nucleotides in plants; review with historical and general perspective. Front. Biosci. 2004, 9, 1398–1411. [Google Scholar] [CrossRef] [PubMed]
- Despotovic, D.; Brandis, A.; Savidor, A.; Levin, Y.; Fumagalli, L.; Tawfik, D.S. Diadenosine tetraphosphate (Ap4A)—An E. coli alarmone or a damage metabolite? FEBS J. 2017, 284, 2194–2215. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.N.; Nechushtan, H.; Figov, N.; Razin, E. The function of lysyl-tRNA synthetase and Ap4A as signaling regulators of MITF activity in FcepsilonRI-activated mast cells. Immunity 2004, 20, 145–151. [Google Scholar] [CrossRef]
- Huebner, K.; Saldivar, J.C.; Sun, J.; Shibata, H.; Druck, T. Hits, Fhits and Nits: Beyond enzymatic function. Adv. Enzyme Regul. 2011, 51, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Brenner, C. Hint, Fhit, and GalT: Function, structure, evolution, and mechanism of three branches of the histidine triad superfamily of nucleotide hydrolases and transferases. Biochemistry 2002, 41, 9003–9014. [Google Scholar] [CrossRef] [PubMed]
- Motzik, A.; Amir, E.; Erlich, T.; Wang, J.; Kim, B.G.; Han, J.M.; Kim, J.H.; Nechushtan, H.; Guo, M.; Razin, E.; et al. Post-translational modification of HINT1 mediates activation of MITF transcriptional activity in human melanoma cells. Oncogene 2017, 36, 4732–4738. [Google Scholar] [CrossRef] [PubMed]
- Marriott, A.S.; Vasieva, O.; Fang, Y.; Copeland, N.A.; McLennan, A.G.; Jones, N.J. NUDT2 disruption elevates diadenosine tetraphosphate (Ap4A) and down-regulates immune response and cancer promotion genes. PLoS ONE 2016, 11, e0154674. [Google Scholar] [CrossRef] [PubMed]
- Crooke, A.; Guzman-Aranguez, A.; Carracedo, G.; de Lara, M.J.P.; Pintor, J. Understanding the presence and roles of Ap4A (Diadenosine tetraphosphate) in the eye. J. Ocul. Pharmacol. Ther. 2017, 33, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Reigada, D.; Navarro-Ruiz, R.M.; Caballero-López, M.J.; Del Águila, Á.; Muñoz-Galdeano, T.; Maza, R.M.; Nieto-Díaz, M. Diadenosine tetraphosphate (Ap4A) inhibits ATP-induced excitotoxicity: A neuroprotective strategy for traumatic spinal cord injury treatment. Purinergic Signal. 2017, 13, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, J.B.; Kjær, K.H.; Justesen, J.; Martensen, P.M. Enzyme assays for synthesis and degradation of 2-5As and other 2’-5’ oligonucleotides. BMC Biochem. 2015, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Tóth, B.; Iordanov, I.; Csanády, L. Ruling out pyridine dinucleotides as true TRPM2 channel activators reveals novel direct agonist ADP-ribose-2’-phosphate. J. Gen. Physiol. 2015, 145, 419–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kühn, F.; Kühn, C.; Lückhoff, A. Different principles of ADP-ribose-mediated activation and opposite roles of the NUDT9 Homology Domain in the TRPM2 orthologs of man and sea anemone. Front. Physiol. 2017, 8, 879. [Google Scholar] [CrossRef] [PubMed]
- Di, A.; Kiya, T.; Gong, H.; Gao, X.; Malik, A.B. Role of the phagosomal redox-sensitive TRP channel TRPM2 in regulating bactericidal activity of macrophages. J. Cell Sci. 2017, 130, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Higashida, H.; Yokoyama, S.; Huang, J.J.; Liu, L.; Ma, W.J.; Akther, S.; Higashida, C.; Kikuchi, M.; Minabe, Y.; Munesue, T. Social memory, amnesia, and autism: Brain oxytocin secretion is regulated by NAD+ metabolites and single nucleotide polymorphisms of CD38. Neurochem. Int. 2012, 61, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Pereira, J.; Tarragó, M.G.; Chini, C.C.; Nin, V.; Escande, C.; Warner, G.M.; Puranik, A.S.; Schoon, R.A.; Reid, J.M.; Galina, A.; Chini, E.N. CD38 dictates age-related NAD decline and mitochondrial dysfunction through an SIRT3-dependent mechanism. Cell Metab. 2016, 23, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Mendelsohn, A.R.; Larrick, J.W. The NAD+/PARP1/SIRT1 Axis in Aging. Rejuvenation Res. 2017, 20, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Dölle, C.; Rack, J.G.; Ziegler, M. NAD and ADP-ribose metabolism in mitochondria. FEBS J. 2013, 280, 3530–3541. [Google Scholar] [CrossRef] [PubMed]
- Diani-Moore, S.; Shoots, J.; Singh, R.; Zuk, J.B.; Rifkind, A.B. NAD+ loss, a new player in AhR biology: Prevention of thymus atrophy and hepatosteatosis by NAD+ repletion. Sci. Rep. 2017, 7, 2268. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yang, T.; Baur, J.A.; Perez, E.; Matsui, T.; Carmona, J.J.; Lamming, D.W.; Souza-Pinto, N.C.; Bohr, V.A.; Rosenzweig, A.; et al. Nutrient-sensitive mitochondrial NAD+ levels dictate cell survival. Cell 2007, 130, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Martín-Guerrero, S.M.; Muñoz-Gámez, J.A.; Carrasco, M.-C.; Salmerón, J.; Martín-Estebané, M.; Cuadros, M.A.; Martín-Oliva, D. Poly(ADP-ribose)polymerases inhibitors prevent early mitochondrial fragmentation and hepatocyte cell death induced by H2O2. PLoS ONE 2017, 12, e0187130. [Google Scholar] [CrossRef] [PubMed]
- Liou, G.G.; Tanny, J.C.; Kruger, R.G.; Walz, T.; Moazed, D. Assembly of the SIR complex and its regulation by O-acetyl-ADP-ribose, a product of NAD-dependent histone deacetylation. Cell 2005, 121, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Denu, J.M. Function and metabolism of sirtuin metabolite O-acetyl-ADP-ribose. Biochim. Biophys. Acta 2010, 1804, 1617–1625. [Google Scholar] [CrossRef] [PubMed]
- Billington, R.A.; Travelli, C.; Ercolano, E.; Galli, U.; Roman, C.B.; Grolla, A.A.; Canonico, P.L.; Condorelli, F.; Genazzani, A.A. Characterization of NAD uptake in mammalian cells. J. Biol. Chem. 2008, 283, 6367–6374. [Google Scholar] [CrossRef] [PubMed]
- Calcraft, P.J.; Ruas, M.; Pan, Z.; Cheng, X.; Arredouani, A.; Hao, X.; Tang, J.; Rietdorf, K.; Teboul, L.; Chuang, K.-T.; et al. NAADP mobilizes calcium from acidic organelles through two-pore channels. Nature 2009, 459, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Virág, L.; Robaszkiewicz, A.; Rodriguez-Vargas, J.M.; Oliver, F.J. Poly(ADP-ribose) signaling in cell death. Mol. Asp. Med. 2013, 34, 1153–1167. [Google Scholar] [CrossRef] [PubMed]
- Henderson, D.J.P.; Miranda, J.L.; Emerson, B.M. The ß-NAD salvage pathway and PKC-mediated signaling influence localized PARP-1 activity and CTCF poly(ADP) ribosylation. Oncotarget 2017, 8, 64698–64713. [Google Scholar] [CrossRef] [PubMed]
- Krietsch, J.; Rouleau, M.; Pic, É.; Ethier, C.; Dawson, T.M.; Dawson, V.L.; Masson, J.Y.; Poirier, G.G.; Gagné, J.P. Reprogramming cellular events by poly(ADP-ribose)-binding proteins. Mol. Asp. Med. 2013, 34, 1066–1087. [Google Scholar] [CrossRef] [PubMed]
- Till, S.; Ladurner, A.G. Sensing NAD metabolites through macrodomains. Front. Biosci. 2009, 14, 3246–3258. [Google Scholar] [CrossRef]
- Nakano, T.; Takahashi-Nakaguchi, A.; Yamamoto, M.; Watanabe, M. Pierisins and CARP-1: ADP-ribosylation of DNA by ARTCs in butterflies and shellfish. In Endogenous ADP-Ribosylation; Koch-Nolte, F., Ed.; Springer International Publishing: New York, NY, USA, 2015; pp. 127–149. [Google Scholar]
- Watanabe, M.; Kono, T.; Matsushima-Hibiya, Y.; Kanazawa, T.; Nishisaka, N.; Kishimoto, T.; Koyama, K.; Sugimura, T.; Wakabayashi, K. Molecular cloning of an apoptosis-inducing protein, pierisin, from cabbage butterfly: Possible involvement of ADP-ribosylation in its activity. Proc. Natl. Acad. Sci. USA 1999, 96, 10608–10613. [Google Scholar] [CrossRef] [PubMed]
- Lamb, R.S.; Citarelli, M.; Teotia, S. Functions of the poly(ADP-ribose) polymerase superfamily in plants. Cell. Mol. Life Sci. 2012, 69, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Jaspers, P.; Overmyer, K.; Wrzaczek, M.; Vainonen, J.P.; Blomster, T.; Salojärvi, J.; Reddy, R.A.; Kangasjärvi, J. The RST and PARP-like domain containing SRO protein family: Analysis of protein structure, function and conservation in land plants. BMC Genom. 2010, 11, 170. [Google Scholar] [CrossRef] [PubMed]
- Lamb, R.S. Abiotic stress responses in plants: A focus on the SRO family. In Selected Plant Physiology Aspects; Montanaro, G., Ed.; InTech: Rijeka, Croatia, 2010; pp. 3–22. [Google Scholar]
- Adams-Phillips, L.; Briggs, A.G.; Bent, A.F. Disruption of poly(ADP-ribosyl)ation mechanisms alters responses of Arabidopsis thaliana to biotic stress. Plant Physiol. 2010, 152, 267–280. [Google Scholar] [CrossRef] [PubMed]
- Rissel, D.; Heym, P.P.; Thor, K.; Brandt, W.; Wessjohann, L.A.; Peiter, E. No silver bullet—Canonical Poly(ADP-Ribose) Polymerases (PARPs) are no universal factors of biotic and biotic stress resistance of Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Liu, C.; Shan, L.; He, P. Protein ADP-ribosylation takes control in plant-bacterium interactions. PLoS Pathog. 2016, 12, e1005941. [Google Scholar] [CrossRef] [PubMed]
- Menke, F.L.H. Plants get on PAR with poly(ADP-ribosyl)ation. EMBO Rep. 2016, 17, 1677–1678. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Ma, S.; Chen, S.; Zhu, N.; Zhang, S.; Yu, B.; Yu, Y.; Le, B.; Chen, X.; Dinesh-Kumar, S.P.; et al. PARylation of the forkhead-associated domain protein DAWDLE regulates plant immunity. EMBO Rep. 2016, 17, 1799–1813. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Mou, Z. Extracellular pyridine nucleotides induce PR gene expression and disease resistance in Arabidopsis. Plant J. 2009, 57, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhou, M.; Zhang, X.; Yao, J.; Zhang, Y.; Mou, Z. A lectin receptor kinase as a potential sensor for extracellular nicotinamide adenine dinucleotide in Arabidopsis thaliana. eLife 2017, 6, e25474. [Google Scholar] [CrossRef] [PubMed]
- Pétriacq, P.; Bont, L.; Tcherkez, G.; Gakière, B. Not just a pawn on the board of plant-pathogen interaction. Plant Signal. Behav. 2013, 8, e22477. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Queval, G.; Gakiere, B. NAD(P) synthesis and pyridine nucleotide cycling in plants and their potential importance in stress conditions. J. Exp. Bot. 2006, 57, 1603–1620. [Google Scholar] [CrossRef] [PubMed]
- Pétriacq, P.; Tcherkez, G.; Gakière, B. Pyridine nucleotides induce changes in cytosolic pools of calcium in Arabidopsis. Plant Signal. Behav. 2016, 11, e1249082. [Google Scholar] [CrossRef] [PubMed]
- Pétriacq, P.; de Bont, L.; Hager, J.; Didierlaurent, L.; Mauve, C.; Guérard, F.; Noctor, G.; Pelletier, S.; Renou, J.P.; Tcherkez, G.; Gakière, B. Inducible NAD overproduction in Arabidopsis alters metabolic pools and gene expression correlated with increased salicylate content and resistance to Pst-AvrRpm1. Plant J. 2012, 70, 650–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pétriacq, P.; Ton, J.; Patrit, O.; Tcherkez, G.; Gakière, B. NAD acts as an integral regulator of multiple defense layers. Plant Physiol. 2016, 172, 1465–1479. [Google Scholar] [CrossRef] [PubMed]
- Miwa, A.; Sawada, Y.; Tamaoki, D.; Yokota Hirai, M.; Kimura, M.; Sato, K.; Nishiuchi, T. Nicotinamide mononucleotide and related metabolites induce disease resistance against fungal phytopathogens in Arabidopsis and barley. Sci. Rep. 2017, 7, 6389. [Google Scholar] [CrossRef] [PubMed]
- Adams-Phillips, L.; Wan, J.; Tan, X.; Dunning, F.M.; Meyers, B.C.; Michelmore, R.W.; Bent, A.F. Discovery of ADP-ribosylation and other plant defense pathway elements through expression profiling of four different Arabidopsis-Pseudomonas R-avr interactions. Mol. Plant Microbe Interact. 2008, 21, 646–665. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, Y.; Zhou, D.X. Rice NAD+-dependent histone deacetylase OsSRT1 represses glycolysis and regulates the moonlighting function of GAPDH as a transcriptional activator of glycolytic genes. Nucleic Acids Res. 2017, 45, 12241–12255. [Google Scholar] [CrossRef] [PubMed]
- Ashihara, H.; Ludwig, I.A.; Katahira, R.; Yokota, T.; Fujimura, T.; Crozier, A. Trigonelline and related nicotinic acid metabolites: Occurrence, biosynthesis, taxonomic considerations, and their roles in planta and in human health. Phytochem. Rev. 2015, 14, 765–798. [Google Scholar] [CrossRef]
- Ishikawa, K.; Ogawa, T.; Hirosue, E.; Nakayama, Y.; Harada, K.; Fukusaki, E.; Yoshimura, K.; Shigeoka, S. Modulation of the poly(ADP-ribosyl)ation reaction via the Arabidopsis ADP-ribose/NADH pyrophosphohydrolase, AtNUDX7, is involved in the response to oxidative stress. Plant Physiol. 2009, 151, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.; Yoshimura, K.; Harada, K.; Fukusaki, E.; Ogawa, T.; Tamoi, M.; Shigeoka, S. AtNUDX6, an ADP-ribose/NADH pyrophosphohydrolase in Arabidopsis, positively regulates NPR1-dependent salicylic acid signaling. Plant Physiol. 2010, 152, 2000–2012. [Google Scholar] [CrossRef] [PubMed]
- Shidore, T.; Broeckling, C.D.; Kirkwood, J.S.; Long, J.J.; Miao, J.; Zhao, B.; Leach, J.E.; Triplett, L.R. The effector AvrRxo1 phosphorylates NAD in planta. PLoS Pathog. 2017, 13, e1006442. [Google Scholar] [CrossRef] [PubMed]
- Luna, E.; van Hulten, M.; Zhang, Y.; Berkowitz, O.; López, A.; Pétriacq, P.; Sellwood, M.A.; Chen, B.; Burrell, M.; van de Meene, A.; et al. Plant perception of β-aminobutyric acid is mediated by an aspartyl-tRNAsynthetase. Nat. Chem. Biol. 2014, 10, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Baccelli, I.; Glauser, G.; Mauch-Mani, B. The accumulation of β-aminobutyric acid is controlled by the plant’s immune system. Planta 2017. [Google Scholar] [CrossRef] [PubMed]
- Thevenet, D.; Pastor, V.; Baccelli, I.; Balmer, A.; Vallat, A.; Neier, R.; Glauser, G.; Mauch-Mani, B. The priming molecule β-aminobutyric acid is naturally present in plants and is induced by stress. New Phytol. 2017, 213, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Ton, J.; Mauch-Mani, B. Beta-amino-butyric acid-induced resistance against necrotrophic pathogens is based on ABA-dependent priming for callose. Plant J. 2004, 38, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbacher, R.E.; Luna, E.; Ton, J. The discovery of the BABA receptor: Scientific implications and application potential. Front. Plant Sci. 2014, 5, 304. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Scheme of the binding to MAR and PAR regions by different reader proteins: Macrodomain (recognising the terminal ADP ribose), WWE, PBZ (binding to ADP ribose-ADP ribose junctions), PBM and FHA (recognising the ribose-diphosphate-ribose adenine region), OB fold, BRCT, and PIN domain, RRM, SR, KR and RGG repeats: docking to mono and poly ADP ribose regions. Figure modified from Verheugd et al., 2016 [5].
Figure 1.
Scheme of the binding to MAR and PAR regions by different reader proteins: Macrodomain (recognising the terminal ADP ribose), WWE, PBZ (binding to ADP ribose-ADP ribose junctions), PBM and FHA (recognising the ribose-diphosphate-ribose adenine region), OB fold, BRCT, and PIN domain, RRM, SR, KR and RGG repeats: docking to mono and poly ADP ribose regions. Figure modified from Verheugd et al., 2016 [5].

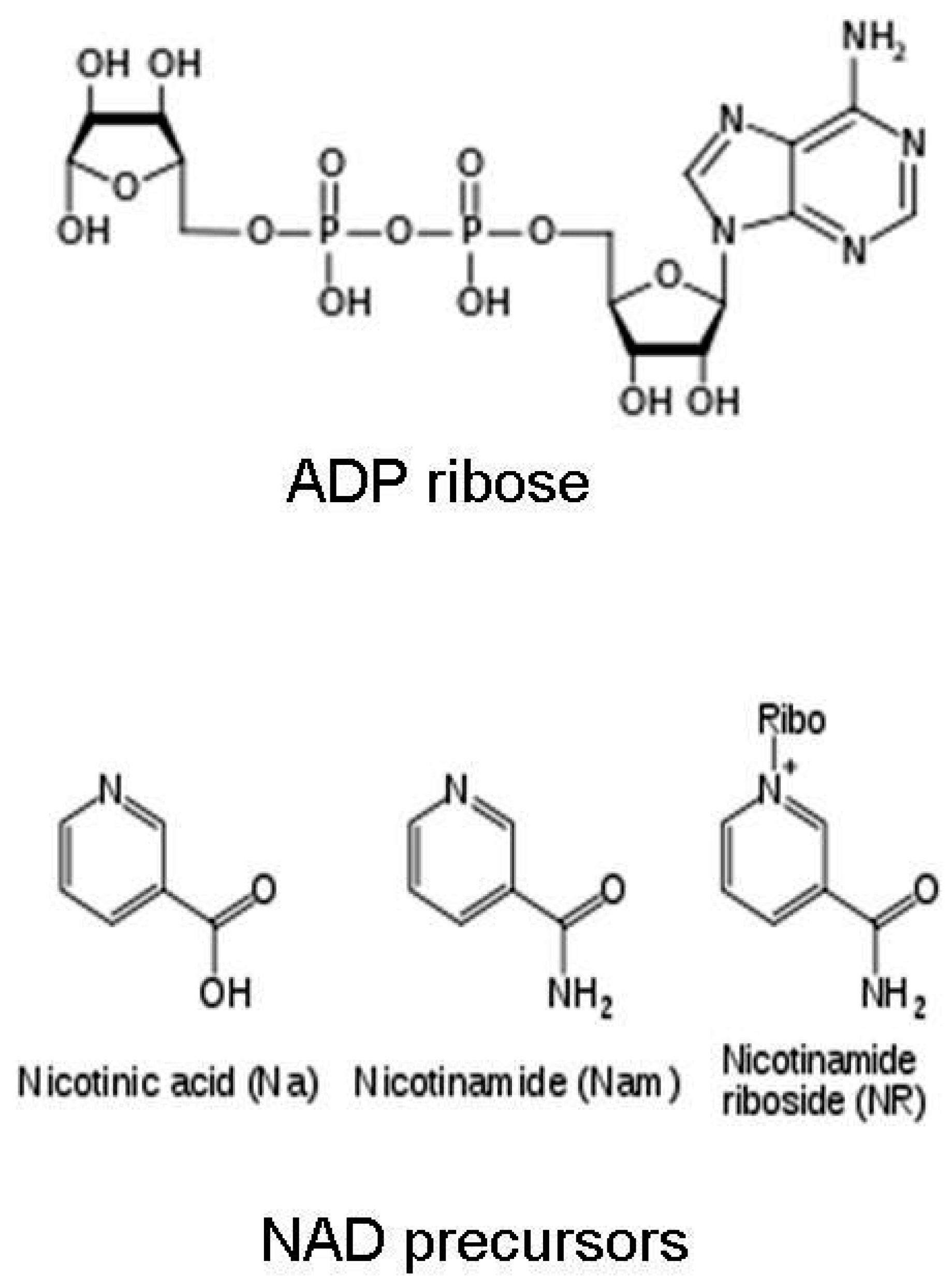
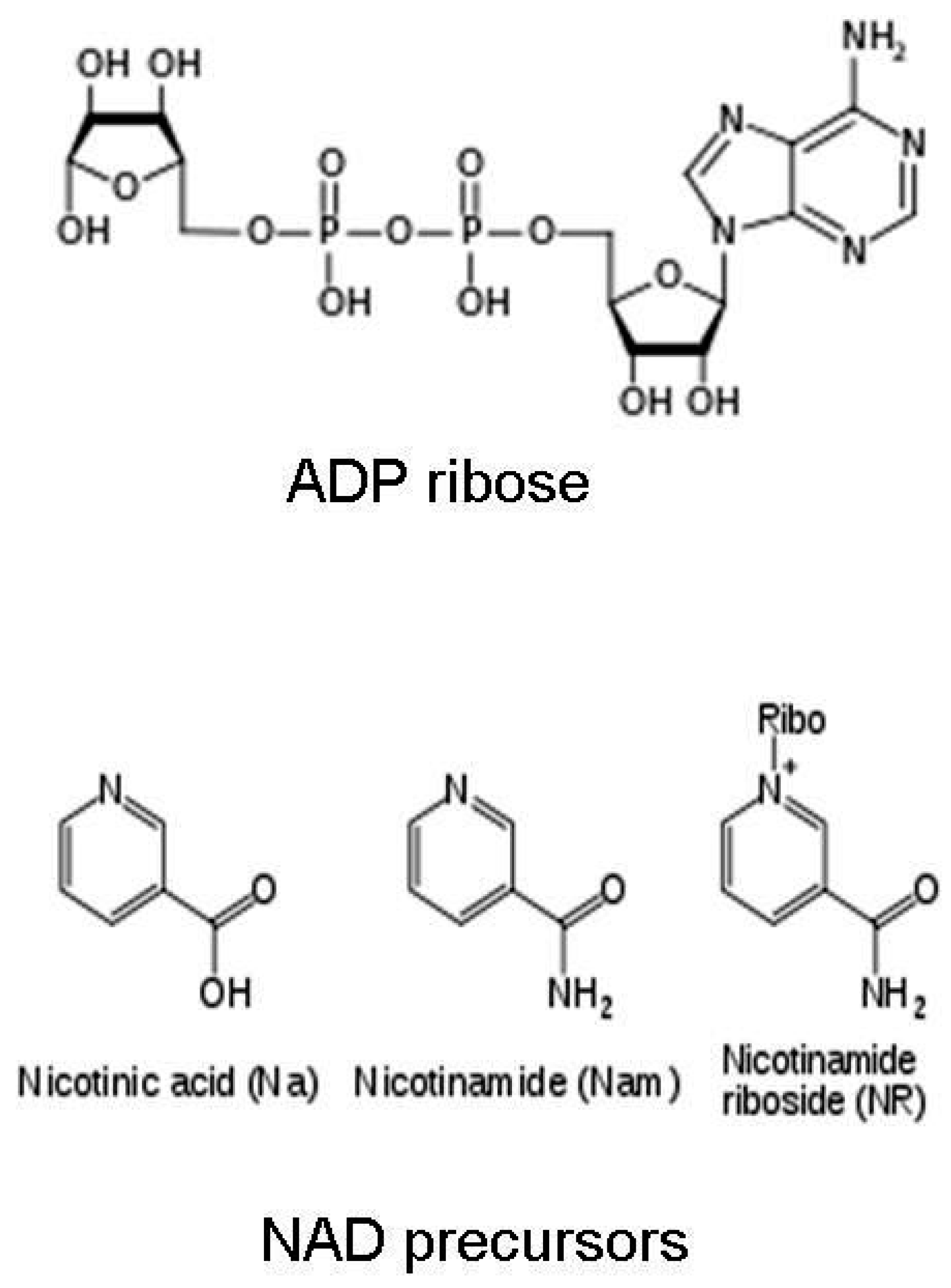
Figure 2.
Chemical structures of nicotinic acid, nicotinamide, nicotinamide riboside, and ADP ribose.
Figure 2.
Chemical structures of nicotinic acid, nicotinamide, nicotinamide riboside, and ADP ribose.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Poltronieri, P.; Čerekovic, N. Roles of Nicotinamide Adenine Dinucleotide (NAD+) in Biological Systems. Challenges 2018, 9, 3. https://doi.org/10.3390/challe9010003
AMA Style
Poltronieri P, Čerekovic N. Roles of Nicotinamide Adenine Dinucleotide (NAD+) in Biological Systems. Challenges. 2018; 9(1):3. https://doi.org/10.3390/challe9010003
Chicago/Turabian StylePoltronieri, Palmiro, and Nataša Čerekovic. 2018. "Roles of Nicotinamide Adenine Dinucleotide (NAD+) in Biological Systems" Challenges 9, no. 1: 3. https://doi.org/10.3390/challe9010003
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.