1. Introduction
Phosphorus (P) is an essential element for all forms of life, from single to multicellular organisms. It is required for normal muscle growth and egg formation, is an important component of nucleic acids, the genetic code, and phospholipids, and is also a co-factor of many enzyme systems. Phosphorus plays a vital role in maintaining osmotic and acid-base balance, energy metabolism, amino acid metabolism and protein synthesis. Phosphorus was named “life’s bottleneck” by chemist and science writer Isaac Asimov [
1], who claims: “Life can multiply until all the phosphorus has gone, and then there is an inexorable halt which nothing can prevent,” also writing, “We may be able to substitute nuclear power for coal, and plastic for wood, and yeast for meat, and friendliness for isolation—but for P there is neither substitute nor replacement.”
The main ingredients of non-ruminant diets are cereal grains and their by-products. These diets usually require supplementation with inorganic P, because of the low concentrations, and low availability of total P in cereal feed ingredients. Meat and bone meal has relatively high bioavailable P content but is banned from use in animal feed in European countries. This policy increases the demand for inorganic feed phosphates [
2]. The majority of feed phosphate is derived from phosphate rock that is a non-renewable resource and becoming increasingly scarce and expensive [
3]. Current global P reserves may be depleted in 50–100 years [
4,
5]. This poses the challenge of securing future P supply for the international and national animal feed industries. Poultry account for approximately 50% of animal feed phosphate consumption worldwide [
6]. Feed phosphates make a significant contribution to the P content of any broiler diet; generally providing as much as 60% of the bird’s P requirements [
7]. There is limited information on biologically determined P values in feed ingredients for poultry [
8,
9,
10]. Therefore, nutritionists have to formulate diets with substantial safety margins to guarantee that birds do not become P deficient. Greater dietary P concentrations increase the cost of poultry production, and P excretion into manure. Excessive amounts of P in manure contribute to the pollution of lakes, streams and wetlands leading to surface water eutrophication. Eutrophication of fresh water globally is accelerated by P accumulation [
11,
12].
All these concerns have attracted much global attention and stimulated the re-examination of the use of inorganic P in animal diets. This issue is pertinent to all non-ruminant animals that require dietary P supplementation, especially poultry (broilers, layers and breeders) and pigs. In this review, the focus is on meat chickens or broilers as much of the P used in animal nutrition is in poultry diets. In an effort to optimise P use by poultry, the P requirements of the modern broiler have been re-examined [
13,
14,
15,
16,
17,
18] and defining P bioavailability in feedstuffs has commenced [
2,
19]. Mention of bioavailability, which encompasses digestion, absorption and utilisation of P, evokes much debate and is the thrust of this review.
In many instances, the terms “availability” and “bioavailability” are interchangeable when referring to nutrients. However, to avoid confusion in this review, we use the term bioavailability to encompass the digestion, absorption and utilisation of P, as the term “available P” has a specific connotation in P nutrition as described below. We contend that bioavailability is central to the sustainable use of P in animal agriculture. A thorough understanding of P bioavailability will permit the development of strategies to enhance efficiency of P utilisation, reduce wastage of P resources and reduce environmental pollution by P.
2. Phosphorus Metabolism
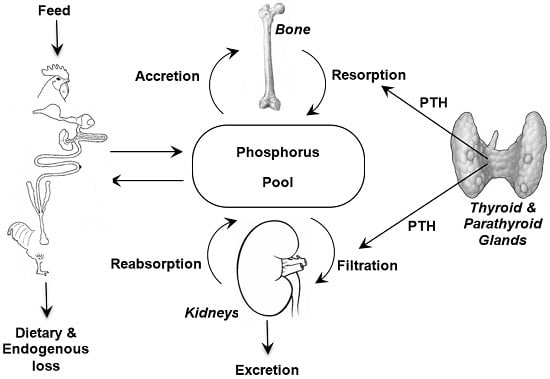
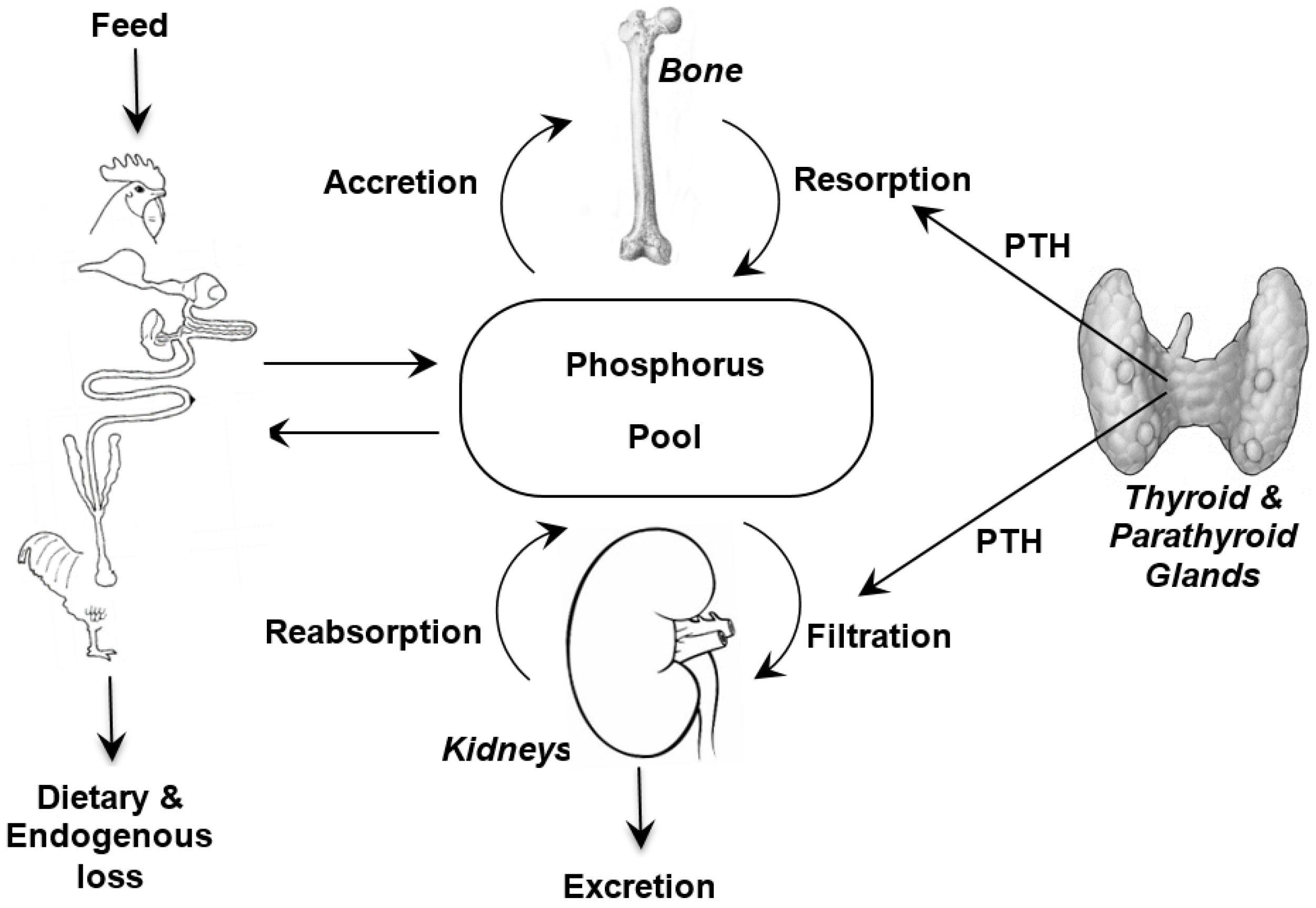
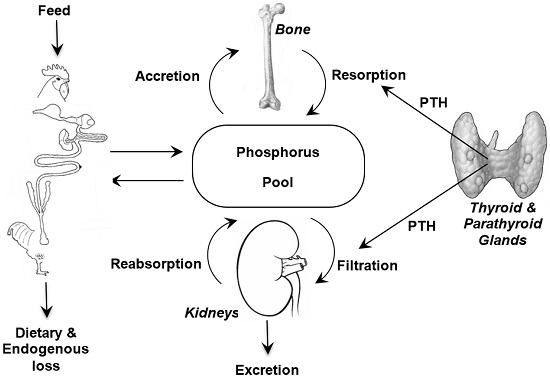
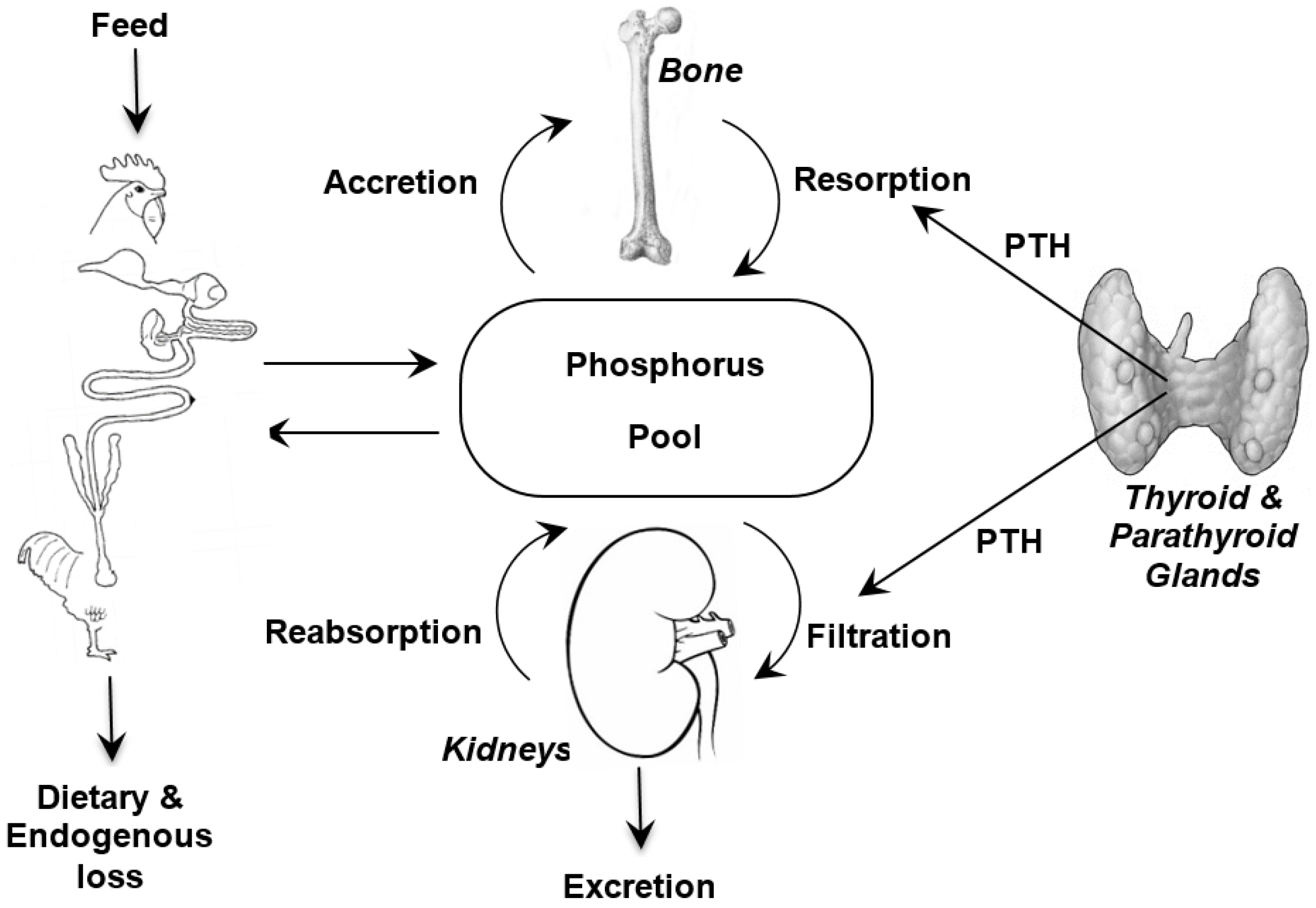
It is important when discussing phosphorus requirements and bioavailability to appreciate the metabolism of phosphorus (
Figure 1) and to remember the close association that phosphorus homeostasis has with the metabolism of two other nutrients; calcium (Ca) and vitamin D. The major features of these interactions are briefly reviewed here and the interested reader is referred to detailed reviews on the topic [
20,
21]. The major store of Ca and P is the skeleton where the cationic and anionic forms, respectively, of these minerals connect to form hydroxyapatite, which confers rigidity on the bone matrix [
2]. Bone is continually turned over and to ensure that the biological demands for both Ca and P are met, metabolism of these essential minerals is tightly coordinated.
Utilisation of Ca and P is modulated by the relative amounts of each in the diet [
2]. However, the quantities of each available for their respective metabolic functions is determined by efficiencies of intestinal absorption, glomerular filtration, renal tubular reabsorption, rates of transfer from blood to bone, and intestinal endogenous losses [
20]. The efficiencies of these processes in the different tissues are modulated by several hormones, chiefly parathyroid hormone (PTH) and the hormonal form of vitamin D
3 (1,25 dihydroxycholecalciferol; calcitriol) [
20,
21]. The control of P metabolism differs from that of Ca, with less control in the gut and greater control in the kidneys. Moreover, there is increasing evidence that dietary protein and carbohydrate affect intestinal metabolism of Ca and P [
21]. Together, the factors that determine intestinal absorption and kidney excretion along with endocrine regulation of absorption and reabsorption in the kidneys facilitate P homeostasis.
3. Phosphorus Requirements
Research on the P requirement of broilers has been the subject of numerous investigations for many decades; however, the minimum requirement for this nutrient has still not been established with any certainty. The published results are variable as different breeds of birds, age of birds, feed ingredients (with or without phytase supplementation), Ca and P sources were used in different studies. In some instances, the differences observed may reflect true differences in requirements, not merely variation resulting from experimental conditions. To further complicate interpretation of results, different criteria, including body weight gain, feed conversion ratio (FCR), tibia breaking strength, tibia ash, or toe ash contents have been used to assess responses to P (
Table 1).
Evidence in the literature clearly demonstrates that the P requirement of broilers is much lower than NRC [
29] recommendations and the values currently used by the industry [
2,
7,
13,
15,
17,
18,
22,
23,
24,
25,
27,
28,
31]. The greatest challenge in applying a lower dietary P strategy in practice is to decide what dietary concentrations of P to use in a formulation. This is due to a lack of information on biologically determined P values in feed ingredients for poultry and an agreed P evaluation system [
8,
9,
10].
5. Terms Used to Describe Bioavailable Phosphorus
Over the years, a number of terms have been suggested to define certain aspects of P utilisation as described by Rodehutscord and WPSA [
60] and outlined below. These terms have largely arisen to reflect differing approaches to the determination of bioavailable P.
Digestible P is the portion of dietary total P that is not recovered in faeces. The determination of digestible P requires in vivo studies with quantitative determination of dietary P intake and P excretion in faeces. Alternatively, digestible P can be determined by using an indigestible marker. However, as bird excreta consists of faeces and urine, determination of digestible P requires colostomised birds to exclude urinary P content.
Retainable P (RP) indicates the proportion of dietary total P that is retained in the body and can be determined quantitatively following measurement of P intake and P excretion in faeces [
60]. It is also called total tract P digestibility and is easy to measure. However, as urine is included in the determination of digestible P, this term is not an appropriate description of the measurement in poultry. Importantly, if a P intake is lower than the requirement, the P content of the urine can be ignored. In these circumstances, the determination of RP should be similar to the determination of digestible P.
Available P (AP) is the quantity of P that is absorbed from the diet by the animal. This definition of AP is different from previous interpretations [
29,
60] where it has been equated to NPP and RP. In the currently acceptable definition of AP [
60], it is determined by including an indigestible marker in the feed and is the proportion of total dietary P that is not recovered in the terminal ileum. It is also called precaecal digestible P or ileal P digestibility [
60]. The major advantage of this method is that if P intake exceeds requirements, urinary excretion of P does not confound the results.
There is now general agreement that ileal digestible P is the preferred approach for determining AP as a quantitative estimate of P bioavailability. Rodehutscord and WPSA [
60] recommend that retainable P values not be used in the future to avoid confusion. Retainable P reflects both P digestibility and post-absorptive utilisation since bird excreta includes faeces and urine. Dietary supply of P relative to requirement and body status affects total tract P utilisation [
63]. In several studies of individual feed ingredients and mixed diets, the RP in total tract is substantially different from AP in the ileum (
Table 2). Based on these and other publications that demonstrate a similar trend, both utilisation of P or Ca in feeds and requirement in birds should be expressed on an ileal digestible basis [
63]. Furthermore, AP can be used to measure the marginal level of P supplied in animal diets to meet the P requirement of the animal [
60].
7. Factors Affecting Phosphorus Availability
Determination of the bioavailability of P in feedstuffs is a major challenge. There are many factors affecting P bioavailability in birds, including: experimental techniques; the chemical form of P; dietary concentrations of Ca, vitamin D3, energy, protein, fat, and vitamin K; the availability and interaction of other nutrients in the diet, feed processing, and particle size; bird physiological and health factors, such as feed consumption, growth rate, sex and age; management factors, including ambient temperature and lighting program. Some of these factors have been investigated in detail [
14] and the major factors are briefly described below.
7.1. Phosphorus and Phytate
High levels of dietary inorganic P reduce PP availability [
49,
77] due to either the inhibitory effect of P on phytase activity [
78], a negative feedback mechanism by excessive amounts of dietary P, or inhibition by products of phytate hydrolysis [
49,
58].
Phytate is the principal storage form of P in plant feedstuffs and is considered to be an anti-nutritive factor [
79]. Phytate carries a strong negative charge and is capable of binding di- and trivalent cations such as calcium, cobalt, copper, iron, magnesium, manganese, nickel and zinc in very stable complexes [
58,
80], reducing the availability of P as well as these minerals to the animal [
2,
54,
55,
81].
Phytate has been shown to inhibit activities of some digestive enzymes such as pepsin, α-amylase and trypsin. Phytate may inhibit proteolysis by changing the protein configuration of digestive enzymes [
82]. Inhibition may also result from the chelation of Ca ions which are essential co-factors for the activity of trypsin and α-amylase leading to reduced protein and starch digestion [
50,
83,
84,
85,
86].
It has also been reported that phytate can reduce fat digestibility by forming insoluble Ca-phytate complexes with fatty acids in the gut lumen [
87]. In its chelated form, the phytate molecule is difficult to hydrolyse by phytase. The pH affects the solubility of phytate. Most phytate mineral complexes are soluble at low pHs (less than 3.5) with maximum insolubility occurring between 4 and 7 [
88]. Champagne [
89] found that Ca-phytate complexes precipitate at pH between 4 and 6, which is the approximate pH of the intestine where the Ca ions should be absorbed. Taylor [
90] has suggested that the primary factor affecting PP utilisation is the Ca ion concentration in the small intestine where insoluble Ca-phytate complexes form. A precipitated phytate mineral complex would not be accessible for hydrolysis or absorption in the intestine.
7.2. Calcium
Calcium is an essential element for bone and egg shell formation, blood clotting, muscle contraction and transmission of nerve impulses. Calcium is also an important co-factor for many enzymes and hormones.
Increases in dietary P and Ca concentrations may affect apparent digestion of these nutrients. High levels of dietary Ca and low levels of P had detrimental effects on broiler performance [
16,
17,
91,
92]. Elevated dietary Ca levels increase pH in the gut and, as a result, P absorption [
93] and retention [
91] are decreased. High dietary Ca concentration can significantly increase gastrointestinal pH [
94] and decrease not only phytate hydrolysis, but also pepsin activity in the proventriculus/gizzard and reduce apparent ileal protein digestibility [
51]. High plasma P levels decrease Ca absorption from the gut [
95]. The optimum dietary Ca-to-P ratio is of utmost importance for broiler production. With the Ca: P ratio, there will be a range of values that can meet metabolic demands for any production outcome. When dietary P concentration is high, birds can tolerate a wider range of Ca levels than when dietary P marginally meets the requirement (Li
et al. unpublished data [
96]).
Total Ca values are currently used for feed formulation since the bioavailability of Ca for feed ingredients have not been determined. Bioavailable Ca from plant raw materials is expected to be low. This is due to the high phytate content of these raw materials and the low contribution of Ca to the diet. Also, to some extent, phytate will negatively influence the availability of Ca from this dietary source by binding Ca in phytate-mineral complexes. Nevertheless, there is a need to determine the bioavailability of Ca in feed ingredients to further delineate the interaction of this nutrient with other nutrients and to improve the precision of feed formulation.
7.3. Vitamin D
Vitamin D
3 is essential for absorption of Ca and P utilization as depicted in
Figure 1. Several reports have demonstrated that dietary addition of vitamin D
3 can significantly enhance the retention of P in birds [
52]. However, excess vitamin D
3 and its metabolites have not been shown to have a further beneficial effect on bird productivity when birds are already consuming adequate vitamin D
3. Like many other minerals, understanding of the molecular mechanisms of P absorption, especially the role of the hormonal form of vitamin D
3, is still limited.
7.4. Phytase
Numerous studies have demonstrated that dietary addition of phytase increases hydrolysis of PP and improves P utilisation in pigs and poultry [
15,
16,
17,
18,
53]. Maximum utilisation of phytate by broilers fed diets with supplemental phytase was approximately 50% on average [
97] following dietary P supplementation. There are many factors affecting the efficacy of microbial phytase such as phytase source and dose [
98], physical factors of feedstuffs, feed particle size, Ca to P ratio [
99], Ca bioavailability and inorganic P supplements, vitamin D status [
100], bird age [
101] and body weight [
102], gut pH, digesta retention time, and Ca and P status. Phytase was found to be most efficacious when added to diets containing low AP and adequate Ca concentrations (Li
et al. unpublished data [
96]).
Dietary Ca level affects the efficacy of phytase in broilers [
103], but not to the same extent for all phytases [
104]. The same source of phytase can produce different responses in different flocks [
105,
106]. Moreover, the efficacy of feed phytase may be improved by the simultaneous dietary addition of other exogenous enzymes. All these variables make predicting phytase responses very difficult and lead to variation in P availability.
7.5. Birds
The ability of poultry to absorb dietary P is known to be influenced by age and physiological state. Young chicks with a rapidly developing skeletal system are critically sensitive to P deficiency [
107] and tend to use P more efficiently than do mature birds [
108]. Older birds have a greater ability to use PP, possibly due to more phytase activity in their intestinal tract [
56]. A concern often raised by commercial nutritionists is the relevance of digestibility values generated with birds of one age group to birds of another age or other species.
Applegate
et al. [
106] conducted two experiments in which chicks were fed the same diet and found that the apparent PP hydrolysis varied nearly two-fold between the first and second experiment. Similar results have been reported by Nelson and Kirby [
109] and Tamim
et al. [
103]. Bird variations make it difficult to predict treatment responses of different flocks. The variation between experiments and between flocks arises from many sources, some of which are summarised in
Table 6.
8. Conclusions
Re-examination of the phosphorus requirement of modern broilers has demonstrated that it is lower than the NRC [
29] recommendations and the values currently used by the chicken meat industry. However, there are practical limitations in applying this information to industry as there is limited data on the bioavailability and variability of P in feed ingredients; there are few reports that have measured P bioavailability of feedstuffs for poultry. Furthermore, the different methods used have made comparisons between the studies and application to practice extremely difficult. It is only when commercial poultry nutritionists have this information that they will be in a position to reduce the safety margins for P when formulating diets. To tackle these challenges, cooperative research on a global scale is needed to standardise measurement procedures in order to produce a robust and reliable database which can be used by nutritionists to formulate diets to meet the bird’s P requirement precisely; this approach is appropriate as only a couple broiler strains dominate international markets. As a starting point, the universal adoption of ileal P digestibility [
60] as the preferred procedure to determine AP would be a major step forward. Moreover, to make sense of the data derived and to allow comparison of the results, it will be necessary to report ingredient composition of experimental diets, dietary concentrations of Ca and non-phytate P, dietary electrolyte balance and mineral content, all of which influence P absorption. Achievement of these goals will assist endeavours to sustain the global supply of phosphorus.



{kind=link}
{kind=link}