Elicitors: A Tool for Improving Fruit Phenolic Content

Department of Food Science and Technology, Faculty of Veterinary Science, University of Murcia, Campus de Espinardo, 30071 Murcia, Spain
*
Author to whom correspondence should be addressed.
Agriculture 2013, 3(1), 33-52; https://doi.org/10.3390/agriculture3010033
Submission received: 30 November 2012
/
Revised: 11 January 2013
/
Accepted: 12 January 2013
/
Published: 25 January 2013
(This article belongs to the Special Issue Healthy Benefits of Nutraceutical, Phytochemicals in Agricultural Products)
Abstract
:Fruits are one of the most important sources of polyphenols for humans, whether they are consumed fresh or as processed products. To improve the phenolic content of fruits, a novel field of interest is based on results obtained using elicitors, agrochemicals which were primarily designed to improve resistance to plant pathogens. Although elicitors do not kill pathogens, they trigger plant defense mechanisms, one of which is to increase the levels of phenolic compounds. Therefore, their application not only allows us to control plant disease but also to increase the phenolic content of plant foodstuffs. Pre- or post-harvest application of the most commonly used elicitors to several fruits is discussed in this review.
1. The Importance of Plant Phenolic Compounds
Fruits, consumed fresh or as processed products, are among the most important sources of polyphenols for humans. These compounds are secondary metabolites with various chemical structures, although a benzene ring with one or more hydroxyl groups is a common feature. They are usually classified as flavonoids (anthocyanins, flavonols, flavan-3-ols, proanthocyanidins or non- hydrolyzable tannins, flavones, isoflavones, and flavanones) and non-flavonoids (hydroxycinnamic acids, hydrolyzable tannins, hydroxybenzoic acids and stilbenes) [1].
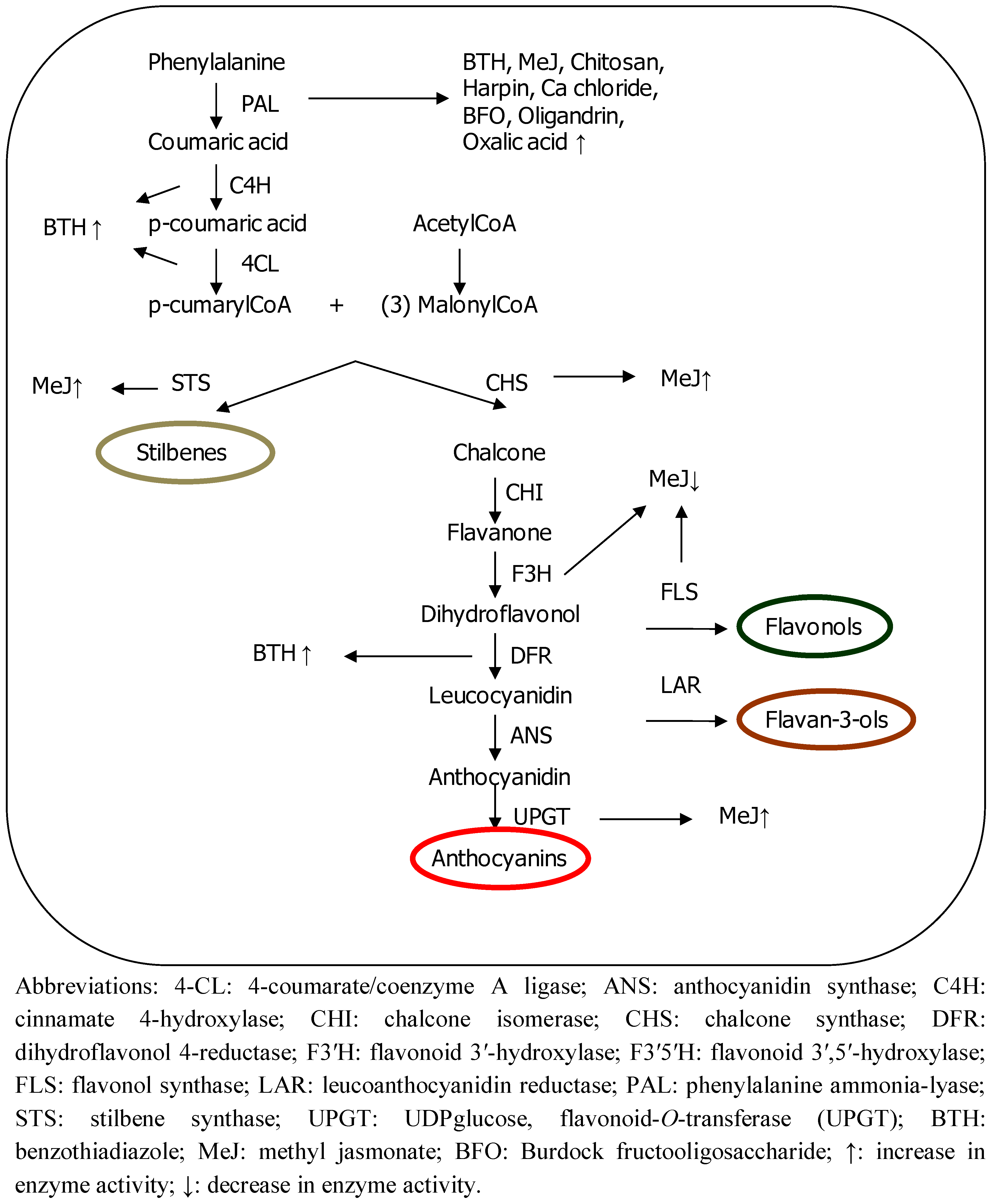
Polyphenols are synthesized from the phenylalanine produced by the shikimic acid pathway (see Figure 1). The deamination of phenylalanine catalyzed by enzyme phenylalanine ammonia-lyase (PAL) is the first step in this biosynthetic pathway. Two of the most important classes of compound, flavonoids and stilbenes, are produced through a bifurcation of this pathway. The enzymes chalcone synthase (CHS) and stilbene synthase (STS) convert the phenylpropanoid structure cumarylCoA into a polyphenol through the formation of an aromatic ring, involving the addition of three more units of C2 (consisting of two C atoms).
Figure 1.
Summary of polyphenol synthesis and enzyme regulation by elicitors.
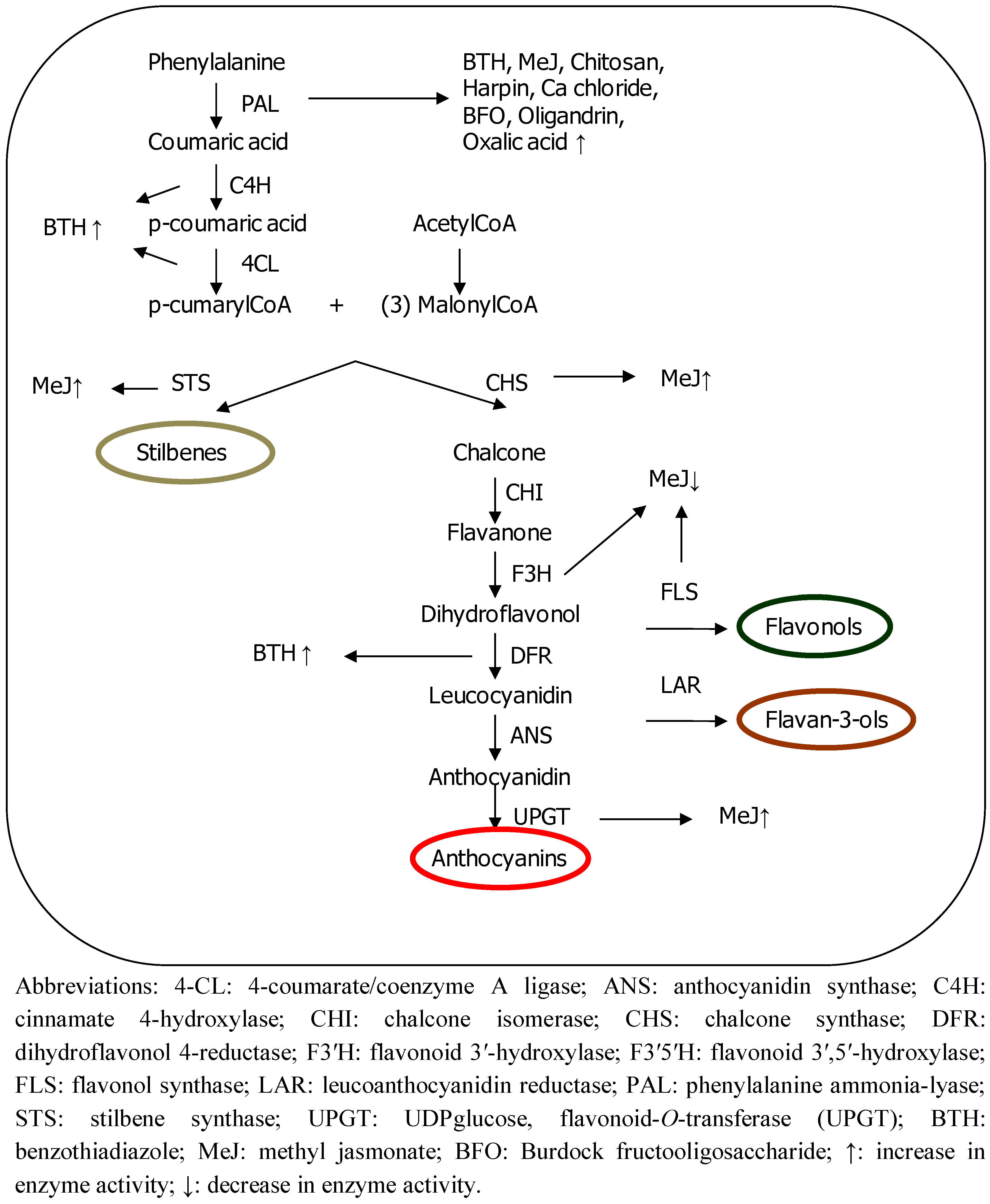
Polyphenolic compounds are important for both plants and humans for several reasons. Firstly, they protect plants from biotic and abiotic stress factors. Indeed, some of these phenolic compounds are only induced when stress factors are present, among them, the so-called phytoalexins, which are specifically involved in defense mechanisms and are synthesized after pathogen or predator attack or injury [2]. Secondly, most of these metabolites are responsible for the organoleptic and qualitative properties of foods originating from such plants. For example, anthocyanins, constitute a pigment group responsible for the color of a great variety of fruits, flowers and leaves [3], and flavan-3-ols are polyphenols involved in the bitterness and astringency of tea, grapes and wine [4,5]. Thirdly, these compounds are unique sources of industrial material in the form of food additives, pharmaceuticals and flavors [6]. Finally, they are considered to be beneficial for health, mainly due to their antioxidant activity. Many studies have suggested that a high intake of polyphenol-rich foods may have cardiovascular benefits, and provide some level of cancer chemopreventive activities and beneficial effects against other less prevalent but devastating illnesses, such as Alzheimer’s disease and urinary bladder dysfunctions [7,8,9,10]. Phenols act as antioxidants through different mechanisms [11]: (1) hydroxyl groups with π electrons of the phenyl can capture free radicals; (2) the generation of free radicals catalyzed by metals is diminished since they chelate metallic ions; (3) the cycle of generating new radicals is stopped through the donation of a proton from the phenolic compounds to the radicals and (4) polyphenols inhibit pro-oxidant enzymes that generate free radicals, such as lipoxygenases, cyclo-oxygenases and xanthine oxidase.
2. Methods for Improving Plant Phenolic Content
For all the above reasons, different methods to improve the polyphenol content of plants have been developed. The most common techniques affect cultural practices such as pruning, cluster thinning or the use of deficit irrigation. Perez-Lamela et al. [12] studied the influence of the training system and pruning of three varieties of red grape (Vitis vinifera L.) on the color intensity, total polyphenol content, total anthocyanins and tannins in wine made from them. The effect of pruning intensity on total phenolics was also investigated in mangoes. Moderate pruning significantly increased total phenolics, while the lowest contents were recorded in non-pruned trees [13]. Cluster thinning was found to increase the total anthocyanin concentration and pruning the total anthocyanin potential in Tannat and Malbec red grapes [14,15]. Soufleros et al. [16] also found that cluster thinning increased total anthocyanins and skin tannin content of red grape during ripening from 52% to 89% and from 56.2% to 114%, respectively, depending on the plot studied.
Another cultural practice that may modify the plant phenolic content is deficit irrigation. When applied pre-veraison, it caused an increase in skin anthocyanin concentration in Cabernet Sauvignon grapes [17]. Basile et al. [18] studied different irrigation regimes at three development stages of the grapevine, finding that the concentration of anthocyanins and polyphenols was improved when no water stress occurred from anthesis to fruit set, followed by mild water stress between fruit set and veraison and moderate to severe water stress post-veraison. Similar results were found by Romero et al. [19], also in grapevine. When regulated deficit irrigation strategies were applied to olive trees during fruit ripening and at different harvesting times to study the effect on oil yield and oil composition, the polyphenol content and stability of oils increased on all picking dates, especially during the first stages of the ripening period [20].
Classical breeding methods to improve certain characteristics in plants have been practiced for many years. Plant breeding can be accomplished by different techniques like simply selecting plants with desirable characteristics for vegetative propagation, the deliberate crossing of closely or distantly related individuals to produce new crop varieties or lines (called hybrids) with desirable properties, and selecting clones from the same variety that express interesting attributes. Genetic factors within crop populations may have important effects on the phenolic content, most observed changes usually being quantitative rather than qualitative [11]. In this way, the anthocyanin profile and color characteristic of a collection of 143 plants arising from crosses from Monastrell × Cabernet Sauvignon grapes was studied to select plants showing the most interesting enological characteristics. As a result, seedlings with a very interesting phenolic profile were identified, due to their composition and concentration [21]. The grape skin and seed proanthocyanidin profiles from Monastrell × Syrah grape (Vitis vinifera L.) crosses were determined by Hernandez-Jimenez et al. [22], who found that the proanthocyanidin composition of crosses was qualitatively similar to that of Monastrell and Syrah, but, quantitatively, grapes with very high procyanidin content were observed. Crosses of other types of fruit have also been performed in order to improve their polyphenolic content. For instance, in citrus, Tusa et al. [23] found ten flavonoids and four hydroxycinnamic acids in all hybrids of orange x lemon analyzed, one of the hybrids showing an intermediate polyphenol composition with respect to that of the parents. The quality of the juices from mandarin-like hybrids was studied to identify plants with high content of polyphenols [24]. Also, the content of ascorbic acid, total phenolics, flavanones, anthocyanins and phenolic acids were investigated in orange hybrids and their juices. The so called OTA 9 hybrid was found to be the richest in polyphenols [25]. Other examples include hybrids of different berries such as blackberries, raspberries, red currants, gooseberries and cornelian cherries that were assayed for antioxidant activity, ascorbic acid, phenol, and anthocyanin contents. In the above study, Pantelidis et al. [26] found that the anthocyanin and phenol contents varied widely, depending on the particular hybrid. Bugaud et al. [27] studied the total polyphenol content of bananas, finding that the total polyphenol contents of the hybrids were three-fold higher than bananas of the Cavendish variety.
Clone selection is also an important tool for improving fruit traits such as their phenolic content. In a study of different clones of Tempranillo grapes, Revilla et al. [28] observed differences in their phenolic content. The phenolic composition of clones from a given variety may be so different that can be used as discriminant factor among the clones. For example, anthocyanins, flavonols and hydorxycinnamic acids were used to discriminate between Vitis vinifera L. cv. “Barbera” clones [29]. In addition, some clones have been shown to have the capacity to produce wines with a distinct color, chromatic profile and phenolic content [30]. Burin et al. [31] studied Cabernet Sauvignon wines made from two different clones, observing differences in their chromatic characteristics. In lemon, improvements in bioactive compounds in the juice, such as vitamin C and flavonoids, may be achieved by selecting specific clones [32]. In a screening of 15 olive tree clones, some were found to have high total polyphenol, oleic and linoleic acid contents [33]. The vitamin C, total carotenoid, total anthocyanin, yellow flavonoids and polyphenol contents and total antioxidant capacity were analyzed in clones of cashew apples, and differences were found between clones [34,35]. Dietary fiber and phenolic compounds were also found to be high in selected clones of acerola and cashew apple [36]. Also in acerola, six commercial clones were investigated by Sampaio et al. [37], who found that the clone II 47/1 had the highest anthocyanin content. Oliveira et al. [38], studying five acerola clones, established that clones II 47/1, BRS 237, and BRS 236 presented outstanding results for vitamin C, phenols and antioxidant enzyme activity.
Genetic engineering has been used to modify flavonoid biosynthesis in plant tissues [39]. Using structural flavonoid genes from different plant sources, Schijlen et al. [40] were able to produce transgenic tomatoes that accumulated new phytochemicals such as high levels of stilbenes, deoxychalcones, flavones and flavonols. The resveratrol content of tomatoes has been increased by metabolic engineering in order to improve their nutritional value. Tomato plants synthesizing resveratrol were obtained via the heterologous expression of a grape (Vitis vinifera L.) cDNA encoding for the enzyme stilbene synthase. The transgenic plants accumulated trans-resveratrol and trans-piceid in their skin and had a higher antioxidant capacity and ascorbate content than the wild tomato [41]. In addition, metabolic engineering of the flavonoid biosynthesis pathways in apple was performed by overexpressing the maize leaf color regulatory gene in the mentioned fruit. Higher levels of the anthocyanin idaein (12-fold), of epicatechin (14-fold) and of catechin (41-fold) were found in the transgenic lines [42]. However, genetic transformation is tedious and is expensive, and involves many regulatory issues, not to mention the problem of public acceptance.
3. Phenolic Compounds and Induced Resistance in Plants
Although elicitors were first used to increase plant resistance to pathogens, it was found that the mechanism involved increased polyphenol levels. Consequently, elicitors can be regarded as an interesting alternative for obtaining plants with higher polyphenol content.
Disease resistance in plants is dependent on both pre-existing physical or chemical barriers (such as thick cell walls or high quantities of lignin or tannins) and inducible defense mechanisms. Upon recognition of the attacker, inducible defenses are activated at the site of infection as well as in distant uninfected tissues. Depending on the type of attack, the plant activates different signaling pathways to synthesize a specific set of defensive compounds [43].
The resistance process, mediated by the accumulation of endogenous salicylic acid (SA), a metabolite downstream the biosynthetic pathway initiated by phenylalanine ammonialyase (PAL), is called systemic acquired resistance (SAR) and is based on the induction of secondary metabolic pathways and the increased synthesis of products, phenolic compounds among them, by this metabolism as a response to pathogen attack [44]. It is assumed that SA acts by inducing: (1) pathogenesis-related proteins such as glucanases, peroxidases and chitinases that may contribute to the resistance via hydrolysis of the pathogen cell wall; (2) an oxidative burst, which triggers the elicitation of phytoalexins with antifungal properties; and (3) the phenylpropanoid pathway by activation of PAL, which leads to the formation of phytoalexins, lignins, and SA. Lignification of the plant cell walls is a mechanism to resist pathogen invasion [45,46].
Besides defenses that are dependent on SA, defense-signaling pathways that are independent of this molecule have also been described. Oxygenated fatty acids (oxylipins) can be potent regulators of defense signaling, especially those known as jasmonates (JA), which orchestrate a large set of defense responses, including the synthesis of new phenolic compounds [43].
Although there are many examples of interaction between the pathways involved in the defense response of plants, the complete process is not clearly understood. JA and SA induce the expression of pathogenesis-related genes and increase resistance to chilling injury in tomato fruit [47]. Methyl jasmonate and SA treatments confer resistance to citrus green and blue molds caused by P. digitatum and P. italicum in sweet oranges [48]. Tomato fruits treated with SA and ethephon (often used as a substitute of ethylene in triggering the ethylene signaling pathway) showed less decay and disease incidence caused by Botritis cinerea [49].
4. The Use of Elicitors
In the absence of any attack, these defense mechanisms may be induced by physical or chemical elicitation. Physical elicitors include, for example, high and low temperatures, and ultraviolet and gamma radiation. The stilbene content of Monastrell grapes irradiated with UV-C light was found to be higher than in control grapes and the final wine made from UV-C-irradiated grapes was about 2- and 1.5-fold enriched in resveratrol and piceatannol, respectively, compared with the control wine [2]. Postharvest treatments involving UV irradiation increased the level of trans-resveratrol in apples [50] and of volatile and non-volatile phenols in blueberries [51]. Cold stress in blood oranges induced transcriptomic modifications directed towards increased flavonoid biosynthesis [52]. Similarly, postharvest carbon dioxide treatments induced proanthocyanidin synthesis in grapes [53].
Chemical elicitors, such as chitosan, benzothiadiazole (BTH), harpin, and 1-methylciclopropane, among others, are agrochemicals that can mimic the action of the signaling molecules SA and JA and their derivates, or simulate the attack of a pathogen. These molecules may interact with receptors in the plant, activating defense responses and triggering, in some cases, a hypersensitive reaction. For example, the main resistance substances, such as total phenolics, flavonoids, lignin and hydroxyproline-rich glycoproteins, increased after post-harvest BTH treatment in mangos [54]. The incidence of disease caused by Penicillium expansum in BTH-treated peaches was lower than in non-treated ones. Besides the higher resistance, the treated fruits presented higher levels of phenolic compounds, lignin, and chlorogenic acid [55]. The oxidative burst in tomatoes can be elicited by hyphal wall components isolated from Phytophthora spp. as an internal emergency signal to induce the metabolic cascade involved in active defense [56]. Foliar application of harpin protein was found to reduce bacterial spots in tomatoes [57]. Chitosan oligosaccharide and Burdock fructooligosaccharide (BFO) were used as elicitors to inhibit natural postharvest diseases and reduce the incidence of disease resulting from inoculation with Botrytis cinerea in tomato fruits [58].
For this reason, elicitors were primary designed to improve plant resistance against pathogens. These compounds do not kill pathogens but trigger plant defense mechanisms, among them, the production of increased levels of phenolic compounds. The effect of the application of different elicitors to plants also proved a useful technique for improving their phenolic content. A summary of the effects of different elicitors on plant phenolic composition and phenol-related enzymes is shown in Table 1.
4.1. Benzo(1,2,3)thiadiazole-7-Carbothioic Acid S-methyl Ester (BTH)
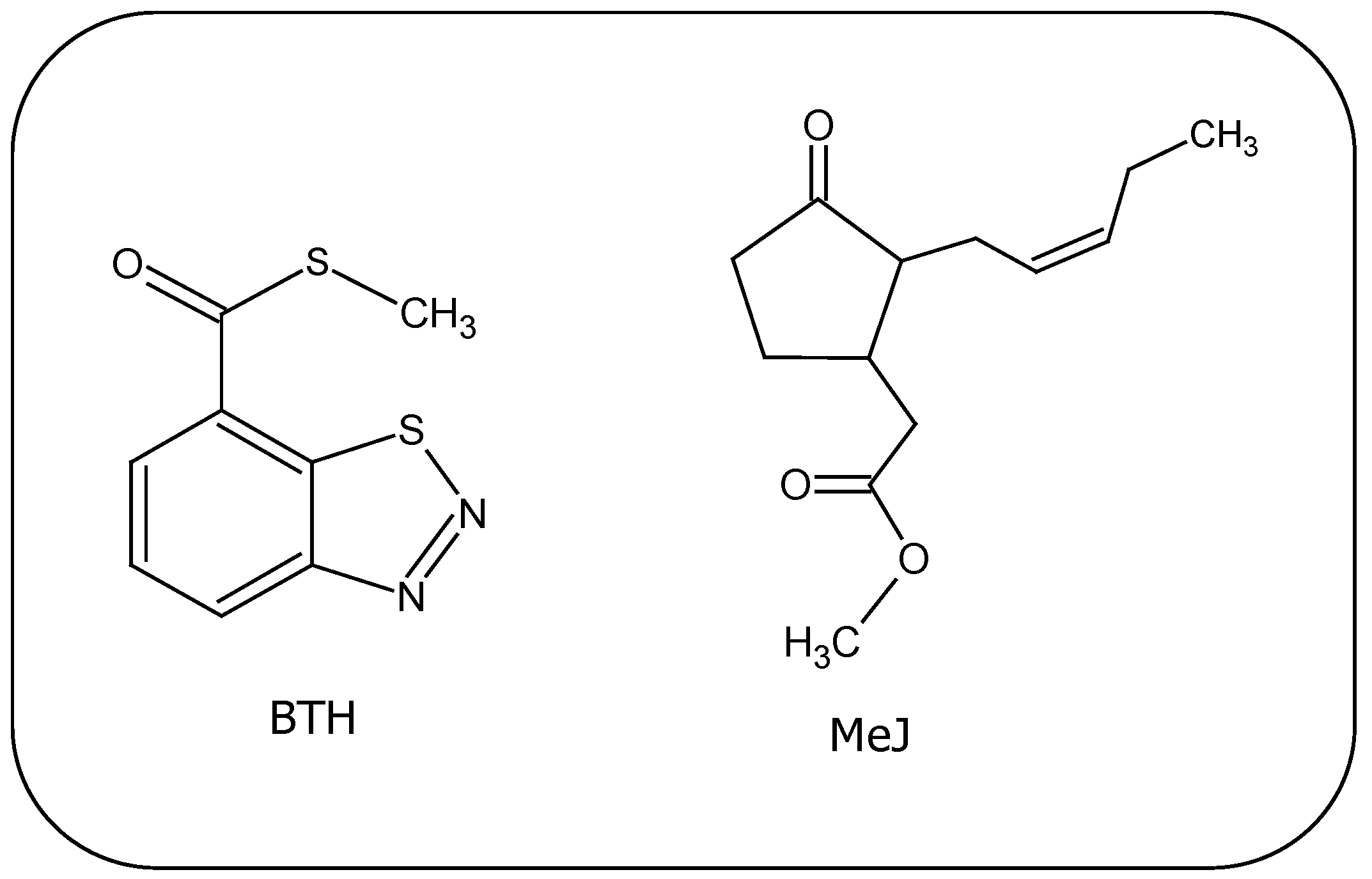
This compound is an analog of salicylic acid with a molecular weight of 136.17 and with the chemical formula shown in Figure 2. It was synthesized during a project directed at synthesizing sulfonylurea herbicides, where the formation of 2-benzylthio-3-furanylbenzoic acid methyl ester was expected rather than BTH. The ability of BTH in triggering SAR was soon discovered [59].
Figure 2.
Chemical structures of the elicitors, benzothiadiazole and methyl jasmonate.
BTH induces the activation of the enzyme PAL, as observed after postharvest treatment in mangoes [60] and peaches [55] (see Figure 1). In both studies, an increase in total phenolic compounds was also observed. Other enzymes from the plant metabolism were activated by this elicitor, including glucose-6-phosphate dehydrogenase, shikimate dehydrogenase, tyrosine ammonia lyase, PAL, cinnamate-4-hydroxylase (C4H), 4-coumarate/coenzyme A ligase (4-CL), and dihydroflavonol 4-reductase (DFR) [61]. Postharvest treatments in bananas and mangoes also resulted in the activation of polyphenol oxidase (PPO) and peroxidases (POD) and an increased total phenolic content [62,63]. However, the effect on flavonoid metabolism might be species-dependent since PAL was inhibited, whereas POD and PPO were activated by postharvest BTH treatment in loquat [64].
Besides the above mentioned studies on enzyme activities, the overall effect of BTH on polyphenolic compounds has been investigated in a variety of fruits. For instance, pre-harvest treatment with BTH of strawberries in greenhouses has proved to be useful for preventing powdery mildew and increasing the content of quercetin and kaempferol [65]; it also enhanced the accumulation of ellagic acid, ellagitannins, p-coumaric acid, gallic acid, and kaempferol hexose in leaves, and kaempferol malonylglucoside in fruits [66], while increasing the amount of quercetin and kaempferol in berries [67]. Field treatments of grapevine with BTH improve resistance to B. cinerea and enhance resveratrol and anthocyanin biosynthesis [68]. Similar studies also found an increase in total polyphenols in berry skin, particularly, the proanthocyanidin fraction [44,69], and in the anthocyanin, flavonol, and proanthocyanidin content of grapes and the color of the corresponding wines [70]. Postharvest treatment with BTH resulted in an enhancement of the phenolic and anthocyanin contents of strawberries [71].

{kind=link}
{kind=link}
Table 1.
Summary of the effects of the application of different elicitors on polyphenol content and activity of polyphenol-related enzymes.
| Elicitor | Fruit | Preharvest | Postharvest | Activated enzyme and/or increased compound | References |
|---|---|---|---|---|---|
| BTH | Mango; peach | Yes | PAL; total phenol content | [55,60] | |
| Strawberry | Yes | G6PDH, SKDH, TAL, PAL, C4H, DFR, anthocyanins; | [61,71] | ||
| Strawberry | Yes | quercetin and kaempferol; ellagic, p-coumaric, and gallic acids | [65,66,67] | ||
| Banana; mango | Yes | total phenolic content | [62,63] | ||
| Loquat | Yes | POD and PPO | [64] | ||
| Grapevine | Yes | resveratrol, anthocyanins; proanthocyanidins; flavonols | [44,68,69,70] | ||
| MeJ | Lychee | Yes | PAL | [72] | |
| Peach | Yes | PAL; total phenols | [73] | ||
| Apple, plum, table grape, strawberry | Yes | PAL; total phenols | [74] | ||
| Grapevine | Yes | CHS, STS, UPGT, stilbenes and anthocyanins | [75] | ||
| Red raspberry | Yes | myricetin, quercetin and kaempferol | [76] | ||
| Tomato | Yes | total phenols | [77] | ||
| Pomegranate, strawberry | Yes | total phenols and anthocyanins | [78,79] | ||
| Bayberry | Yes | total phenols | [80] | ||
| Berries; apple; grape | Yes | flavonoids | [70,81,82,83] | ||
| Strawberry | Yes | resveratrol | [84] | ||
| Grapevine | Yes | resveratrol and viniferins | [85,86] | ||
| Harpin | Melon | Yes | PAL, total phenol content, flavonoids, and lignins | [87] | |
| Jujube | Yes | PAL | [88] | ||
| Peach | Yes | PAL, total phenols | [89] | ||
| Chitosan | Table grape | Yes | Yes | PAL | [90,91] |
| Banana | Yes | PAL | [92] | ||
| Strawberry | Yes | total phenols | [93] | ||
| Grapevine | Yes | total phenols in grapes and wine | [94] | ||
| Oxalic acid and calcium chloride | Pear | Yes | PAL | [95] | |
| BFO | Tomato | Yes | PAL | [58] | |
| Oligandrin | Tomato | Yes | PAL | [96] | |
| Phosphite and acibenzolar- S-methyl | Apple | Yes | trans-resveratrol | [50] | |
| Potassium silicate | Avocado | Yes | catechin and epicatechin | [97] |
Abbreviations: G6PDH: glucose-6-phosphate dehydrogenase; SKDH: shikimate dehydrogenase; TAL: tyrosine ammonia lyase; C4H: cinnamate 4-hydroxylase; CHS: chalcone synthase; DFR: dihydroflavonol 4-reductase; PAL: phenylalanine ammonia-lyase; STS: stilbene synthase; UPGT: UDPglucose: flavonoid-O-transferase (UPGT); BTH: benzothiadiazole; MeJ: methyl jasmonate; BFO: Burdock fructooligosaccharide.
4.2. Methyl Jasmonate (MeJ)
MeJ is a plant volatile derived from JA with a molecular weight of 224.3, whose chemical structure is shown in Figure 2. It has similar activity to JA in plants and so is able to activate the enzymes responsible for the biosynthesis of polyphenols, such as the PAL enzyme (see Figure 1). The activation of PAL following postharvest application of the elicitor has been confirmed in many studies in fruits such as lychees [72], peaches [73], apples, plums, table grapes, strawberries [74] with a subsequent increase of total phenols. The activation of CHS, STS, UDP glucose: flavonoid-O-transferase (UPGT), proteinase inhibitors and chitinase gene expression has also been reported in pre-harvest treatments of grapevine with MeJ. Such activations triggered the accumulation of both stilbenes and anthocyanins in cells [75]. In a different fruit, red raspberry, the enhancement in the levels of myricetin, quercetin and kaempferol has also been reported after postharvest treatment with MeJ [76].
Several other studies on different fruits describe how MeJ affects polyphenol compounds. For example, postharvest treatment with MeJ resulted in higher amounts of total phenols and anthocyanins in tomatoes [77], pomegranates [78], strawberries [79], and bayberries, in which an increase of other flavonoids was also found [80]. Finally, pre-harvest treatment with MeJ has been shown to enhance the levels of flavonoids in blueberries [81], blackberries [82], apples [83], and grapes [70]; and resveratrol levels in strawberries [84] and grapevine cultivars [85,86].
4.3. Harpin
Harpin is a heat-stable, glycine-rich protein of bacterial origin. This protein was first described as being produced by the plant pathogen Erwinia amylovora that causes fire blight in pear, apple and other rosaceous plants [98]. Harpin is able to provoke a hypersensitive response in non-host plants. This response is characterized by a rapid localized cell death at the site of the invasion [99], for which reason, it is able to act as a chemical elicitor. Moreover, this protein activates ROS burst, SA and the JA/ethylene signal transduction pathways that confer SAR to different plants [100,101,102]. Harpin has been applied as an effective postharvest treatment to prevent decay in oranges [103], melons [104], apples [105] and pears [106]. In addition, field applications demonstrated its usefulness for controlling pathogen-borne diseases in passion fruits [107], pears [108], quince and loquat [109].
Harpin has also been described as an elicitor able to activate enzymes such as PAL from the polyphenol biosynthesis pathway. Examples of this have been found in several fruits; for instance, in postharvest-treated peaches and jujube, with a subsequent increase in total phenols [88,89]. In addition, in melon, field application produced enhanced levels of phenolic compounds, flavonoids and lignin [87].
4.4. Chitosan
Chitosan is a polysaccharide resulting from the deacetylation of chitin, the linear polymer of (1-4)-β-linked N-acetyl-d-glucosamine. It is obtained from the outer shell of crustaceans such as crabs and shrimps. This polysaccharide has a positive charge that confers specific physiological and biological properties that are found useful in different industries such as the cosmetics, food, biotechnology, pharmacology, medicine and agriculture industries [110]. Even though it has antimicrobial properties, there is strong evidence that it can act as an elicitor by inducing the production of callose and phenolics in susceptible plants [46].
The extent of the antimicrobial action of chitosan is influenced by factors such as its molecular weight (MW) and degree of acetylation (DA). However, it is difficult to find a clear correlation between these two characteristics and the antimicrobial activity. In general, as the DA increases, the antimicrobial activity is enhanced, since chitosan with a high DA dissolves in water completely, leading to an increased chance of interaction between chitosan and the negatively charged cell walls of micro-organisms. Similarly, as the MW increases, chitosan activity against pathogens increases, but, above a certain value, the effect is reversed [111,112].
Applications of chitosan in the field proved to be effective in controlling postharvest diseases in strawberries [113,114] and in jujubes, where it activated defense-related enzymes to reduce postharvest decay [90]. Postharvest applications of a coating composed of chitosan and Origanum vulgare L. essential oil at sub-inhibitory concentrations were able to control Rhizopus stolonifer and Aspergillus niger in grapes [91].
Many pre- and post-harvest treatments with chitosan have demonstrated that this compound can activate the enzyme PAL (see Figure 1) and increase total polyphenols in table grapes, controlling storage gray mold [92], and activating PPO [93]. It may also enhance the activity of defense-related enzymes in bananas [94] and increase the amount of total polyphenols in strawberries [95]. In addition, chitosan has proved to be effective at controlling powdery mildew and at increasing the total polyphenol content of grapes. Moreover, wines made from chitosan-treated grapes showed a higher total polyphenol content and antiradical power than those made from fungicide-treated and untreated grapes [96].
4.5. Other Elicitors
Many other chemical substances have been studied as possible elicitors in different fruits. For example, oxalic acid and calcium chloride enhanced defense-related enzyme activities, such as β-1, 3-glucanase, PAL, POD and PPO, and reduced disease incidence caused by Alternaria alternata in pears [97]; Burdock fluctooligosaccharide and oligandrin inhibited postharvest disease caused by B. cinerea in tomatoes and activated PAL, enhancing the biosynthesis of phenolic compounds [58,115]; phosphite and acibenzolar-S-methyl induced the synthesis of trans-resveratrol in apples [50], and finally, potassium silicate increased the amounts of catechin and epicatechin in avocados [116].
5. Conclusions
Polyphenolic compounds are important for plants and humans for many reasons: they protect the plant from biotic and abiotic stressors, they contribute to some organoleptic and quality properties in food, they are a unique source of industrial material for uses as food additives, pharmaceuticals, and flavors; and they are considered beneficial for health because of their antioxidant capacity. Fruits are one of the most important sources of polyphenols for humans. The use of elicitors may be regarded as a simple and useful technique to increase the phenolic content of fruit, protecting, at the same time, both plants and fruits from biotic and abiotic stresses, without the disadvantage of the environmental impact and risk of creating resistant pathogen strains that may result from using conventional herbicides and antifungal or antimicrobial compounds.
Acknowledgments
This work was made possible by financial assistance from the Fundación Séneca, Project 08659/PI/08. Yolanda Ruiz-García was supported by a pre-doctoral fellowship from the Fundación Séneca.
References
- Waterhouse, A.L. Wine phenolics. Ann. N. Y. Acad. Sci. 2002, 957, 21–36. [Google Scholar] [CrossRef]
- Cantos, E.; Espín, J.C.; Fernández, M.J.; Oliva, J.; Tomás-Barberán, A. Postharvest UV-C-Irradiated grapes as a potencial source for producing stilbene-enriched red wines. J. Agric. Food Chem. 2003, 51, 1208–1214. [Google Scholar] [CrossRef]
- Harborne, J.B.; Williams, C.A. Advances in flavonoids research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Noble, A.C. Bitterness in wine. Physiol. Behav. 1994, 56, 1251–1255. [Google Scholar] [CrossRef]
- Halsam, E.; Lilley, T.H. Natural astringency in foodstuffs—A molecular interpretation. Crit. Rev. Food Sci. Nutr. 1988, 27, 1–40. [Google Scholar]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar]
- Leifert, W.R.; Abeywardena, M.Y. Cardioprotective actions of grape polyphenols. Nutr. Res. 2008, 28, 729–737. [Google Scholar] [CrossRef]
- Croizier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1046. [Google Scholar] [CrossRef]
- De Pascual-Teresa, S.; Moreno, D.A.; Garcia-Viguera, C. Flavanols and anthocyanins in cardiovascular health: A review of current evidence. Int. J. Mol. Sci. 2010, 11, 1679–1703. [Google Scholar] [CrossRef] [Green Version]
- Pezzuto, J.M. Grapes and human health: A perspective. J. Agric. Food Chem. 2008, 56, 6777–6784. [Google Scholar] [CrossRef]
- Parr, A.J.; Bolwell, G.P. Phenols in the plant and in man: The potential for possible nutritional enhancement of the diet by modifying the phenols content or profile. J. Sci. Food Agric. 2000, 80, 985–1012. [Google Scholar] [CrossRef]
- Perez-Lamela, C.; García-Falcon, M.S.; Simal-Gándara, J.; Orriols-Fernández, I. Influence of grape variety, vine system and enological treatments on the colour stability of young red wines. Food Chem. 2007, 101, 601–606. [Google Scholar] [CrossRef]
- Singh, S.K.; Singh, S.K.; Sharma, R.R. Effects of pruning intensity on the biochemical status of shoot buds in three mango (Mangifera indica L.) cultivars planted at high density. J. Hortic. Sci. Biotechnol. 2010, 85, 483–490. [Google Scholar]
- González-Neves, G.; Gil, G.; Ferrer, M. Effect of different vineyard treatments on the phenolic contents in tannat (Vitis vinifera L.) grapes and their respective wines. Food Sci. Technol. Int. 2002, 8, 315–321. [Google Scholar]
- Fanzone, M.; Zamora, F.; Jofre, V.; Assof, M.; Peña-Neira, A. Phenolic composition of malbec grape skins and seeds from Valle de Uco (Mendoza, Argentina) during Ripening: Effect of cluster thinning. J. Agric. Food Chem. 2011, 59, 6120–6136. [Google Scholar] [CrossRef]
- Soufleros, E.H.; Stavridou, K.; Dagkli, V. The effect of cluster thinning on phenolic maturity of Vitis vinifera cv. Xinomavro grapes. Int. J. Vine Wine Sci. 2011, 45, 171–179. [Google Scholar]
- Koundouras, S.; Hatzidimitriou, E.; Karamolegkou, M.; Dimopoulou, E.; Kallithraka, S.; Tsialtas, J.T.; Zioziou, E.; Nikolaou, N.; Kotseridis, Y. Irrigation and rootstock effects on the phenolic concentration and aroma potential of Vitis vinifera L. cv. Cabernet Sauvignon grapes. J. Agric. Food Chem. 2009, 57, 7805–7813. [Google Scholar] [CrossRef]
- Basile, B.; Marsal, J.; Mata, M.; Vallverdu, X.; Bellvert, J.; Girona, J. Phenological sensitivity of Cabernet Sauvignon to water stress: Vine physiology and berry composition. Am. J. Enol. Vitic. 2011, 62, 452–461. [Google Scholar] [CrossRef]
- Romero, P.; Fernandez-Fernandez, J.I.; Martinez-Cutillas, A. Physiological thresholds for efficient regulated deficit-irrigation management in winegrapes grown under semiarid conditions. Am. J. Enol. Vitic. 2010, 61, 300–312. [Google Scholar]
- Motilva, M.J.; Tovar, M.J.; Romero, M.P.; Alegre, S.; Girona, J. Influence of regulated deficit irrigation strategies applied to olive trees (Arbequina cultivar) on oil yield and oil composition during the fruit ripening period. J. Sci. Food Agric. 2000, 80, 2037–2043. [Google Scholar] [CrossRef]
- Gómez-Plaza, E.; Gil-Muñoz, R.; Hernández-Jiménez, A.; López-Roca, J.M.; Ortega-Regules, A.; Martínez-Cutillas, A. Studies on the anthocyanin profile of Vitis Vinifera intraspecific hybrids (Monastrell × Cabernet Sauvignon). Eur. Food Res. Technol. 2008, 227, 479–484. [Google Scholar] [CrossRef]
- Hernández-Jiménez, A.; Gómez-Plaza, E.; Martínez-Cutillas, A.; Kennedy, J. Grape skin and seed proanthocyanidins from Monastrell × Syrah grapes. J. Agric. Food Chem. 2009, 57, 10798–10803. [Google Scholar] [CrossRef]
- Tusa, N.; Abbate, L.; Renda, A.; Ruberto, G. Polyphenols distribution in juices from citrus allotetraploid somatic hybrids and their sexual hybrids. J. Agric. Food Chem. 2007, 55, 9089–9094. [Google Scholar] [CrossRef]
- Rapisarda, P.; Bellon, S.E.; Fabroni, S.; Russo, G. Juice quality of two new mandarin-like hybrids (Citrus clementina Hort. ex Tan × Citrus sinensis L. Osbeck) containing antocyanins. J. Agric. Food Chem. 2008, 56, 2074–2078. [Google Scholar] [CrossRef]
- Rapisarda, P.; Fabroni, S.; Peterek, S.; Russo, G.; Mock, H.P. Juice of New citrus hybrids (Citrus clementina Hort. ex Tan × C. sinensis L. Osbeck) as a source of natural antioxidants. Food Chem. 2009, 117, 212–218. [Google Scholar] [CrossRef]
- Pantelidis, G.E.; Vasilakakis, M.; Manganaris, G.A.; Diamantidis, G. Antioxidant capacity, phenol, anthocyanin and ascorbic acid contents in raspberries, blackberries, red currants, gooseberries and cornelian cherries. Food Chem. 2007, 102, 777–783. [Google Scholar] [CrossRef]
- Bugaud, C.; Alter, P.; Daribo, M.O.; Brillouet, J.M. Comparison of the physico-chemical characteristics of a new triploid banana hybrid, FLHORBAN 920, and the Cavendish variety. J. Sci. Food Agric. 2009, 89, 407–413. [Google Scholar] [CrossRef]
- Revilla, E.; Garcia-Beneytez, E.; Cabello, F. Anthocyanin fingerprint of clones of Tempranillo grapes and wines made with them. Aust. J. Grape Wine Res. 2009, 15, 70–78. [Google Scholar] [CrossRef]
- Ferrandino, A.; Guidoni, S. Anthocyanins, flavonols and hydroxycinnamates: An attempt to use them to discriminate Vitis vinifera L. cv “Barbera” clones. Eur. Food Res. Technol. 2010, 230, 417–427. [Google Scholar]
- Gómez-Plaza, E.; Gil-Muñoz, R.; Martínez-Cutillas, A. Multivariate classification of wines from seven clones of Monastrell grapes. J. Sci. Food Agric. 2000, 80, 497–501. [Google Scholar] [CrossRef]
- Burin, V.M.; Costa, L.L.F.; Rosier, J.P.; Bordignon-Luiz, M.T. Cabernet Sauvignon wines from two different clones, characterization and evolution during bottle ageing. Food Sci. Technol. 2011, 44, 1931–1938. [Google Scholar]
- Gonzalez-Molina, E.; Moreno, D.A.; Garcia-Viguera, C. Comparison of Verna lemon juice quality for new ingredients and food products. Sci. Hort. 2009, 120, 353–359. [Google Scholar] [CrossRef]
- Romero, A.; Tous, J.; Diaz, I. Vigin oil characteristics for selected clones from Arbequina variety. Acta Hort. 2008, 791, 713–717. [Google Scholar]
- Lopes, M.M.D.; de Miranda, M.R.A.; Moura, C.F.H.; Eneas, J. Bioactive compounds and total antioxidant capacity of cashew apples (Anacardium occidentale L.) during the ripening of early dwarf cashew clones. Cienc. Agrotec. 2012, 36, 325–332. [Google Scholar] [CrossRef]
- Abreu, C.R.A.; Maia, G.A.; Figueiredo, R.W.; Sousa, P.H.M.; Alves, R.E.; Brito, E.S.; Moura, C.F.H.; Rufino, M.S.M. Bioactive compounds and antioxidant activity of cashew apple (Anacardium occidentale L.) from commercial early dwaf clones. Acta Hort. 2009, 841, 451–454. [Google Scholar]
- Rufino, M.D.M.; Perez-Jimenez, J.; Tabernero, M.; Alves, R.E.; de Brito, E.S.; Saura-Calixto, F. Acerola and cashew apple as sources of antioxidants and dietary fibre. Int. J. Food Sci. Technol. 2010, 45, 2227–2233. [Google Scholar] [CrossRef]
- Sampaio, C.G.; Morais, S.M.; Rufino, M.S.M.; Alves, R.E.; Brito, E.S. Quality, bioactive compound content, and antioxidant activity in fruits of Brazilian Acerola clone. Acta Hort. 2009, 841, 463–466. [Google Scholar]
- Oliveira, L.D.; Moura, C.F.H.; de Brito, E.S.; Mamede, R.V.S.; de Miranda, M.R.A. Antioxidant metabolism during fruit development of different Acerola (Malpighia emarginata D.C) clones. J. Agric. Food Chem. 2012, 60, 7957–7964. [Google Scholar]
- Martens, S.; Knott, J.; Seitz, C.A.; Janvari, L.; Yu, S.N.; Forkmann, G. Impact of biochemical pre-studies on specific metabolic engineering strategies of flavonoid biosynthesis in plant tissues. Biochem. Eng. J. 2003, 14, 227–235. [Google Scholar] [CrossRef]
- Schijlen, E.; de Vos, C.H.R.; Jonker, H.; van den Broeck, H.; Molthoff, J.; van Tunen, A.; Martens, S.; Bovy, A. Pathway engineering for healthy phytochemicals leading to the production of novel flavonoids in tomato fruit. Plant Biotechnol. J. 2006, 4, 433–444. [Google Scholar] [CrossRef]
- D’Introno, A.; Paradiso, A.; Scoditti, E.; D’Amico, L.; de Paolis, A.; Carluccio, M.A.; Nicoletti, I.; DeGara, L.; Santino, A.; Giovinazzo, G. Antioxidant and anti-inflammatory properties of tomato fruits synthesizing different amounts of stilbenes. Plant Biotechnol. J. 2009, 7, 422–429. [Google Scholar] [CrossRef]
- Li, H.H.; Flachowsky, H.; Fischer, T.C.; Hanke, M.V.; Forkmmann, G.; Treutter, D.; Schwab, W.; Hoffmann, T.; Szankowski, I. Maize Lc transcription factor enhances biosynthesis of anthocyanins, distinct proanthocyanidins and phenylpropanoids in apple (Malus domestica Borkh.). Planta 2007, 226, 1243–1254. [Google Scholar] [CrossRef]
- Beckers, G.J.M.; Spoel, S.H. Fine-tuning plant defence signalling: Salicylate versus jasmonate. Plant Biol. 2006, 8, 1–10. [Google Scholar] [CrossRef]
- Iriti, M.; Rossoni, M.; Borgo, M.; Ferrara, L.; Faoro, F. Induction of resistance to gray mold with benzothiadiazole modifies amino acid profile and increases proanthocyanidins in grape: Primary versus secondary metabolism. J. Agric. Food Chem. 2005, 53, 9133–9139. [Google Scholar]
- Sticher, L.; Mauch-Mani, B.; Métraux, J.P. Systemic acquired resistance. Ann. Rev. Phytopathol. 1997, 35, 235–270. [Google Scholar] [CrossRef]
- Gozzo, F. Systemic acquired resistance in crop protection: From nature to a chemical approach. J. Agric. Food Chem. 2003, 51, 4487–4503. [Google Scholar] [CrossRef]
- Ding, C.K.; Wang, C.Y.; Gross, K.C.; Smith, D.L. Jasmonate and salicylate induce the expression of pathogenesis-related-protein genes and increase resistance to chilling injury in tomato fruit. Planta 2002, 214, 895–901. [Google Scholar] [CrossRef]
- Iqbal, Z.; Singh, Z.; Khangura, R.; Ahmad, S. Management of citrus blue and green moulds through application of organic elicitors. Aust. Plant Pathol. 2012, 41, 69–77. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Li, B.Q.; Qin, G.Z.; Li, L.; Tian, S.P. Defense response of tomato fruit at different maturity stages to salicylic acid and ethephon. Sci. Hortic. 2011, 129, 183–188. [Google Scholar]
- Sautter, C.K.; Storck, L.; Rizzatti, M.R.; Mallmann, C.A.; Brackmann, A. Synthesis of trans-resveratrol and rotting control in apples with use of elicitors in post-harvest. Pesq. Agropec. Bras. 2008, 43, 1097–1103. [Google Scholar] [CrossRef]
- Eichholz, I.; Huyskens-Keil, S.; Keller, A.; Ulrich, D.; Kroh, L.W.; Rohn, S. UV-B-induced changes of volatile metabolites and phenolic compounds in blueberries (Vaccinium corymbosum L.). Food Chem. 2011, 126, 60–64. [Google Scholar] [CrossRef]
- Crifo, T.; Puglisi, I.; Petrone, G.; Recupero, G.R.; Lo Piero, A.R. Expression analysis in response to low temperature stress in blood oranges: Implication of the flavonoid biosynthetic pathway. Gene 2011, 476, 1–9. [Google Scholar] [CrossRef]
- Becatti, E.; Chkaiban, L.; Tonutti, P.; Forcato, C.; Bonghi, C.; Ranieri, A.M. Short-term postharvest carbon dioxide treatments induce selective molecular and metabolic changes in grape berries. J. Agric. Food Chem. 2010, 58, 8012–8020. [Google Scholar] [CrossRef]
- Pan, Y.G.; Liu, X.H. Effect of benzo-thiadiazole-7-carbothioic acid S-methyl ester (BTH) treatment on the resistant substance in postharvest mango fruits of different varieties. Afr. J. Biotechnol. 2011, 10, 15521–15528. [Google Scholar]
- Liu, H.X.; Jiang, W.B.; Bi, Y.; Luo, Y.B. Postharvest BTH treatment induces resistance of peach (Prunus persica L. cv. Jiubao) fruit to infection by Penicillium expansum and enhances activity of fruit defense mechanisms. Postharvest Biol. Technol. 2005, 35, 263–269. [Google Scholar] [CrossRef]
- Doke, N.; Miura, Y.; Sanchez, L.M.; Park, H.J.; Noritake, T.; Yoshioka, H.; Kawakita, K. The oxidative burst protects plants against pathogen attack: Mechanism and role as an emergency signal for plant bio-defence—A review. Gene 1996, 179, 45–51. [Google Scholar] [CrossRef]
- Obradovic, A.; Jones, J.B.; Momol, M.T.; Balogh, B.; Olson, S.M. Management of tomato bacterial spot in the field by foliar applications of bacteriophages and SAR inducers. Plant Dis. 2004, 88, 736–740. [Google Scholar] [CrossRef]
- Wang, F.D.; Feng, G.H.; Chen, K.S. Defense responses of harvested tomato fruit to burdock fructooligosaccharide, a novel potential elicitor. Postharvest Biol. Technol. 2009, 52, 110–116. [Google Scholar] [CrossRef]
- Kunz, W.; Schurter, R.; Maetzke, T. The chemistry of benzothiadiazole plant activators. Pestic. Sci. 1997, 50, 275–282. [Google Scholar] [CrossRef]
- Zhu, X.; Cao, J.; Wang, Q.; Jiang, W. Postharvest infiltration of BTH reduces of mango fruits (Mangifera indica L. cv. Tainong) by Colletotrichum gloeosporioides and enhances resistance inducing compounds. J. Phytopathol. 2008, 156, 68–74. [Google Scholar] [CrossRef]
- Cao, S.F.; Hu, Z.C.; Zheng, Y.H.; Lu, B.H. Effect of BTH on anthocyanin content and activities of related enzymes in strawberry after harvest. J. Agric. Food Chem. 2010, 58, 5801–5805. [Google Scholar]
- Zhu, S.J.; Ma, B.C. Benzothiadiazole- or methyl jasmonate-induced resistance to Colletotrichum musae in harvested banana fruit is related to elevated defense enzyme activities. J. Hortic. Sci. Biotechnol. 2007, 82, 500–506. [Google Scholar]
- Lin, J.H.; Gong, D.Q.; Zhu, S.J.; Zhang, L.J.; Zhang, L.B. Expression of PPO and POD genes and contents of polyphenolic compounds in harvested mango fruits in relation to Benzothiadiazole-induced defense against anthracnose. Sci. Hortic. 2011, 130, 85–89. [Google Scholar]
- Zhu, S.J.; Zhang, Z.W.; Xu, J.W.; Ma, L.Y.; Tang, W.L. Effect of BTH treatment on storability and activity of related enzymes of harvested loquat fruit. Acta Hortic. 2007, 750, 445–450. [Google Scholar]
- Anttonen, M.; Hukkanen, A.; Tiilikkala, K.; Karjalainen, R. Benzothiadiazole induces defense responses in berry crops. Acta Hortic. 2003, 567, 177–182. [Google Scholar]
- Hukkanen, A.T.; Kokko, H.I.; Buchala, A.J.; Mcdougall, G.J.; Stewart, D.; Karenlampi, S.O.; Karjalainen, R.O. Benzothiadiazole induces the accumulation of phenolics and improves resistance to powdery mildew in strawberries. J. Agric. Food Chem. 2007, 55, 1862–1870. [Google Scholar]
- Karjalainen, R.; Lehtinen, A.; Keinanen, M.; Julkunen-Tiitto, R.; Hietaniemi, V.; Pihlava, J.M.; Tiilikkala, K.; Jokinen, K. Benzothiadiazole and glycine betaine treatments enhance phenolic compound production in strawberry. Acta Hortic. 2002, 567, 353–356. [Google Scholar]
- Iriti, M.; Rossoni, M.; Borgo, M.; Faoro, F. Benzothiadiazole enhances resveratrol and anthocyanin biosynthesis in grapevine, meanwhile improving resistance to Botrytis cinerea. J. Agric. Food Chem. 2004, 52, 4406–4413. [Google Scholar]
- Fumagalli, F.; Rossoni, M.; Iriti, M.; di Gennaro, A.; Faoro, F.; Borroni, E.; Borgo, M.; Scienza, A.; Sala, A.; Folco, G. From field to health: A simple way to increase the nutraceutical content of grape as shown by NO-dependent vascular relaxation. J. Agric. Food Chem. 2006, 54, 5344–5349. [Google Scholar]
- Ruiz-Garcia, Y.; Romero Cascales, I.; Gil-Muñoz, R.; Fernandez-Fernandez, J.I.; Lopez-Roca, J.M.; Gómez-Plaza, E. Improving grape phenolic content and wine chromatic characteristics through the use of two different elicitors: Methyl Jasmonate versus Benzothiadiazole. J. Agric. Food Chem. 2012, 60, 1283–1290. [Google Scholar]
- Cao, S.F.; Hu, Z.C.; Zheng, Y.H.; Yang, Z.F.; Lu, B.H. Effect of BTH on antioxidant enzymes, radical-scavenging activity and decay in strawberry fruit. Food Chem. 2011, 125, 145–149. [Google Scholar] [CrossRef]
- Yang, S.Y.; Chen, Y.L.; Feng, L.Y.; Yang, E.; Su, X.G.; Jiang, Y.M. Effect of Methyl jasmonate on pericarp browning of postharvest lychees. J. Food Process. Preserv. 2011, 35, 417–422. [Google Scholar] [CrossRef]
- Jin, P.; Zheng, Y.H.; Tang, S.S.; Rui, H.J.; Wang, C.Y. Enhancing disease resistance in peach fruit with methyl jasmonate. J. Sci. Food Agric. 2009, 89, 802–808. [Google Scholar] [CrossRef]
- Heredia, J.B.; Cisneros-Zevallos, L. The effects of exogenous ethylene and methyl jasmonate on the accumulation of phenolic antioxidants in selected whole and wounded fresh produce. Food Chem. 2009, 115, 1500–1508. [Google Scholar] [CrossRef]
- Belhadj, A.; Telef, N.; Saigne, C.; Cluzet, S.; Barrieu, F.; Hamdi, S.; Merillon, J.M. Effect of methyl jasmonate in combination with carbohydrates on gene expression of PR proteins, stilbene and anthocyanin accumulation in grapevine cell cultures. Plant Physiol. Biochem. 2008, 46, 493–499. [Google Scholar] [CrossRef]
- Moreno, F.D.; Blanch, G.P.; del Castillo, M.L.R. Methyl jasmonate-induced bioformation of myricetin, quercetin and kaempferol in red raspberries. J. Agric. Food Chem. 2010, 58, 11639–11644. [Google Scholar] [CrossRef]
- Tzortzakis, N.G.; Economakis, C.D. Maintaining postharvest quality of the tomato fruit by employing methyl jasmonate and ethanol vapor treatment. J. Food Qual. 2007, 30, 567–580. [Google Scholar] [CrossRef]
- Sayyari, M.; Babalar, M.; Kalantari, S.; Martinez-Romero, D.; Guillen, F.; Serrano, M.; Valero, D. Vapour treatments with methyl salicylate or methyl jasmonate alleviated chilling injury and enhanced antioxidant potential during postharvest storage of pomegranates. Food Chem. 2011, 124, 964–970. [Google Scholar] [CrossRef]
- Moreno, F.D.; Monagas, M.; Blanch, G.P.; Bartolome, B.; del Castillo, M.L.R. Enhancement of anthocyanins and selected aroma compounds in strawberry fruits through methyl jasmonate vapor treatment. Eur. Food Res. Technol. 2010, 230, 989–999. [Google Scholar] [CrossRef]
- Wang, K.T.; Jin, P.; Shang, H.T.; Zheng, Y.H. Effect of methyl jasmonate in combination with ethanol treatment on postharvest decay and antioxidant capacity in Chinese bayberries. J. Agric. Food Chem. 2010, 58, 9597–9604. [Google Scholar] [CrossRef]
- Percival, D.; MacKenzie, J.L. Use of plant growth regulators to increase polyphenolic compounds in the wild blueberry. Can. J. Plant Sci. 2007, 87, 333–336. [Google Scholar] [CrossRef]
- Wang, S.Y.; Bowman, L.; Ding, M. Methyl jasmonate enchances antioxidant activity and flavonoid content in blackberries (Rubus sp.) and promotes antiproliferation of human cancer cells. Food Chem. 2008, 107, 1261–1269. [Google Scholar]
- Shafid, M.; Singh, Z.; Khan, A.S. Pre-harvest spray application of methyl jasmonate improves red blush and flavonoid content in Cripps Pink apple. J. Hortic. Sci. Biotechnol. 2011, 86, 422–430. [Google Scholar]
- Wang, S.Y.; Chen, C.T.; Wang, C.Y.; Chen, P. Resveratrol content in strawberry fruit is affected by preharvest conditions. J. Agric. Food Chem. 2007, 55, 8269–8274. [Google Scholar] [CrossRef]
- Esna-Ashari, M.; Pour, A.M. Effect of methyl jasmonate on resveratrol production in organs and cell suspension cultures of two Iranian grapevine (Vitis vinifera L.) cultivars. J. Hortic. Sci. Biotechnol. 2011, 86, 557–562. [Google Scholar]
- Vezzulli, S.; Civardi, S.; Ferrari, F.; Bavaresco, L. Methyl jasmonate treatment as a trigger of resveratrol synthesis in cultivated grapevine. Am. J. Enol. Vitic. 2007, 58, 530–533. [Google Scholar]
- Wang, J.; Yang, B.; Zhang, Z.; Zhang, H.; Ge, Y.H. Reduction of Latent infection and enhancement of disease resistance in muskmelon by preharvest application of harpin. J. Agric. Food Chem. 2011, 59, 12527–12533. [Google Scholar]
- Danner, M.A.; Sasso, S.A.Z.; Medeiros, J.G.S.; Marchese, J.A.; Mazaro, S.M. Induction of resistance to brown-rot on peaches by elicitors use in post-harvest. Pesq. Agropecu. Bras. 2008, 43, 793–799. [Google Scholar] [CrossRef]
- Li, M.; Yu, M.L.; Zhang, Z.Q.; Liu, Z.G.; Pan, Y. Control of black spot disease caused by Alternaria alternata on jujube (Ziziphus jujuba Mill. cv. Dongzao) using HarpinXoo protein. J. Hortic. Sci. Biotechnol. 2012, 87, 250–254. [Google Scholar]
- Yan, J.Q.; Cao, J.K.; Jiang, W.B.; Zhao, Y.M. Effects of preharvest oligochitosan sprays on portharvest fungal disease, storage quality, and defense responses in jujube (Zizyphus jujuba Mill. cv. Dongzao) fruit. Sci. Hortic. 2012, 142, 196–204. [Google Scholar] [CrossRef]
- Dos Santos, N.S.T.; Athayde Aguiar, A.J.A.; de Oliveira, C.E.V.; Verissimo de Sales, C.; de Melo E Silva, S.; Sousa da Silva, R.; Stamford, T.C.M.; de Souza, E.L. Efficacy of the application of a coating composed of chitosan and Origanum vulgare L. essential oil to control Rhizopus stolonifer and Aspergillus niger in grapes (Vitis labrusca L.). Food Microbiol. 2012, 32, 345–353. [Google Scholar] [CrossRef]
- Romanazzi, G.; Nigro, E.; Ippolito, A.; Di Venere, D. Effects of pre- and postharvest chitosan treatments to control storage grey mold of table grapes. J. Food Sci. 2002, 67, 1862–1867. [Google Scholar] [CrossRef]
- Meng, X.H.; Qin, G.Z.; Tian, S.P. Influences of preharvest spraying Cryptococcus laurentii combined with postharvest chitosan coating on postharvest diseases and quality of table grapes in storage. Food Sci. Technol. 2010, 43, 596–601. [Google Scholar]
- Meng, X.C.; Tang, Y.X.; Zhang, A.Y.; Huang, X.M.; Zhang, Z.Q. Effect of oligochitosan on development of Colletotrichum musae in vitro and in situ and its role in protection of banana fruits. Fruits 2012, 67, 147–155. [Google Scholar] [CrossRef]
- Mazaro, S.M.; Deschamps, C.; de Mio, L.L.; Biasi, L.A.; de Gouvea, A.; Sautter, C.K. Post harvest behavior of strawberry fruits after pre harvest treatment with chitosan and acibenzolar-S-methyl. Revista Brasileira de Fruticultura 2012, 30, 185–190. [Google Scholar]
- Iriti, M.; Vitalini, S.; Di Tommaso, G.; D’Amico, S.; Borgo, M.; Faoro, F. New chitosan formulation prevents grapevine powdery mildew infection and improves polyphenol content and free radical scavenging activity of grape and wine. Aust. J. Grape Wine Res. 2011, 17, 263–269. [Google Scholar] [CrossRef]
- Tian, S.P.; Qin, G.Z.; Xu, Y. Induction of defense responses against Alternaria rot by different elicitors in harvested pear fruit. Appl. Microbiol. Biotechnol. 2006, 70, 729–734. [Google Scholar] [CrossRef]
- Wei, Z.M.; Laby, R.J.; Zumoff, C.H.; Bauer, D.W.; He, S.Y.; Collmer, A.; Beer, S.V. Harpin, elicitor of the hypersensitive response produced by the plant pathogen Erwinia amylovora. Science 1992, 257, 85–88. [Google Scholar]
- He, S.Y.; Huang, H.-C.; Collmer, A. Pseudomonas syringae pv. syringae harpinPss: A protein that is secreted via the HRP pathway and elicits the hypersensitive response in plants. Cell 1993, 73, 1255–1266. [Google Scholar] [CrossRef]
- Dong, H.S.; Delaney, T.P.; Bauer, D.W.; Beer, S.V. Harpin induces disease resistance in Arabidopsis through the systemic acquired resistance pathway mediated by salicylic acid and the NIM1 gene. Plant J. 1999, 20, 207–215. [Google Scholar] [CrossRef]
- Dong, H.P.; Peng, J.L.; Bao, Z.L.; Meng, X.D.; Bonasera, J.M.; Chen, G.Y.; Beer, S.V.; Dong, H.S. Downstream divergence of the ethylene signaling pathway for harpin-stimulated Arabidopsis growth and insect defense. Plant Physiol. 2004, 136, 3628–3638. [Google Scholar] [CrossRef]
- Li, W.Q.; Shao, M.; Zhong, W.G.; Yang, J.; Okada, K.; Yamane, H.; Zhang, L.; Wang, G.; Wang, D.; Xiao, S.S.; et al. Ectopic expression of Hrf1 enhances bacterial resistance via regulation of diterpene phytoalexins, silicon and reactive oxygen species burst in rice. PloS One 2012, 7, 1–10. [Google Scholar]
- Lucon, C.M.M.; Guzzo, S.D.; de Jesus, C.O.; Pascholati, S.F. Postharvest harpin or Bacillus thuringiensis treatments suppress citrus black spot in Valencia oranges. Crop Prot. 2010, 29, 766–772. [Google Scholar] [CrossRef]
- Bi, Y.; Li, Y.C.; Ge, Y.H.; Wang, Y. Induced Resistance in melons by elicitors for the control of postharvest diseases. Postharvest Pathol. 2010, 2, 31–41. [Google Scholar]
- De Capdeville, G.; Beer, S.V.; Watkins, C.B.; Wilson, C.L.; Tedeschi, L.O.; Aist, J.R. Pre- and post-harvest harpin treatments of apples induce resistance to blue mold. Plant Dis. 2003, 87, 39–44. [Google Scholar] [CrossRef]
- Wang, J.J.; Wang, Y.; Ge, Y.H.; Bi, Y. Inhibiting effect of postharvest harpin treatment on Alternaria rot and induction to resistance enzymes of Pyrus bretschneideri cv Pingguoli. J. Gansu Agric. Univ. 2006, 41, 114–117. [Google Scholar]
- Boro, M.C.; Beriam, L.O.S.; Guzzo, S.D. Induced resistance against Xanthomonas axonopodis pv. passiflorae in passion fruit plants. Trop. Plant Pathol. 2011, 36, 74–80. [Google Scholar]
- Saour, G.; Ismail, H.; Hashem, A. Impact of kaolin particle film, spirodiclofen acaricide, harpin protein, and an organic biostimulant on pear psylla Cacopsylla pyri (Hemiptera: Psyllidae). Int. J. Pest Manag. 2010, 56, 75–79. [Google Scholar] [CrossRef]
- Bastas, K.K.; Maden, S. Evaluation of host resistance inducers and conventional products for fire blight management in loquat and quince. Phytoprotection 2007, 88, 93–101. [Google Scholar] [CrossRef]
- Bautista-Baños, S.; Hernanadez-Lauzardo, A.N.; Velazquez-del Valle, M.G.; Hernandez-Lopez, M.; Barka, E.A.; Bosquez-Molina, E.; Wilson, C.L. Chitosan as a potential natural compound to control pre and postharvest diseases of horticultural commodities. Crop Prot. 2006, 25, 108–118. [Google Scholar] [CrossRef]
- Zhang, H.; Li, R.; Liu, W. Effects of chitin and its derivative chitosan on postharvest decay of fruits: A review. Int. J. Mol. Sci. 2011, 12, 917–934. [Google Scholar] [CrossRef]
- Badawyl, M.E.; Rabea, E.I. A biopolymer chitosan and its derivatives as promising antimicrobial agents against plant pathogens and their applications in crop protection. Int. J. Carbohyd. Chem. 2011. [Google Scholar] [CrossRef]
- Reddy, M.V.B.; Belkacemi, K.; Corcuff, R.; Castaigne, F.; Arul, J. Effect of pre-harvest chitosan sprays on post-harvest infection by Botrytis cinerea and quality of strawberry fruit. Postharvest Biol. Technol. 2000, 20, 39–51. [Google Scholar] [CrossRef]
- Romanazzi, G.; Nigro, F.; Ippolito, A. Effectiveness of pre and postharvest chitosan treatments on storage decay of strawberries. Revista di Frutticoltura e di Ortofloricoltura 2000, 62, 71–75. [Google Scholar]
- Wang, A.Y.; Lou, B.G.; Xu, T.; Lin, C. Defense responses in tomato fruit induced by oligandrin against Botrytis cinerea. Afr. J. Biotechnol. 2011, 10, 4596–4601. [Google Scholar]
- Tesfay, S.Z.; Bertling, I.; Bower, J.P. Effects of postharvest potassium silicate application on phenolics and other anti-oxidant systems aligned to avocado fruit quality. Postharvest Biol. Technol. 2011, 60, 92–99. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Ruiz-García, Y.; Gómez-Plaza, E. Elicitors: A Tool for Improving Fruit Phenolic Content. Agriculture 2013, 3, 33-52. https://doi.org/10.3390/agriculture3010033
AMA Style
Ruiz-García Y, Gómez-Plaza E. Elicitors: A Tool for Improving Fruit Phenolic Content. Agriculture. 2013; 3(1):33-52. https://doi.org/10.3390/agriculture3010033
Chicago/Turabian StyleRuiz-García, Yolanda, and Encarna Gómez-Plaza. 2013. "Elicitors: A Tool for Improving Fruit Phenolic Content" Agriculture 3, no. 1: 33-52. https://doi.org/10.3390/agriculture3010033