Photosynthesis and Yellow Vine Syndrome of American Cranberry

Department of Physical Sciences, Alabama State University, 915 South Jackson Street, Montgomery, AL 36104, USA
Agriculture 2012, 2(2), 125-138; https://doi.org/10.3390/agriculture2020125
Submission received: 23 April 2012
/
Revised: 1 June 2012
/
Accepted: 2 June 2012
/
Published: 7 June 2012
(This article belongs to the Special Issue The Impact of Plant Disease on Food Security)
Abstract
:The American cranberry (Vaccinium macrocarpon Ait.) contains rich antioxidants and has significant health benefits in fighting a variety of human diseases. In the past ten years, cranberry growers have reported yellow vine syndrome, which is associated with reduced photosynthetic performance, in the cranberry bogs. It has been found that the yellow vine syndrome of cranberry is associated with nutritional imbalance; it might be an issue for cranberry quality and food security as well as the crop production. This review evaluates the present state of knowledge of yellow vine syndrome, together with recent advances that are resulting from an improved mechanistic understanding and a possible solution that will be of considerable value to cranberry growers. This review also includes results from the author’s own laboratory. Water stress, nutritional imbalance, and photoinhibition are the likely reasons for producing yellow vine of cranberry. Future endeavors should be placed on the combination of genetic, biochemical, and biophysical techniques at the molecular level and plant physiology at the field and greenhouse level. This may provide specific information in order to understand the molecular details of yellow vine of cranberry as well as a tool for guiding future breeding efforts and management practices.
1. Introduction
The American cranberry (Vaccinium macrocarpon Ait.) is a long-lived woody evergreen trailing vine and grows in temperate climate zones in United States and Canada. Cranberry is a nutritious food and beverage source. In Massachusetts, cranberry is an important horticultural crop that has been cultivated on approximately 14,000 acres [1]. Cranberry fruit contains abundant antioxidants, such as flavonoid phenolic acids, both in quality and quantity and has noteworthy health benefits in fighting a variety of human diseases including cardiovascular problem and cancer [2,3,4,5,6]. Cranberry juice has long been consumed for the prevention of urinary tract infections. The in vitro studies using a variety of tumor models showed that polyphenolic extracts from cranberries inhibited the growth and proliferation of breast, colon, prostate, lung, and other tumors.
Environmental stress plays an important role in the growth and production of agricultural crops. Response mechanisms of plants to stress factors have been proposed in the past decade, such as nutritional deficiency [7,8,9], drought stress [10], heat-induced inactivation [11,12], UV and visible effects [13,14], tolerance to salinity [15,16], water stress [17,18,19], and chilling sensitivity [20]. The production and quality of cranberries may be affected by long-term or short-term environmental stress, such as water, temperature, humidity, nutrient, and light intensity.
Under normal light conditions, cranberry growers observed yellow vine syndrome, which produced yellow coloration along the leaf margins while the area along the vein remained green (Figure 1). Each year numerous reports of yellow vine syndrome of cranberry were received from cranberry growers in Massachusetts, USA [21]. Typically the symptoms show up in the old leaves and then move up the stem into the newly developed leaves. The most common season for the symptoms to become severe is around or just after fruit set when demand for resources in the plants is high. However, the molecular mechanism for developing yellow vine syndrome in cranberries has been poorly studied and is largely unknown. Complete understanding of the mechanisms of yellow vine syndrome development in cranberry plants may offer an opportunity to minimize its effect. In this review, the recent advances in probing the mechanism for the formation of yellow vine syndrome, possible solutions and future efforts have been summarized and discussed.
Figure 1.
Images of normal cranberry leaves (a) and yellow vine syndrome cranberry leaves (b).

2. Photosynthesis
Photosynthesis, which harvests, transfers, converts and stores the solar energy in the form of chemical bonding energy and supports all life on earth, is one of the most important chemical reactions in science. In the photosynthetic process, the most amazing chemistry is embodied in the water splitting reaction, which occurs in the photosystem II (PS II) protein complex embedded in the thylakoid membranes of higher plants, green algae, and cyanobacteria [22,23,24]. However the molecular basis of photosynthetic water oxidation has remained one of the major mysteries in bioenergetics research.
Light is the energy source for photosynthesis; however, too much of it may cause damage to plants and algae by producing highly reactive intermediates such as P680+ and toxic reactive oxygen species such as singlet oxygen [25]. In particular, environmental stress may enhance the extent of photoinhibition [26]. To comply with a rapidly fluctuating light environment, the light harvesting system in plants and algae uses non-photochemical quenching (NPQ) to dissipate the excess light energy into harmless heat radiation to protect the photosynthetic apparatus from photodamage [27]. The reversible phosphorylation of thylakoid proteins modulates the membrane dynamics and affinity in response to high light stress [28].
Photosynthesis serves as the principal method of energy storage in cranberry plants and affects cranberry yield. The effect of radiation on net photosynthetic rate was examined to develop a model for understanding the impact of radiation on photosynthesis of the cranberry plant [29]. Saturating light level for both cranberry cultivars was in the range of 600 to 800 μmol m2 s−1, depending on temperature. In the field, the predicted values from the nonrectangular hyperbola model fitted only the early morning data. By midday, the observed net photosynthetic rate was consistently lower than the predicted values, but rose again by late afternoon to levels closer to the predicted data. The data indicated that cranberry leaves experience midday depression [29].
Light may affect carbon production in cranberries. There are several indications that cranberry vines are constantly under carbohydrate stress, such as low fruit set which ranges from 30% to 40%, biennial bearing of uprights (vertical, fruit-bearing stems), and competition between developing fruits for resources. Cranberry stores little carbohydrate in below-ground tissue and has no major permanent wood for the storage of carbohydrates. The effect of light on carbon production and partitioning in cranberry showed that the light compensation point differed between upright and runner tissue. The light compensation point for runners was much higher than those for fruiting uprights and vegetative uprights [30]. However, use of greenhouse vines simplified the relationships between multiple sinks that were present in the field and further experiments in cranberry bog may improve our understanding of the effects.
The effect of light treatment on the post-harvest ripening of late-harvested cranberries resulted in 28% more phenolics and 24% higher antioxidant capacity [31]. The concentration of glucose, the predominant sugar, was not significantly affected by light. However, light increased fructose and sucrose concentration by 202% and 167%, respectively. Similarly, light increased the concentration of quinic, malic, and shikimic acids by 62%, 100%, and 140%, respectively. In addition, antioxidant content was dependant on the use of light wavelength, suggesting the expressions of enzymes that catalyze anthocyanin biosynthesis are regulated by different environments [32].
3. Yellow Vine Syndrome of Cranberry
3.1. Effect of Shade Treatment
Yellow vine syndrome on cranberry leaves appears as yellowing along leaf margins. The chlorophyll content is expected to be lower than in normal cranberry leaves. The shading of the yellow vine plants seems to passively affect chlorophyll concentrations and may not have any effect on nutrient concentrations [33]. The chlorophyll content of the yellow vine samples in the absence and presence of shade was determined by spectrometric analysis (Table 1). Shading increased Chl a and Chl b content by 11.1% and 13.9%, respectively [34]. HPLC analysis of shaded and unshaded yellow vine leaves using a C18 column with a photodiode array detector, following published procedures, was conducted and confirmed the spectrometric data.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Analytical results of Chl a, Chl b, and the Chl a/Chl b ratio in the methanol extracts of yellow vine samples in the absence and presence of shade [34].
| Unshaded yellow vine leaves | Shaded yellow vine leaves | Change (%) | |
|---|---|---|---|
| Chl a (mg/g) | 0.99 ± 0.05 | 1.10 ± 0.06 | +11.1 |
| Chl b (mg/g) | 0.72 ± 0.04 | 0.82 ± 0.04 | +13.9 |
| Chl a/ Chl b | 1.37 ± 0.06 | 1.34 ± 0.06 | −0.02 |
The above observation can be explained by the increase in the rate of chlorophyll biosynthesis in yellow vine cranberry leaves. For example, shading activates the photoreceptor and turns on the genes involved in the chlorophyll biosynthetic pathway [35,36,37]. Alternatively, the partial recovery of chlorophyll could be due to slow photodegradation and photodamage of chlorophylls although this would be expected to be more extensive under bright light. It has been demonstrated that chlorophyll is photosensitive to damage from excess light and degrades rapidly [38,39,40,41,42]. The photodamage and photodegradation of chlorophyll is associated with the highly active singlet oxygen species formed by triplet chlorophylls [39,43,44].
As shown in Table 1, the Chl a/Chl b ratio in the unshaded yellow vine samples was 1.36, while the Chl a/Chl b ratio was 1.34 in the shaded leaves. The photosynthetic reaction centers of PS I and PS II in higher plants contained the Chl a without Chl b [22]. In contrast, the light harvesting system contained both Chl a and Chl b. The comparable Chl a/Chl b ratio in both samples suggested that the organization of photosynthetic centers in cranberry leaves was unchanged. It suggests that the organization of photosynthetic machinery in yellow vine cranberry plants might not be affected by shading. An increase in Chl a content would imply an increase in the number of reaction centers in both photosystem II and light-harvesting complex.
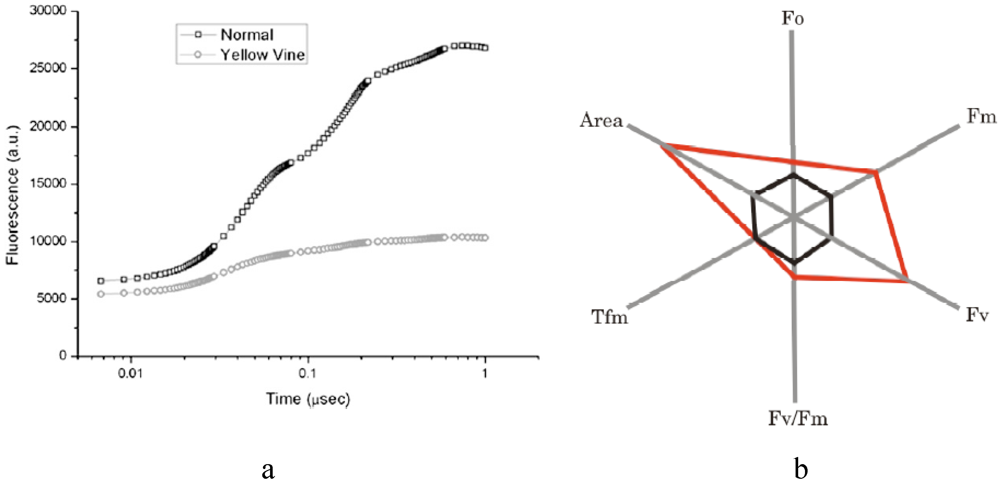
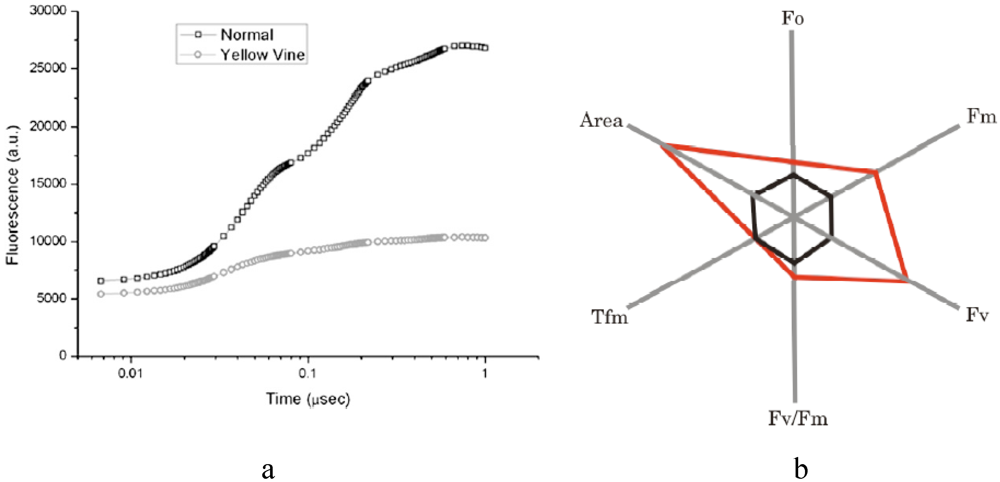
Fluorescence by chlorophyll is very sensitive to each step of the PS II electron transfer reactions in vivo. Therefore chlorophyll fluorescence has been proven to be an informative tool and reveals information on plant performance and responses via non-invasive measurements, especially addressing the effects of plant leaves under environmental stress conditions [45,46,47,48,49]. Chlorophyll fluorescence analysis provides the structural and functional parameters quantifying PS II behavior, including initial fluorescence (Fo), variable fluorescence (Fv), maximum fluorescence (Fm), lifetime of maximum fluorescence (Tfm), numbers of reaction centers per absorption cross section (RC/ABS), and PS II maximum quantum yield (Fv/Fm) [47]. The size of the PS II quinone pool is expressed as “Area,” which is the measurement of fluorescence area above the transient [47]. The photosynthesis performance index, PI, is defined as a driving force of the primary photosynthetic reaction [50]. The spider plots of chlorophyll fluorescence data provide an overall picture of photosynthetic efficiency in plants in vivo [47]. A spider plot of chlorophyll fluorescence for cranberry samples revealed that five of the six parameters of shaded yellow vine leaves were increased dramatically over those of unshaded samples (Figure 2). This observation agreed with the notion that PS II organization was unaffected by the shade. The Fv/Fm ratio was increased by about ~20%. The rise in Fv/Fm indicated the enhancement of PS II electron transfer activity, which supports the hypothesis that the number of PS II was increased based on chlorophyll analysis. Similar behavior was observed in Camellia leaves with development of an energy pipeline model of the photosynthetic apparatus [51,52]. In addition, the “Area” above the fluorescence transient for the shaded sample was also increased drastically, indicating the presence of a larger quinone pool [34,53].
Figure 2.
Typical chlorophyll transient curves (a) and a spider plot (b) of the chlorophyll fluorescence parameters for yellow vine cranberry leaves in the absence (black) and presence (red) of shade. (Adapted from [34], with permission from the American Society for Horticultural Science).
Figure 2.
Typical chlorophyll transient curves (a) and a spider plot (b) of the chlorophyll fluorescence parameters for yellow vine cranberry leaves in the absence (black) and presence (red) of shade. (Adapted from [34], with permission from the American Society for Horticultural Science).
To clarify the discrepancy between the chlorophyll fluorescence assay and chlorophyll determination by the spectrometric and HPLC method, a three-week chlorophyll fluorescence experiment was conducted. The three chlorophyll fluorescence parameters including Fv/Fm, “Area”, and photosynthesis index factor were averaged over the three weeks and listed in Table 2. The data showed; (1) the chlorophyll fluorescence parameters of unshaded yellow vine samples varied appreciably with the time (see the measured error in Table 2). Yellow vine leaves might be more sensitive to environmental stress. This could be due to the variation of several factors, such as temperature, weather, water, and use of herbicides in the bogs over the three weeks. The other possible reason could be variation due to sampling; (2) shaded yellow vine samples showed a relatively steady and constant value over time. The standard deviations of shaded yellow vine syndrome leaves were decreased by a factor of 2~10 (Table 2). This indicated the partial recovery of yellow vine syndrome under shade and enhancement of the capability to respond to the stress. As the PS II is the main target of photoinhibition, the decrease in PS II activity suggested a possible role of photoinhibition associated with the yellow vine syndrome in cranberry plants [34]. However, the data cannot rule out the involvement of other outcomes of shading.
Table 2.
Photosynthetic parameters of the shaded and unshaded cranberry leaves determined by chlorophyll fluorescence kinetics [34].
| Sample Conditions | Fv/Fm | “Area” (×105 unit) | PI |
|---|---|---|---|
| Unshaded yellow vine leaves | 0.72 ± 0.10 | 1.9 ± 0.9 | 0.32 ± 0.20 |
| Shaded yellow vine leaves | 0.85 ± 0.01 | 2.1 ± 0.4 | 0.35 ± 0.06 |
3.2. Photosynthetic Activity
Chlorophyll content in the plant was easily determined by extraction using organic solvents followed by spectrometric and HPLC methods [54,55,56,57]. The relatively lower chlorophyll content in these leaves compared to the normal healthy counterpart would be anticipated, which was supported by the spectrometric analytical data of chlorophyll analysis in the extracts of leaves by methanol and acetone shown in Table 3. The contents of both chlorophyll a and chlorophyll b were decreased from that of the normal leaves by approximately 22~24%. The low content of chlorophyll a and chlorophyll b was confirmed by HPLC analysis (data not shown). The Chl a/Chl b ratios in both yellow vine syndrome and normal leaf samples were almost identical. Chlorophyll a is primarily in the reaction centers, and chlorophyll b is predominantly found in light harvesting complexes in photosynthetic membrane proteins. Therefore, the low content of chlorophyll in yellow vine syndrome leaves was due to the smaller number of photosynthetic reaction centers associated with their intact light harvesting systems.
Table 3.
Analytical results of Chl a, Chl b, and Chl a/Chl b ratio in yellow vine and normal cranberry leaves.
| Normal leaves (mg/g fresh weight) | Yellow vine syndrome leaves (mg/g fresh weight) | Change (%) | |
|---|---|---|---|
| Chl a | 1.30 ± 0.09 | 0.99 ± 0.10 | 23.8 |
| Chl b | 0.93 ± 0.06 | 0.72 ± 0.09 | 22.6 |
| Chl a/Chl b ratio | 1.40 | 1.40 | 0 |
The PSII maximum quantum yield (Fv/Fm), the size of quinone pool (AREA), the numbers of reaction center per absorption cross section (RC/ABS), and the photosynthesis performance index (PI) were determined by chlorophyll fluorescence transient analysis. As shown in Table 4, the PS II maximum quantum yield was decreased by 28%, which is similar to the level of chlorophyll a and chlorophyll b (~25%). It suggested that the number of PS II in yellow vine leaves was lower than that of normal leaves by 25~30%. The size of the quinone pool and the number of reaction centers per chlorophyll in yellow vine samples were decreased by 57~60%. The discrepancy between these numbers and Fv/Fm might occur because the distribution of PS II in yellow vine leaves was not optimized. In particular, the much smaller size of the quinone pool would imply the vulnerability and sensitivity of yellow vine leaves to other environmental stress factors. This was supported by the observation that water stress significantly worsen the symptom in cranberry bogs [21,58]. The PI value of yellow vine leaves was almost completely diminished with a loss of ~90%. This suggested that the impairment and destruction of the photosynthetic apparatus in yellow vine syndrome of cranberry leaves is multi-targeted and complex, in which PS II is only one of the important factors.
Table 4.
Photosynthetic parameters of yellow vine syndrome and normal cranberry leaves; PSII maximum quantum yield (Fv/Fm), the size of quinone pool (Area), the numbers of reaction center per absorption cross section (RC/ABS), and the photosynthesis performance index (PI).
| FV/FM | Area | RC/ABS | PI | |
|---|---|---|---|---|
| Normal leave | 100 ± 5 | 100 ± 15 | 100 ± 8 | 100 ± 25 |
| Yellow vine | 72 ± 4 | 40 ± 6 | 43 ± 5 | 11 ± 3 |
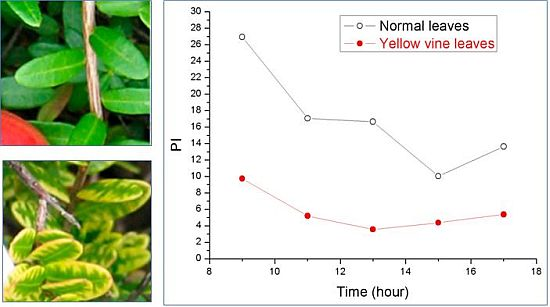
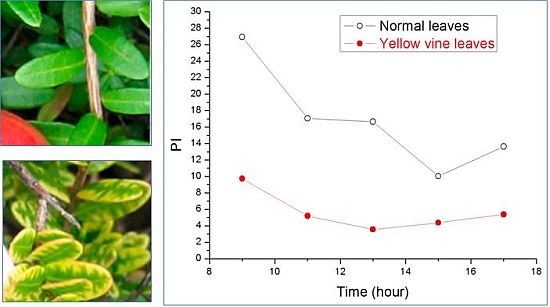
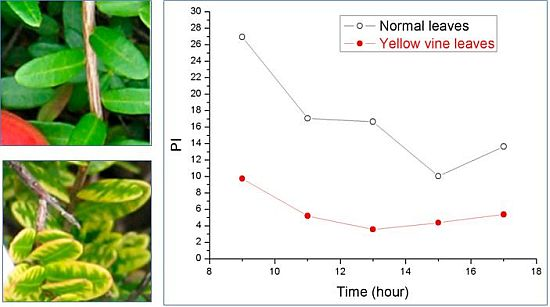
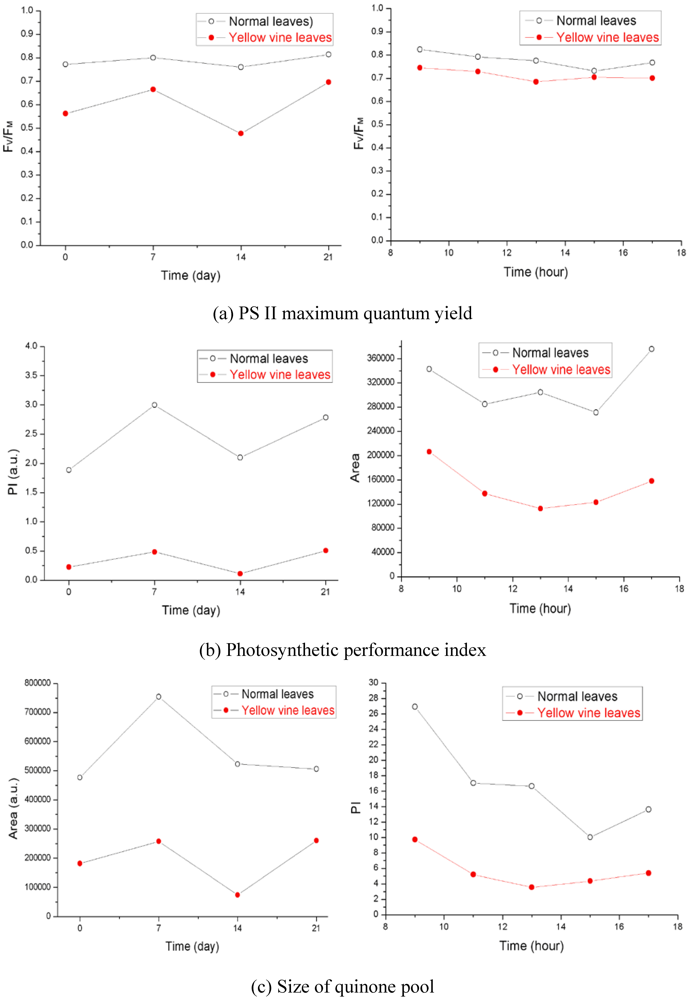
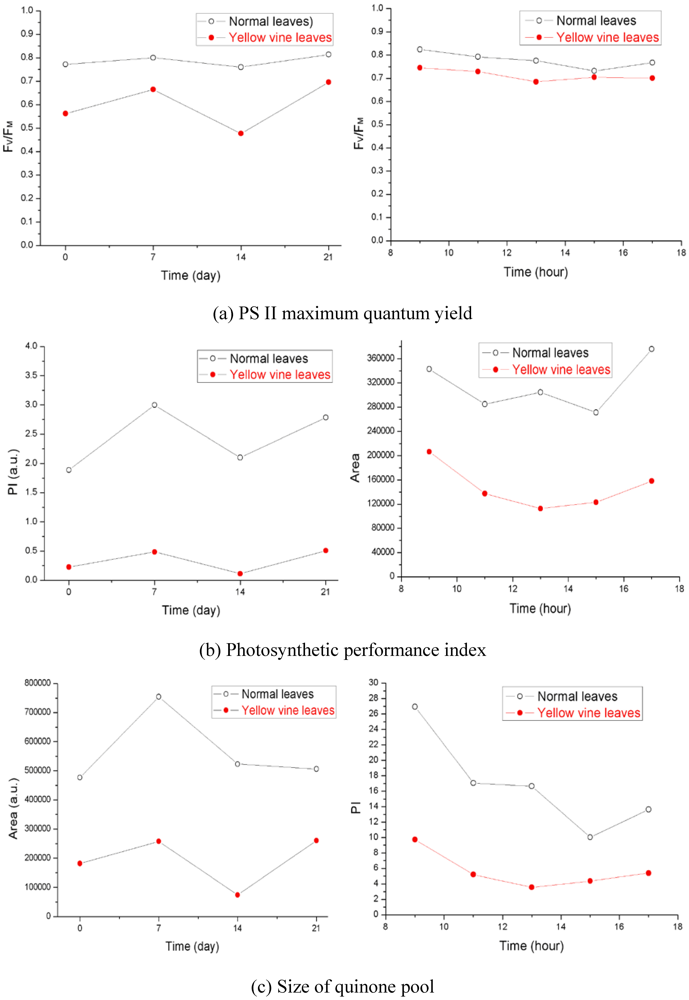
Using chlorophyll fluorescence analysis, the photosynthetic activity of yellow vine syndrome leaves was examined over a period of three weeks or of one day. As shown in Figure 3, the PS II maximum quantum yield of yellow vine samples was consistently smaller than in the normal leaves. The result was in line with the data of yellow vine leaves, which was 26~28% less in chlorophyll than the normal leaves revealed by spectrometric and HPLC analysis. The results indicated that yellow vine syndrome was associated with poor photosynthetic activity and was a threat for the long-term growth and crop production of cranberries.
Figure 3.
Photosynthetic activity of cranberry leaves obtained during a period of three weeks (left) and of one day (right): (a) PS II maximum quantum yield; (b) Photosynthetic performance index; (c) Size of quinone pool. (From [59], with permission from the American Society for Horticultural Science).
Figure 3.
Photosynthetic activity of cranberry leaves obtained during a period of three weeks (left) and of one day (right): (a) PS II maximum quantum yield; (b) Photosynthetic performance index; (c) Size of quinone pool. (From [59], with permission from the American Society for Horticultural Science).
3.3. Working Model and Hypothesis
Previous nutritional analysis showed that nutritional imbalance might be associated with yellow vine development in cranberry [60]. However fertilizer management was not the cause of the problem. It was possible that nutrient imbalance was secondary to root problems. Additionally, yellow vine syndrome often worsens in bogs with drainage problems, indicating that water stress is another factor in the formation of yellow vine in cranberries [60]. Water stress conditions might lead to poor root development.
The experiment of shade treatment revealed that the shading of cranberry plants appeared to reduce the syndrome by improving the photosynthetic activity and increasing the chlorophyll content [34]. The yellow vine leaves were associated with 11 ± 5% and 14 ± 5% increase in Chl a/Chl b ratio after shading, respectively. The electron transport efficiency in PSII and the size of the quinone pool were increased. In addition, the overall photosynthesis performance index was drastically improved by shading. These results suggested that the shade effect increases the numbers of PS II in the cells of yellow vine cranberry leaves. As PS II was involved directly in photoinhibition, a possible role of photoinhibition might be associated with the yellow vine syndrome in cranberry plants.
Yellow vine formation might be due to multiple factors. As environmental stress enhances the photoinhibitory effect on photosynthetic machineries [26], the combination of photoinhibition, water stress, and nutritional imbalance appears to cause yellow vine syndrome of cranberries. A possible model has been proposed to explain yellow vine syndrome of cranberry. When normal healthy cranberry plants encountered issues of nutritional imbalance, water stress, and photodamage, they developed yellow vine syndrome by forming less PS II complexes. Chlorophyll biosynthesis might be inhibited and negatively regulated. Alternatively, degradation of chlorophyll might be activated and positively regulated. This is likely to be associated with D1 protein turnover and the dissembling of PS II complexes.
Therefore, the performance of yellow vine and normal cranberry leaves by chlorophyll fluorescence analysis over periods of one day and of three weeks indicated that the photosynthetic parameters of the yellow vine samples were substantially lower than those of the normal cranberry leaves. Spectrometric and HPLC analyses revealed that yellow vine leaves contained 26~28% less chlorophyll than the normal cranberry leaves. These data sets demonstrate that yellow vine syndrome is associated with a poor photosynthetic activity and is problematic for the long-term growth and crop production of cranberries.
4. Conclusions
As the yellow vine syndrome of cranberry is associated with nutritional imbalance, it could be a concern for cranberry quality and food security. The yellow vine syndrome is most likely multifaceted (1) nutritional imbalance; (2) fertilizer management; (3) water stress; (4) herbicide usage; (5) light inhibition. Spectrometric, HPLC, and chlorophyll fluorescence assays confirmed the loss of chlorophyll content and photosynthetic activity in cranberry leaves exhibiting yellow vine syndrome [59]. Recent HPLC analysis revealed a link between the yellow vine and xanthophyll cycle. In addition, chlorophyll fluorescence data on the effect of diverse environmental factors such as pH and nutrition ions on yellow vine syndrome showed that the yellow vine syndrome of cranberry is associated with poor performance to stress factors. Further study using a variety of fertilizers available on the market and herbicides with various chemical structures on yellow vine syndrome would be particularly worthwhile, in order to provide chemical understanding of yellow vine formation.
Although great progress in plant stress physiology had been made, the understanding of mechanisms and relationships between cranberry plants and stress factors is relatively limited and poorly understood. We do not have a solution to solve the problem yet, and the current recommendation to treat yellow vine of cranberry is simply water and nutrition management. Recently recovery of photoinhibited plant leaves has been examined, and PS II mobility in thylakoid membranes may play a key role [61]. The recovery experiments might provide further insights into the mechanisms of yellow vine syndrome development in cranberry bogs and offer an opportunity to solve the problem. It would be interesting to conduct experiments on recovery from the syndrome under optimized experimental conditions.
As documented in the review, the effects of yellow vine syndrome in terms of changes in photosynthetic activities is rather thorough; However, a complete understanding of its etiology is needed, which may lead to a solution of this important problem. It is worth noting that bacteria or viruses might be a factor in the development of yellow vine syndrome. The appearance and pattern of yellow vine syndrome in cranberry bogs implies that the involvement of bacteria or viruses is less likely. Since the causes remain rather mysterious, the detection and identification of bacteria or viruses in yellow vine of cranberry are of interest and remain to be investigated.
Most experiments and measurements were based on observations in the fields and greenhouses. The biochemical, molecular genetics, and state-of-the art spectroscopic techniques have been rarely utilized in the research of cranberry plant physiology. It is here proposed that the combination of genetic, biochemical, and biophysical techniques at the molecular level and plant physiology at the field and greenhouse level will provide insight to understand the molecular details in cranberry stress physiology. For example, the 454-cDNA sequencing of cranberry cDNA samples under stress conditions may be used to identify the possible yellow vine genes of cranberry. Similarly, the use of LC/MS and MS/MS (including MALDI) to analyze the organic solvent extracts or purified protein fractions of cranberry leaves under stress conditions will enable the key small molecules and peptides or proteins associated with stress factors, respectively, to be identified. By determining the underlying genetic and physiological factors associated with stress factors, these research activities have potential to provide models and to elucidate the molecular mechanisms in cranberry stress physiology as well as being a tool for guiding future breeding efforts and management practices.
Acknowledgments
The work was supported by Alabama State University, UMass Dartmouth, and a grant from the US Department of Agriculture. I am grateful to Carolyn De Moranville and Peter Jeranyama for their support. I thank Douglas Strout for critical reading of the manuscript and insightful discussion and the three anonymous reviewers for their constructive comments. I also thank my students Fan Zhang, Zi Wei, Lien-Yang Chou, Wanshu He, Xuejing Hou, Ndi Geh, Sean Cederlund, Joy Patel, Robert Mulkern, and Aaron Raposo for participation in the project.
References
- Cranberry production: A guide for Massachusetts, 2008. UMass Cranberry Station Web site. Available online: http://scholarworks.umass.edu/cgi/viewcontent.cgi?article=1000&context=cranberry_prod_guide (accessed on 18 January 2011).
- Kalgaonkar, S.; Gross, H.B.; Yokoyama, W.; Keen, C.L. Effects of a flavonol-rich diet on select cardiovascular parameters in a golden syrian hamster model. J. Med. Food 2010, 13, 108–115. [Google Scholar] [CrossRef]
- Neto, C.C.; Amoroso Jon, W.; Liberty Anne, M. Anticancer activities of cranberry phytochemicals: An update. Mol. Nutr. Food Res. 2008, 52, S18–S27. [Google Scholar]
- Neto, C.C. Cranberry and its phytochemicals: A review of in vitro anticancer studies. J. Nutr. 2007, 137, 186–193. [Google Scholar]
- Lipson, S.M.; Cohen, P.; Zhou, J.; Burdowski, A.; Stotzky, G. Cranberry cocktail juice, cranberry concentrates, and proanthocyanidins reduce reovirus infectivity titers in African green monkey kidney epithelial cell cultures. Mol. Nutr. Food Res. 2007, 51, 752–758. [Google Scholar] [CrossRef]
- Deyhim, F.; Patil, B.S.; Villarreal, A.; Lopez, E.; Garcia, K.; Rios, R.; Garcia, C.; Gonzales, C.; Mandadi, K. Cranberry juice increases antioxidant status without affecting cholesterol homeostasis in orchidectomized rats. J. Med. Food 2007, 10, 49–53. [Google Scholar] [CrossRef]
- Imsande, J. Iron, sulfur, and chlorophyll deficiencies: A need for an integrative approach in plant physiology. Physiol. Plant 1998, 103, 139–144. [Google Scholar]
- Abadia, J. Leaf responses to iron deficiency: A review. J. Plant Nutr. 1992, 15, 1699–1713. [Google Scholar] [CrossRef]
- Davies, J.P.; Grossman, A.R. Responses to deficiencies in macronutrients. In Molecular Biology of Chloroplasts and Mitochondria in Chlamydomonas; Rochaix, J.-D., Goldschmidt-Clermont, M., Merchant, S., Eds.; Springer: New York, NY, USA, 1998; pp. 613–635. [Google Scholar]
- Cornic, G.; Massacci, A. Leaf photosynthesis under drought stress. In Photosynthesis and the Environment; Baker, N.R., Ed.; Springer: New York, NY, USA, 1996; pp. 347–366. [Google Scholar]
- Pshybytko, N.L.; Kruk, J.; Kabashnikova, L.F.; Strzalka, K. Function of plastoquinone in heat stress reactions of plants. Biochim. Biophys. Acta 2008, 1777, 1393–1399. [Google Scholar] [CrossRef]
- Gombos, Z.; Murata, N. Genetic engineering of the unsaturation of membrane glycerolipid: effects on the ability of the photosynthetic machinery to tolerate temperature stress. In Lipids in Photosynthesis: Structure, Function and Genetics; Siegenthaler, P.-A., Murata, N., Eds.; Springer: New York, NY, USA, 1998; pp. 249–262. [Google Scholar]
- Kasahara, M.; Wada, M. Chloroplast avoidance movement. Ann. Rev. Plant Biol. 2005, 13, 267–282. [Google Scholar]
- Tevini, M. Plant responses to ultraviolet radiation stress. In Chlorophyll a Fluorescence; Papageorgiou, G.C., Govindjee, Eds.; Springer: New York, NY, USA, 2004; pp. 605–621. [Google Scholar]
- Demetriou, G.; Neonaki, C.; Navakoudis, E.; Kotzabasis, K. Salt stress impact on the molecular structure and function of the photosynthetic apparatus—The protective role of polyamines. Biochim. Biophys. Acta 1767, 272–280. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Ann. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Mommer, L.; Visser, E.J.W. Underwater photosynthesis in flooded terrestrial plants: A matter of leaf plasticity. Anna. Bot. 2005, 96, 581–589. [Google Scholar] [CrossRef]
- Sack, L.; Holbrook, N.M. Leaf hydraulics. Ann. Rev. Plant Biol. 2006, 57, 361–381. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Voesenek, L.A.C.J. Flooding stress: Acclimations and genetic diversity. Ann. Rev. Plant Biol. 2008, 59, 313–339. [Google Scholar] [CrossRef]
- Nishida, I.; Murata, N. Chilling sensitivity in plants and cyanobacteria: The crucial contribution of membrane lipids. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 541–568. [Google Scholar] [CrossRef]
- DeMoranville, C.; Lampinen, B. Cranberry Station Newsletter; UMass Cranberry Station: East Wareham, MA, USA, 1999; Volume 8, pp. 2–3. [Google Scholar]
- Blankenship, R.E. Molecular Mechanisms of Photosynthesis; Blackwell Science: Bristol, UK, 2002. [Google Scholar]
- Brudvig, G.W. Water oxidation chemistry of photosystem II. Philos. Trans. R. Soc. B 2008, 363, 1211–1219. [Google Scholar] [CrossRef]
- Rutherford, A.W.; Boussac, A. Biochemistry: Water photolysis in biology. Science 2004, 303, 1782–1784. [Google Scholar] [CrossRef]
- Kramer, D.M. The photonic smart grid of the chloroplast in action. Proc. Natl. Acad. Sci. USA 2010, 107, 2729–2730. [Google Scholar] [CrossRef]
- Takahashi, S.; Murata, N. How do environmental stresses accelerate photoinhibition? Trends Plant Sci. 2008, 13, 178–182. [Google Scholar] [CrossRef]
- Bailey, S.; Grossman, A. Photoprotection in cyanobacteria: regulation of light harvesting. Photochem. Photobiol. 2008, 84, 1410–1420. [Google Scholar] [CrossRef]
- Vener, A.V. Environmentally modulated phosphorylation and dynamics of proteins in photosynthetic membranes. Biochim. Biophys. Acta 2007, 1767, 449–457. [Google Scholar]
- Kumudini, S. Effect of radiation and temperature on cranberry photosynthesis and characterization of diurnal change in photosynthesis. J. Am. Soc. Hort. Sci. 2004, 129, 106–111. [Google Scholar]
- Vanden Heuvel, J.E.; Davenport, J.R. Effect of light, temperature, defoliation, and fruiting on carbon assimilation and partitioning in potted cranberry. HortScience 2005, 40, 1699–1704. [Google Scholar]
- Forney, C.F.; Kalt, W.; Abrams, S.R.; Owen, S.J. Effects of postharvest light and ABA treatments on the composition of late-harvested white cranberry fruit. Acta Hortic. 2009, 810, 799–806. [Google Scholar]
- Zhou, Y.; Singh, B.R. Effect of light on anthocyanin levels in submerged, harvested cranberry fruit. J. Biomed. Biotechnol. 2004, 5, 259–263. [Google Scholar] [CrossRef]
- Sicuranza, J.; Jeranyama, P.; Hou, H.J.M.; DeMoranville, C. Shade effects on chlorophyll content and nutrient content of cranberry vines exhibiting yellow vine symptoms. HortScience 2009, 44, 553. [Google Scholar]
- Wei, Z.; Jeranyama, P.; Zhang, F.; DeMoranville, C.; Hou, H.J.M. Probing the mechanisms of the yellow vine syndrome development in cranberry: Shade effect. HortScience 2010, 45, 1345–1348. [Google Scholar]
- Tanaka, R.; Tanaka, A. Tetrapyrrole biosynthesis in higher plants. Annu. Rev. Plant Biol. 2007, 58, 321–346. [Google Scholar] [CrossRef]
- Ito, H.; Yokono, M.; Tanaka, R.; Tanaka, A. Identification of a novel vinyl reductase gene essential for the biosynthesis of monovinyl chlorophyll in Synechocystis sp. PCC6803. J. Biol. Chem. 2008, 283, 9002–9011. [Google Scholar] [CrossRef]
- Masuda, T. Recent overview of the Mg branch of the tetrapyrrole biosynthesis leading to chlorophylls. Photosynth. Res. 2008, 96, 121–143. [Google Scholar] [CrossRef]
- Nedbal, L.; Samson, G.; Whitmarsh, J. Redox state of a one-electron component controls the rate of photoinhibition of photosystem II. Proc. Natl. Acad. Sci. USA 1992, 89, 7929–7933. [Google Scholar]
- Tracewell, C.A.; Cua, A.; Stewart, D.H.; Bocian, D.F.; Brudvig, G.W. Characterization of carotenoid and chlorophyll photooxidation in photosystem II. Biochemistry 2001, 40, 193–203. [Google Scholar]
- Telfer, A.; Barber, J. Evidence for the photoinduced oxidation of the primary electron donor P680 in the isolated photosystem II reaction center. FEBS Lett. 1989, 246, 223–228. [Google Scholar] [CrossRef]
- Hou, J.; Kuang, T.; Peng, D.; Tang, C.; Tang, P. The photodamage and protective role of pheophytin a in the photosystem II reaction center against light-induced damage. Prog. Nat. Sci. 1996, 6, 489–493. [Google Scholar]
- Melis, A. Photosystem-II damage and repair cycle in chloroplasts: What modulates the rate of photodamage in vivo? Trends Plant Sci. 1999, 4, 130–135. [Google Scholar] [CrossRef]
- Hou, J.-M.; Kuang, T.-Y.; Peng, D.-C.; Tang, C.-Q.; Tang, P.-S. Photoinduced damage of the photosystem II primary electron donor P680. In Photosynthesis: Mechanisms and Effects; Garab, G., Ed.; Kluwers Academic Publishers: Norwell, MA, USA, 1998; Volume 3, pp. 2119–2122. [Google Scholar]
- Telfer, A.; Bishop, S.M.; Phillips, D.; Barber, J. Isolated photosynthetic reaction center of photosystem II as a sensitizer for the formation of singlet oxygen. Detection and quantum yield determination using a chemical trapping technique. J. Biol. Chem. 1994, 269, 13244–13253. [Google Scholar]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev.Plant Physiol. Plant Mol. Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Bazzaz, F.A. From leaves to ecosystems: Using chlorophyll fluorescence to assess photosynthesis and plant function in ecological studies. In Chlorophyll a Fluorescence; Papageorgiou, G.C., Govindjee, Eds.; Springer: New York, NY, USA, 2004; pp. 737–755. [Google Scholar]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence; Papageorgiou, G.C., Govindjee, Eds.; Springer: New York, NY, USA, 2004; pp. 321–362. [Google Scholar]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef]
- Adams, W.W., III; Demmig-Adams, B. Chlorophyll fluorescence as a tool to monitor plant response to the environment. In Chlorophyll a Fluorescence; Papageorgiou, G.C., Govindjee, Eds.; Springer: New York, NY, USA, 2004; pp. 583–604. [Google Scholar]
- Srivastava, A.; Strasser, R.J.; Govindjee. Greening of peas: Parallel measurements of 77 K emission spectra, OJIP chlorophyll a fluorescence transient, period four oscillation of the initial fluorescence level, delayed light emission, and P700. Photosynthetica 1999, 37, 365–392. [Google Scholar] [CrossRef]
- Kruger, G.H.J.; Tsimilli-Michael, M.; Strasser, R.J. Light stress provokes plastic and elastic modifications in structure and function of photosystem II in camellia leaves. Physiol. Plant. 1997, 101, 265–277. [Google Scholar] [CrossRef]
- Strasser Reto, J. Energy pipeline model. Prog. Bot. Res. 1987, 2, 717–720. [Google Scholar]
- Toth, S.Z.; Schansker, G.; Strasser, R.J. A noninvasive assay of the plastoquinone pool redox state based on the OJIP transient. Photosynth. Res. 2007, 93, 193–203. [Google Scholar] [CrossRef]
- De las Rivas, J.; Abadia, A.; Abadia, J. A new reversed-phase HPLC method resolving all major higher plant photosynthetic pigments. Plant Physiol. 1989, 91, 190–192. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Borrmann, D.; Castelhano de Andrade, J.; Lanfer-Marquez, U.M. Chlorophyll degradation and formation of colorless chlorophyll derivatives during soybean (glycine max L. Merill) seed maturation. J. Agric. Food Chem. 2009, 57, 2030–2034. [Google Scholar]
- Shioi, Y.; Fukae, R.; Sasa, T. Chlorophyll analysis by high-performance liquid chromatography. Biochim. Biophys. Acta 1983, 722, 72–79. [Google Scholar] [CrossRef]
- DeMoranville, C. Cranbery best management practice adoption and conservation farm planning in Massachusetts. Hort. Tech. 2006, 16, 393–397. [Google Scholar]
- Zhang, F.; Wei, Z.; Jeranyama, P.; Demoranville, C.; Hou, H.J.M. A significant loss in photosynthetic activity associated with the yellow vine syndrome of cranberry. HortScience 2011, 49, 901–907. [Google Scholar]
- DeMoranville, C.; Howes, B.; Schlezinger, D.; White, D. Cranberry phosphorus management: How changes in practice can reduce output in drainage water. Acta Hortic. 2009, 810, 633–640. [Google Scholar]
- Oguchi, R.; Jia, H.; Barber, J.; Chow, W.S. Recovery of photoinactivated photosystem II in leaves: Retardation due to restricted mobility of photosystem II in the thylakoid membrane. Photosynth. Res. 2008, 98, 621–629. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Hou, H.J.M. Photosynthesis and Yellow Vine Syndrome of American Cranberry. Agriculture 2012, 2, 125-138. https://doi.org/10.3390/agriculture2020125
AMA Style
Hou HJM. Photosynthesis and Yellow Vine Syndrome of American Cranberry. Agriculture. 2012; 2(2):125-138. https://doi.org/10.3390/agriculture2020125
Chicago/Turabian StyleHou, Harvey J. M. 2012. "Photosynthesis and Yellow Vine Syndrome of American Cranberry" Agriculture 2, no. 2: 125-138. https://doi.org/10.3390/agriculture2020125