
Lipidomics to Assess Omega 3 Bioactivity


Department of Molecular Medicine, University of Padova, Via 8 Febbraio, 2-35122 Padova, Italy
J. Clin. Med. 2015, 4(9), 1753-1760; https://doi.org/10.3390/jcm4091753
Submission received: 15 June 2015
/
Revised: 19 August 2015
/
Accepted: 31 August 2015
/
Published: 7 September 2015
(This article belongs to the Special Issue Omega-3 Fatty Acids in Health and Disease)
{kind=link}
Abstract
:How can we resolve the conflict between the strong epidemiological evidence pointing to the usefulness of fish—and, thus, omega 3—consumption with the debacle of supplementation trials? One potential explanation is that the null results obtained thus far are the consequences of ill-contrived investigations that do not allow us to conclude on the effects (or lack thereof) of omega 3 fatty acid supplementation. One potential solution is through the use of lipidomics, which should prove very useful to screen suitable patients and to correlate plasma (or red blood cells, or whole blood, or phospholipid) fatty acid profile with outcomes. This has never been done in omega 3 trials. The wise use of lipidomics should be essential part of future omega 3 trials and would help in untangling this current riddle.
1. Introduction
Fish consumption is directly associated with better cardiovascular prognosis [1]. From the first observations made in Greenland to the latest randomized controlled trials (RCT), long-chain omega 3 fatty acids, namely docosahexaenoic (DHA) and eicosapentaenoic (EPA) fatty acids have been held responsible for the health effects of fish [1]. One notable example is that of the omega 3 index, which is influenced by omega 3 consumption and is a strong predictor of cardiovascular risk [2]. Consequently, much research has been devoted to elucidating, in vitro and in vivo, the protective activities of EPA and DHA; indeed, the general perception is that supplementary intake of these fatty acids would protect from cardiovascular disease and other degenerative pathologies. Suggesting consumption of adequate amounts of omega 3 fatty acids actually makes biological sense; these fatty acids are important in that mammals cannot efficiently synthesize them and they are indispensable to a number of biological processes, especially those that involve excitable cells such as cardiomyocytes, neurons, retinal cells, etc. [1]. In addition, nearly every randomized clinical trial reported that omega 3 fatty acids positively modulate surrogate markers of cardiovascular disease and in vitro studies explored a wide range of mechanisms underlying the purported beneficial activities of EPA and DHA. The sad truth is that, when given as supplements, long chain omega 3 fatty acids are—very likely—ineffective in affording better health [3]. The big question is why [4]? What is wrong [5]? Indeed, this is probably the major current conundrum in pharma-nutrition research, with fish oils threatening to join the vitamin E, beta-carotene, resveratrol, etc. in the dustbin of promising nutritional therapies that failed to show benefit in RCTs [6,7,8].
2. Omega 3 Fatty Acids in Cardiovascular Therapy
How can we reconcile this apparent failure with strong epidemiological evidence pointing to the usefulness of fish—and, thus, omega 3—consumption? One potential explanation is that the null results obtained thus far are the consequences of ill-contrived trials that do not allow one to draw firm conclusions on the effects (or lack thereof) of omega 3 fatty acid supplementation [9].
In this respect, one paradigmatic example is that of folate in stroke prevention. In a well-designed trial recently published by Huo et al. [10], the highest risk of stroke and the greatest benefit of folic acid therapy were seen in patients with the lowest baseline folate levels. In addition, Huo et al. [10] suggest that individuals with the TT genotype may require a higher dosage of folic acid supplementation to overcome biologically-insufficient levels (as reflected in the relatively greater folate requirement for subjects with the TT genotype).
Unfortunately, neither basal plasma concentrations nor genotype/nutrigenomics issues have ever been taken into consideration in omega 3 supplementation studies [9]. The natural consequence is that the near totality of omega 3 trials could not really conclude that modulation of omega 3 status is ineffective. This situation is unheard of in classic drug research, where surrogate markers, e.g., blood pressure of cholesterol concentrations, are mandatorily assessed before and after interventions on these risk factors.
In this respect, the use of lipidomics in RCTs should prove very useful to screen patients first and to correlate plasma (or red blood cells, or whole blood, or phospholipid) fatty acid profile with outcomes.
3. Lipidomics
From a technical viewpoint, lipidomics can be conveniently classified into as two distinct approaches: A “shotgun”, i.e., electrospray ionization coupled with single stage or tandem mass spectrometry approach and a liquid chromatography-mass spectrometry (LC-MS)-based one. The shotgun approach does not require separation prior to MS analyses. The shotgun approach is being considered as the best method for analysis of lipid extracts. Its major advantage over other methods is that mixtures of lipids are directly infused and sprayed into the mass spectrometer, therefore analyzing all the molecules collectively [11]. In this way, shotgun lipidomics, when applied to whole tissue lipid extracts, generates molecular ions without extensive fragmentation of the molecules. This type of lipidomics allows the identification and quantitation of the lipids in complex mixtures and provides helpful global information about cellular and tissue lipidomes. The procedures generate quasi-molecular ions in high yields. The major limitation of shotgun lipidomics is that the most abundant or most efficiently ionized molecules “quench” the signal of other ones, in turn causing ion suppression and hampering detection of lipids that are present in lower concentrations [12]. Therefore, intra-source separation is often employed to overcome this limitation. Mostly, the analytical setting of lipidomics involves a triple-quadrupole mass spectrometer with electrospray ionization (ESI, primarily negative mode) in selective reaction monitoring (SRM) or multiple reaction monitoring (MRM) to maximize sensitivity [12]. Whereas reversed-phase chromatography, i.e., C18, is the first separation choice, hydrophilic interaction liquid chromatography (HILIC) is best for phospholipids, sphingolipids, and phosphatidylcholines [12]. In summary, the most appropriate methodological approach should be based on the goals of the analysis and, in the future, we might witness a combination of techniques that would match ease of use with accuracy.
When mass spectral equipment is available in clinical laboratories, lipidomics can be applied to clinical studies. Current bottlenecks include sample handling methods and data reduction and analysis of the enormous volume of data generated by contemporary mass spectrometric analysis. However, this rapidly evolving field can rapidly elucidate the lipidome in medical samples of small size. Of note, current techniques can identify thousands of distinct lipid molecular species, of which the most biologically important ones are fatty acyls, glycerolipids (neutral), glycerophospholipids (polar), sphingolipids, sterols, and prenols [13,14].
4. Lipidomics Applied to Fatty Acid Research
As mentioned, the lipidome provides useful information on the basal omega 3 status of patients or healthy volunteers and can greatly help resolve the current omega 3 conundrum in cardiovascular research/clinic. To date, lipidomics have been applied to observational and preclinical models of cardiovascular disease, including mice, hamsters, rabbits, and pigs, and nonhuman primates.
Example of observational research include the use of lipid profiling in diabetes mellitus [15,16], cardiovascular disease [17,18], metabolic disorders [19], dietary habits, and obesity [20], as well as in determining response to drug therapy (elegantly reviewed by Hinterwirth et al. [13]). These techniques can provides useful information on the lipid composition and profile of whole blood or of tissues e.g., the retina [21], atherosclerotic plaques [22], muscles [23], or adipose tissue [24].
In terms of cardiovascular disease and lipid profiling, some examples are worth our attention. The use of olive oil as a predominant source of fat has been consistently associated with better cardiovascular prognosis. Most researchers are still attributing the purported healthful properties of olive oil to its relatively high monounsaturated fatty acids (MUFAs) content [25]. The theory behind this is that MUFAs, namely oleic acid, favorably modifies the high-density lipoprotein/low-density lipoprotein (HDL/LDL) ratio and, therefore, afford cardiovascular protection [25]. Yet, accumulated evidence clearly indicates that MUFAs only mildly modify the HDL/LDL ratio, to an extent that should minimally influence cardiovascular risk [26]. Moreover, at least two lipidomic studies [27,28] show that the proportion of MUFAs is higher in red blood cells and whole blood, respectively, of myocardial infarct patients as compared with controls, confirming findings from the large dataset of Stegeman et al. [17]. In summary, the hypothesis that MUFAs are cardioprotective and that increasing their intake would benefit the cardiovascular system is still unproven.
Lipidomics have also been employed in pharmaceutical research [11], namely to assess the effects of selected drugs on hypertension [29] and cardiovascular disease, in particular when recombinant HDL [16] and statins [30] were employed.
Within the context of this paper, it is worth reiterating that lipidomics have never been applied to omega 3 intervention trials and that precisely this improper approach is leading to pallid conclusions on their therapeutic roles.
5. Examples of Lipidomics Applied to Omega 3 Research
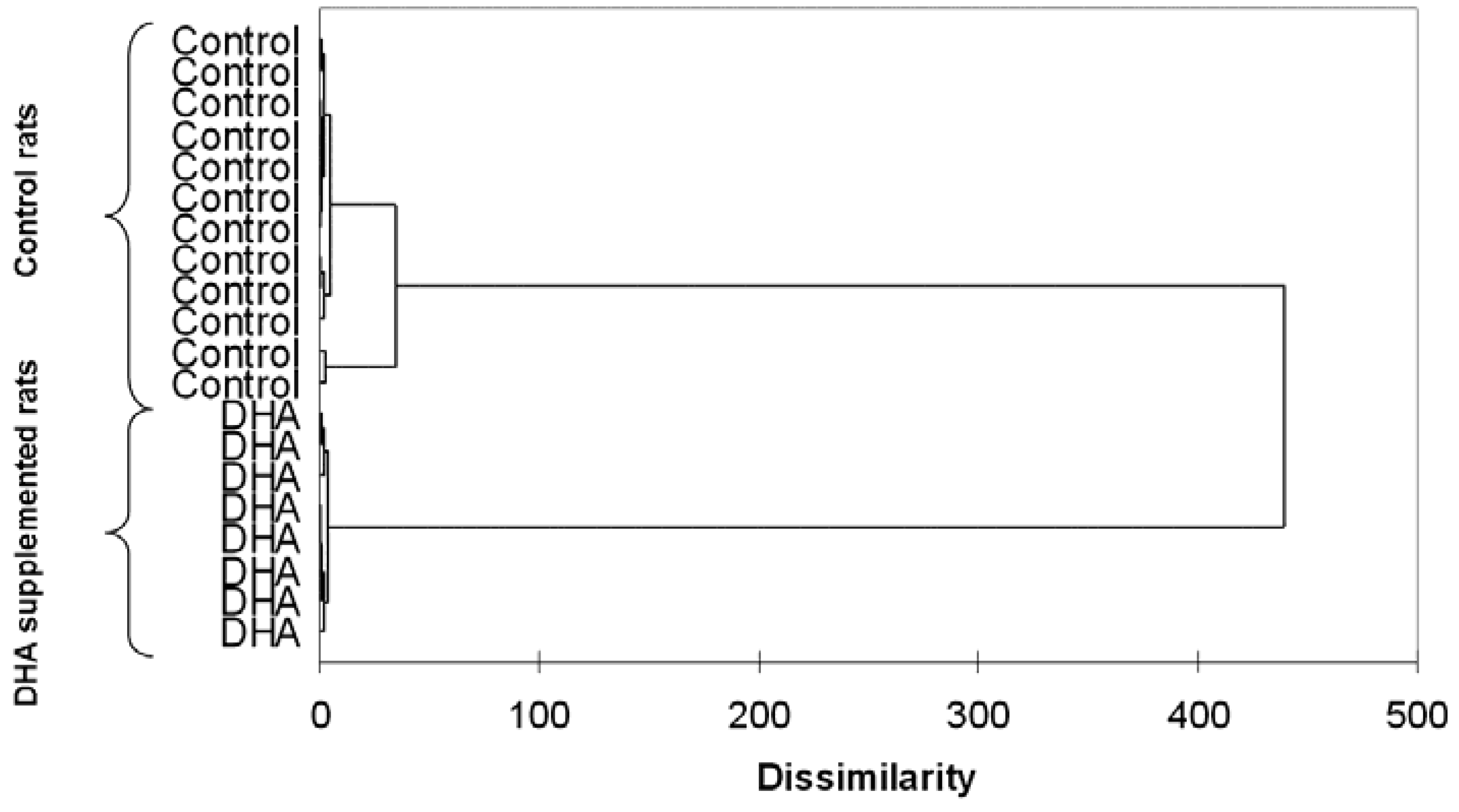
As mentioned, the near totality of omega 3 trials did not include baseline or post-treatment evaluations of omega 3 status in patients of healthy volunteers. In rats, lipidomics have been successfully employed by Lamaziere and coworkers to assess the effects of omega 3 supplementation on hepatic lipogenesis [31] and brain composition [32]. Additionally, this technique allowed the identification of the in vitro concentrations to be employed in cardiac biochemistry studies [33]. It is worth noting that, even though rats are given identical feed, their tissutal omega 3 status varies greatly [31,32,33]. One can only speculate that these differences are enhanced in humans who consume varied diets. Additionally, omega 3 supplementation reduces such inter-individual variations (Figure 1) and renders the omega 3 composition of organs and tissues more homogeneous [31,32,33].
Figure 1.
Example of lipidomics applied to omega 3 research: dendrogram resulting from the hierarchical cluster analysis of atrial rat cardiomyocytes isolated from control and docosahexaenoic (DHA)-treated rats. Unpublished data. Methodological details can be found in Lamaziere et al. [33].
Figure 1.
Example of lipidomics applied to omega 3 research: dendrogram resulting from the hierarchical cluster analysis of atrial rat cardiomyocytes isolated from control and docosahexaenoic (DHA)-treated rats. Unpublished data. Methodological details can be found in Lamaziere et al. [33].
Another mechanistic application of lipidomics to omega 3 research concerns the differential distribution of DHA and EPA in the various lipid classes. As an example, the former mostly accumulates into phospholipids whereas the latter contributes to cholesterol esters [34]. Likely, the biological importance of phospholipids is higher than that of cholesterol esters because they originate from important lipid mediators; their compositional modulation by diet or supplement should result into more relevant biological actions.
Finally, we should not forget that the in vivo effects of omega 3 fatty acids likely depend on capsules’ formulation, i.e., tryacylglycerols vs. ethyl esters vs. phospholipids [35] and on whether they are taken as supplements or as part of food items (either food as such [36] or functional food such as milk [37,38]). It is conceivable that omega 3 fatty acids’ bioavailability and subsequent distribution in the body might be different depending on the matrix [35]. This can be explored by proper lipidomic approaches.
In recent years, much investigation has been dedicated to bioactive small molecules that activate the resolution of acute inflammation [39]. These molecules are autacoids that include resolvins, protectins, and maresins, which are formed from EPA and DHA during acute inflammation. Interestingly, resolvins and protectins can currently be synthesized [40], which paves the way to their future use in therapy. Lipid-mediator lipidomic profiling of self-resolving exudates (after solid-phase extraction) allowed framing the time course of inflammation resolution and elucidating the roles played by these important omega 3-derived molecules [40]. Alas, the lipidomic methods that have been successfully employed to clarify the lipid mediator profiles of phagocytes and inflammatory exudates [41] have never been employed in omega 3 clinical trials, thus preventing drawing firm conclusions on their real contribution to the resolution of inflammation in humans.
6. Conclusions
While the assessment of relevant biomarkers, e.g., blood pressure, glycaemia, or cholesterol is habitual in pharmacological research, research in the omega 3 area has been hampered by the lack of baseline and post-treatment measurements of fatty acid levels. This prevents drawing solid conclusions on the true healthful role of these essential fatty acids. While it is tempting—based on cumulated evidence—to state that the omega 3 myth has been debunked, we need much more focused research before solid conclusions be reached and before we can educe evidence-based recommendations for or against using omega 3 supplements. This is even more relevant for cardiovascular patients who, as reviewed elsewhere [9], are treated with effective and potent drugs and might enjoy additional benefits from omega 3 supplementation only if their basal concentrations are low, i.e., with an omega 3 index <4%.
The wise use of lipidomics should be essential part of future omega 3 trials and would help untangle this current riddle.
Conflicts of Interest
The author declares no conflict of interest.
References
- Richard, D.; Bausero, P.; Schneider, C.; Visioli, F. Polyunsaturated fatty acids and cardiovascular disease. Cell. Mol. Life Sci. 2009, 66, 3277–3288. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; von Schacky, C. The omega-3 index: A new risk factor for death from coronary heart disease? Prev. Med. 2004, 39, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Messori, A.; Fadda, V.; Maratea, D.; Trippoli, S. Omega-3 fatty acid supplements for secondary prevention of cardiovascular disease: From “no proof of effectiveness” to “proof of no effectiveness”. JAMA Intern. Med. 2013, 173, 1466–1468. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, N.A.; Tappia, P.S. Why are there inconsistencies in the outcomes of some omega-3 fatty acid trials for the management of cardiovascular disease? Clin. Lipidol. 2015, 10, 27–32. [Google Scholar] [CrossRef]
- Visioli, F. Cover story: What is wrong with omega 3 fatty acids? PharmaNutrition 2014, 2, A1. [Google Scholar] [CrossRef]
- Moyer, V.A. Vitamin, mineral, and multivitamin supplements for the primary prevention of cardiovascular disease and cancer: U.S. preventive services task force recommendation statement. Ann. Intern. Med. 2014, 160, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Bjelakovic, G.; Nikolova, D.; Gluud, C. Antioxidant supplements to prevent mortality. JAMA 2013, 310, 1178–1179. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F. The resveratrol fiasco. Pharmacol. Res. 2014, 90, 87. [Google Scholar] [CrossRef] [PubMed]
- Von Schacky, C. Omega-3 fatty acids in cardiovascular disease—An uphill battle. Prostaglandins Leukot. Essent. Fatty Acids 2015, 92, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Huo, Y.; Li, J.; Qin, X.; Huang, Y.; Wang, X.; Gottesman, R.F.; Tang, G.; Wang, B.; Chen, D.; He, M.; et al. Efficacy of folic acid therapy in primary prevention of stroke among adults with hypertension in china: The csppt randomized clinical trial. JAMA 2015, 313, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Lamaziere, A.; Wolf, C.; Quinn, P.J. Application of lipidomics to assess lipogenesis in drug development and pre-clinical trials. Curr. Pharm. Biotechnol. 2012, 13, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Kasuga, K.; Suga, T.; Mano, N. Bioanalytical insights into mediator lipidomics. J. Pharm. Biomed. Anal. 2015, 113, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Hinterwirth, H.; Stegemann, C.; Mayr, M. Lipidomics: Quest for molecular lipid biomarkers in cardiovascular disease. Circ. Cardiovasc. Genet. 2014, 7, 941–954. [Google Scholar] [CrossRef] [PubMed]
- Quehenberger, O.; Armando, A.M.; Brown, A.H.; Milne, S.B.; Myers, D.S.; Merrill, A.H.; Bandyopadhyay, S.; Jones, K.N.; Kelly, S.; Shaner, R.L.; et al. Lipidomics reveals a remarkable diversity of lipids in human plasma. J. Lipid Res. 2010, 51, 3299–3305. [Google Scholar] [CrossRef] [PubMed]
- Rhee, E.P.; Cheng, S.; Larson, M.G.; Walford, G.A.; Lewis, G.D.; McCabe, E.; Yang, E.; Farrell, L.; Fox, C.S.; O’Donnell, C.J.; et al. Lipid profiling identifies a triacylglycerol signature of insulin resistance and improves diabetes prediction in humans. J. Clin. Investig. 2011, 121, 1402–1411. [Google Scholar] [CrossRef] [PubMed]
- Drew, B.G.; Carey, A.L.; Natoli, A.K.; Formosa, M.F.; Vizi, D.; Reddy-Luthmoodoo, M.; Weir, J.M.; Barlow, C.K.; van Hall, G.; Meikle, P.J.; et al. Reconstituted high-density lipoprotein infusion modulates fatty acid metabolism in patients with type 2 diabetes mellitus. J. Lipid Res. 2011, 52, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Stegemann, C.; Pechlaner, R.; Willeit, P.; Langley, S.R.; Mangino, M.; Mayr, U.; Menni, C.; Moayyeri, A.; Santer, P.; Rungger, G.; et al. Lipidomics profiling and risk of cardiovascular disease in the prospective population-based bruneck study. Circulation 2014, 129, 1821–1831. [Google Scholar] [CrossRef] [PubMed]
- Sysi-Aho, M.; Koikkalainen, J.; Seppanen-Laakso, T.; Kaartinen, M.; Kuusisto, J.; Peuhkurinen, K.; Karkkainen, S.; Antila, M.; Lauerma, K.; Reissell, E.; et al. Serum lipidomics meets cardiac magnetic resonance imaging: Profiling of subjects at risk of dilated cardiomyopathy. PLoS ONE 2011, 6, e15744. [Google Scholar] [CrossRef] [PubMed]
- Stubiger, G.; Aldover-Macasaet, E.; Bicker, W.; Sobal, G.; Willfort-Ehringer, A.; Pock, K.; Bochkov, V.; Widhalm, K.; Belgacem, O. Targeted profiling of atherogenic phospholipids in human plasma and lipoproteins of hyperlipidemic patients using MALDI-QIT-TOF-MS/MS. Atherosclerosis 2012, 224, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.P.; Montoliu, I.; Collino, S.; Scherer, M.; Guy, P.; Tavazzi, I.; Thorimbert, A.; Moco, S.; Rothney, M.P.; Ergun, D.L.; et al. Topographical body fat distribution links to amino acid and lipid metabolism in healthy obese women (corrected). PLoS ONE 2013, 8, e73445. [Google Scholar] [CrossRef] [PubMed]
- Schwartzman, M.L.; Iserovich, P.; Gotlinger, K.; Bellner, L.; Dunn, M.W.; Sartore, M.; Grazia Pertile, M.; Leonardi, A.; Sathe, S.; Beaton, A.; et al. Profile of lipid and protein autacoids in diabetic vitreous correlates with the progression of diabetic retinopathy. Diabetes 2010, 59, 1780–1788. [Google Scholar] [CrossRef] [PubMed]
- Spijkers, L.J.; van den Akker, R.F.; Janssen, B.J.; Debets, J.J.; de Mey, J.G.; Stroes, E.S.; van den Born, B.J.; Wijesinghe, D.S.; Chalfant, C.E.; MacAleese, L.; et al. Hypertension is associated with marked alterations in sphingolipid biology: A potential role for ceramide. PLoS ONE 2011, 6, e21817. [Google Scholar] [CrossRef] [PubMed]
- Kien, C.L.; Bunn, J.Y.; Poynter, M.E.; Stevens, R.; Bain, J.; Ikayeva, O.; Fukagawa, N.K.; Champagne, C.M.; Crain, K.I.; Koves, T.R.; et al. A lipidomics analysis of the relationship between dietary fatty acid composition and insulin sensitivity in young adults. Diabetes 2013, 62, 1054–1063. [Google Scholar] [CrossRef] [PubMed]
- Pietilainen, K.H.; Rog, T.; Seppanen-Laakso, T.; Virtue, S.; Gopalacharyulu, P.; Tang, J.; Rodriguez-Cuenca, S.; Maciejewski, A.; Naukkarinen, J.; Ruskeepaa, A.L.; et al. Association of lipidome remodeling in the adipocyte membrane with acquired obesity in humans. PLoS Biol. 2011, 9, e1000623. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Bernardini, E. Extra virgin olive oil’s polyphenols: Biological activities. Curr. Pharm. Des. 2011, 17, 786–804. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Micha, R.; Wallace, S. Effects on coronary heart disease of increasing polyunsaturated fat in place of saturated fat: A systematic review and meta-analysis of randomized controlled trials. PLoS Med. 2010, 7, e1000252. [Google Scholar] [CrossRef] [PubMed]
- Marangoni, F.; Novo, G.; Perna, G.; Perrone Filardi, P.; Pirelli, S.; Ceroti, M.; Querci, A.; Poli, A. Omega-6 and omega-3 polyunsaturated fatty acid levels are reduced in whole blood of italian patients with a recent myocardial infarction: The AGE-IM study. Atherosclerosis 2014, 232, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Block, R.C.; Harris, W.S.; Reid, K.J.; Spertus, J.A. Omega-6 and trans fatty acids in blood cell membranes: A risk factor for acute coronary syndromes? Am. Heart J. 2008, 156, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Kong, H.; Qu, F.; Li, Y.; Yu, Z.; Gao, P.; Peng, S.; Xu, G. Application of plasma lipidomics in studying the response of patients with essential hypertension to antihypertensive drug therapy. Mol. Biosyst. 2011, 7, 3271–3279. [Google Scholar] [CrossRef] [PubMed]
- Kaddurah-Daouk, R.; Baillie, R.A.; Zhu, H.; Zeng, Z.B.; Wiest, M.M.; Nguyen, U.T.; Watkins, S.M.; Krauss, R.M. Lipidomic analysis of variation in response to simvastatin in the cholesterol and pharmacogenetics study. Metabolomics 2010, 6, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Lamaziere, A.; Wolf, C.; Barbe, U.; Bausero, P.; Visioli, F. Lipidomics of hepatic lipogenesis inhibition by omega 3 fatty acids. Prostaglandins Leukot. Essent. Fatty Acids 2013, 88, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Lamaziere, A.; Richard, D.; Barbe, U.; Kefi, K.; Bausero, P.; Wolf, C.; Visioli, F. Differential distribution of dha-phospholipids in rat brain after feeding: A lipidomic approach. Prostaglandins Leukot. Essent. Fatty Acids 2011, 84, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Lamaziere, A.; Richard, D.; Bausero, P.; Barbe, U.; Kefi, K.; Wolf, C.; Visioli, F. Comparison of docosahexaenoic acid uptake in murine cardiomyocyte culture and tissue: Significance to physiologically relevant studies. Prostaglandins Leukot. Essent. Fatty Acids 2015, 94, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Vidgren, H.M.; Agren, J.J.; Schwab, U.; Rissanen, T.; Hanninen, O.; Uusitupa, M.I. Incorporation of n-3 fatty acids into plasma lipid fractions, and erythrocyte membranes and platelets during dietary supplementation with fish, fish oil, and docosahexaenoic acid-rich oil among healthy young men. Lipids 1997, 32, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, J.P.; Hahn, A. Bioavailability of long-chain omega-3 fatty acids. Prostaglandins Leukot. Essent. Fatty Acids 2013, 89, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Rise, P.; Barassi, M.C.; Marangoni, F.; Galli, C. Dietary intake of fish vs. formulations leads to higher plasma concentrations of n-3 fatty acids. Lipids 2003, 38, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Molinari, C.; Risé, P.; Guerra, C.; Mauro, N.; Piani, C.; Bosi, E.; Galli, C.; Scavini, M. Eight-week consumption of milk enriched with omega 3 fatty acids raises their blood concentrations yet does not affect lipids and cardiovascular disease risk factors in adult healthy volunteers. Pharmanutrition 2014, 2, 141–148. [Google Scholar] [CrossRef]
- Visioli, F.; Rise, P.; Plasmati, E.; Pazzucconi, F.; Sirtori, C.R.; Galli, C. Very low intakes of n-3 fatty acids incorporated into bovine milk reduce plasma triacylglycerol and increase HDL-cholesterol concentrations in healthy subjects. Pharmacol. Res. 2000, 41, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Dalli, J. The resolution code of acute inflammation: Novel pro-resolving lipid mediators in resolution. Semin. Immunol. 2015, 27, 200–215. [Google Scholar] [CrossRef] [PubMed]
- Dalli, J.; Winkler, J.W.; Colas, R.A.; Arnardottir, H.; Cheng, C.Y.; Chiang, N.; Petasis, N.A.; Serhan, C.N. Resolvin D3 and aspirin-triggered resolvin D3 are potent immunoresolvents. Chem. Biol. 2013, 20, 188–201. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Chiang, N.; Oh, S.F.; Serhan, C.N. Metabolomics-lipidomics of eicosanoids and docosanoids generated by phagocytes. Curr. Protoc. Immunol. 2011. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Visioli, F. Lipidomics to Assess Omega 3 Bioactivity. J. Clin. Med. 2015, 4, 1753-1760. https://doi.org/10.3390/jcm4091753
AMA Style
Visioli F. Lipidomics to Assess Omega 3 Bioactivity. Journal of Clinical Medicine. 2015; 4(9):1753-1760. https://doi.org/10.3390/jcm4091753
Chicago/Turabian StyleVisioli, Francesco. 2015. "Lipidomics to Assess Omega 3 Bioactivity" Journal of Clinical Medicine 4, no. 9: 1753-1760. https://doi.org/10.3390/jcm4091753