
Role of MicroRNAs-221/222 in Digestive Systems


1
Center for Preventive Medicine, Keio University Hospital, Tokyo 160-0016, Japan
2
Division of Gastroenterology and Hepatology, Department of Internal Medicine, Keio University School of Medicine, 35 Shinanomachi, Shinjuku-ku, Tokyo 160-8582, Japan
*
Author to whom correspondence should be addressed.
J. Clin. Med. 2015, 4(8), 1566-1577; https://doi.org/10.3390/jcm4081566
Submission received: 29 June 2015
/
Revised: 17 July 2015
/
Accepted: 22 July 2015
/
Published: 6 August 2015
(This article belongs to the Special Issue MicroRNAs: Novel Biomarkers and Therapeutic Targets for Human Cancers)
Abstract
:MiR-221 and miR-222 (miR-221/222) are well-studied oncogenic microRNAs that are frequently upregulated in several types of human tumors, such as esophageal adenocarcinoma, gastric adenocarcinoma, colorectal adenocarcinoma, hepatocellular carcinoma, and pancreatic ductal adenocarcinoma. In these cancers, silencing miR-221/222 could represent a novel anti-tumor approach to inhibit tumor growth and metastasis. On the other hand, miR-221/222 also play onco-suppressive roles in cholangiocarcinoma and gastrointestinal stromal tumors (GISTs). Here we will review the roles of miR-221/222 in digestive systems and their possibility as prognostic and therapeutic tools.
1. Introduction
MicroRNAs (miRs) are ~22 nucleotide noncoding RNAs that can downregulate various gene products by translational repression when partially complementary sequences are present in the 3′ untranslated regions (3′ UTR) of the target mRNAs or by directing mRNA degradation. Increasing evidence has demonstrated that miRs are involved in cancer initiation, progression, and metastasis, and may serve as diagnostic and prognostic biomarkers for cancers. Among the many miRNAs already identified as regulators of neoplastic transformation, invasion, and metastasis, miR-221 and miR-222 (miR-221/222) have emerged as key miRNAs deregulated in many cancers, such as gastrointestinal cancers, breast cancer, prostate cancer, thyroid cancer, and glioma [1,2,3,4,5]. MiR-221 and miR-222 are encoded in tandem from a gene cluster located on chromosome Xp11.3. Several reports indicated that miR-221/222 could be used as a therapeutic tool to decrease cell proliferation or modulate sensitivity to anti-cancer agents [6,7,8]. Here we review the current knowledge about the role of miR-221/222 in digestive systems, including hepatobiliary and pancreatic cancers.
2. Direct Targets of miR-221/222
The identification of target mRNAs is a key step for assessing the role of aberrantly expressed microRNAs in human cancer. To date, various direct targets of miR-221/222 have been reported, even in the digestive system, as shown in Table 1. Among them, regulation of p27Kip1 by miR-221/222 is well studied. Downregulation of p27Kip1 is required for cell cycle entry after growth factor stimulation. MiR-221/222 are underactive towards p27Kip1-3′ UTRs in quiescent cells, as a result of target site hindrance. Pumilio-1 (PUM1) is a ubiquitously expressed RNA-binding protein (RBP) that interacts with p27Kip1-3′ UTR. In response to growth factor stimulation, PUM1 is upregulated and phosphorylated for optimal induction of its RNA-binding activity towards the p27Kip1-3′ UTR [9]. PUM1 binding induces a local change in RNA structure that favors association with miR-221/222, efficient suppression of p27Kip1 expression, and rapid entry to the cell cycle.

{kind=link}
| Target | Cancer Type | Reference |
|---|---|---|
| p27Kip1 | Esophageal adenocarcinoma | Matsuzaki et al. (2013) |
| Hepatocellular carcinoma | Pineau et al. (2010), Fu et al. (2011), Callegari et al. (2012) | |
| Pancreatic ductal adenocarcinoma | Park et al. (2009), Sarkar et al. (2013), Tanaka et al. (2015) | |
| p57Kip2 | Colorectal adenocarcinoma | Sun et al. (2011) |
| Pancreatic ductal adenocarcinoma | Sarkar et al. (2013) | |
| PTEN | Gastric adenocarcinoma | Chun-Zhi et al. (2010) |
| Hepatocellular carcinoma | Fornari et al. (2008), Callegari et al. (2012), Garofalo et al. (2009) | |
| Colorectal adenocarcinoma | Tsunoda et al. (2011), Xue et al. (2013) | |
| Pancreatic ductal adenocarcinoma | Sarkar et al. (2013) | |
| RelA | Colorectal adenocarcinoma | Liu et al. (2014) |
| PDLIM2 | Colorectal adenocarcinoma | Liu et al. (2014) |
| RECK | Colorectal adenocarcinoma | Qin et al. (2014) |
| BMF | Hepatocellular carcinoma | Gramantieri et al. (2009), Callegari et al. (2012), He et al. (2014) |
| BBC3 | Hepatocellular carcinoma | He et al. (2014) |
| ANGPTL2 | Hepatocellular carcinoma | He et al. (2014) |
| HDAC6 | Hepatocellular carcinoma | Bae et al. (2015) |
| ERα | Hepatocellular carcinoma | Chen et al. (2015) |
| SOCS1 | Hepatocellular carcinoma | Xu et al. (2014) |
| SOCS3 | Hepatocellular carcinoma | Xu et al. (2014) |
| MDM2 | Hepatocellular carcinoma | Fornari et al. (2014) |
| DDIT4 | Hepatocellular carcinoma | Pineau et al. (2010) |
| TIMP3 | Hepatocellular carcinoma | Garofalo et al. (2009) |
| TIMP2 | Pancreatic ductal adenocarcinoma | Xu et al. (2015) |
| PIK3R1 | Colangiocarcinoma | Okamoto et al. (2013) |
| PUMA | Pancreatic ductal adenocarcinoma | Sarkar et al. (2013) |
| TRPS1 | Pancreatic ductal adenocarcinoma | Su et al. (2013) |
| KIT | Gastrointestinal stromal tumor | Koelz et al. (2011), Gits et al. (2013), Ihle et al. (2015) |
3. Esophageal Cancer
Duodeno-gastro-esophageal bile reflux contributes to development of esophageal adenocarcinoma. We recently reported that expression levels of miR-221/222 increased, along with the activity of nuclear bile acid receptor/farnesoid X receptor (FXR), when cultured esophageal epithelial cells were exposed to bile acids [10]. Furthermore, miR-221/222 expression was higher in esophageal adenocarcinoma than in the surrounding Barrett’s esophagus, a precursor lesion of esophageal adenocarcinoma. p27Kip1 is known to inhibit the proteasomal protein degradation of the transcription factor CDX2. We also confirmed that the levels of p27Kip1 and CDX2 were lower in areas of esophageal adenocarcinoma than in those of Barrett’s esophagus. Incubation of cells with bile acids increased degradation of CDX2; this process was reduced when cells were also incubated with proteasome inhibitors. Overexpression of miR-221/222 reduced levels of p27Kip1 and CDX2, and knockdown of miR-221/222 increased levels of these proteins in cultured cells. In addition, inhibitors of miR-221/222 reduced growth of xenograft tumors in immunodeficient mice.
4. Gastric Cancer
Liu et al. reported that miR-221 was upregulated in 88% of gastric cancer tissue samples compared with their paired adjacent non-tumor tissue samples [11]. High expression of miR-221 showed a significant correlation with advanced tumor-node-metastasis stage, local invasion, and lymphatic metastasis. MiR-221 overexpression was an unfavorable prognostic factor for overall survival in patients with gastric cancer.
In gastric cancer cells, upregulation of miR-221/222 induced the malignant phenotype, whereas knockdown of miR-221/222 reversed this phenotype via induction of PTEN, a direct target of miR-221/222 [12]. In addition, knockdown of miR-221/222 inhibited cell growth and invasion and increased the radiosensitivity.
5. Colorectal Cancer
MiR-221 was upregulated in 90% of colorectal cancer (CRC) tissue samples compared to that in the adjacent non-tumorous tissue, and the expression level was positively correlated to an advanced TNM stage and local invasion [13,14,15,16,17,18]. A survival analysis indicated that high expression of miR-221 was closely associated with a shorter survival time [14,19]. In CRC cells, miR-221 overexpression enhances, whereas miR-221 depletion reduces CRC cell proliferation, migration, invasion, and colony formation [16,17]. In mice with colitis, injection of lentiviruses expressing miR-221/222 sponges led to formation of fewer tumors than injection of control lentiviruses [16]. Protein expressions of p57Kip2 and RECK, direct targets of miR-221, were decreased in the CRC tissues, and promoted CRC occurrence and progress [15,17].
Liu et al. reported that mimics of miR-221/222 activated NF-κB and STAT3 in CRC cells [16]. MiR-221/222 also reduced the ubiquitination and degradation of the RelA and STAT3 proteins by binding to the 3′ untranslated region of PDLIM2 mRNA (PDLIM2 is a nuclear ubiquitin E3 ligase for RelA and STAT3). In human CRC tissues, levels of miR-221/222 positively correlated with levels of RelA and STAT3 mRNAs. Levels of PDLIM2 mRNA were lower than non-tumor tissues.
Xue et al. investigated the regulative effect of miR-221 on CRC cell radiosensitivity [20]. X-ray radiation had an effect on the expression of miR-221 in CRC cells in a dose-dependent manner. The protein levels of PTEN, a direct target of miR-221, reduced gradually during exposure to X-rays. Inhibition of miR-221 upregulated expression of PTEN protein and enhanced the radiosensitivity. Moreover, the inhibitory effect was dramatically abolished by pretreatment with anti-PTEN-siRNA, suggesting that the enhancement of radiosensitivity was mediated by PTEN.
Tsunoda et al. reported that the increased expression of miR-221/222 was observed in 3D culture as compared with 2D culture [18]. They showed that miR-221/222 was regulated by oncogenic KRAS, which plays several key roles in 3D culture. The protein expression level of PTEN was reduced under the control of KRAS in a 3D-specific manner.
The plasma concentration of miR-221 is a potential biomarker for differentiating CRC patients from controls [21]. Kaplan–Meier curve assessment shows that the elevated plasma miR-221 level is a significant prognostic factor for poor overall survival in CRC patients. The immunohistochemistry analysis demonstrates a significant correlation between plasma miR-221 level and p53 expression.
Stool-based miR-221 can also be used as a non-invasive biomarker for the detection of CRC [13]. In stool samples, miR-221 showed a significant increasing trend from normal controls to late stages of CRC. The AUC of stool miR-221 was 0.73 for CRC patients as compared with normal controls. No significant differences in stool miR-221 levels were found between patients with proximal and distal CRCs. The use of antibiotics did not influence stool miR-221 levels.
6. Hepatocellular Carcinoma
MiR-221/222 is a critical modulator in the hepatocellular carcinoma (HCC) signaling pathway [22]. MiR-221/222 was upregulated in the human liver in a fibrosis progression-dependent manner with upregulation of α1 (I) collagen (COL1A1) and α-smooth muscle actin (αSMA) [23,24,25]. Upregulation of miR-221 and downregulation of p27Kip1 and p57Kip2 were associated with tumor stages, local recurrence, metastasis, and poor prognosis [24,25,26,27,28,29]. In a mouse model of liver cancer, miR-221 overexpression stimulated growth of tumorigenic murine hepatic progenitor cells [30,31]. Inhibition of miR-221 decreased liver cancer cell proliferation, clonogenicity, migration, and invasion and also induced G1 arrest and apoptosis in vitro and in vivo [22,27,32].
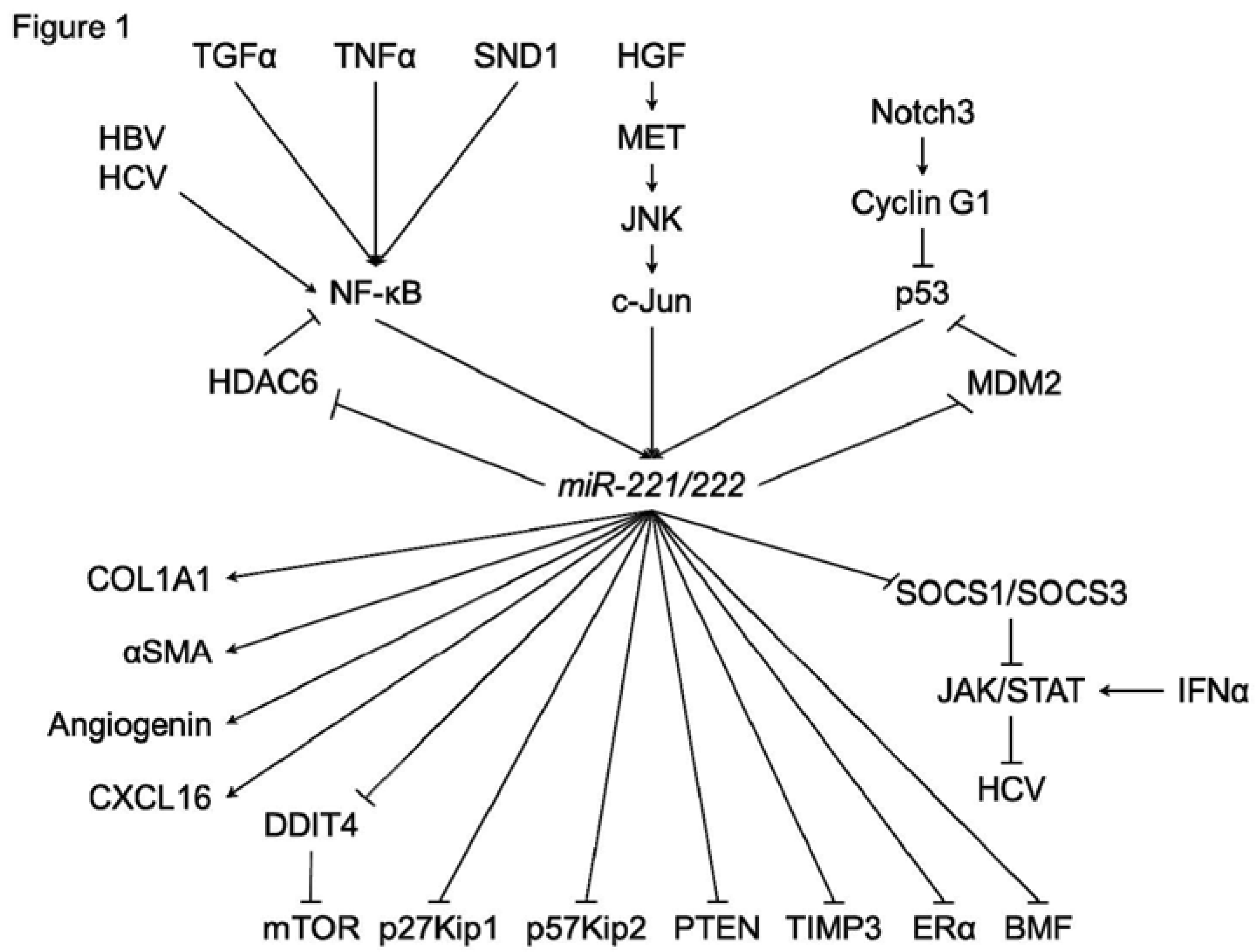
In HCC cells or hepatocyte, various functions of miR-221/222 have been investigated (Figure 1). In addition to p27Kip1 and p57Kip2, several direct targets of miR-221/222 were identified, such as estrogen receptor-alpha (ERα) and a proapoptotic BH3-only protein (BMF) [33,34]. DNA damage-inducible transcript 4 (DDIT4), a modulator of mTOR pathway, was also a direct target of miR-221 [30]. Garofalo et al. showed that miR-221/222, by targeting PTEN and TIMP3 tumor suppressors, induce TRAIL resistance and enhance cellular migration through the activation of the AKT pathway and metallopeptidases. Xu et al. reported that miR-221 was upregulated by HCV infection [35]. In addition, an miR-221 mimic could accentuate the anti-HCV effect of IFN-α in an HCV model, through the inhibition of two members of the suppressor of cytokine signaling (SOCS) family, SOCS1 and SOCS3.
In HCC cells, regulation systems of miR-221/222 have also been investigated (Figure 1). JNK/c-Jun activation and NF-κB nuclear translocation were reported to be essential for the transcription of miR-221/222 [23,36,37]. Hepatitis B virus X protein (HBx) leads to the promotion of cell proliferation and cell growth viability with overexpression of miR-221 [33]. HCV infection could also upregulate the expression of miR-221 in an NF-κB dependent manner [35,38]. Staphylococcal nuclease domain-containing 1 (SND1) is a multifunctional protein that is overexpressed in multiple cancers, including hepatocellular carcinoma (HCC). Santheladur et al. reported that SND1-induced activation of NF-κB resulted in induction of miR-221 and subsequent induction of angiogenic factors Angiogenin and CXCL16 [39].
Figure 1.
A schematic of the regulatory mechanisms of miR-221/222 in hepatocarcinogenesis.
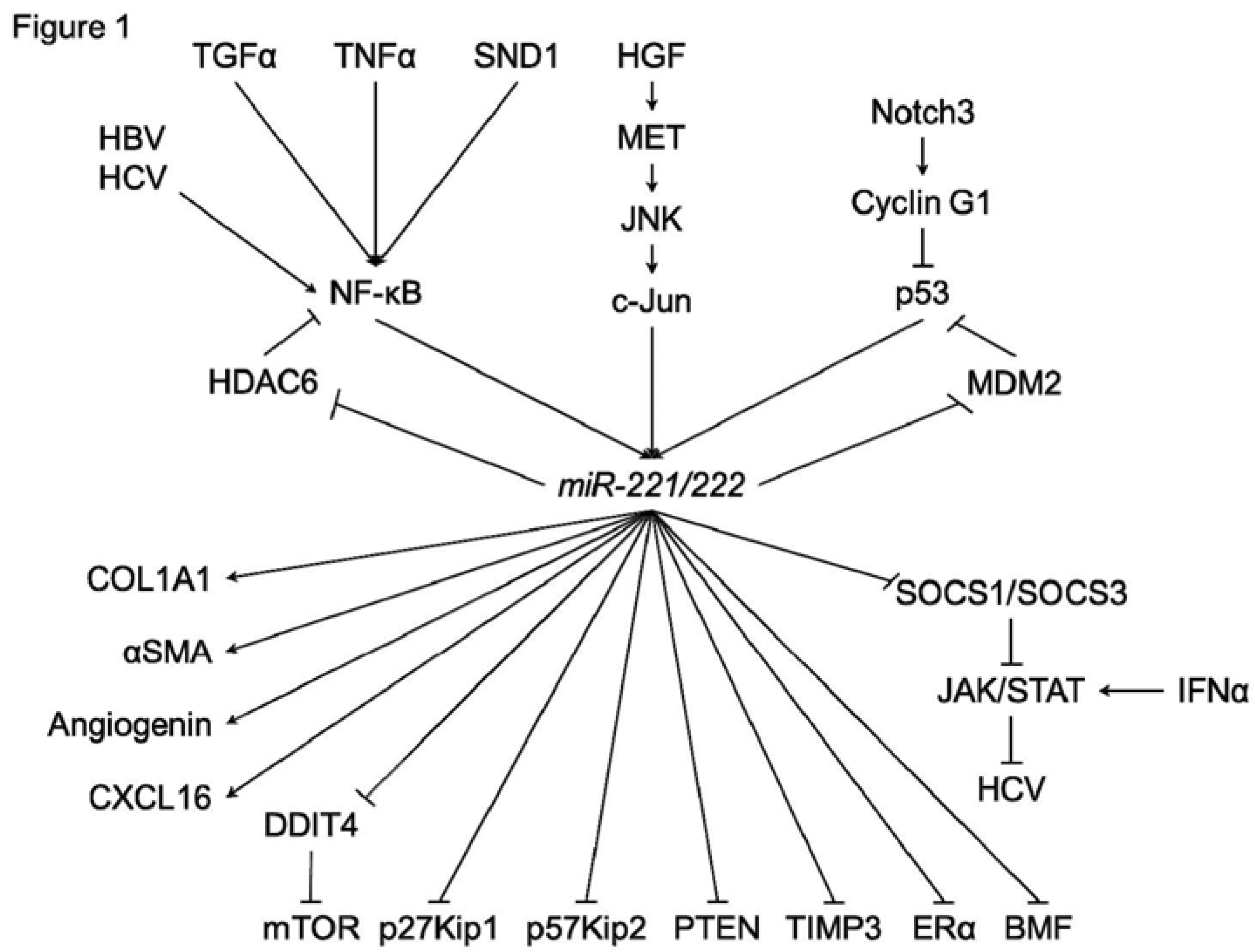
Bae et al. showed that the direct suppression of HDAC6 (histone deacetylase 6) by miR-221 was induced by JNK/c-Jun signaling in liver cancer cells but not in normal hepatic cells [36]. In addition, NF-κB could be activated by miR-221, since HDAC6 suppressed the translocation of NF-κB.
Fornari et al. reported that MDM2 (E3 ubiquitin-protein ligase homolog), a known p53 (TP53) modulator, is identified as a direct target of miR-221 [40]. MiR-221 can activate the p53/mdm2 axis by inhibiting MDM2 and, in turn, p53 activation contributes to miR-221 enhanced expression. Giovannini et al. reported that Notch3 silencing in HCC resulted in p53 upregulation [41]. They found that Notch3 regulated p53 at post-transcriptional level controlling both Cyclin G1 expression and the feed-forward circuit involving p53, miR-221, and MDM2.
7. Pancreatic Cancer
Expression of miR-221/222 is upregulated in pancreatic cancer as compared with normal pancreatic duct epithelial cells or normal pancreas tissues [42,43,44]. Pancreatic cancer patients with high miR-221 expression had a relatively shorter survival compared to those with lower expression [42]. Antisense to miR-221 suppressed the proliferative capacity, increased the amount of apoptosis, and sensitized the effects of gemcitabine in pancreatic cancer cells with concomitant up-regulation of PTEN, p27Kip1, p57Kip2, and PUMA, which are the tumor suppressors and the predicted targets of miR-221 [42,45,46].
Tanaka et al. reported that metformin suppressed the expression of miR-221 in human pancreatic cancer cells, leading to G1-phase arrest via the upregulation of p27Kip1 [47]. In addition, Sarker et al. reported that the treatment of pancreatic cancer cells with isoflavone mixture (G2535), formulated 3,3′-diindolylmethane (BR-DIM), or synthetic curcumin analogue (CDF) could downregulate the expression of miR-221 and consequently upregulate the expression of PTEN, p27Kip1, p57Kip2, and PUMA, leading to the inhibition of proliferation and migration of pancreatic cancer cells [42]. Therefore, these agents combined with conventional chemotherapeutics could be useful in designing novel targeted therapeutic strategy for the treatment of pancreatic cancer.
Matrix metalloproteinases (MMPs) are closely related to cell migration and invasion. Among the MMPs, MMP-2 and MMP-9 have been implicated in human cancer invasion. Xu et al. reported that the tissue inhibitor of metalloproteinase (TIMP)-2 was directly regulated by miR-221/222 [43]. They also showed that miR-221/222 mimic directly inhibited TIMP-2 expression, leading to the upregulation of MMP-2 and MMP-9.
The platelet-derived growth factor (PDGF) signaling pathway has been found to play important roles in the development and progression of human cancers by regulating the processes of cell proliferation, apoptosis, migration, invasion, metastasis, and the acquisition of the epithelial-mesenchymal transition (EMT) phenotype. Su et al. reported that miR-221 expression was activated by PDGF signaling [48]. After the inhibition of miR-221, PDGF did not alter the levels of cell migration, proliferation, and acquisition of the EMT phenotype. These results showed that miR-221 is essential for the PDGF-mediated EMT phenotype, migration, and growth of pancreatic cancer cells. Downregulation of TRPS1 by miR-221 is critical for PDGF-mediated acquisition of the EMT phenotype.
Plasma miR-221 concentration could be a useful biomarker for cancer detection, monitoring tumor dynamics, and predicting malignant outcomes in pancreatic cancer patients [44]. Plasma miR-221 levels were higher in pancreatic cancer patients than in benign pancreatic tumors and controls, and were correlated with distant metastasis. In addition, plasma miR-221 levels were reduced in postoperative samples.
Pancreatic cysts are a group of lesions with heterogeneous malignant potential. MiR-221 concentration in the endoscopically acquired pancreatic cyst fluid samples could be useful for the diagnosis of pancreatic cysts. MiR-221 was expressed at higher levels in malignant cysts compared with benign or premalignant cysts [49].
8. Cholangiocarcinoma
In contrast to the other epithelial cancers, miR-221/222 was downregulated in intrahepatic cholangiocarcinoma tissues, suggesting that miR-221/222 would play onco-suppressive roles [25]. Okamoto et al. reported a relationship between miR-221 expression and the sensitivity of cholangiocarcinoma (CCA) cells to gemcitabine [50]. Microarray analysis was used to determine the miRNA expression profiles of two CCA cell lines, HuCCT1 and HuH28. HuCCT1 cells were more sensitive to gemcitabine than were HuH28 cells, and 18 miRNAs were differentially expressed between HuH28 and HuCCT1. To determine the effect of candidate miRNAs on gemcitabine sensitivity, expression of each candidate miRNA was modified via either transfection of a miRNA mimic or transfection of an anti-oligonucleotide. Among these 18 miRNAs, ectopic overexpression of each of three downregulated miRNAs in HuH28 (miR-29b, miR-205, and miR-221) restored gemcitabine sensitivity to HuH28. Selective siRNA-mediated downregulation of either of two software-predicted targets, PIK3R1 (target of miR-29b and miR-221) or MMP-2 (target of miR-29b), also conferred gemcitabine sensitivity to HuH28.
9. Gastrointestinal Stromal Tumor (GIST)
Gastrointestinal stromal tumors (GISTs) are characterized by high expression of the KIT receptor tyrosine kinase protein, resulting from oncogenic mutations in the extracellular, juxtamembrane, or kinase domains. KIT is known to be directly regulated by miR-221/222, suggesting that miR-221/222 would also play onco-suppressive roles in GISTs [51]. In fact, expression of miR-221/222 is reduced in GISTs compared to control tissue and other sarcomas [51,52,53]. Overexpression of miR-221/222 in GIST cells inhibited cell proliferation, affected cell cycle progression, and induced apoptosis [51,52].
Ihle et al. analyzed expression of miR-221/222 in six KIT exon 9, three KIT exon 11 mutated, and nine wild-type GISTs [52]. MiRNA expression was lower for the wild-type compared to mutated GISTs. Transient transfection of miR-221/222 reduced viability and induced apoptosis by inhibition of KIT expression and its phosphorylation and activation of caspases 3 and 7 in GIST cells. p-AKT, AKT, and BCL2 expression were also reduced after miR-221/222 transfection.
10. Conclusions and Prospects
The discovery of the important role of miRNAs in cancer has opened up a new era of cancer investigations that take into account new and emerging knowledge regarding the RNA signaling systems. The unraveling of miR-221/222 signaling pathways and networks will be key to understanding the role that deregulated miRNA functioning can play in oncogenic or onco-suppressive processes and may be important for defining novel therapeutic molecules.
Recently miRNAs contained in exosomes have been shown to be released and to act as a signal transducer. However, the function of secretory miR-221/222 has never been reported. Previous reports showed that miR-221/222 play various roles not only in cancer but also in vascular smooth muscle cells, vascular endothelial cells, and adipose tissue [54,55]. These suggest that interactions between cancers and blood vessels or adipose tissue would be mediated by secretory miR-221/222. Revealing the inter-organic functions of miRNAs will also help us to better understand cancer biology.
Acknowledgments
This study was funded by a Grant-in-Aid for Young Scientists (B) (26860527, to Juntaro Matsuzaki), a Grant-in-Aid for Scientific Research (B) (25293178, to Hidekazu Suzuki), Grant-in-aid for Challenging Exploratory Research (26670065, to Hidekazu Suzuki) from the Japan Society for the Promotion of Science (JSPS), the Translational Research Network Program from Ministry of Education, Culture, Sports, Science and Technology of Japan (to Hidekazu Suzuki), Princess Takamatsu Cancer Research grants (to Hidekazu Suzuki), a grant from Takeda Science Foundation (to Juntaro Matsuzaki) and the Keio Gijuku Academic Development Fund (to Hidekazu Suzuki).
Author Contributions
Juntaro Matsuzaki wrote and Hidekazu Suzuki supervised and revised the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Galardi, S.; Mercatelli, N.; Giorda, E.; Massalini, S.; Frajese, G.V.; Ciafre, S.A.; Farace, M.G. MiR-221 and miR-222 expression affects the proliferation potential of human prostate carcinoma cell lines by targeting p27kip1. J. Biol. Chem. 2007, 282, 23716–23724. [Google Scholar] [CrossRef] [PubMed]
- Kneitz, B.; Krebs, M.; Kalogirou, C.; Schubert, M.; Joniau, S.; van Poppel, H.; Lerut, E.; Kneitz, S.; Scholz, C.J.; Strobel, P.; et al. Survival in patients with high-risk prostate cancer is predicted by miR-221, which regulates proliferation, apoptosis, and invasion of prostate cancer cells by inhibiting IRF2 and SOCS3. Cancer Res. 2014, 74, 2591–2603. [Google Scholar] [CrossRef] [PubMed]
- Falkenberg, N.; Anastasov, N.; Rappl, K.; Braselmann, H.; Auer, G.; Walch, A.; Huber, M.; Hofig, I.; Schmitt, M.; Hofler, H.; et al. MiR-221/-222 differentiate prognostic groups in advanced breast cancers and influence cell invasion. Br. J. Cancer 2013, 109, 2714–2723. [Google Scholar] [PubMed]
- Quintavalle, C.; Garofalo, M.; Zanca, C.; Romano, G.; Iaboni, M.; del Basso De Caro, M.; Martinez-Montero, J.C.; Incoronato, M.; Nuovo, G.; Croce, C.M.; et al. MiR-221/222 overexpession in human glioblastoma increases invasiveness by targeting the protein phosphate ptpmu. Oncogene 2012, 31, 858–868. [Google Scholar] [PubMed]
- He, H.; Jazdzewski, K.; Li, W.; Liyanarachchi, S.; Nagy, R.; Volinia, S.; Calin, G.A.; Liu, C.G.; Franssila, K.; Suster, S.; et al. The role of microRNA genes in papillary thyroid carcinoma. Proc. Natl. Acad. Sci. USA 2005, 102, 19075–19080. [Google Scholar] [CrossRef] [PubMed]
- Moshiri, F.; Callegari, E.; D’Abundo, L.; Corra, F.; Lupini, L.; Sabbioni, S.; Negrini, M. Inhibiting the oncogenic miR-221 by microRNA sponge: Toward microrna-based therapeutics for hepatocellular carcinoma. Gastroenterol. Hepatol. Bed Bench 2014, 7, 43–54. [Google Scholar] [PubMed]
- Liu, Y.; Cui, H.; Wang, W.; Li, L.; Wang, Z.; Yang, S.; Zhang, X. Construction of circular miRNA sponges targeting miR-21 or miR-221 and demonstration of their excellent anticancer effects on malignant melanoma cells. Int. J. Biochem. Cell Biol. 2013, 45, 2643–2650. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Han, L.; Zhang, A.; Wang, G.; Jia, Z.; Yang, Y.; Yue, X.; Pu, P.; Shen, C.; Kang, C. Adenovirus-mediated shrnas for co-repression of miR-221 and miR-222 expression and function in glioblastoma cells. Oncol. Rep. 2011, 25, 97–105. [Google Scholar] [PubMed]
- Kedde, M.; van Kouwenhove, M.; Zwart, W.; Oude Vrielink, J.A.; Elkon, R.; Agami, R. A pumilio-induced RNA structure switch in p27-3′ UTR controls miR-221 and miR-222 accessibility. Nat. Cell Biol. 2010, 12, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, J.; Suzuki, H.; Tsugawa, H.; Watanabe, M.; Hossain, S.; Arai, E.; Saito, Y.; Sekine, S.; Akaike, T.; Kanai, Y.; et al. Bile acids increase levels of microRNAs 221 and 222, leading to degradation of CDX2 during esophageal carcinogenesis. Gastroenterology 2013, 145, 1300–1311. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Li, G.; Fan, C.; Diao, Y.; Wu, B.; Li, J. Increased expression of microRNA-221 in gastric cancer and its clinical significance. J. Int. Med. Res. 2012, 40, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Chun-Zhi, Z.; Lei, H.; An-Ling, Z.; Yan-Chao, F.; Xiao, Y.; Guang-Xiu, W.; Zhi-Fan, J.; Pei-Yu, P.; Qing-Yu, Z.; Chun-Sheng, K. MicroRNA-221 and microRNA-222 regulate gastric carcinoma cell proliferation and radioresistance by targeting pten. BMC Cancer 2010, 10, 367. [Google Scholar] [PubMed]
- Yau, T.O.; Wu, C.W.; Dong, Y.; Tang, C.M.; Ng, S.S.; Chan, F.K.; Sung, J.J.; Yu, J. MicroRNA-221 and microRNA-18a identification in stool as potential biomarkers for the non-invasive diagnosis of colorectal carcinoma. Br. J. Cancer 2014, 111, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Tao, K.; Yang, J.; Guo, Z.; Hu, Y.; Sheng, H.; Gao, H.; Yu, H. Prognostic value of miR-221–3p, miR-342–3p and miR-491–5p expression in colon cancer. Am. J. Transl. Res. 2014, 6, 391–401. [Google Scholar] [PubMed]
- Sun, K.; Wang, W.; Zeng, J.J.; Wu, C.T.; Lei, S.T.; Li, G.X. MicroRNA-221 inhibits cdkn1c/p57 expression in human colorectal carcinoma. Acta Pharmacol. Sin. 2011, 32, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Sun, X.; Wang, M.; Hou, Y.; Zhan, Y.; Jiang, Y.; Liu, Z.; Cao, X.; Chen, P.; Chen, X.; et al. A microRNA-221- and 222-mediated feedback loop maintains constitutive activation of nfkappab and STAT3 in colorectal cancer cells. Gastroenterology 2014, 147, 847–859.e11. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Luo, M. MicroRNA-221 promotes colorectal cancer cell invasion and metastasis by targeting RECK. FEBS Lett 2014, 588, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Tsunoda, T.; Takashima, Y.; Yoshida, Y.; Doi, K.; Tanaka, Y.; Fujimoto, T.; Machida, T.; Ota, T.; Koyanagi, M.; Kuroki, M.; et al. Oncogenic KRAS regulates miR-200c and miR-221/222 in a 3D-specific manner in colorectal cancer cells. Anticancer Res. 2011, 31, 2453–2459. [Google Scholar] [PubMed]
- Cai, K.; Shen, F.; Cui, J.H.; Yu, Y.; Pan, H.Q. Expression of miR-221 in colon cancer correlates with prognosis. Int. J. Clin. Exp. Med. 2015, 8, 2794–2798. [Google Scholar] [PubMed]
- Xue, Q.; Sun, K.; Deng, H.J.; Lei, S.T.; Dong, J.Q.; Li, G.X. Anti-mirna-221 sensitizes human colorectal carcinoma cells to radiation by upregulating pten. World J. Gastroenterol. 2013, 19, 9307–9317. [Google Scholar] [CrossRef] [PubMed]
- Pu, X.X.; Huang, G.L.; Guo, H.Q.; Guo, C.C.; Li, H.; Ye, S.; Ling, S.; Jiang, L.; Tian, Y.; Lin, T.Y. Circulating miR-221 directly amplified from plasma is a potential diagnostic and prognostic marker of colorectal cancer and is correlated with p53 expression. J. Gastroenterol. Hepatol. 2010, 25, 1674–1680. [Google Scholar] [CrossRef] [PubMed]
- He, X.X.; Guo, A.Y.; Xu, C.R.; Chang, Y.; Xiang, G.Y.; Gong, J.; Dan, Z.L.; Tian, D.A.; Liao, J.Z.; Lin, J.S. Bioinformatics analysis identifies miR-221 as a core regulator in hepatocellular carcinoma and its silencing suppresses tumor properties. Oncol. Rep. 2014, 32, 1200–1210. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Enomoto, M.; Fujii, H.; Sekiya, Y.; Yoshizato, K.; Ikeda, K.; Kawada, N. MicroRNA-221/222 upregulation indicates the activation of stellate cells and the progression of liver fibrosis. Gut 2012, 61, 1600–1609. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.O.; Chun, S.M.; Han, E.H.; Choi, J.; Jang, S.J.; Koh, S.A.; Hwang, S.; Yu, E. Deregulated expression of microRNA-221 with the potential for prognostic biomarkers in surgically resected hepatocellular carcinoma. Hum. Pathol. 2011, 42, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Karakatsanis, A.; Papaconstantinou, I.; Gazouli, M.; Lyberopoulou, A.; Polymeneas, G.; Voros, D. Expression of microRNAs, miR-21, miR-31, miR-122, miR-145, miR-146a, miR-200c, miR-221, miR-222, and miR-223 in patients with hepatocellular carcinoma or intrahepatic cholangiocarcinoma and its prognostic significance. Mol. Carcinog 2013, 52, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Wang, Q.; Chen, J.; Huang, X.; Chen, X.; Cao, L.; Tan, H.; Li, W.; Zhang, L.; Bi, J.; et al. Clinical significance of miR-221 and its inverse correlation with p27kip(1) in hepatocellular carcinoma. Mol. Biol. Rep. 2011, 38, 3029–3035. [Google Scholar] [CrossRef] [PubMed]
- Rong, M.; Chen, G.; Dang, Y. Increased miR-221 expression in hepatocellular carcinoma tissues and its role in enhancing cell growth and inhibiting apoptosis in vitro. BMC Cancer 2013, 13, 21. [Google Scholar] [CrossRef] [PubMed]
- Fornari, F.; Gramantieri, L.; Ferracin, M.; Veronese, A.; Sabbioni, S.; Calin, G.A.; Grazi, G.L.; Giovannini, C.; Croce, C.M.; Bolondi, L.; et al. MiR-221 controls CDKN1C/p57 and CDKN1B/p27 expression in human hepatocellular carcinoma. Oncogene 2008, 27, 5651–5661. [Google Scholar] [PubMed]
- Li, J.; Wang, Y.; Yu, W.; Chen, J.; Luo, J. Expression of serum miR-221 in human hepatocellular carcinoma and its prognostic significance. Biochem. Biophys. Res. Commun. 2011, 406, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Pineau, P.; Volinia, S.; McJunkin, K.; Marchio, A.; Battiston, C.; Terris, B.; Mazzaferro, V.; Lowe, S.W.; Croce, C.M.; Dejean, A. MiR-221 overexpression contributes to liver tumorigenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Callegari, E.; Elamin, B.K.; Giannone, F.; Milazzo, M.; Altavilla, G.; Fornari, F.; Giacomelli, L.; D’Abundo, L.; Ferracin, M.; Bassi, C.; et al. Liver tumorigenicity promoted by microRNA-221 in a mouse transgenic model. Hepatology 2012, 56, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Park, J.K.; Kogure, T.; Nuovo, G.J.; Jiang, J.; He, L.; Kim, J.H.; Phelps, M.A.; Papenfuss, T.L.; Croce, C.M.; Patel, T.; et al. MiR-221 silencing blocks hepatocellular carcinoma and promotes survival. Cancer Res. 2011, 71, 7608–7616. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Tang, Y.S.; Huang, S.F.; Ai, J.G.; Wang, H.X.; Zhang, L.P. HBx protein-induced upregulation of microRNA-221 promotes aberrant proliferation in HBV-related hepatocellular carcinoma by targeting estrogen receptor-alpha. Oncol. Rep. 2015, 33, 792–798. [Google Scholar] [PubMed]
- Gramantieri, L.; Fornari, F.; Ferracin, M.; Veronese, A.; Sabbioni, S.; Calin, G.A.; Grazi, G.L.; Croce, C.M.; Bolondi, L.; Negrini, M. MicroRNA-221 targets Bmf in hepatocellular carcinoma and correlates with tumor multifocality. Clin. Cancer Res. 2009, 15, 5073–5081. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Yang, F.; Ding, C.L.; Wang, J.; Zhao, P.; Wang, W.; Ren, H. MiR-221 accentuates IFNs anti-HCV effect by downregulating SOCS1 and SOCS3. Virology 2014, 462–463, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.J.; Jung, K.H.; Eun, J.W.; Shen, Q.; Kim, H.S.; Park, S.J.; Shin, W.C.; Yang, H.D.; Park, W.S.; Lee, J.Y.; et al. MicroRNA-221 governs tumor suppressor HDAC6 to potentiate malignant progression of liver cancer. J. Hepatol. 2015, 63, 408–419. [Google Scholar] [PubMed]
- Garofalo, M.; di Leva, G.; Romano, G.; Nuovo, G.; Suh, S.S.; Ngankeu, A.; Taccioli, C.; Pichiorri, F.; Alder, H.; Secchiero, P.; et al. MiR-221&222 regulate TRAIL resistance and enhance tumorigenicity through PTEN and TIMP3 downregulation. Cancer Cell 2009, 16, 498–509. [Google Scholar] [PubMed]
- Ding, C.L.; Xu, G.; Ren, H.; Zhao, L.J.; Zhao, P.; Qi, Z.T.; Wang, W. HCV infection induces the upregulation of miR-221 in nf-kappab dependent manner. Virus Res. 2015, 196, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Santhekadur, P.K.; Das, S.K.; Gredler, R.; Chen, D.; Srivastava, J.; Robertson, C.; Baldwin, A.S., Jr.; Fisher, P.B.; Sarkar, D. Multifunction protein staphylococcal nuclease domain containing 1 (SND1) promotes tumor angiogenesis in human hepatocellular carcinoma through novel pathway that involves nuclear factor kappab and miR-221. J. Biol. Chem. 2012, 287, 13952–13958. [Google Scholar] [CrossRef] [PubMed]
- Fornari, F.; Milazzo, M.; Galassi, M.; Callegari, E.; Veronese, A.; Miyaaki, H.; Sabbioni, S.; Mantovani, V.; Marasco, E.; Chieco, P.; et al. P53/mdm2 feedback loop sustains miR-221 expression and dictates the response to anticancer treatments in hepatocellular carcinoma. Mol. Cancer Res. 2014, 12, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Giovannini, C.; Minguzzi, M.; Baglioni, M.; Fornari, F.; Giannone, F.; Ravaioli, M.; Cescon, M.; Chieco, P.; Bolondi, L.; Gramantieri, L. Suppression of p53 by Notch3 is mediated by Cyclin G1 and sustained by MDM2 and miR-221 axis in hepatocellular carcinoma. Oncotarget 2014, 5, 10607–10620. [Google Scholar] [PubMed]
- Sarkar, S.; Dubaybo, H.; Ali, S.; Goncalves, P.; Kollepara, S.L.; Sethi, S.; Philip, P.A.; Li, Y. Down-regulation of miR-221 inhibits proliferation of pancreatic cancer cells through up-regulation of PTEN, p27(kip1), p57(kip2), and PUMA. Am. J. Cancer Res. 2013, 3, 465–477. [Google Scholar] [PubMed]
- Xu, Q.; Li, P.; Chen, X.; Zong, L.; Jiang, Z.; Nan, L.; Lei, J.; Duan, W.; Zhang, D.; Li, X.; et al. MiR-221/222 induces pancreatic cancer progression through the regulation of matrix metalloproteinases. Oncotarget 2015, 6, 14153–14164. [Google Scholar] [PubMed]
- Kawaguchi, T.; Komatsu, S.; Ichikawa, D.; Morimura, R.; Tsujiura, M.; Konishi, H.; Takeshita, H.; Nagata, H.; Arita, T.; Hirajima, S.; et al. Clinical impact of circulating miR-221 in plasma of patients with pancreatic cancer. Br. J. Cancer 2013, 108, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Park, J.K.; Lee, E.J.; Esau, C.; Schmittgen, T.D. Antisense inhibition of microRNA-21 or -221 arrests cell cycle, induces apoptosis, and sensitizes the effects of gemcitabine in pancreatic adenocarcinoma. Pancreas 2009, 38, e190–e199. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Alder, H.; Khiyami, A.; Leahy, P.; Croce, C.M.; Haldar, S. MicroRNA-375 and microRNA-221: Potential noncoding RNAs associated with antiproliferative activity of benzyl isothiocyanate in pancreatic cancer. Genes Cancer 2011, 2, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, R.; Tomosugi, M.; Horinaka, M.; Sowa, Y.; Sakai, T. Metformin causes G1-phase arrest via down-regulation of miR-221 and enhances trail sensitivity through DR5 up-regulation in pancreatic cancer cells. PLoS ONE 2015, 10, e0125779. [Google Scholar] [CrossRef] [PubMed]
- Su, A.; He, S.; Tian, B.; Hu, W.; Zhang, Z. MicroRNA-221 mediates the effects of pdgf-bb on migration, proliferation, and the epithelial-mesenchymal transition in pancreatic cancer cells. PLoS ONE 2013, 8, e71309. [Google Scholar] [CrossRef] [PubMed]
- Farrell, J.J.; Toste, P.; Wu, N.; Li, L.; Wong, J.; Malkhassian, D.; Tran, L.M.; Wu, X.; Li, X.; Dawson, D.; et al. Endoscopically acquired pancreatic cyst fluid microRNA 21 and 221 are associated with invasive cancer. Am. J. Gastroenterol. 2013, 108, 1352–1359. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Miyoshi, K.; Murawaki, Y. MiR-29b, miR-205 and miR-221 enhance chemosensitivity to gemcitabine in HuH28 human cholangiocarcinoma cells. PLoS ONE 2013, 8, e77623. [Google Scholar] [CrossRef] [PubMed]
- Gits, C.M.; van Kuijk, P.F.; Jonkers, M.B.; Boersma, A.W.; van Ijcken, W.F.; Wozniak, A.; Sciot, R.; Rutkowski, P.; Schoffski, P.; Taguchi, T.; et al. MiR-17-92 and miR-221/222 cluster members target KIT and ETV1 in human gastrointestinal stromal tumours. Br. J. Cancer 2013, 109, 1625–1635. [Google Scholar] [PubMed]
- Ihle, M.A.; Trautmann, M.; Kuenstlinger, H.; Huss, S.; Heydt, C.; Fassunke, J.; Wardelmann, E.; Bauer, S.; Schildhaus, H.U.; Buettner, R.; et al. MiRNA-221 and miRNA-222 induce apoptosis via the kit/akt signalling pathway in gastrointestinal stromal tumours. Mol. Oncol. 2015. [Google Scholar] [CrossRef]
- Koelz, M.; Lense, J.; Wrba, F.; Scheffler, M.; Dienes, H.P.; Odenthal, M. Down-regulation of miR-221 and miR-222 correlates with pronounced Kit expression in gastrointestinal stromal tumors. Int. J. Oncol. 2011, 38, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Meerson, A.; Traurig, M.; Ossowski, V.; Fleming, J.M.; Mullins, M.; Baier, L.J. Human adipose microRNA-221 is upregulated in obesity and affects fat metabolism downstream of leptin and TNF-alpha. Diabetologia 2013, 56, 1971–1979. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cheng, Y.; Yang, J.; Xu, L.; Zhang, C. Cell-specific effects of miR-221/222 in vessels: Molecular mechanism and therapeutic application. J. Mol. Cell. Cardiol. 2012, 52, 245–255. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Matsuzaki, J.; Suzuki, H. Role of MicroRNAs-221/222 in Digestive Systems. J. Clin. Med. 2015, 4, 1566-1577. https://doi.org/10.3390/jcm4081566
AMA Style
Matsuzaki J, Suzuki H. Role of MicroRNAs-221/222 in Digestive Systems. Journal of Clinical Medicine. 2015; 4(8):1566-1577. https://doi.org/10.3390/jcm4081566
Chicago/Turabian StyleMatsuzaki, Juntaro, and Hidekazu Suzuki. 2015. "Role of MicroRNAs-221/222 in Digestive Systems" Journal of Clinical Medicine 4, no. 8: 1566-1577. https://doi.org/10.3390/jcm4081566