Genetic Predisposition and Disease Expression of Bipolar Disorder Reflected in Shape Changes of the Anterior Limbic Network

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Imaging Data Acquisition and Preprocessing
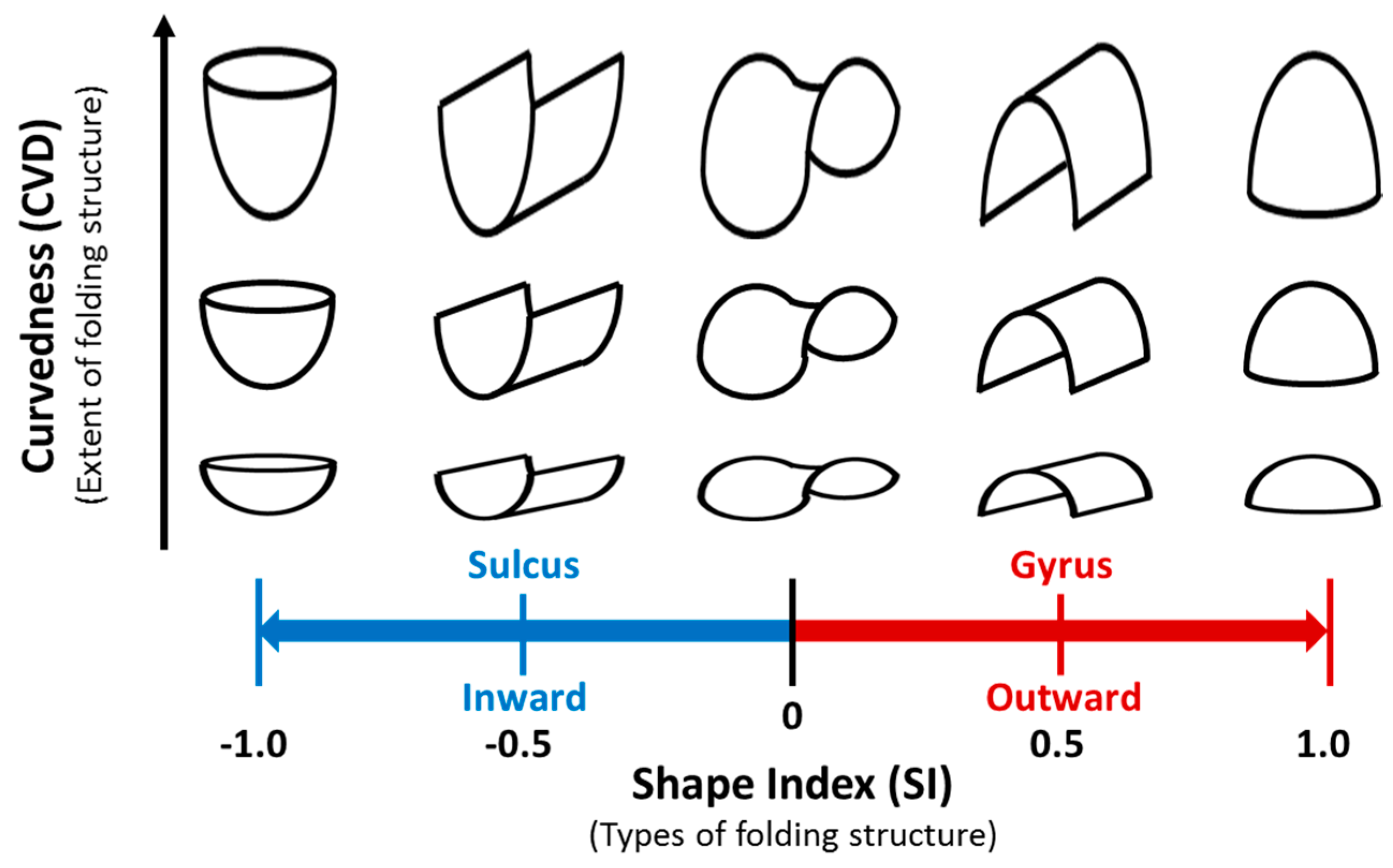
2.3. Cortical Surface Extraction and Shape Analysis
2.4. Statistical Analysis for Group Comparison
3. Results
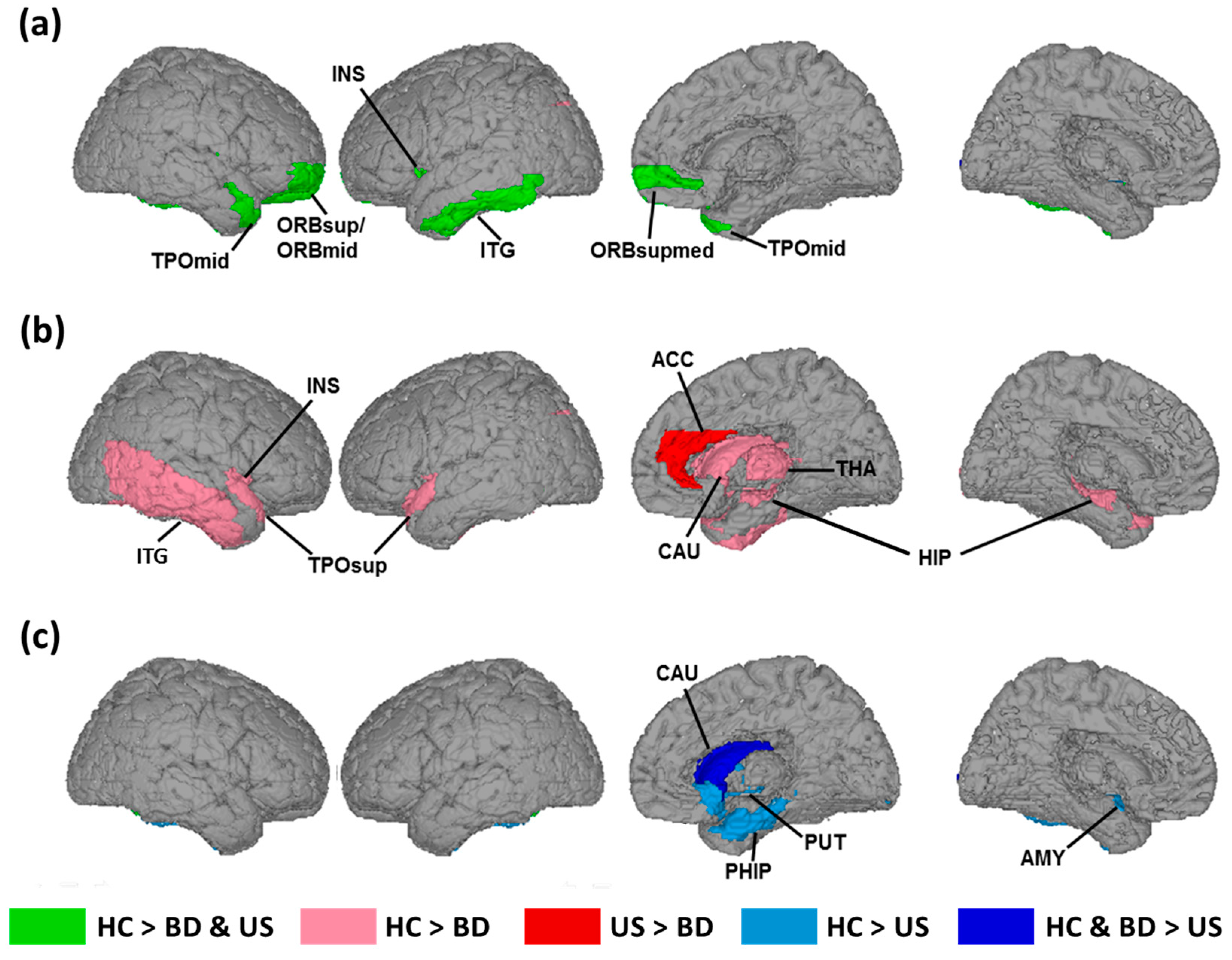
3.1. Changes in the Cortical Folding associated with Hereditary Predisposition to Bipolar Disorder
3.2. Changes in Cortical Folding associated with Disease Expression of BD
3.3. Changes in Cortical Folding Associated with the Absence of Bipolar Disorder Diagnosis
4. Discussion
4.1. Neurobiological Basis of Changes in Cortical Folding Structure
4.2. Changes in Cortical Folding Associated with Hereditary Predisposition to Bipolar Disorder
4.3. Changes in Cortical Folding Associated with Disease Expression of Bipolar Disorder
4.4. Cortical Folding Structures Associated with an Absence of Diagnosis for Unaffected Siblings
4.5. Further Considerations
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Goes, F.S.; Pirooznia, M.; Parla, J.S.; Kramer, M.; Ghiban, E.; Mavruk, S.; Chen, Y.-C.; Monson, E.T.; Willour, V.L.; Karchin, R. Exome sequencing of familial bipolar disorder. JAMA Psychiatry 2016, 73, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Smoller, J.W.; Finn, C.T. Family, twin, and adoption studies of bipolar disorder. Am. J. Med. Genet. Part C 2003, 123, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, F.; Jamison, K. Manic-Depressive Illness. Bipolar Disorders and Recurrent Depression, 2nd ed.; Oxford University Press: New York, NY, USA, 2007. [Google Scholar]
- Heun, R.; Maier, W. The distinction of bipolar II disorder from bipolar I and recurrent unipolar depression: Results of a controlled family study. Acta Psychiatr. Scand. 1993, 87, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Cardno, A.G.; Marshall, E.J.; Coid, B.; Macdonald, A.M.; Ribchester, T.R.; Davies, N.J.; Venturi, P.; Jones, L.A.; Lewis, S.W.; Sham, P.C. Heritability estimates for psychotic disorders: The Maudsley twin psychosis series. Arch. Gen. Psychiatry 1999, 56, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Martinez, D.; Millischer, V.; Michelon, L.; Camilo, C.; Brentani, H.; Esteban, N.; Krieger, J.; Pereira, A.; Lavebratt, C.; Schalling, M. High Heritability of Telomere Length In Families With Bipolar Disorder. Eur. Neuropsychopharmacol. 2017, 27, S387. [Google Scholar] [CrossRef]
- Stahl, E.A.; Breen, G.; Forstner, A.J.; McQuillin, A.; Ripke, S.; Trubetskoy, V.; Mattheisen, M.; Wang, Y.; Coleman, J.R.; Gaspar, H.A. Genome-wide association study identifies 30 loci associated with bipolar disorder. Nat. Genet. 2019, 51, 793. [Google Scholar] [CrossRef] [PubMed]
- Shih, R.A.; Belmonte, P.L.; Zandi, P.P. A review of the evidence from family, twin and adoption studies for a genetic contribution to adult psychiatric disorders. Int. Rev. Psychiatry 2004, 16, 260–283. [Google Scholar] [CrossRef] [PubMed]
- Hibar, D.; Westlye, L.T.; Doan, N.T.; Jahanshad, N.; Cheung, J.; Ching, C.R.; Versace, A.; Bilderbeck, A.; Uhlmann, A.; Mwangi, B. Cortical abnormalities in bipolar disorder: An MRI analysis of 6503 individuals from the ENIGMA Bipolar Disorder Working Group. Mol. Psychiatry 2018, 23, 932. [Google Scholar] [CrossRef]
- Arnone, D.; Cavanagh, J.; Gerber, D.; Lawrie, S.M.; Ebmeier, K.P.; McIntosh, A.M. Magnetic resonance imaging studies in bipolar disorder and schizophrenia: Meta-analysis. Br. J. Psychiatry 2009, 195, 194–201. [Google Scholar] [CrossRef]
- Kempton, M.J.; Geddes, J.R.; Ettinger, U.; Williams, S.R.; Grasby, P.M. Meta-analysis, database, and meta-regression of 98 structural imaging studies in bipolar disorder. Arch. Gen. Psychiatry 2008, 65, 1017–1032. [Google Scholar] [CrossRef]
- Ellison-Wright, I.; Bullmore, E. Anatomy of bipolar disorder and schizophrenia: A meta-analysis. Schizophr. Res. 2010, 117, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Frangou, S. Neuroimaging Markers of Risk, Disease Expression, and Resilience to Bipolar Disorder. Curr. Psychiatry Rep. 2019, 21, 52. [Google Scholar] [CrossRef] [PubMed]
- McDonald, C.; Bullmore, E.T.; Sham, P.C.; Chitnis, X.; Wickham, H.; Bramon, E.; Murray, R.M. Association of genetic risks for schizophrenia and bipolar disorder with specific and generic brain structural endophenotypes. Arch. Gen. Psychiatry 2004, 61, 974. [Google Scholar] [CrossRef] [PubMed]
- Hajek, T.; Cullis, J.; Novak, T.; Kopecek, M.; Blagdon, R.; Propper, L.; Stopkova, P.; Duffy, A.; Hoschl, C.; Uher, R. Brain structural signature of familial predisposition for bipolar disorder: Replicable evidence for involvement of the right inferior frontal gyrus. Biol. Psychiatry 2013, 73, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Kempton, M.J.; Haldane, M.; Jogia, J.; Grasby, P.M.; Collier, D.; Frangou, S. Dissociable Brain Structural Changes Associated with Predisposition, Resilience, and Disease Expression in Bipolar Disorder. J. Neurosci. 2009, 29, 10863–10868. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Kopecek, M.; Nicoletti, M.; Hatch, J.; Watanabe, Y.; Nery, F.; Zunta-Soares, G.; Soares, J. New structural brain imaging endophenotype in bipolar disorder. Mol. Psychiatry 2011, 17, 412–420. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, A.M.; Job, D.E.; Moorhead, T.W.J.; Harrison, L.K.; Forrester, K.; Lawrie, S.M.; Johnstone, E.C. Voxel-based morphometry of patients with schizophrenia or bipolar disorder and their unaffected relatives. Biol. Psychiatry 2004, 56, 544–552. [Google Scholar] [CrossRef]
- Noga, J.T.; Vladar, K.; Torrey, E.F. A volumetric magnetic resonance imaging study of monozygotic twins discordant for bipolar disorder. Psychiatry Res. 2001, 106, 25–34. [Google Scholar] [CrossRef]
- Chaddock, C.A.; Barker, G.J.; Marshall, N.; Schulze, K.; Hall, M.H.; Fern, A.; Walshe, M.; Bramon, E.; Chitnis, X.A.; Murray, R.; et al. White matter microstructural impairments and genetic liability to familial bipolar I disorder. Br. J. Psychiatry 2009, 194, 527–534. [Google Scholar] [CrossRef] [Green Version]
- Sprooten, E.; Sussmann, J.E.; Clugston, A.; Peel, A.; McKirdy, J.; Moorhead, T.W.J.; Anderson, S.; Shand, A.J.; Giles, S.; Bastin, M.E.; et al. White Matter Integrity in Individuals at High Genetic Risk of Bipolar Disorder. Biol. Psychiatry 2011, 70, 350–356. [Google Scholar] [CrossRef]
- Zilles, K.; Palomero-Gallagher, N.; Amunts, K. Development of cortical folding during evolution and ontogeny. Trends Neurosci. 2013, 36, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Kriegstein, A.; Noctor, S.; Martínez-Cerdeño, V. Patterns of neural stem and progenitor cell division may underlie evolutionary cortical expansion. Nat. Rev. Neurosci. 2006, 7, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Mota, B.; Herculano-Houzel, S. How the cortex gets its folds: An inside-out, connectivity-driven model for the scaling of mammalian cortical folding. Front. Neuroanat. 2012, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Van Essen, D.C. A tension-based theory of morphogenesis and compact wiring in the central nervous system. Nature 1997, 385, 313–318. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, A.; Moorhead, T.; McKirdy, J.; Hall, J.; Sussmann, J.; Stanfield, A.; Harris, J.; Johnstone, E.; Lawrie, S. Prefrontal gyral folding and its cognitive correlates in bipolar disorder and schizophrenia. Acta Psychiatr. Scand. 2009, 119, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Penttilä, J.; Cachia, A.; Martinot, J.L.; Ringuenet, D.; Wessa, M.; Houenou, J.; Galinowski, A.; Bellivier, F.; Gallarda, T.; Duchesnay, E. Cortical folding difference between patients with early-onset and patients with intermediate-onset bipolar disorder. Bipolar Disord. 2009, 11, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.P.V.; Bücker, J.; Burke, K.P.; Czepielewski, L.S.; Santos, B.T.; Fijtman, A.; Passos, I.C.; Kunz, M.; Bonnin, C.D.M.; Vieta, E. Cognitive performance and psychosocial functioning in patients with bipolar disorder, unaffected siblings, and healthy controls. Braz. J. Psychiatry 2016, 38, 275–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivedi, J.K.; Goel, D.; Dhyani, M.; Sharma, S.; Singh, A.P.; Sinha, P.K.; Tandon, R. Neurocognition in first-degree healthy relatives (siblings) of bipolar affective disorder patients. Psychiatry Clin. Neurosci. 2008, 62, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.; Jain, S.; Janardhan Reddy, Y.; Kumar, K.J.; Kandavel, T. Impairment of verbal learning and memory and executive function in unaffected siblings of probands with bipolar disorder. Bipolar Disord. 2010, 12, 647–656. [Google Scholar] [CrossRef]
- Bora, E.; Yucel, M.; Pantelis, C. Cognitive endophenotypes of bipolar disorder: A meta-analysis of neuropsychological deficits in euthymic patients and their first-degree relatives. J. Affect. Disord. 2009, 113, 1–20. [Google Scholar] [CrossRef]
- Arts, B.; Jabben, N.; Krabbendam, L.; Van Os, J. Meta-analyses of cognitive functioning in euthymic bipolar patients and their first-degree relatives. Psychol. Med. 2008, 38, 771–786. [Google Scholar] [CrossRef] [PubMed]
- Adler, C.M.; DelBello, M.P.; Strakowski, S.M. Brain network dysfunction in bipolar disorder. CNS Spectr. 2006, 11, 312. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.-H.; Chen, H.-Y.; Hung, C.-I.; Guo, W.-Y.; Wu, Y.-T. Shape and curvedness analysis of brain morphology using human fetal magnetic resonance images in utero. Brain Struct. Funct. 2012, 218, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.-F.; Wang, P.-S.; Lao, Y.-L.; Wu, H.-M.; Soong, B.-W.; Wu, Y.-T. Medullo-ponto-cerebellar white matter degeneration altered brain network organization and cortical morphology in multiple system atrophy. Brain Struct. Funct. 2014, 219, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.-L.; Wang, P.-S.; Lu, C.-F.; Hung, C.-I.; Li, C.-T.; Lin, C.-P.; Hsieh, J.-C.; Su, T.-P.; Wu, Y.-T. Cortical Shape and Curvedness Analysis of Structural Deficits in Remitting and Non-Remitting Depression. PLoS ONE 2013, 8, e68625. [Google Scholar] [CrossRef]
- Awate, S.P.; Yushkevich, P.A.; Song, Z.; Licht, D.J.; Gee, J.C. Cerebral cortical folding analysis with multivariate modeling and testing: Studies on gender differences and neonatal development. Neuroimage 2010, 53, 450–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segonne, F.; Dale, A.; Busa, E.; Glessner, M.; Salat, D.; Hahn, H.; Fischl, B. A hybrid approach to the skull stripping problem in MRI. Neuroimage 2004, 22, 1060–1075. [Google Scholar] [CrossRef]
- Sled, J.G.; Zijdenbos, A.P.; Evans, A.C. A nonparametric method for automatic correction of intensity nonuniformity in MRI data. IEEE Trans. Med. Imaging 1998, 17, 87–97. [Google Scholar] [CrossRef]
- Li, K.; Guo, L.; Li, G.; Nie, J.; Faraco, C.; Cui, G.; Zhao, Q.; Miller, L.S.; Liu, T. Gyral folding pattern analysis via surface profiling. Neuroimage 2010, 52, 1202–1214. [Google Scholar] [CrossRef] [Green Version]
- Koenderink, J.J.; van Doorn, A.J. Surface shape and curvature scales. Image Vis. Comput. 1992, 10, 557–564. [Google Scholar] [CrossRef]
- Thirion, J.-P.; Gourdon, A. Computing the differential characteristics of isointensity surfaces. Comput. Vis. Image Underst. 1995, 61, 190–202. [Google Scholar] [CrossRef]
- Rajmohan, V.; Mohandas, E. The limbic system. Indian J. Psychiatry 2007, 49, 132. [Google Scholar] [CrossRef] [PubMed]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyer, B.; Joliot, M. Automated Anatomical Labeling of Activations in SPM Using a Macroscopic Anatomical Parcellation of the MNI MRI Single-Subject Brain. NeuroImage 2002, 15, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, S.; Cachia, A.; Hozer, F.; McDonald, C.; Emsell, L.; Cannon, D.M.; Wessa, M.; Linke, J.; Versace, A.; Hamdani, N. Neurodevelopmental subtypes of bipolar disorder are related to cortical folding patterns: An international multicenter study. Bipolar Disord. 2018, 20, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Penttilä, J.; Paillère-Martinot, M.-L.; Martinot, J.-L.; Ringuenet, D.; Wessa, M.; Houenou, J.; Gallarda, T.; Bellivier, F.; Galinowski, A.; Bruguière, P. Cortical folding in patients with bipolar disorder or unipolar depression. J. Psychiatry Neurosci. 2009, 34, 127. [Google Scholar] [PubMed]
- Rogers, J.; Kochunov, P.; Zilles, K.; Shelledy, W.; Lancaster, J.; Thompson, P.; Duggirala, R.; Blangero, J.; Fox, P.T.; Glahn, D.C. On the genetic architecture of cortical folding and brain volume in primates. Neuroimage 2010, 53, 1103–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, J.; Benders, M.; Cachia, A.; Lazeyras, F.; Leuchter, R.H.-V.; Sizonenko, S.; Borradori-Tolsa, C.; Mangin, J.; Hüppi, P.S. Mapping the early cortical folding process in the preterm newborn brain. Cereb. Cortex 2008, 18, 1444–1454. [Google Scholar] [CrossRef] [PubMed]
- Frazier, J.A.; Breeze, J.L.; Papadimitriou, G.; Kennedy, D.N.; Hodge, S.M.; Moore, C.M.; Howard, J.D.; Rohan, M.P.; Caviness, V.S.; Makris, N. White matter abnormalities in children with and at risk for bipolar disorder. Bipolar Disord. 2007, 9, 799–809. [Google Scholar] [CrossRef]
- Peper, J.S.; Brouwer, R.M.; Boomsma, D.I.; Kahn, R.S.; Pol, H.; Hilleke, E. Genetic influences on human brain structure: A review of brain imaging studies in twins. Hum. Brain Mapp. 2007, 28, 464–473. [Google Scholar] [CrossRef]
- Wright, I.C.; Sham, P.; Murray, R.M.; Weinberger, D.R.; Bullmore, E.T. Genetic Contributions to Regional Variability in Human Brain Structure: Methods and Preliminary Results. Neuroimage 2002, 17, 256–271. [Google Scholar] [CrossRef]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews-Hanna, J.R.; Reidler, J.S.; Sepulcre, J.; Poulin, R.; Buckner, R.L. Functional-Anatomic Fractionation of the Brain’s Default Network. Neuron 2010, 65, 550–562. [Google Scholar] [CrossRef] [PubMed]
- Meda, S.A.; Gill, A.; Stevens, M.C.; Lorenzoni, R.P.; Glahn, D.C.; Calhoun, V.D.; Sweeney, J.A.; Tamminga, C.A.; Keshavan, M.S.; Thaker, G.; et al. Differences in Resting-State Functional Magnetic Resonance Imaging Functional Network Connectivity Between Schizophrenia and Psychotic Bipolar Probands and Their Unaffected First-Degree Relatives. Biol. Psychiatry 2012, 71, 881–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, L.Q.; Nomi, J.S.; Hébert-Seropian, B.; Ghaziri, J.; Boucher, O. Structure and function of the human insula. J. Clin. Neurophysiol. 2017, 34, 300. [Google Scholar] [CrossRef] [PubMed]
- Uddin, L.Q. Salience processing and insular cortical function and dysfunction. Nat. Rev. Neurosci. 2015, 16, 55. [Google Scholar] [CrossRef] [PubMed]
- Kelley, A.E.; Baldo, B.A.; Pratt, W.E. A proposed hypothalamic–thalamic–striatal axis for the integration of energy balance, arousal, and food reward. J. Comp. Neurol. 2005, 493, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Olson, I.R.; Plotzker, A.; Ezzyat, Y. The Enigmatic temporal pole: A review of findings on social and emotional processing. Brain 2007, 130, 1718–1731. [Google Scholar] [CrossRef]
- Teng, S.; Lu, C.-F.; Wang, P.-S.; Li, C.-T.; Tu, P.-C.; Hung, C.-I.; Su, T.-P.; Wu, Y.-T. Altered Resting-State Functional Connectivity of Striatal-Thalamic Circuit in Bipolar Disorder. PLoS ONE 2014, 9, e96422. [Google Scholar] [CrossRef]
- Carter, C.S.; Braver, T.S.; Barch, D.M.; Botvinick, M.M.; Noll, D.; Cohen, J.D. Anterior cingulate cortex, error detection, and the online monitoring of performance. Science 1998, 280, 747–749. [Google Scholar] [CrossRef]
- Bush, G.; Luu, P.; Posner, M.I. Cognitive and emotional influences in anterior cingulate cortex. Trends Cogn. Sci. 2000, 4, 215–222. [Google Scholar] [CrossRef]
- Fornito, A.; Yücel, M.; Wood, S.; Stuart, G.W.; Buchanan, J.-A.; Proffitt, T.; Anderson, V.; Velakoulis, D.; Pantelis, C. Individual differences in anterior cingulate/paracingulate morphology are related to executive functions in healthy males. Cereb. Cortex 2004, 14, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Tissier, C.; Linzarini, A.; Allaire-Duquette, G.; Mevel, K.; Poirel, N.; Dollfus, S.; Etard, O.; Orliac, F.; Peyrin, C.; Charron, S. Sulcal Polymorphisms of the IFC and ACC Contribute to Inhibitory Control Variability in Children and Adults. eNeuro 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Strakowski, S.; Delbello, M.; Adler, C. The functional neuroanatomy of bipolar disorder: A review of neuroimaging findings. Mol. Psychiatry 2005, 10, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, M.E.; DelBello, M.P.; Getz, G.E.; Shear, P.K.; Strakowski, S.M. Anterior cingulate subregion volumes and executive function in bipolar disorder. Bipolar Disord. 2006, 8, 281–288. [Google Scholar] [CrossRef]
- Drevets, W.C.; Savitz, J.; Trimble, M. The subgenual anterior cingulate cortex in mood disorders. CNS Spectr. 2008, 13, 663. [Google Scholar] [CrossRef]
- Fornito, A.; Whittle, S.; Wood, S.J.; Velakoulis, D.; Pantelis, C.; Yücel, M. The influence of sulcal variability on morphometry of the human anterior cingulate and paracingulate cortex. Neuroimage 2006, 33, 843–854. [Google Scholar] [CrossRef]
- Sanches, M.; Amorim, E.; Mwangi, B.; Zunta-Soares, G.B.; Soares, J.C. Smaller left anterior cingulate cortex in non-bipolar relatives of patients with bipolar disorder. Braz. J. Psychiatry 2019, 41, 254–256. [Google Scholar] [CrossRef] [Green Version]
- Surguladze, S.A.; Marshall, N.; Schulze, K.; Hall, M.-H.; Walshe, M.; Bramon, E.; Phillips, M.L.; Murray, R.M.; McDonald, C. Exaggerated neural response to emotional faces in patients with bipolar disorder and their first-degree relatives. Neuroimage 2010, 53, 58–64. [Google Scholar] [CrossRef]
- LaBar, K.S.; Cabeza, R. Cognitive neuroscience of emotional memory. Nat. Rev. Neurosci. 2006, 7, 54–64. [Google Scholar] [CrossRef]
- Phillips, M.L.; Drevets, W.C.; Rauch, S.L.; Lane, R. Neurobiology of emotion perception I: The neural basis of normal emotion perception. Biol. Psychiatry 2003, 54, 504–514. [Google Scholar] [CrossRef]
- Brotman, M.; Guyer, A.; Lawson, E.; Horsey, S.; Rich, B.; Dickstein, D.; Pine, D.; Leibenluft, E. Facial emotion labeling deficits in children and adolescents at risk for bipolar disorder. Am. J. Psychiatry 2008, 165, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Boccardi, M.; Almici, M.; Bresciani, L.; Caroli, A.; Bonetti, M.; Monchieri, S.; Gennarelli, M.; Frisoni, G.B. Clinical and medial temporal features in a family with mood disorders. Neurosci. Lett. 2010, 468, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Ladouceur, C.D.; Almeida, J.R.C.; Birmaher, B.; Axelson, D.A.; Nau, S.; Kalas, C.; Monk, K.; Kupfer, D.J.; Phillips, M.L. Subcortical Gray Matter Volume Abnormalities in Healthy Bipolar Offspring: Potential Neuroanatomical Risk Marker for Bipolar Disorder? J. Am. Acad. Child Adolesc. Psychiatry 2008, 47, 532–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellivier, F.; Golmard, J.-L.; Rietschel, M.; Schulze, T.G.; Malafosse, A.; Preisig, M.; McKeon, P.; Mynett-Johnson, L.; Henry, C.; Leboyer, M. Age at onset in bipolar I affective disorder: Further evidence for three subgroups. Am. J. Psychiatry 2003, 160, 999–1001. [Google Scholar] [CrossRef] [PubMed]
- Yucel, K.; Taylor, V.H.; McKinnon, M.C.; MacDonald, K.; Alda, M.; Young, L.T.; MacQueen, G.M. Bilateral hippocampal volume increase in patients with bipolar disorder and short-term lithium treatment. Neuropsychopharmacology 2008, 33, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Monkul, E.S.; Matsuo, K.; Nicoletti, M.A.; Dierschke, N.; Hatch, J.P.; Dalwani, M.; Brambilla, P.; Caetano, S.; Sassi, R.B.; Mallinger, A.G.; et al. Prefrontal gray matter increases in healthy individuals after lithium treatment: A voxel-based morphometry study. Neurosci. Lett. 2007, 429, 7–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Bipolar I Patients (BD) | Unaffected Siblings (US) | Healthy Controls (HC) | p-Value | |
|---|---|---|---|---|
| Number of subjects | 17 | 17 | 22 | |
| Gender (male/female) | 11/6 | 8/9 | 13/9 | 0.566 a |
| Age (years) | 42.9 ± 10.6 | 42.6 ± 11.2 | 42.1 ± 10.6 | 0.970 b |
| HAMD | 3.3 ± 2.8 | 0.7 ± 1.0 | 1.1 ± 1.4 | <0.001 b |
| YMARS | 2.4 ± 2.5 | 0.6 ± 1.8 | 0.2 ± 0.7 | 0.001 b |
| Duration of illness (years) | 17.3 ± 9.9 | -- | -- | |
| Onset age (years) | 25.5 ± 11.5 | -- | -- | |
| Past manic episodes (times) | 5.8 ± 6.6 | -- | -- | |
| Past depressive episodes (times) | 4.1 ± 3.4 | -- | -- | |
| Number of subjects who had psychosis (%) | 11 (64.7%) | -- | -- | |
| GM volumes (cm3) | 577.6 ± 56.4 | 562.3 ± 44.6 | 569.0 ± 53.8 | 0.692 b |
| WM volumes (cm3) | 509.5 ± 56.3 | 497.8 ± 54.7 | 506.3 ± 56.8 | 0.819 b |
| GM + WM volumes (cm3) | 1087.0 ± 108.4 | 1060.0 ± 85.6 | 1075.2 ± 106.1 | 0.738 b |
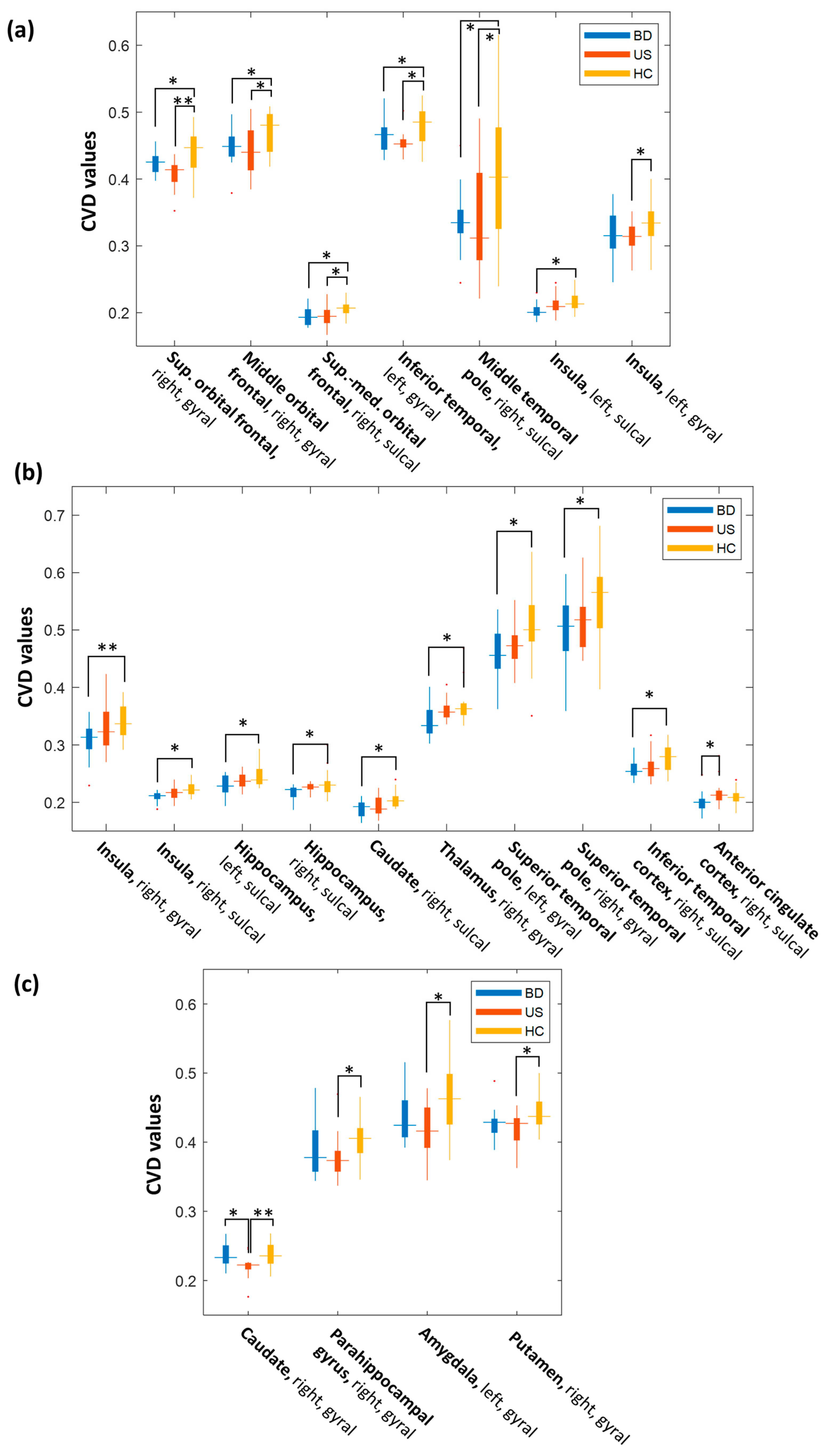
| Cortical Region | Surface Category | CVD in BD | CVD in US | CVD in HC | Pairwise p-Value |
|---|---|---|---|---|---|
| BD < HC and US < HC | |||||
| Superior orbital frontal cortex, right | Gyral | 0.424 ± 0.016 | 0.408 ± 0.022 | 0.444 ± 0.031 | BD vs. HC 0.048 * US vs. HC < 0.001 * BD vs. US 0.228 |
| Middle orbital frontal cortex, right | Gyral | 0.448 ± 0.028 | 0.442 ± 0.037 | 0.473 ± 0.029 | BD vs. HC 0.047 * US vs. HC 0.011 * BD vs. US 1.000 |
| Superior-medial orbital frontal cortex, right | Sulcal | 0.194 ± 0.014 | 0.195 ± 0.014 | 0.206 ± 0.011 | BD vs. HC 0.011 * US vs. HC 0.022 * BD vs. US 1.000 |
| Inferior temporal cortex, left | Gyral | 0.463 ± 0.024 | 0.454 ± 0.016 | 0.482 ± 0.026 | BD vs. HC 0.033 * US vs. HC 0.001 * BD vs. US 0.843 |
| Middle temporal pole, right | Sulcal | 0.336 ± 0.047 | 0.340 ± 0.084 | 0.405 ± 0.010 | BD vs. HC 0.033 * US vs. HC 0.048 * BD vs. US 1.000 |
| Insula, left | Sulcal | 0.204 ± 0.011 | 0.212 ± 0.014 | 0.215 ± 0.013 | BD vs. HC 0.032 * US vs. HC 1.000 BD vs. US 0.197 |
| Insula, left | Gyral | 0.319 ± 0.032 | 0.312 ± 0.024 | 0.339 ± 0.034 | BD vs. HC 0.152 US vs. HC 0.026 * BD vs. US 1.000 |
| Cortical Region | Surface Category | CVD in BD | CVD in US | CVD in HC | Pairwise p-Value |
|---|---|---|---|---|---|
| BD < HC | |||||
| Insula, right | Gyral | 0.308 ± 0.034 | 0.327 ± 0.041 | 0.340 ± 0.029 | BD vs. HC 0.020 * US vs. HC 0.774 BD vs. US 0.355 |
| Insula, right | Sulcal | 0.210 ± 0.009 | 0.216 ± 0.013 | 0.223± 0.012 | BD vs. HC 0.003 * US vs. HC 0.200 BD vs. US 0.374 |
| Hippocampus, left | Sulcal | 0.228 ± 0.019 | 0.238 ± 0.012 | 0.244 ± 0.016 | BD vs. HC 0.008 * US vs. HC 0.657 BD vs. US 0.243 |
| Hippocampus, right | Sulcal | 0.217 ± 0.014 | 0.226 ± 0.008 | 0.233 ± 0.022 | BD vs. HC 0.011 * US vs. HC 0.574 BD vs. US 0.334 |
| Caudate, right | Sulcal | 0.189 ± 0.015 | 0.194 ± 0.017 | 0.205 ± 0.014 | BD vs. HC 0.006 * US vs. HC 0.068 BD vs. US 1.000 |
| Thalamus, right | Gyral | 0.340 ± 0.028 | 0.361 ± 0.018 | 0.367 ± 0.030 | BD vs. HC 0.009 * US vs. HC 1.000 BD vs. US 0.083 |
| Superior temporal pole, left | Gyral | 0.461 ± 0.048 | 0.484 ± 0.039 | 0.506 ± 0.070 | BD vs. HC 0.045 * US vs. HC 0.251 BD vs. US 1.000 |
| Superior temporal pole, right | Gyral | 0.500 ± 0.062 | 0.515 ± 0.050 | 0.554 ± 0.066 | BD vs. HC 0.022 * US vs. HC 0.152 BD vs. US 1.000 |
| Inferior temporal cortex, right | Sulcal | 0.258 ± 0.019 | 0.263 ± 0.023 | 0.279 ± 0.026 | BD vs. HC 0.018 * US vs. HC 0.104 BD vs. US 1.000 |
| BD < US | |||||
| Anterior cingulate cortex, right | Sulcal | 0.201 ± 0.017 | 0.216 ± 0.022 | 0.210 ± 0.013 | BD vs. HC 0.262 US vs. HC 0.883 BD vs. US 0.033 * |
| Cortical Region | Surface Category | CVD in BD | CVD in US | CVD in HC | Pairwise p-Value |
|---|---|---|---|---|---|
| US < HC and US < BD | |||||
| Caudate, right | Gyral | 0.237 ± 0.016 | 0.220 ± 0.016 | 0.238 ± 0.017 | BD vs. HC 1..000 US vs. HC 0.004 * BD vs. US 0.014 * |
| US < HC | |||||
| Parahippocampal gyrus, right | Gyral | 0.386 ± 0.036 | 0.377 ± 0.032 | 0.405 ± 0.031 | BD vs. HC 0.249 US vs. HC 0.036 * BD vs. US 1.000 |
| Amygdala, left | Gyral | 0.436 ± 0.036 | 0.421 ± 0.037 | 0.461 ± 0.052 | BD vs. HC 0.225 US vs. HC 0.016 * BD vs. US 0.918 |
| Putamen, right | Gyral | 0.426 ± 0.023 | 0.419 ± 0.025 | 0.442 ± 0.024 | BD vs. HC 0.136 US vs. HC 0.013 * BD vs. US 1.000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, C.-F.; Wu, Y.-T.; Teng, S.; Wang, P.-S.; Tu, P.-C.; Su, T.-P.; Jao, C.-W.; Li, C.-T. Genetic Predisposition and Disease Expression of Bipolar Disorder Reflected in Shape Changes of the Anterior Limbic Network. Brain Sci. 2019, 9, 240. https://doi.org/10.3390/brainsci9090240
Lu C-F, Wu Y-T, Teng S, Wang P-S, Tu P-C, Su T-P, Jao C-W, Li C-T. Genetic Predisposition and Disease Expression of Bipolar Disorder Reflected in Shape Changes of the Anterior Limbic Network. Brain Sciences. 2019; 9(9):240. https://doi.org/10.3390/brainsci9090240
Chicago/Turabian StyleLu, Chia-Feng, Yu-Te Wu, Shin Teng, Po-Shan Wang, Pei-Chi Tu, Tung-Ping Su, Chi-Wen Jao, and Cheng-Ta Li. 2019. "Genetic Predisposition and Disease Expression of Bipolar Disorder Reflected in Shape Changes of the Anterior Limbic Network" Brain Sciences 9, no. 9: 240. https://doi.org/10.3390/brainsci9090240