Timing-Dependent Protection of Swimming Exercise against d-Galactose-Induced Aging-Like Impairments in Spatial Learning/Memory in Rats




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
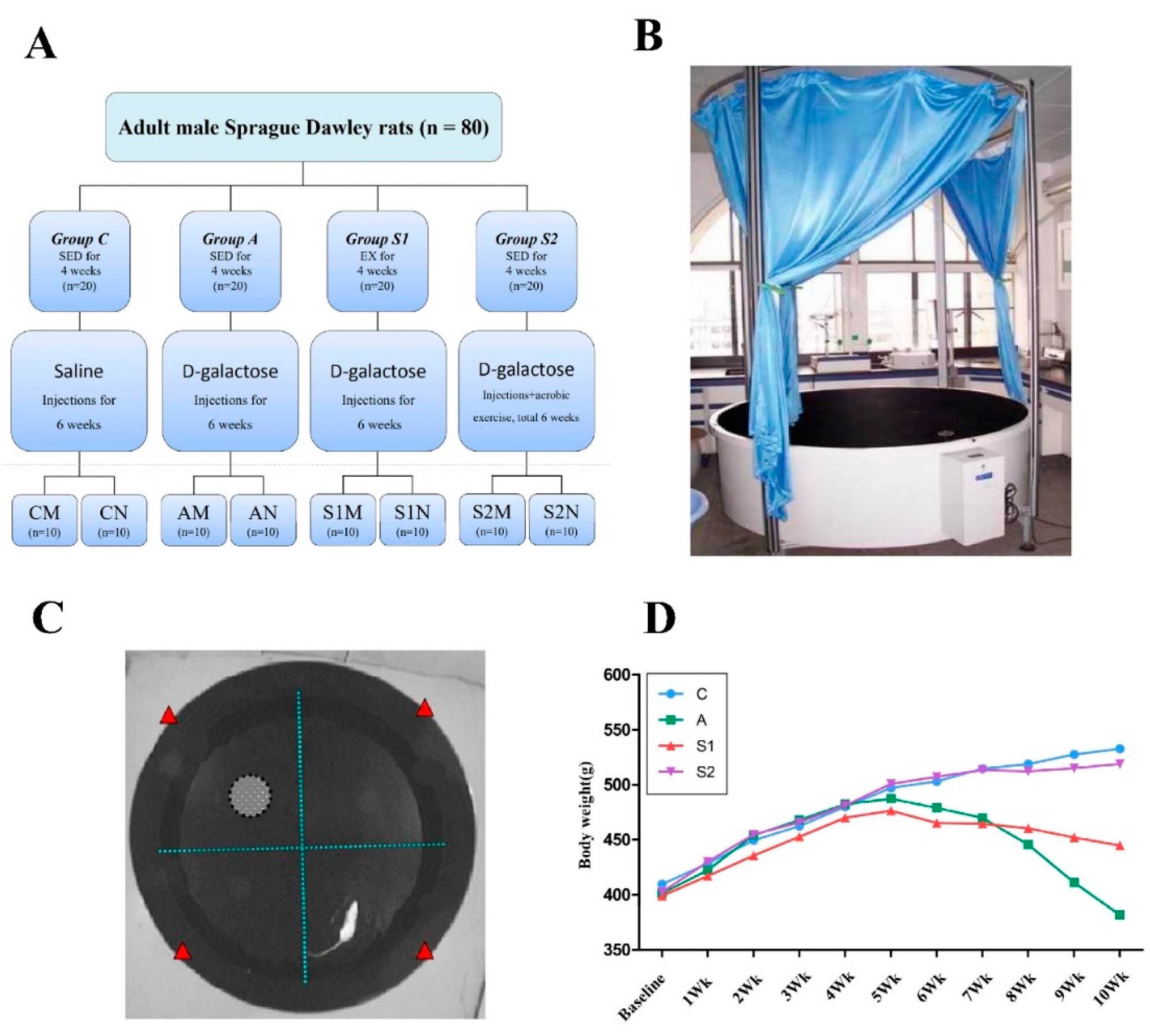
2.1. Animals and Rat Model of Drug (d-Galactose)-Induced Aging
2.2. Exercise Protocols and Subsequent Tests and Tissue Collections
2.3. Morris Water Maze Behavioral Test
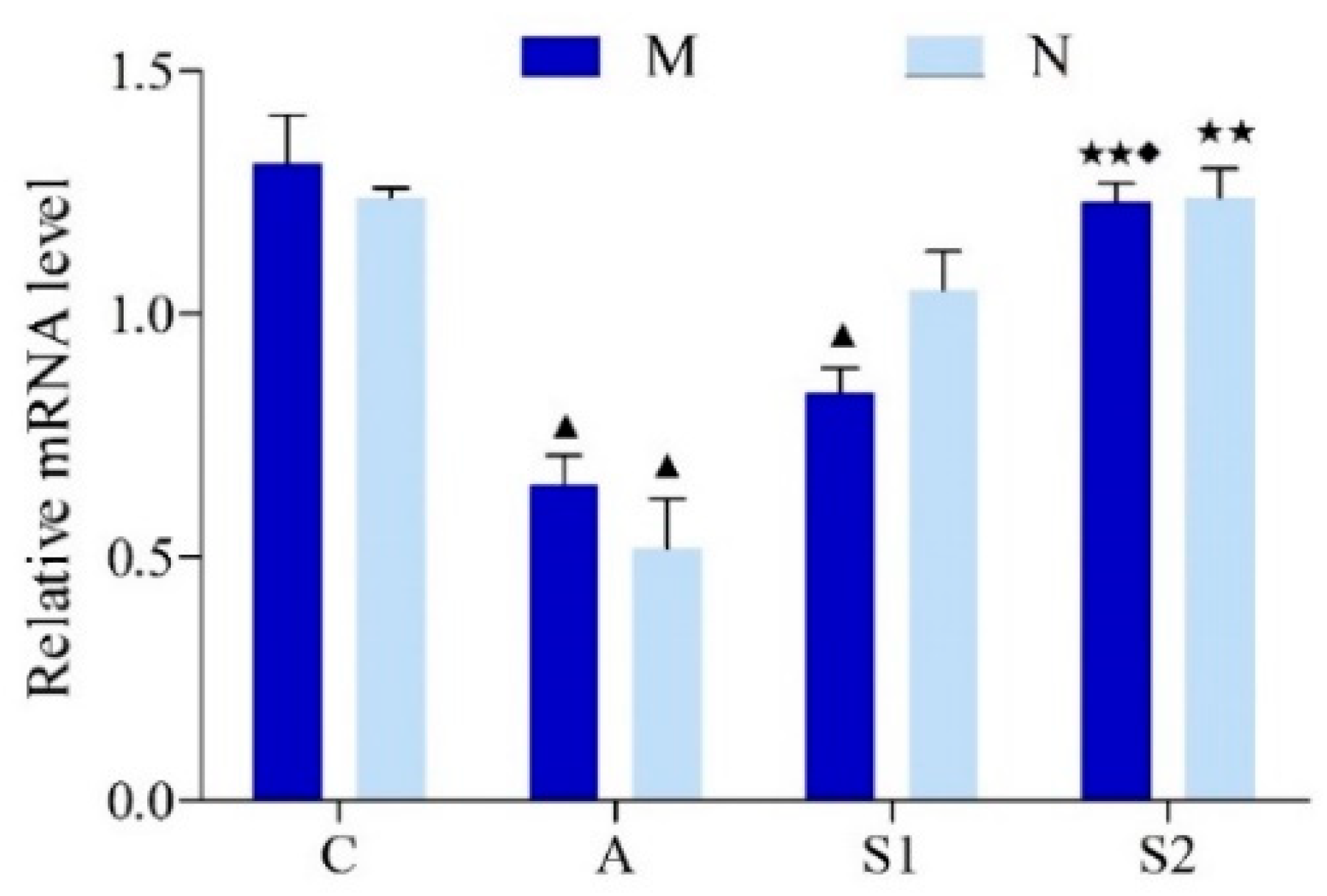
2.4. CREB mRNA Expressions in Hippocampus
2.5. Hippocampal CREB Protein Content and β-Actin as Protein Loading Control
2.6. Epigenetic Markers
2.7. Data Analysis
3. Results
3.1. General Health Status of Animals
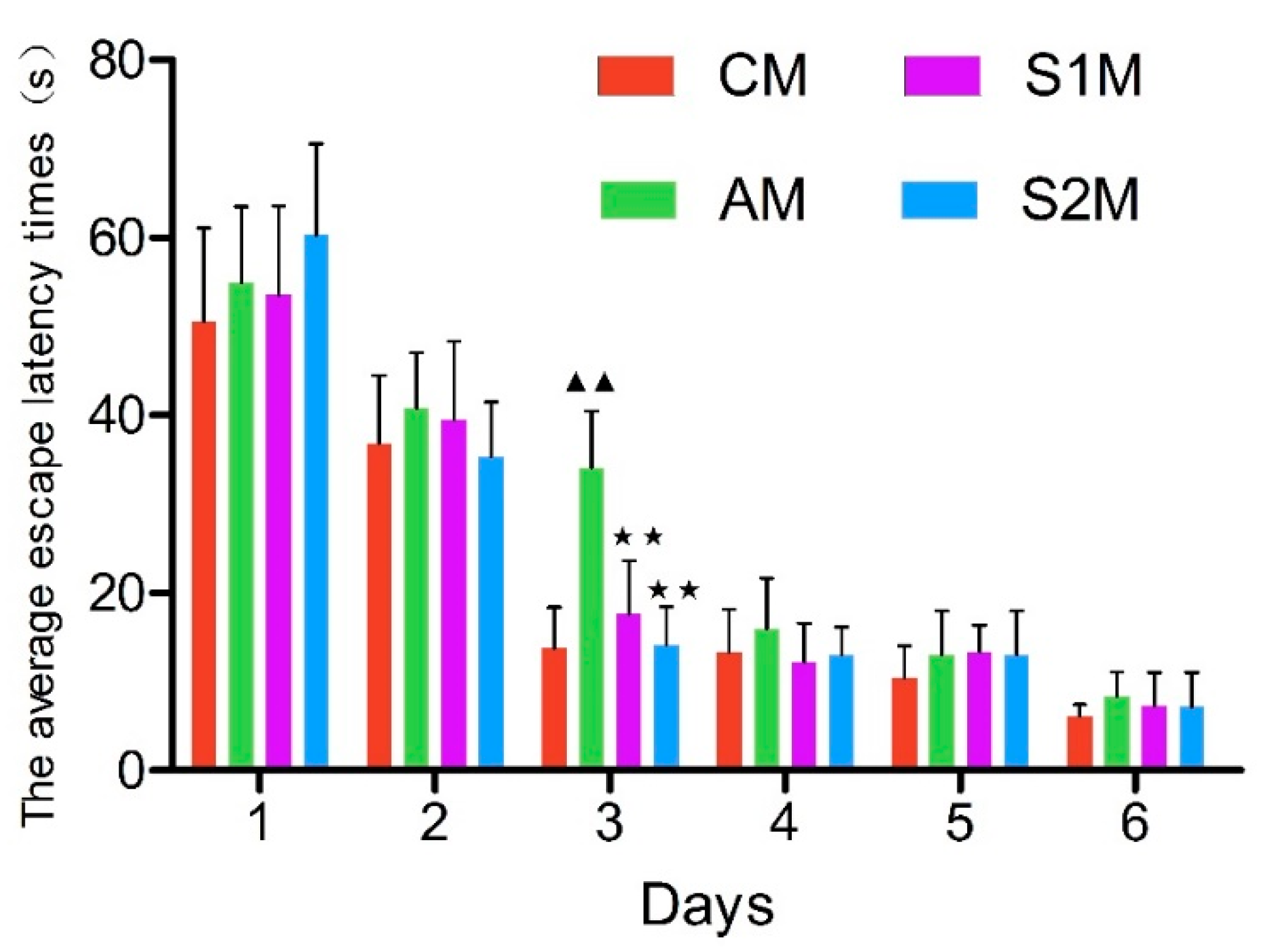
3.2. Morris Water Maze Navigation Test
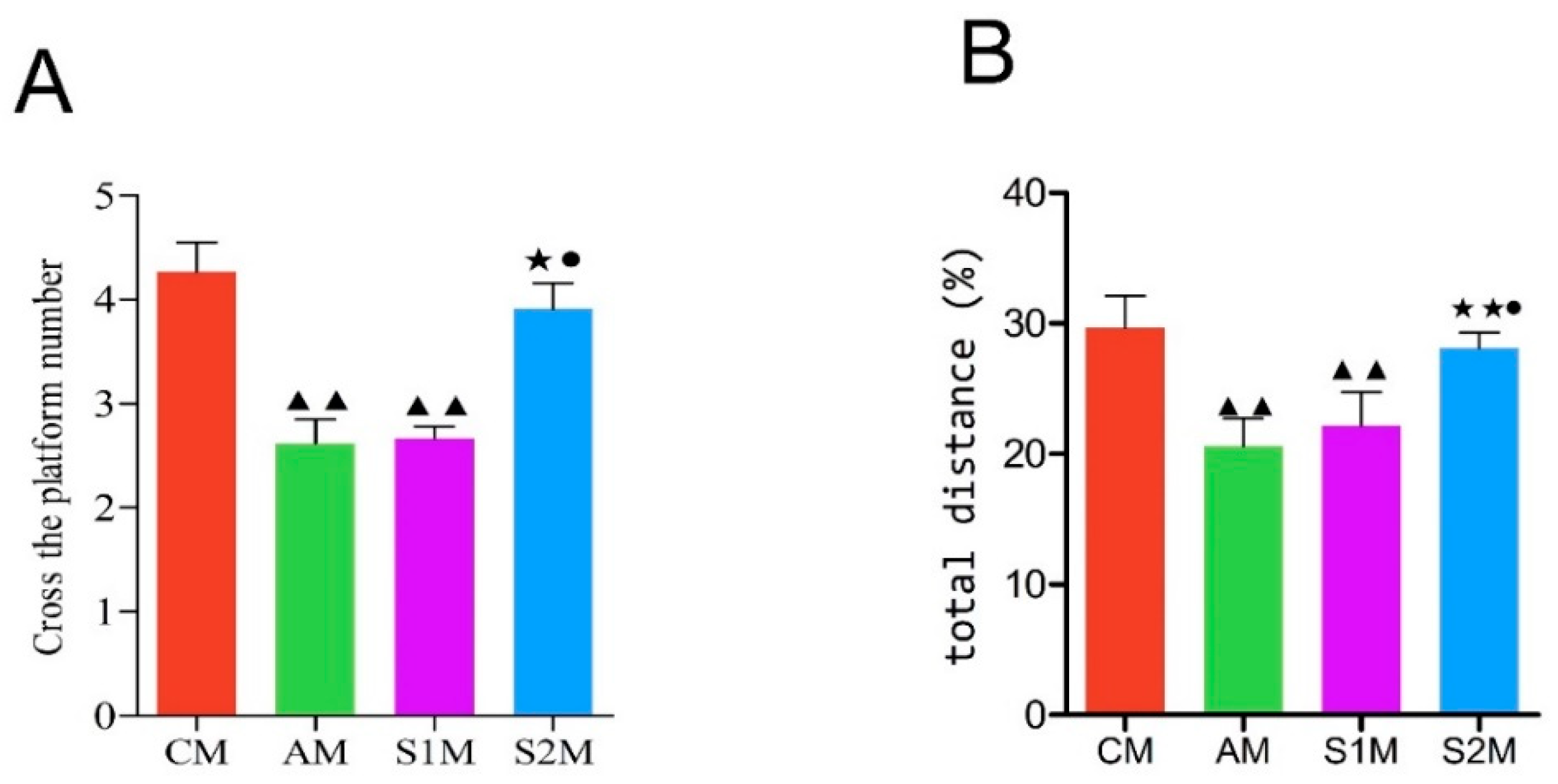
3.3. Spatial Exploration
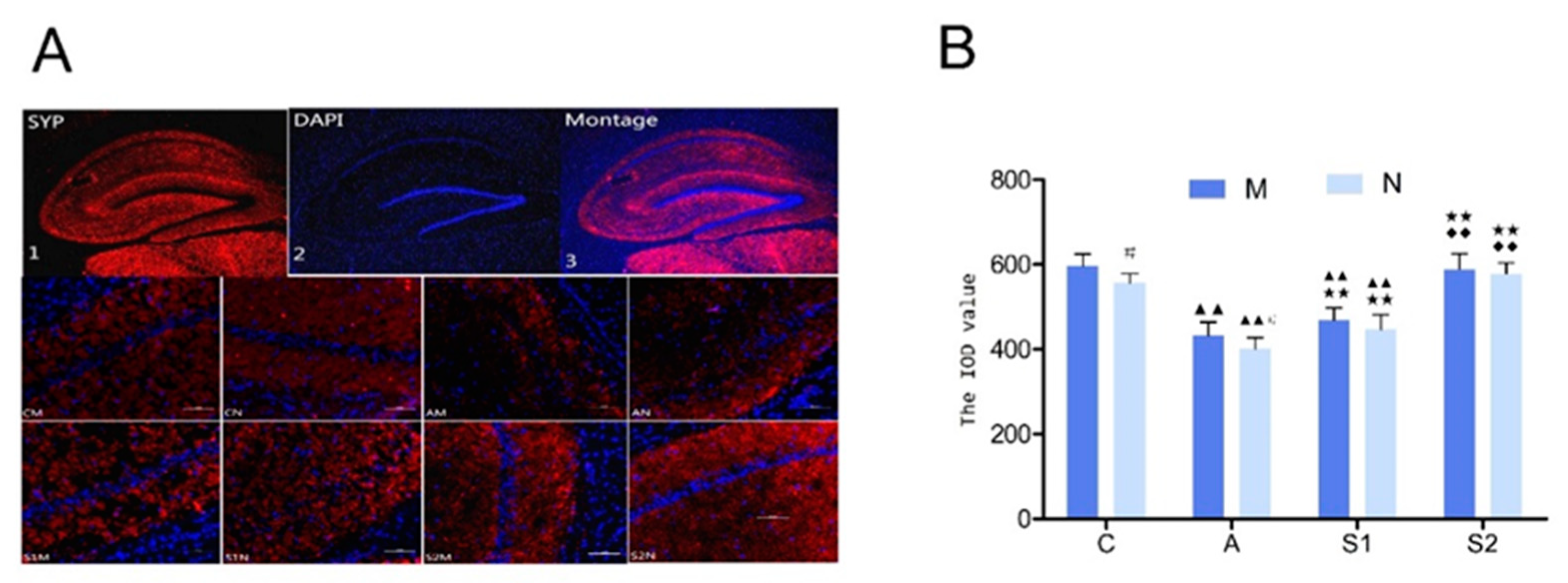
3.4. Immunofluorescence of Synaptophysin (Syp) in Hippocampus
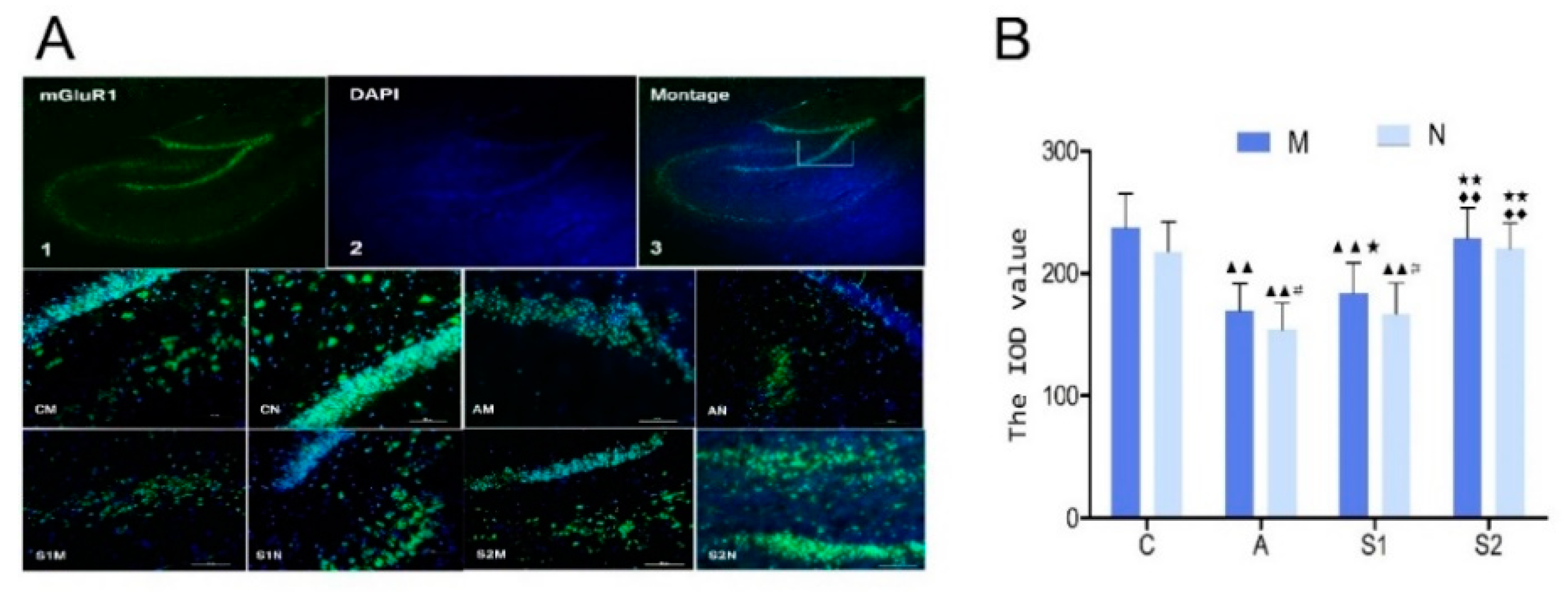
3.5. Immunofluorescence of mGluR1 in Hippocampus
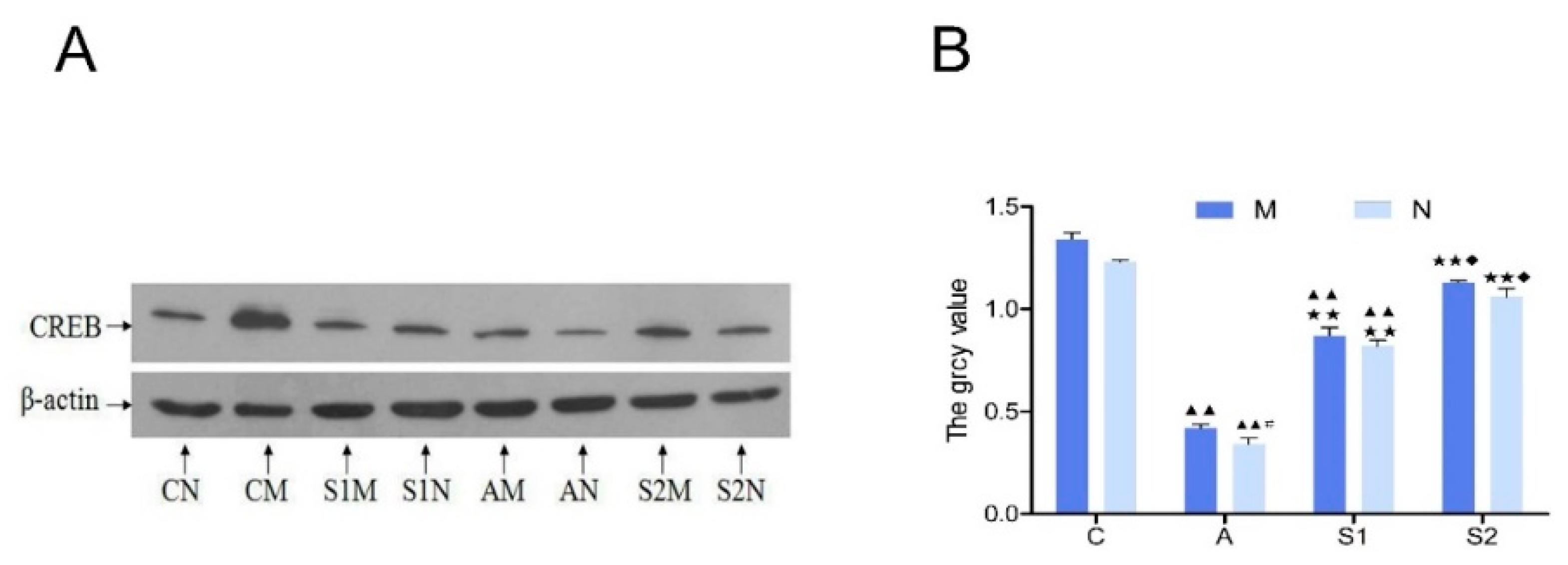
3.6. Hippocampal CREB Protein and mRNA Expression
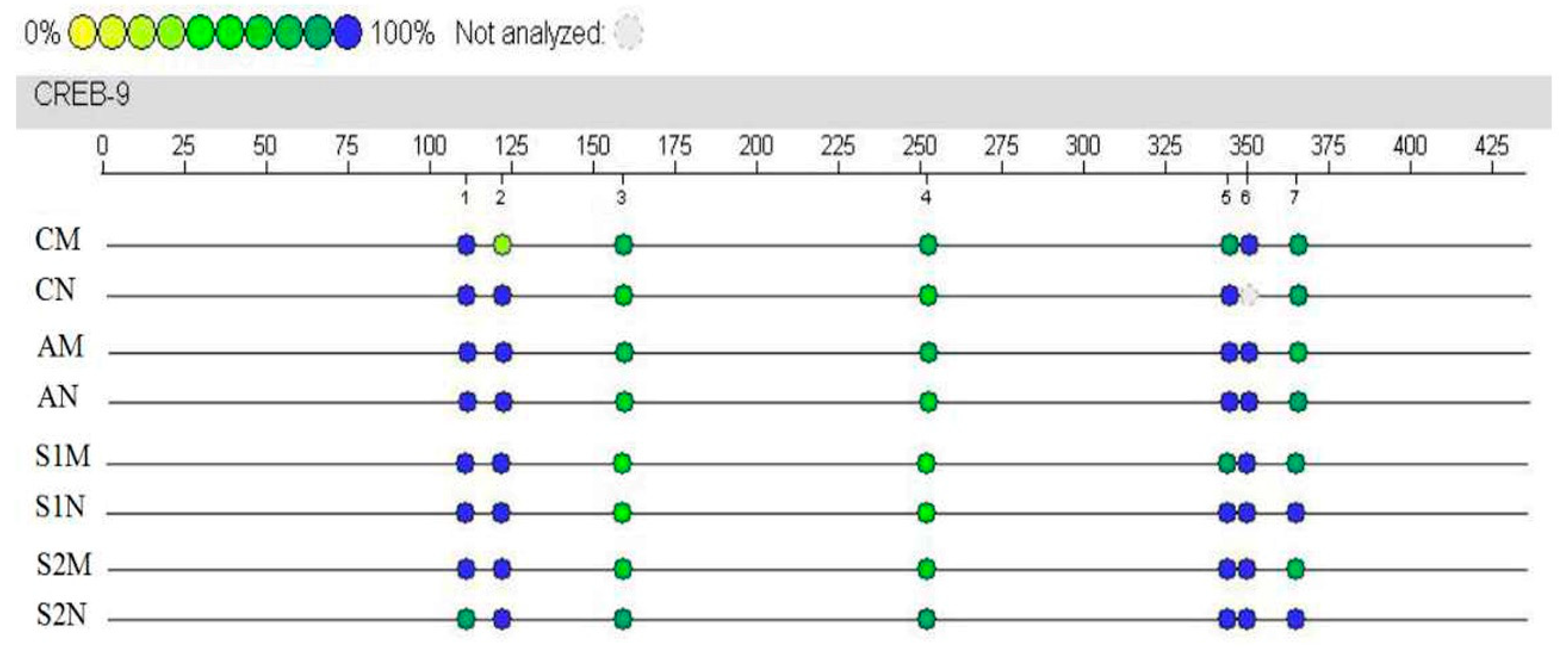
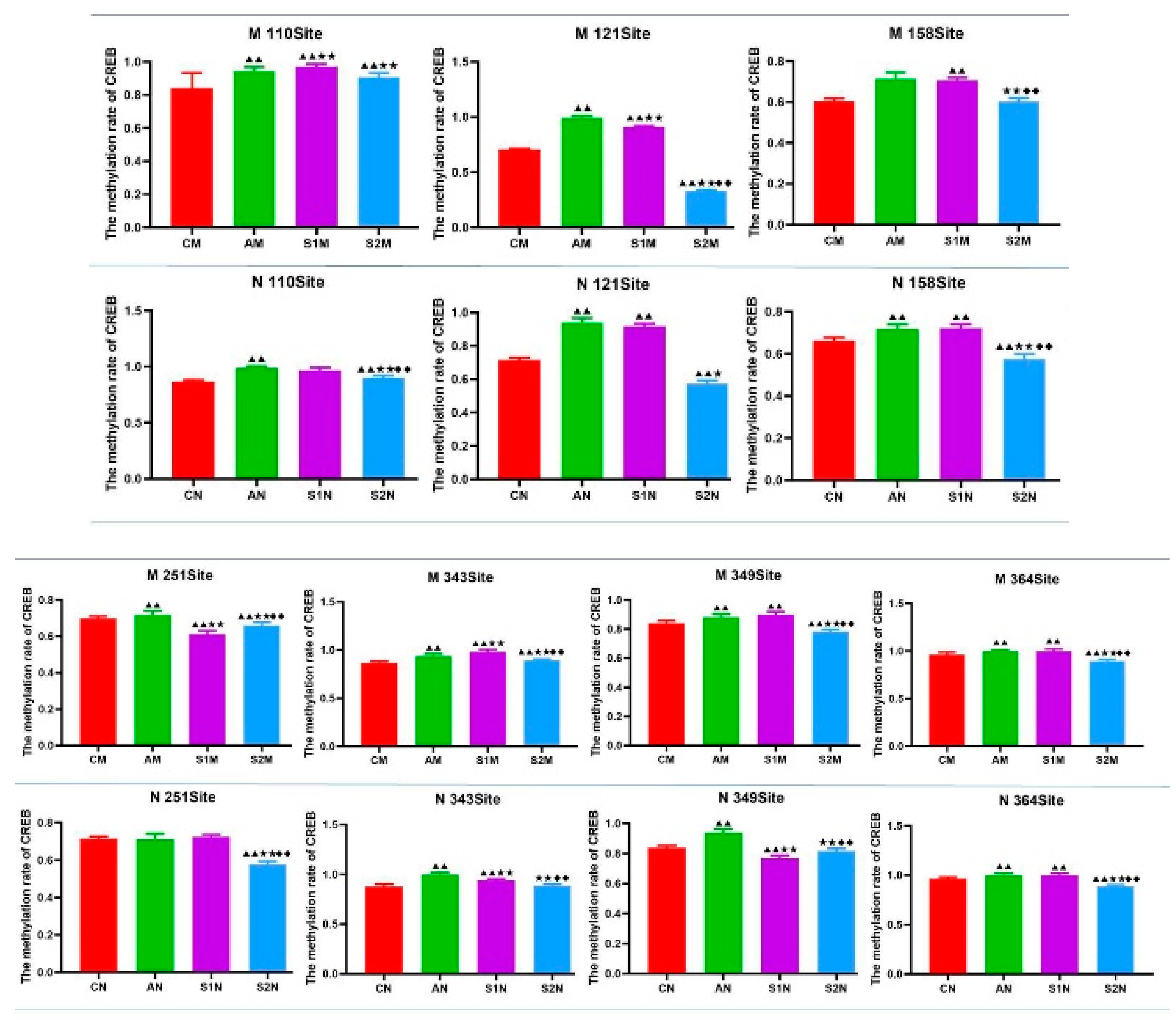
3.7. CREB Gene Methylation Level in Hippocampus
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Donzis, E.J.; Tronson, N.C. Modulation of learning and memory by cytokines: Signaling mechanisms and long term consequences. Neurobiol. Learn. Mem. 2014, 115, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molfese, D.L. Advancing neuroscience through epigenetics: Molecular mechanisms of learning and memory. Dev. Neuropsychol. 2011, 36, 810–827. [Google Scholar] [CrossRef] [PubMed]
- Kennard, J.A.; Woodruff-Pak, D.S. Age sensitivity of behavioral tests and brain substrates of normal aging in mice. Front. Aging Neurosci. 2011, 3, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, K.; Hanaoka, Y.; Nishijima, T.; Okamoto, M.; Chang, H.; Saito, T.; Soya, H. Long-term mild exercise training enhances hippocampus-dependent memory in rats. Int. J. Sports Med. 2015, 36, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Suwabe, K.; Byun, K.; Hyodo, K.; Reagh, Z.M.; Roberts, J.M.; Matsushita, A.; Saotome, K.; Ochi, G.; Fukuie, T.; Suzuki, K.; et al. Rapid stimulation of human dentate gyrus function with acute mild exercise. Proc. Natl. Acad. Sci. USA 2018, 115, 10487–10492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishijima, T.; Torres-Aleman, I.; Soya, H. Exercise and cerebrovascular plasticity. Prog. Brain Res. 2016, 225, 243–268. [Google Scholar]
- Sadigh-Eteghad, S.; Majdi, A.; McCann, S.K.; Mahmoudi, J.; Vafaee, M.S.; Macleod, M.R. d-galactose-induced brain ageing model: A systematic review and meta-analysis on cognitive outcomes and oxidative stress indices. PLoS ONE 2017, 12, e0184122. [Google Scholar] [CrossRef]
- Wei, H.; Li, L.; Song, Q.; Ai, H.; Chu, J.; Li, W. Behavioural study of the d-galactose induced aging model in C57BL/6J mice. Behav. Brain Res. 2005, 157, 245–251. [Google Scholar] [CrossRef]
- Chae, C.H.; Kim, H.T. Forced, moderate-intensity treadmill exercise suppresses apoptosis by increasing the level of NGF and stimulating phosphatidylinositol 3-kinase signaling in the hippocampus of induced aging rats. Neurochem. Int. 2009, 55, 208–213. [Google Scholar] [CrossRef]
- Li, L.; Xu, M.; Shen, B.; Li, M.; Gao, Q.; Wei, S.G. Moderate exercise prevents neurodegeneration in d-galactose-induced aging mice. Neural Regen. Res. 2016, 11, 807–815. [Google Scholar]
- Yu, F.; Xu, B.; Song, C.; Ji, L.; Zhang, X. Treadmill exercise slows cognitive deficits in aging rats by antioxidation and inhibition of amyloid production. Neuroreport 2013, 24, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Berchicci, M.; Lucci, G.; Di, R.F. Benefits of physical exercise on the aging brain: The role of the prefrontal cortex. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1337–1341. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pinilla, F.; Hillman, C. The influence of exercise on cognitive abilities. Compr. Physiol. 2013, 3, 403–428. [Google Scholar] [PubMed]
- Lautenschlager, N.T.; Cox, K.; Cyarto, E.V. The influence of exercise on brain aging and dementia. Biochim. Biophys. Acta 2012, 1822, 474–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baptista, S.; Piloto, N.; Reis, F.; Teixeira-de-Lemos, E.; Garrido, A.P.; Dias, A.; Lourenço, M.; Palmeiro, A.; Ferrer-Antunes, C.; Teixeira, F. Treadmill running and swimming imposes distinct cardiovascular physiological adaptations in the rat: Focus on serotonergic and sympathetic nervous systems modulation. Acta Physiol. Hung. 2008, 95, 365–381. [Google Scholar] [CrossRef] [PubMed]
- Geenen, D.; Buttrick, P.; Scheuer, J. Genetic and epigenetic factors are associated with expression of respiratory chain component NDUFB6 in human skeletal muscle. J. Appl. Physiol. 1988, 65, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Contarteze, R.V.; Manchado Fde, B.; Gobatto, C.A.; De Mello, M.A. Stress biomarkers in rats submitted to swimming and treadmill running exercises. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 151, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.F.; Lang, S.Y.; Zuo, P.P.; Yang, N.; Wang, X.Q.; Xia, C. Effects of d-galactose on the expression of hippocampal peripheral-type benzodiazepine receptor and spatial memory performances in rats. Psychoneuroendocrinology 2006, 31, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Pierozan, P.; Jerneren, F.; Ransome, Y.; Karlsson, O. The choice of euthanasia method affects metabolic serum biomarkers. Basic Clin. Pharmacol. Toxicol. 2017, 121, 113–118. [Google Scholar] [CrossRef]
- Lombroso, P.J.; Ogren, M.P. Learning and memory, part I: Brain regions involved in two types of learning and memory. J. Am. Acad. Child. Adolesc. Psychiatry 2008, 47, 1228–1232. [Google Scholar] [CrossRef]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Anderson, E.M.; Moenk, M.D.; Barbaro, L.; Clarke, D.A.; Matuszewich, L. Effects of pretraining and water temperature on female rats’ performance in the Morris water maze. Physiol. Behav. 2013, 122, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Mameli, M.; Halbout, B.; Creton, C.; Engblom, D.; Parkitna, J.R.; Spanagel, R.; Luscher, C. Cocaine-evoked synaptic plasticity: Persistence in the VTA triggers adaptations in the NAc. Nat. Neurosci. 2009, 12, 1036–1041. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yao, Y.; Wang, L.; Yang, C.; Wang, F.; Guo, J.; Wang, Z.; Yang, Z.; Ming, D. Gastrin-releasing peptide facilitates glutamatergic transmission in the hippocampus and effectively prevents vascular dementia induced cognitive and synaptic plasticity deficits. Exp. Neurol. 2017, 287, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Bartolome, M.V.; Zuluaga, P.; Carricondo, F.; Gil-Loyzaga, P. Immunocytochemical detection of synaptophysin in C57BL/6 mice cochlea during aging process. Brain Res. Rev. 2009, 60, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Simonyi, A.; Ngomba, R.T.; Storto, M.; Catania, M.V.; Miller, L.A.; Youngs, B.; Di Giorgi-Gerevini, V.; Nicoletti, F.; Sun, G.Y. Expression of groups I and II metabotropic glutamate receptors in the rat brain during aging. Brain Res. 2005, 1043, 95–106. [Google Scholar] [CrossRef]
- Chen, Y.C.; Chen, Q.S.; Lei, J.L.; Wang, S.L. Physical training modifies the age-related decrease of GAP-43 and synaptophysin in the hippocampal formation in C57BL/6J mouse. Brain Res. 1998, 806, 238–245. [Google Scholar] [CrossRef]
- Nie, J.; Yang, X.; Tang, Q.; Shen, Q.; Li, S. Willed-movement training reduces brain damage and enhances synaptic plasticity related proteins synthesis after focal ischemia. Brain Res. Bull. 2016, 120, 90–96. [Google Scholar] [CrossRef]
- Seo, H.G.; Kim, D.Y.; Park, H.W.; Lee, S.U.; Park, S.H. Early motor balance and coordination training increased synaptophysin in subcortical regions of the ischemic rat brain. J. Korean Med. Sci. 2010, 25, 1638–1645. [Google Scholar] [CrossRef]
- Cammarota, M.; Bevilaqua, L.R.; Ardenghi, P.; Paratcha, G.; de Stein, M.L.; Izquierdo, I.; Medina, J.H. Learning-associated activation of nuclear MAPK, CREB and Elk-1, along with Fos production, in the rat hippocampus after a one-trial avoidance learning: Abolition by NMDA receptor blockade. Mol. Brain Res. 2000, 76, 36–46. [Google Scholar] [CrossRef]
- Takeo, S.; Niimura, M.; Miyake-Takagi, K.; Nagakura, A.; Fukatsu, T.; Ando, T.; Takagi, N.; Tanonaka, K.; Hara, J. A possible mechanism for improvement by a cognition-enhancer nefiracetam of spatial memory function and cAMP-mediated signal transduction system in sustained cerebral ischaemia in rats. Br. J. Pharmacol. 2003, 138, 642–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waltereit, R.; Weller, M. Signaling from cAMP/PKA to MAPK and synaptic plasticity. Mol. Neurobiol. 2003, 27, 99–106. [Google Scholar] [CrossRef]
- Alzoubi, K.H.; Alkadhi, K.A. Levothyroxin replacement therapy restores hypothyroidism induced impairment of L-LTP induction: Critical role of CREB. Brain Res. Bull. 2014, 100, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Dubnau, J.; Tully, T. Gene discovery in Drosophila: New insights for learning and memory. Annu. Rev. Neurosci. 1998, 21, 407–444. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.C.; Michael, D.; Rose, J.C.; Barad, M.; Casadio, A.; Zhu, H.; Kandel, E.R. MAP kinase translocates into the nucleus of the presynaptic cell and is required for long-term facilitation in Aplysia. Neuron 1997, 18, 899–912. [Google Scholar] [CrossRef]
- Dahl, C.; Guldberg, P. DNA methylation analysis techniques. Biogerontology 2003, 4, 233–250. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, M. DNA methylation in cancer: Too much, but also too little. Oncogene 2002, 21, 5400–5413. [Google Scholar] [CrossRef] [PubMed]
- Horvath, S.; Zhang, Y.; Langfelder, P.; Kahn, R.S.; Boks, M.P.; van, E.K.; van den Berg, L.H.; Ophoff, R.A. Aging effects on DNA methylation modules in human brain and blood tissue. Genome Biol. 2012, 13, R97. [Google Scholar] [CrossRef]
- Ling, C.; Poulsen, P.; Simonsson, S.; Ronn, T.; Holmkvist, J.; Almgren, P.; Hagert, P.; Nilsson, E.; Mabey, A.G.; Nilsson, P.; et al. Genetic and epigenetic factors are associated with expression of respiratory chain component NDUFB6 in human skeletal muscle. J. Clin. Investig. 2007, 117, 3427–3435. [Google Scholar] [CrossRef] [Green Version]
- Ronn, T.; Poulsen, P.; Hansson, O.; Holmkvist, J.; Almgren, P.; Nilsson, P.; Tuomi, T.; Isomaa, B.; Groop, L.; Vaag, A.; et al. Age influences DNA methylation and gene expression of COX7A1 in human skeletal muscle. Diabetologia 2008, 51, 1159–1168. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Pinilla, F.; Zhuang, Y.; Feng, J.; Ying, Z.; Fan, G. Exercise impacts brain-derived neurotrophic factor plasticity by engaging mechanisms of epigenetic regulation. Eur. J. Neurosci. 2011, 33, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, A.S., Jr.; Castro, A.A.; Moreira, E.L.; Glaser, V.; Santos, A.R.; Tasca, C.I.; Latini, A.; Prediger, R.D. Short bouts of mild-intensity physical exercise improve spatial learning and memory in aging rats: Involvement of hippocampal plasticity via AKT, CREB and BDNF signaling. Mech. Ageing Dev. 2011, 132, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Subbanna, S.; Joshi, V.; Basavarajappa, B.S. Activity-dependent signaling and epigenetic abnormalities in mice exposed to postnatal ethanol. Neuroscience 2018, 392, 230–240. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Wang, L.; Zhang, S.; Hu, X.; Yang, H.; Xi, L. Timing-Dependent Protection of Swimming Exercise against d-Galactose-Induced Aging-Like Impairments in Spatial Learning/Memory in Rats. Brain Sci. 2019, 9, 236. https://doi.org/10.3390/brainsci9090236
Li X, Wang L, Zhang S, Hu X, Yang H, Xi L. Timing-Dependent Protection of Swimming Exercise against d-Galactose-Induced Aging-Like Impairments in Spatial Learning/Memory in Rats. Brain Sciences. 2019; 9(9):236. https://doi.org/10.3390/brainsci9090236
Chicago/Turabian StyleLi, Xue, Lu Wang, Shuling Zhang, Xiang Hu, Huijun Yang, and Lei Xi. 2019. "Timing-Dependent Protection of Swimming Exercise against d-Galactose-Induced Aging-Like Impairments in Spatial Learning/Memory in Rats" Brain Sciences 9, no. 9: 236. https://doi.org/10.3390/brainsci9090236