Music Training Positively Influences the Preattentive Perception of Voice Onset Time in Children with Dyslexia: A Longitudinal Study

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Longitudinal Study: Procedure
2.3. MMN Experiment: Procedure
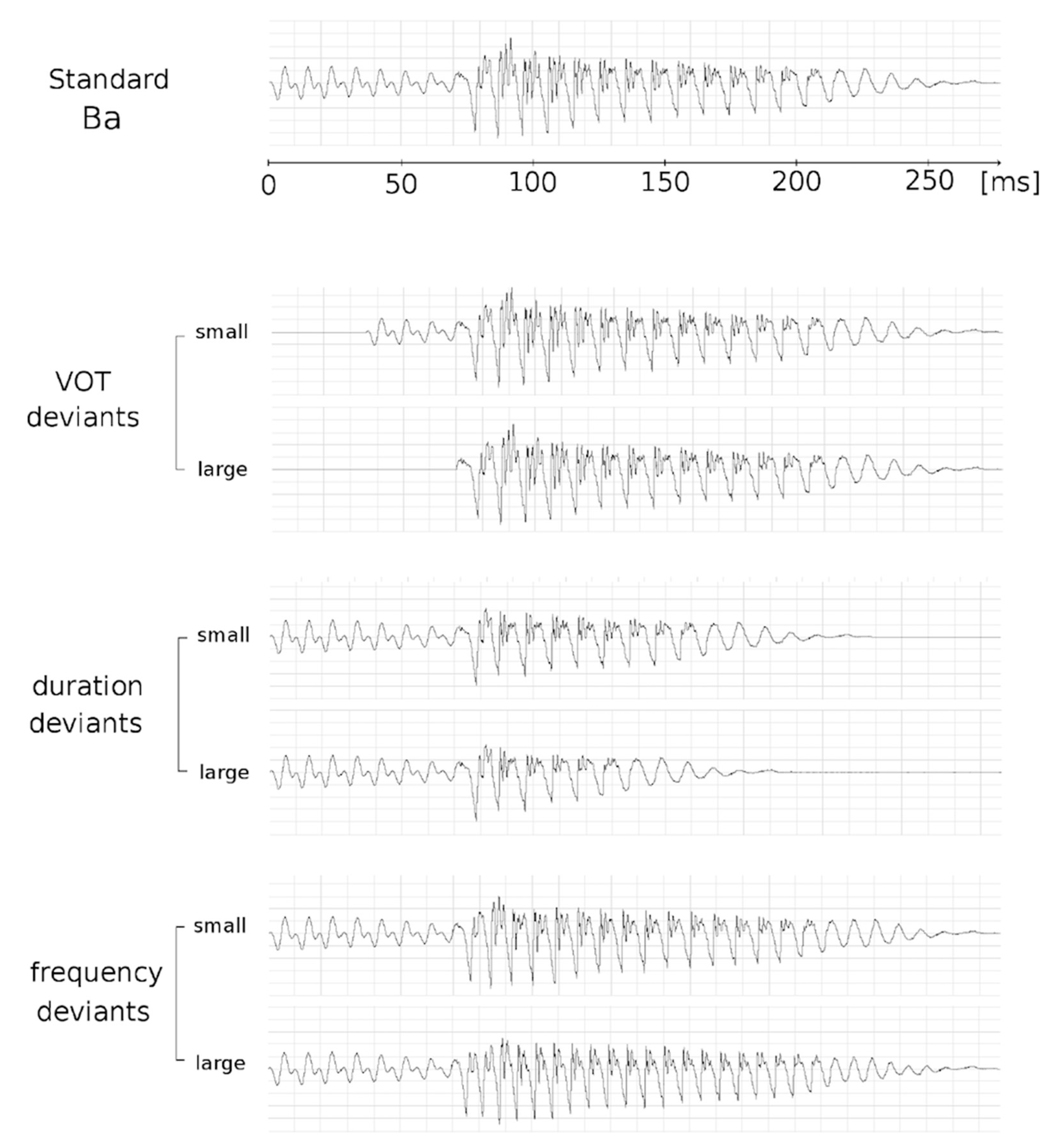
2.4. Stimuli
2.5. ERP Recording and Processing
2.6. Data Analysis
3. Results
3.1. Neuropsychological and Speech Assessments
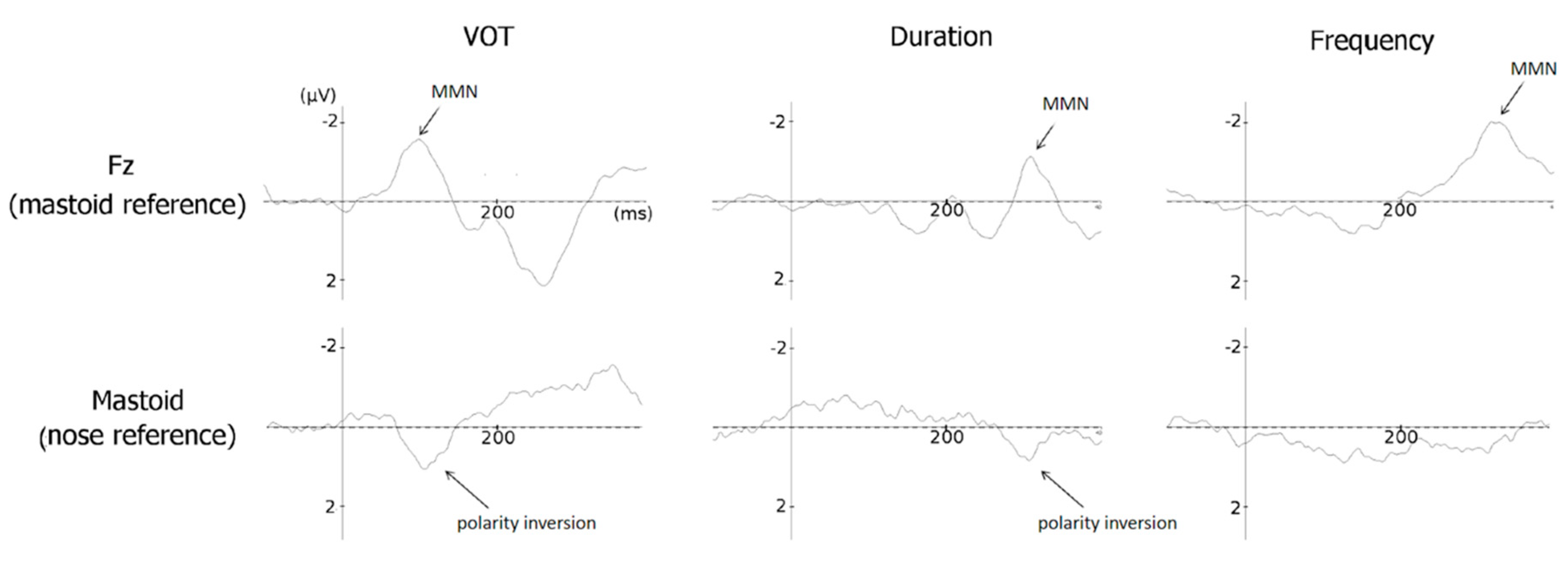
3.2. MMN Amplitude
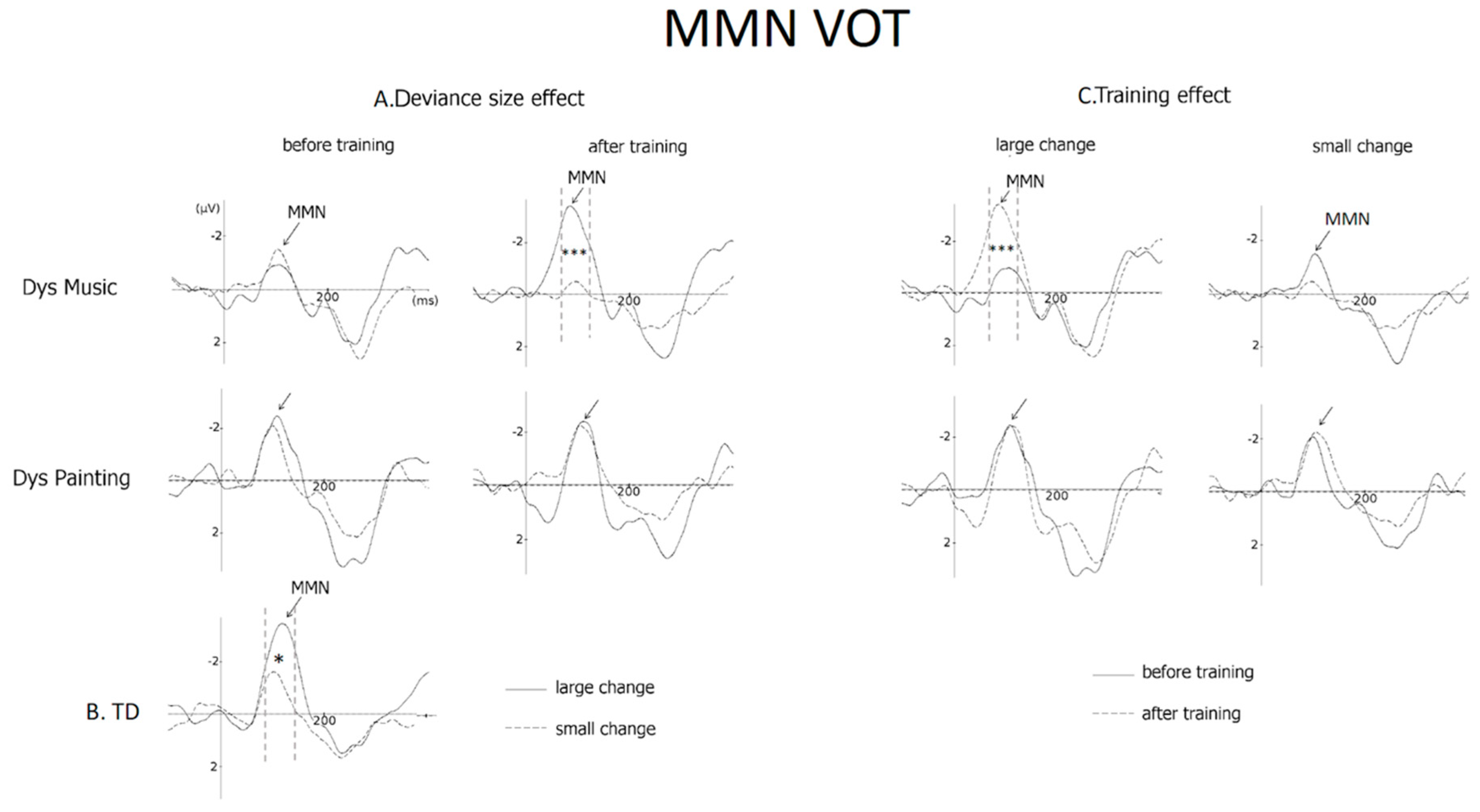
3.2.1. VOT (MMN Amplitude)
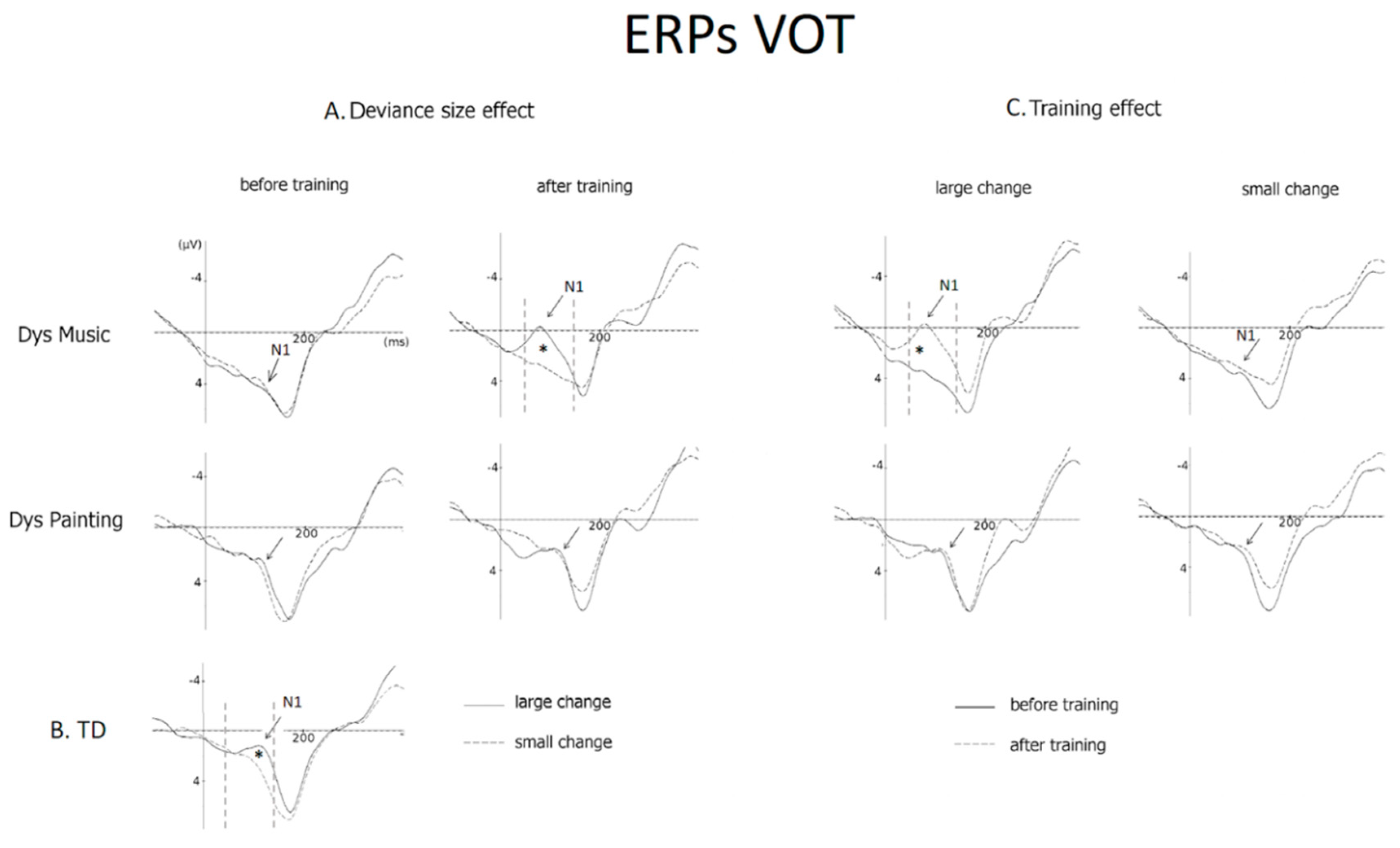
3.2.2. VOT (N1 Amplitude)
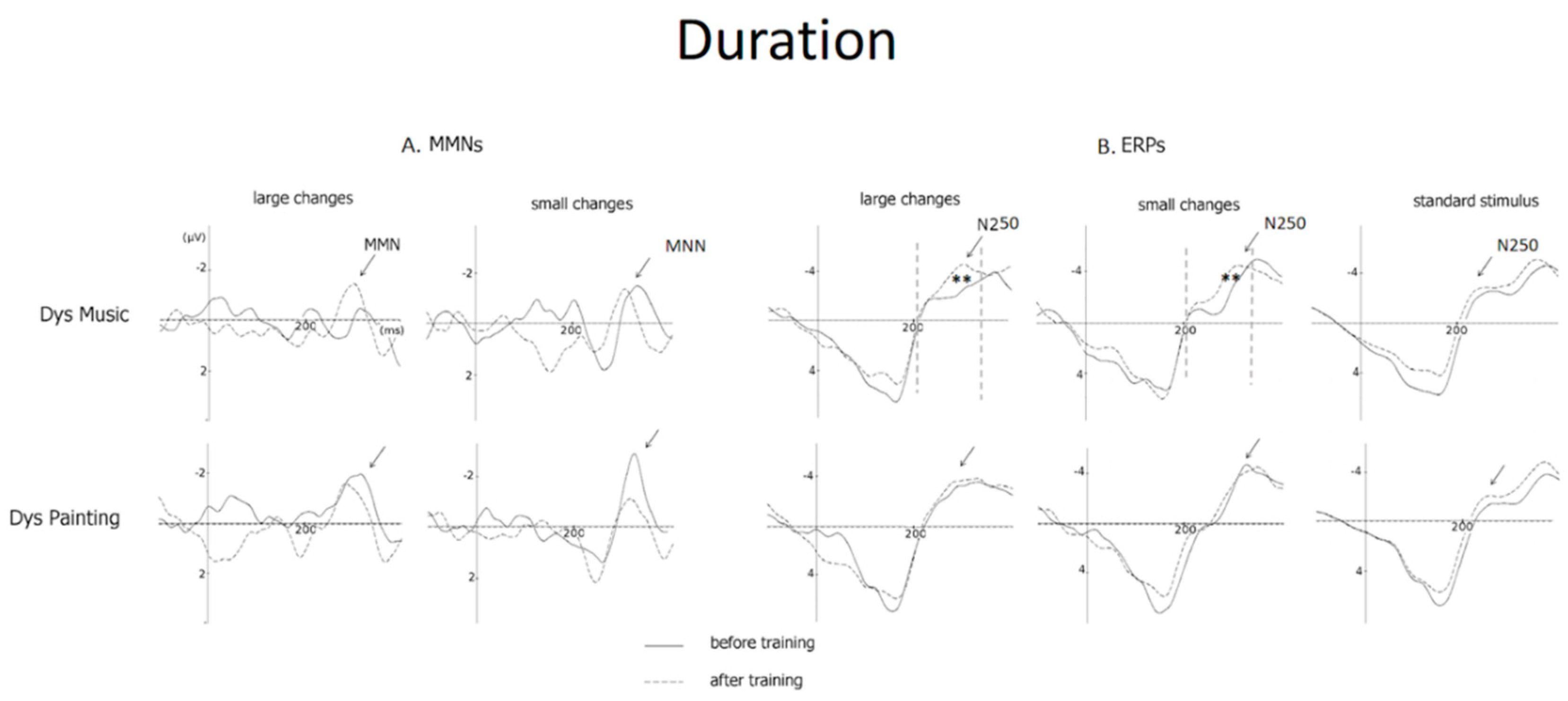
3.3. Duration
3.3.1. Duration (MMN Amplitude)
3.3.2. Duration (N250 Amplitude)
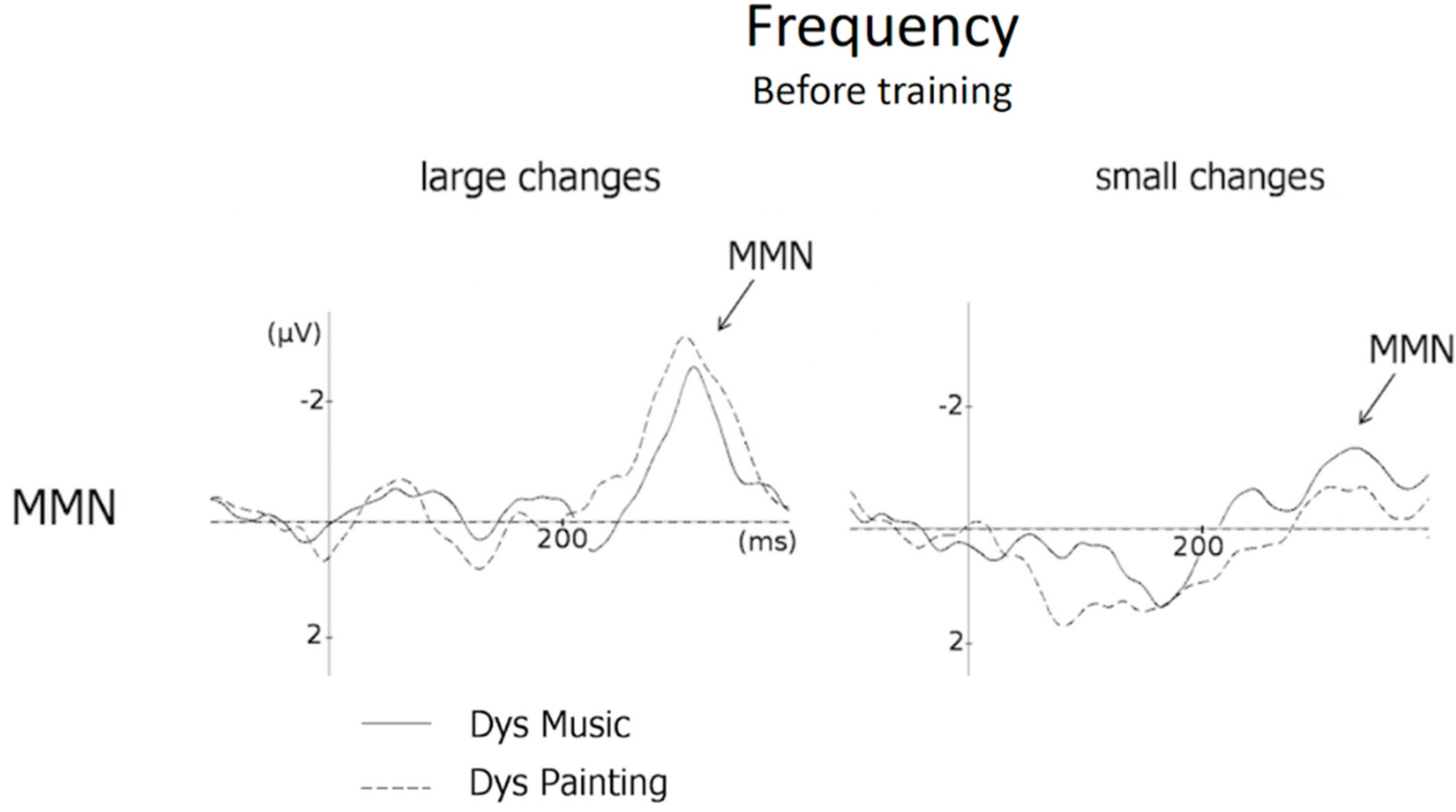
3.4. Frequency (MMN and N250 Amplitude)
4. Discussion
4.1. VOT Deviant Stimuli
4.2. Duration Deviant Stimuli
4.3. Frequency Deviant Stimuli
4.4. Psychometric Tests
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Démonet, J.F.; Taylor, M.J.; Chaix, Y. Developmental dyslexia. Lancet 2004, 363, 1451–1460. [Google Scholar] [CrossRef]
- Habib, M. The neurological basis of developmental dyslexia. Brain 2000, 123, 2373–2399. [Google Scholar] [CrossRef] [PubMed]
- Collective Expertise INSERM, CNDRSDI. Dyslexie, Dysorthographie, Dyscalculie: Bilan des Données Scientifiques; INSERM: Paris, France, 2007. [Google Scholar]
- Norton, E.S.; Beach, S.D.; Gabrieli, J.D. Neurobiology of dyslexia. Curr. Opin. Neurobiol. 2015, 30, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Snowling, M.J. Dyslexia; Blackwell: Oxford, UK, 2000. [Google Scholar]
- Valdois, S.; Bosse, M.L.; Tainturier, M.J. The cognitive deficits responsible for developmental dyslexia: Review of evidence for a selective visual attention disorder. Dyslexia 2004, 10, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.; Zorzi, M.; Ziegler, J.C. Understanding Dyslexia Through Personalized Large-Scale Computational Models. Psychol. Sci. 2019, 30, 386–395. [Google Scholar] [CrossRef]
- Ramus, F. Developmental dyslexia: Specific phonological deficit or general sensorimotor dysfunction? Curr. Opin. Neurobiol. 2003, 13, 212–218. [Google Scholar] [CrossRef]
- Saksida, A.; Iannuzzi, S.; Bogliotti, C.; Chaix, Y.; Demonet, J.F.; Bricout, L.; Billard, C.; Nguyen-Morel, M.A.; Le Heuzey, M.F.; Soares-Boucaud, I.; et al. Phonological skills, visual attention span, and visual stress in de- velopmental dyslexia. Dev. Psychol. 2016, 52, 1503–1516. [Google Scholar] [CrossRef]
- White, S.; Milne, E.; Rosen, S.; Hansen, P.; Swettenham, J.; Frith, U.; Ramus, F. The role of sensorimotor impairments in dyslexia: A multiple case study of dyslexic children. Dev. Sci. 2006, 9, 237–255. [Google Scholar] [CrossRef]
- Ahissar, M.; Lubin, Y.; PutterKatz, H.; Banai, K. Dyslexia and the failure to form a perceptual anchor. Nat. Neurosci. 2006, 9, 1558–1564. [Google Scholar] [CrossRef] [PubMed]
- Kimppa, L.; Shtyrov, Y.; Partanen, E.; Kujala, T. Impaired neural mechanism for online novel word acquisition in dyslexic children. Sci. Rep. 2018, 24, 12779. [Google Scholar] [CrossRef]
- Thomson, J.M.; Goswami, U. Learning novel phonological representations in developmental dyslexia: Associations with basic auditory processing of rise time and phonological awareness. Read. Writ. 2010, 23, 453–473. [Google Scholar] [CrossRef]
- Ramus, F.; Marshall, C.R.; Rosen, S.; van der Lely, H.K. Phonological deficits in specific language impairment and developmental dyslexia: Towards a multidimensional model. Brain 2013, 136, 630–645. [Google Scholar] [CrossRef] [PubMed]
- Boets, B.; de Beeck, H.; Vandermosten, M.; Scott, S.K.; Gillebert, C.R.; Mantini, D.; Bulthé, J.; Sunaert, S.; Wouters, J.; Ghesquière, P. Intact but less accessible phonetic representations in adults with dyslexia. Science 2013, 342, 1251–1254. [Google Scholar] [CrossRef]
- Lovio, R.; Näätänen, R.; Kujala, T. Abnormal pattern of cortical speech feature discrimination in 6-year-old children at risk for dyslexia. Brain Res. 2010, 1335, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, S.; Mahncke, H.; Salz, T.; Tallal, P.; Roberts, T.; Merzinech, M. Cortical auditory signal processing in poor readers. Proc. Natl. Acad. Sci. USA 1999, 25, 6483–6488. [Google Scholar] [CrossRef]
- Ziegler, J.C.; Ferrand, L. Orthography shapes the perception of speech: The consistency effect in auditory word recognition. Psychon. Bull. Rev. 1998, 5, 683–689. [Google Scholar] [CrossRef]
- Hämäläinen, J.A.; Leppänen, P.H.T.; Guttorm, T.K.; Lyytinen, H. N1 and P2 components of auditory event-related potentials in children with and without reading disabilities. Clin. Neurophysiol. 2007, 118, 2263–2275. [Google Scholar] [CrossRef] [PubMed]
- Hämäläinen, J.A.; Leppänen, P.H.T.; Guttorm, T.K.; Lyytinen, H. Event-related potentials to pitch and rise time change in children with reading disabilities and typically reading children. Clin. Neurophysiol. 2008, 119, 100–115. [Google Scholar] [CrossRef] [PubMed]
- Chobert, J.; François, C.; Habib, M.; Besson, M. Deficit in the preattentive processing of syllables in children with dyslexia. Neuropsychologia 2012, 50, 2044–2055. [Google Scholar] [CrossRef]
- Näätänen, R.; Gaillard, A.W.K.; Mäntysalo, S. Early selective-attention effect on evoked potential reinterpreted. Acta Psychol. 1978, 42, 313–329. [Google Scholar] [CrossRef]
- Näätänen, R.; Paavilainen, P.; Rinne, T.; Alho, K. The mismatch negativity (MMN) in basic research of central auditory processing: A review. Clin. Neurophysiol. 2007, 118, 2544–2590. [Google Scholar] [CrossRef]
- Kujala, T.; Tervaniemi, M.; Schröger, E. The mismatch negativity in cognitive and clinical neuroscience: Theoretical and methodological considerations. Biol. Psychol. 2007, 74, 1–19. [Google Scholar] [CrossRef]
- Näätänen, R.; Pakarinen, S.; Rinne, T.; Takegata, R. The mismatch negativity (MMN): Towards the optimal paradigm. Clin. Neurophysiol. 2004, 115, 140–144. [Google Scholar] [CrossRef]
- Lisker, L.; Abramson, A.S. Some effects of context on voice onset time in English stops. Lang. Speech 1967, 10, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Serniclaes, W. Etude Expérimentale de la Perception du Trait de Voisement des Occlusives du Français. Unpublished Doctoral’s Thesis, Université Libre de Bruxelles, Bruxelles, Belgium, 1987. [Google Scholar]
- Banai, K.; Hornickel, J.; Skoe, E.; Nicol, T.; Zecker, S.G.; Kraus, N. Reading and subcortical auditory function. Cereb. Cortex 2009, 19, 2699–2707. [Google Scholar] [CrossRef] [PubMed]
- Hämäläinen, J.; Salminen, H.; Leppanen, P. Basic auditory processing deficits in dyslexia; A systematic review of the behavioural and event-related potential field evidence. J. Learn. Disabil. 2013, 46, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Lovio, R.; Pakarinen, S.; Huotilainen, M.; Alku, P.; Silvennoinen, S.; Näätänen, R.; Kujala, T. Auditory discrimination profiles of speech sound changes in 6-year-old children as determined with the multi-feature MMN paradigm. Clin. Neurophysiol. 2009, 120, 916–921. [Google Scholar] [CrossRef]
- Baldeweg, T.; Richardson, A.; Watkins, S.; Foale, C.; Gruzelier, J. Impaired auditory frequency discrimination in dyslexia detected with mismatch evoked potentials. Ann. Neurol. 1999, 45, 495–503. [Google Scholar] [CrossRef]
- Santos, A.; Joly-Pottuz, B.; Moreno, S.; Habib, M.; Besson, M. Behavioral and event-related potentials evidence for pitch discrimination deficits in dyslexic children: Improvement after intensive phonic intervention. Neuropsychologia 2007, 45, 1080–1090. [Google Scholar] [CrossRef]
- Cutini, S.; Szucs, D.; Mead, N.; Huss, M.; Goswami, U. Atypical right hemisphere response to slow temporal modulations in children with developmental dyslexia. Neuroimage 2016, 143, 40–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Power, A.J.; Colling, L.J.; Mead, N.; Barnes, L.; Goswami, U. Neural encoding of the speech envelope by children with developmental dyslexia. Brain Lang. 2016, 160, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Goswami, U. A temporal sampling framework for developmental dyslexia. Trends Cogn. Sci. 2011, 15, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Goswami, U.; Power, A.J.; Lallier, M.; Facoetti, A. Oscillatory “temporal sampling” and developmental dyslexia: Toward an over-arching theoretical framework. Front. Hum. Neurosci. 2014, 8, 904. [Google Scholar] [CrossRef] [PubMed]
- Hämäläinen, J.; Rupp, A.; Soltész, F.; Szücs, D.; Goswami, U. Reduced phase locking to slow amplitude modulation in adults with dyslexia: An MEG study. NeuroImage 2012, 59, 2952–2961. [Google Scholar] [CrossRef] [PubMed]
- Lehongre, K.; Ramus, F.; Villiermet, N.; Schwartz, D.; Giraud, A.L. Altered low-gamma sampling in auditory cortex accounts for the three main facets of dyslexia. Neuron 2011, 72, 1080–1090. [Google Scholar] [CrossRef]
- Cantiania, C.; Ortiz-Mantillab, S.; Rivaa, V.; Piazzac, C.; Bettonia, R.; Musacchia, G.; Moltenia, M.; Marino, C.; Benasich, A.A. Reduced left-lateralized pattern of event-related EEG oscillations in infants at familial risk for language and learning impairment. Neuroimage Clin. 2019, 22, 101778. [Google Scholar] [CrossRef]
- Halliday, L.F.; Barry, J.G.; Hardiman, M.J.; Bishop, D.V.M. Late, not early mismatch responses to changes in frequency are reduced or deviant in children with dyslexia: An event-related potential study. J. Neurodev. Disord. 2014, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bishop, D.V.M. Using mismatch negativity to study central auditory processing in developmental language and literacy impairments: Where are we, and where should we be going? Psychol. Bull. 2007, 133, 651–672. [Google Scholar] [CrossRef] [PubMed]
- Goswami, U.; Huss, M.; Mead, N.; Fosker, T.; Verney, J.P. Perception of patterns of musical beat distribution in phonological developmental dyslexia: Significant longitudinal relations with word reading and reading comprehension. Cortex 2013, 49, 1363–1376. [Google Scholar] [CrossRef]
- Huss, M.; Verney, J.P.; Fosker, T.; Mead, N.; Goswami, U. Music, rhythm, rise time perception and developmental dyslexia: Perception of musical meter predicts reading and phonology. Cortex 2011, 47, 674–689. [Google Scholar] [CrossRef] [Green Version]
- Forgeard, M.; Schlaug, G.; Norton, A.; Rosam, C.; Iyengar, U.; Winner, E. The relation between music and phonological processing innormal-reading children and children with dyslexia. Music Percept. Interdiscip. J. 2008, 25, 383–390. [Google Scholar] [CrossRef]
- Frey, A.; François, C.; Chobert, J.; Besson, M.; Ziegler, J. Behavioral and electrophysiological investigation of speech perception deficits in silence, noise and envelope conditions in developmental dyslexia. Neuropsychologia 2018. [Google Scholar] [CrossRef] [PubMed]
- Liberman, I.Y.; Shankweiler, D. Phonology and the problems of learning to read and write. Remedial Spec. Educ. 1985, 6, 8–17. [Google Scholar] [CrossRef]
- Overy, K. Dyslexia and music: From timing deficits to musical intervention. Ann. N. Y. Acad. Sci. 2003, 999, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, J.C.; Goswami, U. Reading acquisition, developmental dyslexia, and skilled reading across languages: A psycholinguistic grain size theory. Psychol. Bull. 2005, 131, 3–29. [Google Scholar] [CrossRef]
- Abrams, D.A.; Bhatara, A.; Ryali, S.; Balaban, E.; Levitin, D.J.; Menon, V. Decoding temporal structure in music and speech relies on shared brain resources but elicits different fine-scale spatial patterns. Cereb. Cortex 2011, 21, 1507–1518. [Google Scholar] [CrossRef] [PubMed]
- Besson, M. Meaning, structure and time in language and music. Neural substrates of cognitive processes. Special issue in homage to Jean Requin. Curr. Psychol. Cogn. 1998, 17, 921–951. [Google Scholar]
- Besson, M.; Chobert, J.; Marie, C. Transfer of training between music and speech: Common processing, attention, and memory. Front. Psychol. 2011, 2, 94. [Google Scholar] [CrossRef] [PubMed]
- Kraus, N.; Chandrasekaran, B. Music training for the development of auditory skills. Nat. Rev. Neurosci. 2010, 11, 599–605. [Google Scholar] [CrossRef]
- Maess, B.; Koelsch, S.; Gunter, T.C.; Friederici, A.D. Musical syntax is processed in Broca’s area: An MEG study. Nat. Neurosci. 2001, 4, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.D. Language, music, syntax and the brain. Nat. Neurosci. 2003, 6, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.D. Music, Language, and the Brain; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Zatorre, R.J.; Gandour, J.T. Neural specializations for speech and pitch: Moving beyond the dichotomies. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 1087–1104. [Google Scholar] [CrossRef] [PubMed]
- Kishon-Rabin, L.; Amir, O.; Vexler, Y.; Zaltz, Y. Pitch discrimination: Are professional musicians better than non-musicians? J. Basic Clin. Physiol. Pharmacol. 2001, 12, 125–143. [Google Scholar] [CrossRef] [PubMed]
- Marie, C.; Kujala, T.; Besson, M. Musical and linguistic expertise influence preattentive and attentive processing of non-speech sounds. Cortex 2012, 10, 1016. [Google Scholar]
- Spiegel, M.F.; Watson, C.S. Performance on frequency discrimination tasks by musicians and nonmusicians. J. Acoust. Soc. Am. 1984, 76, 1690–1695. [Google Scholar] [CrossRef]
- Tervaniemi, M.; Just, V.; Koelsch, S.; Widmann, A.; Schröger, E. Pitch discrimination accuracy in musicians vs. nonmusicians: An event-related potential and behavioral study. Exp. Brain Res. 2005, 161, 1–10. [Google Scholar] [CrossRef]
- Kühnis, J.; Elmer, S.; Meyer, M.; Jäncke, L. The encoding of vowels and temporal speech cues in the auditory cortex of professional musicians: An EEG study. Neuropsychologia 2013, 5l, 1608–1618. [Google Scholar] [CrossRef]
- Zuk, J.; Ozernov-Palchik, O.; Kim, H.; Lakshminarayanan, K.; Gabrieli, J.D.E.; Tallal, P.; Gaab, N. Enhanced Syllable Discrimination Thresholds in Musicians. PLoS ONE 2013, 8, e80546. [Google Scholar] [CrossRef]
- Parbery-Clark, A.; Tierney, A.; Strait, D.L.; Kraus, N. Musicians have fine-tuned neural distinction of speech syllables. Neuroscience 2012, 219, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Besson, M.; Dittinger, E.; Barbaroux, M. How music training influences language processing: Evidence against informational encapsulation. L’année Psychol. 2018, 118, 273–288. [Google Scholar]
- Nikjeh, D.A.; Lister, J.J.; Frisch, S.A. The relationship between pitch discrimination and vocal production: Comparison of vocal and instrumental musicians. J. Acoust. Soc. Am. 2009, 125, 328–338. [Google Scholar] [CrossRef]
- Tervaniemi, M.; Rytkönen, M.; Schröger, E.; Ilmoniemi, R.J.; Näätänen, R. Superior formation of cortical memory traces for melodic patterns in musicians. Learn. Mem. 2001, 8, 295–300. [Google Scholar] [CrossRef]
- Milovanov, R.; Huotilainen, M.; Esquef, P.A.A.; Välimäki, V.; Alku, P.; Tervaniemi, M. The role of musical aptitude and language skills in preattentive duration determination in school-aged children. Neurosci. Lett. 2009, 460, 161–165. [Google Scholar] [CrossRef]
- Chobert, J.; Marie, C.; François, C.; Schön, D.; Besson, M. Enhanced passive and active processing of syllables in musician children. J. Cogn. Neurosci. 2011, 23, 3874–3887. [Google Scholar] [CrossRef] [PubMed]
- Anvari, S.H.; Trainor, L.J.; Woodside, J.; Levy, B.A. Relation among musical skills, phonological processing and early reading ability in preschool children. J. Exp. Psychol. 2002, 83, 111–130. [Google Scholar] [CrossRef]
- Degé, F.; Schwarzer, G. The effect of a music program on phonological awareness in preschoolers. Front. Psychol. 2011, 2, 124. [Google Scholar] [CrossRef]
- Slevc, L.R.; Miyake, A. Individual differences in second language proficiency: Does musical ability matter? Psychol. Sci. 2006, 17, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Moreno, S.; Marques, C.; Santos, A.; Santos, M.; Castro, S.L.; Besson, M. Musical training influences linguistic abilities in 8-year-old children: More evidence for brain plasticity. Cereb. Cortex 2009, 19, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Schellenberg, E.G. Music lessons enhance IQ. Psychol. Sci. 2004, 15, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Chobert, J.; François, C.; Velay, J.-L.; Besson, M. Twelve months of active musical training in 8 to 10 year old children enhances the preattentive processing of syllabic duration and Voice Onset Time. Cereb. Cortex 2014, 24, 956–967. [Google Scholar] [CrossRef] [PubMed]
- Rugg, M.D.; Coles, M.G.H. Electrophysiology of Mind; Oxford University Press: New York, NY, USA, 1995. [Google Scholar]
- Donchin, E.; Ritter, W.; McCallum, C. Cognitive psychophysiology: The endogenous components of the ERP. In Event-Related Brain Potentials in Man; Callaway, E., Tueting, P., Koslow, S.H., Eds.; Academic Press: New York, NY, USA, 1978; pp. 349–411. [Google Scholar]
- Ritter, W.; Simon, R.; Vaughan, H.G., Jr. Event-related potential correlates to two stages of information processing in physical and semantic discrimination tasks. Psychophysiology 1983, 20, 168–179. [Google Scholar] [CrossRef]
- Ceponiene, R.; Shestakova, A.; Balan, P.; Alku, P.; Yiaguchi, K.; Näätänen, R. Children’s auditory event-related potentials index sound complexity and “speechness”. Int. J. Neurosci. 2001, 109, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Lefavrais, J. Test de l’Alouette, rev. version; ECPA: Paris, France, 2005. [Google Scholar]
- Bertrand, D.; Fluss, J.; Billard, C.; Ziegler, J.C. Efficacité, sensibilité, spécificité: Comparaison de différents tests de lecture [Efficiency, sensitivity, specificity: Comparison of different reading tests]. Année Psychol. 2010, 110, 299–320. [Google Scholar] [CrossRef]
- Wechsler, D. Wechsler Intelligence Scale for Children, 4th ed.; WISC-IV; The Psychological Corporation: San Antonio, TX, USA, 2003. [Google Scholar]
- Korkman, M.; Kemp, S.L.; Kirk, U. NEPSY: Bilan Neuropsychologique de L’enfant; ECPA (Editions du centre de psychologie appliquée): Paris, France, 2004. [Google Scholar]
- Raven, J.C. Standard Progressive Matrices: Sets A, B, C, D, E; Psychologists Press: Oxford, UK, 1976. [Google Scholar]
- Chavez, M.; Day, R.; Deyell, S.; Ellis, P.; Fazio, S.; Green, P.; Johnston, D.; Jonasson, B.; Levine, J.; Orler, T.; et al. Adobe Audition Software. 2003. Available online: http://www.adobe.com/products/audition.html (accessed on 25 April 2011).
- Boersma, P.; Weenink, D. Praat [Computer Software], Version 4.0. Available online: http://www.fon.hum.uva.nl/praat/ (accessed on 18 April 2001).
- Jasper, H.A. The ten—twenty system of the International Federation. Electroencephologr. Clin. Neurophysiol. 1958, 10, 371–375. [Google Scholar]
- Schröger, E.; Wolff, C. Attentional orienting and reorienting is indicated by human event-related brain potentials. Neuroreport 1998, 9, 3355–3358. [Google Scholar] [CrossRef] [PubMed]
- Dufor, O.; Serniclaes, W.; Sprenger-Charolles, L.; Démonet, J.F. Left premotor cortex and allophonic speech perception in dyslexia: A PET study. Neuroimage 2009, 46, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoonhorst, I.; Medina, V.; Colin, C.; Markessis, E.; Radeau, M.; Deltenre, P.; Serniclaes, W. Categorical perception of voicing, colors and facial expressions: A developmental study. Speech Commun. 2011, 53, 417–430. [Google Scholar] [CrossRef]
- Noordenbos, M.W.; Segers, E.; Serniclaes, W.; Mitterer, H.; Verhoeven, L. Neural evidence of allophonic perception in children at risk for dyslexia. Neuropsychologia 2012, 50, 2010–2017. [Google Scholar] [CrossRef] [PubMed]
- Serniclaes, W.; Heghe, S.V.; Mousty, P.; Carré, R.; Sprenger-Charolles, L. Allophonic mode of speech perception in dyslexia. J. Exp. Child Psychol. 2004, 87, 336–361. [Google Scholar] [CrossRef] [Green Version]
- Habib, M.; Lardy, C.; Desiles, T.; Commeiras, C.; Chobert, J.; Besson, M. Music and dyslexia: A new musical training method to improve reading and related disorders. Front. Psychol. 2016, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Habib, M.; Commeiras, C. «Mélodys»: Remédiation Cognitivo-Musicale des Troubles de L’apprentissage; De Boeck: Bruxelles, Belgium, 2014. [Google Scholar]
- Bidelman, G.M.; Alain, C. Musical training orchestrates coordinated neuroplasticity in auditory brainstem and cortex to counteract age-related declines in categorical vowel perception. J. Neurosci. 2015, 35, 1240–1249. [Google Scholar] [CrossRef]
- Horvath, J.; Czigler, I.; Jacobsen, T.; Maess, B.; Schröger, E.; Winkler, I. MMN or no MMN: No magnitude deviance effect on the MMN amplitude. Psychophysiology 2008, 45, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Näätänen, R.; Alho, K. Mismatch negativity-the measure for central sound representation accuracy. Audiol. Neurotol. 1997, 2, 341–353. [Google Scholar] [CrossRef]
- Marie, C.; Magne, C.; Besson, M. Musicians and the metric structure of words. J. Cogn. Neurosci. 2011, 23, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Jacquier-Roux, M.; Valdois, S.; Zorman, M.O. Outil de Dépistage des Dyslexies; Cogni-Sciences: Grenoble, France, 2005. [Google Scholar]
- Maurer, U.; Bucher, K.; Brem, S.; Brandeis, D. Altered responses to tone and phoneme mismatch in kindergartners at familial dyslexia risk. Neuroreport 2003, 14, 2245–2250. [Google Scholar] [CrossRef]
- Hämäläinen, J.; Leppänen, P.H.T.; Torppa, M.; Müller, K.; Lyytinen, H. Detection of sound rise time by adults with dyslexia. Brain Lang. 2005, 94, 32–42. [Google Scholar] [CrossRef]
- Flaugnacco, E.; Lopez, L.; Terribili, C.; Montico, M.; Zoia, S.; Schön, D. Music Training Increases Phonological Awareness and Reading Skills in Developmental Dyslexia: A Randomized Control Trial. PLoS ONE 2015, 25, e0138715. [Google Scholar] [CrossRef] [PubMed]
- Przybylski, L.; Bedoin, N.; Krifi-Papoz, S.; Herbillon, V.; Roch, D.; Léculier, L.; Kotz, S.A.; Tillmann, B. Rhythmic auditory stimulation influences syntactic processing in children with developmental language disorders. Neuropsychology 2013, 27, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Van der Kleij, S.W.; Groen, M.A.; Segers, E.; Verhoeven, L. Enhanced semantic involvement during word recognition in children with dyslexia. J. Exp. Child Psychol. 2019, 178, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.B.; Ueki, K.; Oliveira, D.G.; Boggio, P.S.; Macedo, E.C. Early Stages of Sensory Processing, but Not Semantic Integration, Are Altered in Dyslexic Adults. Front. Psychol. 2016, 7, 430. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test | TD | DysM. | DysP. | F(2,40) | Post hoc comparisons |
|---|---|---|---|---|---|
| Chronological age (month) | 99.14 | 122.91 | 129.00 | 60.25, p < 0.001 | DysM. vs. TD: p < 0.001 DysP. vs. TD: p < 0.001 DysM. vs. DysP.: ns |
| Reading age a (month) | 92.44 | 87.24 | 86.27 | F < 1 | |
| Memory b Digit Span (/32) | 13.38 | 11.45 | 11.73 | 3.71, p = 0.03 | DysM. vs. TD: p < 0.05 DysP.vs TD: ns DysM. vs. DysP.: ns |
| Verbal IQ Similarities (/44) b | 15.67 | 17.73 | 18.36 | F < 1 | |
| Nonverbal IQ Symboles (/60) b | 16.38 | 20.73 | 16.82 | 2.16, p = 0.13 | |
| Nonverbal IQ Progressive Matrices (/36) c | 26.81 | 28.91 | 27.64 | F < 1 | |
| Phonologyd RAN (seconds) | 28.58 | 27.18 | 29.45 | F < 1 | |
| Phonology d Phoneme Deletion (/10) | 5.58 | 5.54 | 4.82 | F < 1 | |
| Phonology d Phoneme Fusion (/10) | 5.92 | 5.64 | 5.18 | F < 1 | |
| Phonology d Nonword repetition (/20) | 17.58 | 16.64 | 17.09 | F < 1 | |
| Reading irregular words (/20) | 7.91 | 4.45 | 6.90 | 2.24, p = 0.15 | |
| Reading regular words (/20) | 14.27 | 9.27 | 12.36 | 3.07, p = 0.08 | |
| Reading of pseudowords (/20) | 13.60 | 8.82 | 10.08 | 7.84, p < 0.03 | DysM. vs. TD: p < 0.02 DysP. vs. TD: p < 0.05 DysM. vs. DysP.: ns |
| Attention e Visual Attention Score (/45) | 17.08 | 16.00 | 18.09 | F < 1 | |
| Attention e Auditory Attention (/132) | 93.92 | 84.36 | 89.27 | F < 1 | |
| Attention e Orientation (/10) | 6.56 | 7.45 | 7.18 | F < 1 | |
| Attention e Visuomotor precision (/52) | 22.42 | 25.36 | 24.09 | F < 1 | |
| Attention e Arrows (/30) | 19.33 | 20.73 | 20.09 | F < 1 |
| Test | DysM. | DysP. | Main Effect Session F(1,20) | Post hoc Comparisons |
|---|---|---|---|---|
| Reading age a (month) | 90.18 | 90.45 | 18.50 p < 0.001 | T0 = 82.64 < T6 = 90.32 |
| Memory b Digit Span (/32) | 11.73 | 11.18 | F < 1 | |
| Verbal IQ Similarities (/44) b | 23.55 | 24.09 | 34.93 p < 0.001 | T0 = 18.04 < T6 = 23.82 |
| Nonverbal IQ Symboles (/60) b | 22.64 | 19.00 | F = 2.29 p < 0.15 | |
| Nonverbal IQ Progressive Matrices (/36) c | 30.36 | 27.64 | F < 1 | |
| Phonology d RAN (seconds) | 23.27 | 23.64 | 5.62 p < 0.03 | T0 = 32.44 > T6 = 26.40 |
| Phonology d Phoneme Deletion (/10) | 6.00 | 5.18 | F < 1 | |
| Phonology d Phoneme Fusion (/10) | 7.27 | 7.09 | 11.20 p < .003 | T0 = 5.41 < T6 = 7.18 |
| Phonology d Nonword repetition (/20) | 17.27 | 18.00 | F < 1 | |
| Reading irregular words (/20) | 6.82 | 8.45 | 8.52 p < 0.01 | T0 = 5.68 < T6 = 7.64 |
| Reading regular words (/20) | 11.18 | 12.00 | F < 1 | |
| Reading Pseudowords (/20) | 8.64 | 11.73 | F < 1 | |
| Attention e Visual Attention Score (/45) | 18.55 | 17.55 | F < 1 | |
| Attention e Auditory Attention (/132) | 106.18 | 98.91 | 5.76 p < 0.03 | T0 = 86.82 < T6 = 102.55 |
| Attention e Orientation (/10) | 8.18 | 8.18 | 10.87 p < 0.005 | T0 = 7.32 < T6 = 8.18 |
| Attention e Visuomotor precision (/52) | 26.91 | 26.73 | F = 2.13 p < 0.16 | |
| Attention e Arrows (/30) | 20.45 | 19.00 | F < 1 |
| VOT | Duration | Frequency | |
|---|---|---|---|
| Frontal | −1.61 µV | −1.12 µV | −1.85 µV |
| Central | −1.27 µV | −1.16 µV | −1.24 µV |
| Parietal | −0.64 µV | −0.69 µV | −0.59 µV |
| Frontal | Central | ||||
|---|---|---|---|---|---|
| Before | After | Before | After | ||
| Large Dev. | −1.96 | −2.99 | −1.90 | −2.60 | |
| DysMus | Small Dev. | −1.88 | −2.76 | −1.54 | −2.13 |
| Large Dev. | −1.44 | −1.37 | −1.66 | −1.38 | |
| DysPaint | Small Dev. | −0.86 | −1.19 | −0.96 | −1.51 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frey, A.; François, C.; Chobert, J.; Velay, J.-L.; Habib, M.; Besson, M. Music Training Positively Influences the Preattentive Perception of Voice Onset Time in Children with Dyslexia: A Longitudinal Study. Brain Sci. 2019, 9, 91. https://doi.org/10.3390/brainsci9040091
Frey A, François C, Chobert J, Velay J-L, Habib M, Besson M. Music Training Positively Influences the Preattentive Perception of Voice Onset Time in Children with Dyslexia: A Longitudinal Study. Brain Sciences. 2019; 9(4):91. https://doi.org/10.3390/brainsci9040091
Chicago/Turabian StyleFrey, Aline, Clément François, Julie Chobert, Jean-Luc Velay, Michel Habib, and Mireille Besson. 2019. "Music Training Positively Influences the Preattentive Perception of Voice Onset Time in Children with Dyslexia: A Longitudinal Study" Brain Sciences 9, no. 4: 91. https://doi.org/10.3390/brainsci9040091