Antifungal and Antiaflatoxigenic Activities of 1,8-Cineole and t-Cinnamaldehyde on Aspergillus flavus

1
School of Applied Biosciences, Kyungpook National University, Daegu 41566, Korea
2
Department of Pharmaceutical Engineering, Inje University, Gimhae 50834, Korea
*
Authors to whom correspondence should be addressed.
Appl. Sci. 2018, 8(9), 1655; https://doi.org/10.3390/app8091655
Submission received: 8 August 2018
/
Revised: 2 September 2018
/
Accepted: 12 September 2018
/
Published: 14 September 2018
(This article belongs to the Special Issue Pesticide and Emerging Organic Pollutant Analyses and their Ecotoxicological Evaluation in Soil and Aquatic Environments)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Featured Application
The specific application of the research aims to food safety by reduction of aflatoxin contamination.
Abstract
Aspergillus flavus and A. parsiticus produce aflatoxins that are highly toxic to mammals and birds. In this study, the inhibitory effects of 1,8-cineole and t-cinnamaldehyde were examined on the growth of Aspergillus flavus ATCC 22546 and aflatoxin production. 1,8-Cineole showed 50% inhibition of fungal growth at a concentration of 250 ppm, while t-cinnamaldehyde almost completely inhibited fungal growth at a concentration of 50 ppm. Furthermore, no fungal growth was observed when the growth medium was treated with 100 ppm t-cinnamaldehyde. 1,8-Cineole also exhibited 50% inhibition on the production of aflatoxin B1 and aflatoxin B2 at a concentration of 100 ppm, while the addition of 100 ppm t-cinnamaldehyde completely inhibited aflatoxin production. These antiaflatoxigenic activities were related to a dramatic downregulation of the expression of aflE and aflL by 1,8-cineole, but the mode of action for t-cinnamaldehyde was unclear. Collectively, our results suggest that both of the compounds are promising alternatives to the currently used disinfectant, propionic acid, for food and feedstuff preservation.
1. Introduction
Essential oils, which were obtained from several plants, have been considered to be potent fumigational insecticides that can be alternatives to the currently used manmade fumigants, including phosphine and ethyl formate, against stored product insects [1,2,3]. As some stored product insects have developed strong resistance to the currently used fumigants, such as phosphine, essential oils, having different target sites of insecticidal action and presumable selectivity for certain insects [4,5], might help to overcome the problem of resistance.
Recently, these essential oils have been introduced for their fungicidal activities for the control of agricultural pathogenic fungi, such as Aspergillus flavus, A. niger, A. ochraceus, Colletotrichum acutatum, and Fusarium graminearum [6,7,8]. Therefore, these plant oils can be used for dual purposes by controlling both insect pests and agricultural pathogenic microbes.
Mycotoxins produced by fungi spread steadily, contaminating agricultural foods and foodstuffs around the world, and there are several technologies that are available that use chemical and biological treatments and physiological applications to protect the foodstuffs from fungal toxins and to degrade the toxins before they reach the consumer [9,10]. Interestingly, some essential oils can suppress mycotoxin contamination by inhibiting fungal growth and toxin production. Eucalyptus essential oils and cinnamon essential oils are potent inhibitors of A. flavus growth and suppressors of aflatoxin production [11,12]. Their primary constituents are 1,8-cineole (or eucalyptol) and t-cinnamaldehyde, and their antifungal and antiaflatoxigenic activities have been reported previously [12,13]. A previous report demonstrated a dramatic reduction of the expression of some genes that are involved in the biosynthetic pathways of aflatoxin B1 by cinnamaldehyde [14]. However, no reports have described the mode of fungicidal action for the antiaflatoxigenic activity of 1,8-cineole. Additionally, the antifungal and antiaflatoxigenic properties of essential oils and their primary constituents need to be reconsidered before the introduction of these chemicals in agricultural industries, because the indefinite mode of antiaflatoxigenic action may be a major obstacle for their use in controlling A. flavus growth and aflatoxin production.
In the present study, 1,8-cineole and t-cinnamaldehyde were used to treat A. flavus and to examine the growth and suppression of aflatoxin production. Aflatoxin production was quantitatively determined while using high performance-liquid chromatography (HPLC) with a fluorescence detector. An RT-PCR technique was employed to understand how these chemicals decrease the expression of genes that are involved in the aflatoxin biosynthetic pathway, in order to comparatively measure the gene expression levels between the samples and the controls.
2. Materials and Methods
2.1. Chemicals and Microorganisms
1,8-Cineole and t-cinnamaldehyde were purchased from Sigma-Aldrich Co. (St. Louis, MO, USA). Malt extract agar (MEA; Difco, Franklin Lake, NJ, USA) was prepared for the subcultures of the aflatoxin-producing fungi. Potato dextrose broth (PDB; Difco, Franklin Lake, NJ, USA) was also prepared for the liquid culture media. The other chemicals used in this study were of reagent grade quality. In this study, A. flavus ATCC 22546 was purchased from the Korea Culture Center of Microorganisms (Seoul, Korea).
2.2. Measurement of the Antifungal Activities of the Test Compounds
Measurement of the antifungal activities of the two test compounds were conducted according to a previous report [15]. Briefly, spores, equivalent to 106 of A. flavus, were transferred into PDB liquid culture media (25 mL), and the test compounds were added to the liquid media.
As the fungal mycelia and the test compounds were presented together, the liquid culture was left for five days with shaking at 25 °C. Three replicates for each concentration were produced for the test compounds. After completion of the incubation, the mycelial growth rates (%) were obtained by measuring the mycelial and sclerotial dry weights of the experimental samples as compared to the control. The control groups contained only solvent, and no test compounds. The arithmetic means of the three replicates for each concentration were analyzed for any significant differences when compared to the control using one-way ANOVA, followed by Tukey’s test, and a p value < 0.05 was considered to be significant.
2.3. Aflatoxin Measurement Using High-Performance Liquid Chromatography (HPLC)
Measurements of the aflatoxin B- and G-types were conducted by HPLC, and the analytical conditions for HPLC were same as a previous report [16].
2.4. Isolation of Total RNA and Gene Expression Analysis by RT-qPCR
After allowing fungal growth for five days, A. flavus mycelia were carefully collected by filtration with a cell strainer (SPL Life Sciences Co., Ltd., Gyeonggi-do, Korea). The filtered mycelia were left in a mortar and ground with some liquid nitrogen. Total RNA was obtained from the filtered A. flavus mycelia by the addition of the QIAzol Lysis Reagent (QIAGEN Inc., Hilden, Germany). Extracted RNAs were first quantified while using a μDrop™ Plate incorporated with Multiskan™ Go Microplate Spectrophotometer (Thermo Fisher Scientific Inc., Ratastie, Finland). Then, the quantified RNA was examined qualitatively by electrophoresis using a 1% agarose gel with ethidium bromide. Complementary DNA (cDNA) was made using a Maxima First Strand cDNA Synthesis kit (Thermo Fisher Scientific Inc., Ratastie, Finland). For the procedure, extracted RNAs, which were equivalent to 2 μg, were used for cDNA synthesis.
cDNA (100 ng) was used for the analysis of gene expression with a Rotor-Gene SYBR Green PCR kit (QIAGEN Inc., Hilden, Germany) and RT-qPCR analysis. Specific primers for the genes were synthesized by Genotech (Daejeon, Korea) to determine the comparative levels of gene expression. The primers for the genes involved in aflatoxin biosynthesis (yap, aflC, aflD, aflE, aflG, aflK, aflL, erg28, aflR, aflS, and 18S rRNA) are listed in Table S1. The PCR conditions were exactly the same as those reported previously [15].
The RT-qPCR analysis was conducted three times for each sample. Differences in gene expression between the treated samples and the controls, after the addition of the test chemical, were calculated while using the 2−ΔΔCt method [17], followed by one-way ANOVA incorporated with Tukey’s test (p value < 0.05). The results were recalculated after normalization with 18S rRNA expression, and the gene expression comparisons were further analyzed.
3. Results
3.1. Effects of 1,8-Cineole and t-Cinnamaldehyde on A. flavus Growth and Aflatoxin Production
Figure 1 shows the antifungal activities of the two tested compounds on A. flavus growth. 1,8-Cineole showed a relatively lower antifungal activity against A. flavus than t-cinnamaldehyde. 1,8-Cineole demonstrated a 50% inhibition of A. flavus growth at a concentration of 250 ppm in the growth medium, and at 50 ppm, it did not exhibit any inhibitory effects on the growth.
t-Cinnamaldehyde almost completely inhibited A. flavus growth at a concentration of 100 ppm in the growth medium. Furthermore, at a concentration of 50 ppm, the fungal growth rate reached about 5% of that of the control. However, treatment with 10 ppm t-cinnamaldehyde showed no inhibition of A. flavus growth, as the fungal growth rate was over 100% when compared to the control.
Figure 2 shows the inhibitory effects of 1,8-cineole on aflatoxin B1 (AFB1) production in A. flavus. No AFB1 production was measured in A. flavus following treatment with 50 ppm of 1,8-cineole. However, the AFB1 production rate decreased to about 50% after treatment with 1,8-cineole at the concentration of 100 ppm. For AFB2 production, 1,8-cineole suppressed the production to less than 50% at the concentration of 100 ppm.
Figure 3 illustrates the inhibitory effects of t-cinnamaldehyde on AFB1 production by A. flavus. AFB1 production by A. flavus was completely inhibited by treatment with 100 ppm of t-cinnamaldehyde. However, the AFB1 production rate was about 70% following treatment with t-cinnamaldehyde at a concentration of 10 ppm. For AFB2 production, t-cinnamaldehyde totally inhibited production at the concentration of 100 ppm, while it lost its inhibitory effect at the concentration of 10 ppm.
3.2. RT-qPCR Analysis of the Expression of Genes Involved in Aflatoxin Biosynthesis
After the addition of the two test compounds to the A. flavus growth medium, the expression levels of genes that were involved in the AFB1 biosynthetic pathway and ergosterol were examined using RT-qPCR with nine gene primers (Figure 4 and Figure 5). The examined genes express polyketide synthase (aflC), reductase (aflD), norsolorinic acid (NOR) reductase (aflE), P450 monooxygenase (aflK), and versicolorin B (VERB) synthase (aflL). Another three genes, yap, aflR, and aflS, are transcription factors for aflatoxin biosynthesis, and 18S rRNA was used to normalize the gene expression. A gene, erg28, was used to determine the expression of the ergosterol biosynthetic protein 28.
Following treatment with 1,8-cineole, the expression levels of two genes, aflE and aflL, were strongly downregulated from the two treatment concentrations (Figure 4). These two genes express NOR-reductase and VERB synthase. Interestingly, the transcription factors aflR and aflS increased slightly or were unchanged, respectively. The expression of other genes (aflC, aflD, aflG, and aflK) that are involved in aflatoxin biosynthesis exhibited no change at all of the treatment concentrations. However, erg28 expression was about two-fold higher in the 1,8-cineole-treated samples than the control.
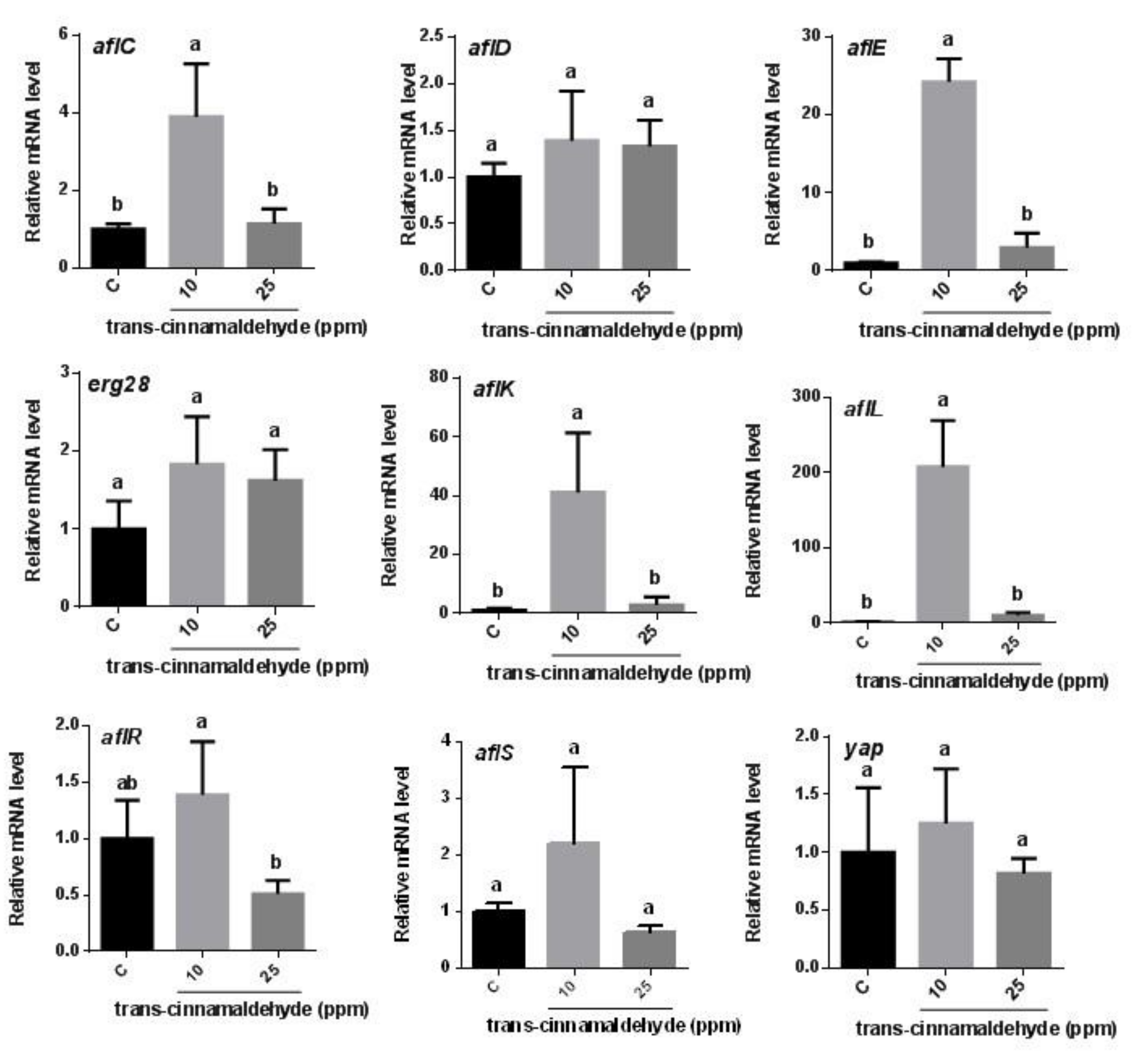
For t-cinnamaldehyde, no genes involved in aflatoxin biosynthesis were differentially regulated in comparison to the control, except for the aflR gene, a transcription factor for aflatoxin biosynthesis (Figure 5). At the treatment concentration of 10 ppm, the expression levels of aflL, aflK, and aflE were upregulated by about 200-fold, 40-fold, and 25-fold, respectively, in the t-cinnamaldehyde-treated A. flavus when compared to the control. However, there was no effect on erg28 gene expression. Therefore, only the aflR gene was expressed differently, indicating that this gene might play an important role in the reduction of aflatoxin biosynthesis during t-cinnamaldehyde treatment.
4. Discussion
Agricultural foods and feedstuffs need to be safe until the customer can consume them properly, and thus, fungal infection and mycotoxin contamination during the storage and/or transport should be periodically monitored [18]. Additionally, effective control strategies for fungal infection and toxin production should be organized for the right place and time [19]. Chemical treatment has been seriously considered as an effective method to reduce fungal infection and toxin production in agricultural foods and feedstuffs [9,15,20].
Recently, Moon et al. [21] reported that five food additives might be used to protect foods and feedstuffs from aflatoxin contamination. These authors demonstrated that benzoic acid up to 0.1% exhibited potent antifungal and antiaflatoxigenic activities against A. flavus at concentration levels below the legal limit for standard use; it can be used as an alternative food additive to propionic acids at those levels, because propionic acid, the currently used disinfectant, cannot be utilized due to the limitations of its standard use.
On the other hand, naturally occurring bioactive compounds, including plant essential oils and their major constituents, have been introduced, due to their strong antifungal activities [7,8,11,22]. Among them, essential oil from Mentha longifolia, which contained 2.2% of 1,8-cineole, showed strong antifungal and antiaflatoxigenic activities towards A. flavus [22]. 1,8-Cineole showed moderate antifungal activity against A. flavus (Figure 1A) at a treatment concentration of up to 250 ppm. However, t-cinnamaldehyde exhibited strong antifungal activity when compared to 1,8-cineole (Figure 1B). In addition to the antifungal activity, the antiaflatoxigenic activity of t-cinnamaldehyde on aflatoxin production was higher than that of 1,8-cineole (Figure 2 and Figure 3). AFB1 and AFB2 decreased with increasing concentrations in the chemical-treated growth medium.
In this study, 1,8-cineole showed antifungal and antiaflatoxigenic activities towards A. flavus growth and AFB1 production at a concentration of 250 ppm. Additionally, a recent report examined 1,8-cineole and showed that its minimal inhibitory concentration (MIC) and minimal fungicidal concentration (MFC) were lower than those of the currently used fungicide Nystatin [13]. They used 1,8-cineole at a concentration of 0.918 mg/mL (about 918 ppm) in the growth medium. Therefore, 1,8-cineole completely inhibited A. flavus growth, at least at a concentration of 918 ppm. With this antifungal activity, 1,8-cineole strongly inhibited AFB1 production without non-phytotoxic effects on chickpea seed germination [13]. However, in the report, they did not explain how 1,8-cineole possessed antiaflatoxignic activity. In our study, 1,8-cineole strongly inhibited aflE and aflL gene expression in A. flavus (Figure 4). These two genes are responsible for the expression of norsolorinic acid reductase and cytochrome P450 monooxygenase/desaturase. Norsolorinic acid (NOR) reductase is responsible for the conversion of NOR to averantin [23]. This enzyme functions in aflatoxin biosynthesis after polyketide synthase [23], which is at a very early part of the biosynthetic pathway. Therefore, 1,8-cineole inhibits the expression of the first enzyme to suppress aflatoxin production.
Moreover, 1,8-cineole downregulated the aflL gene, which encodes cytochrome P450 monooxygenase/desaturase and it presumably participates in the conversion of versicolorin B (VERB) to VERA [23]. This enzyme plays a key role in the separation of the AFB1 subtype from the AFB2 subtype. With the addition of 1,8-cineole, the AFB1 production route will be blocked and the biosynthetic production of AFB2 will be directly enhanced. This is the first reported method for the mode of the antiaflatoxigenic activity of 1,8-cineole.
Alternatively, t-cinnamaldehyde treatment showed strong inhibition of fungal growth and aflatoxin production, but the mode of its antiaflatoxigenic activity was not clear from our study (Figure 5). At a low concentration of t-cinnamaldehyde, some genes were dramatically upregulated, even though the aflatoxin production decreased at the same concentration. Rather, Sun et al. [24] indicated that cinnamaldehyde inhibited fungal growth and AFB1 production by oxidative stress. This reduction of AFB1 production is not related to the downregulation of the biosynthetic genes, but instead, to the significant inhibition of some antioxidant enzymes, including total superoxide dismutase.
However, Liang et al. [14] determined changes in the expression of genes that are involved in aflatoxin biosynthesis, and cinnamaldehyde downregulated the expression of five genes of the tested genes, including aflR, aflT, aflD, aflM, and aflP. They were treating with cinnamladehyde at the concentration of 0.40 mmol/L, which is equivalent to about 132 ppm. At this concentration, cinnamaldehyde suppressed aflM gene expression 5963-fold, followed by aflP, aflR, aflD, and aflT with the average folds of suppression of 55, 18, 6.5, and 5.8, respectively. In our study, we could not collect total RNA from the fungi, because they did not survive properly at the concentration range of 50 to 100 ppm of cinnamaldehyde (Figure 1). It is likely that A. flavus, which was used in this study, may be differently affected by the action of cinnamaldehyde when compared to the fungi used in the study by Liang et al. [14].
5. Conclusions
Two monoterpenoids, 1,8-cineole and t-cinnamaldehyde, which are present in eucalyptus and cinnamon essential oils, were examined for their inhibitory effects on fungal growth and aflatoxin production in A. flavus. 1,8-Cineole possessed antifungal and antiaflatoxigenic activities at the concentration of 250 ppm, while t-cinnamaldehyde at the concentration of 50 ppm showed potent inhibitory activities. However, a clear mode for the antiaflatoxigenic activity of 1,8-cineole was found from the dramatic downregulation of aflE and aflL gene expression, whereas t-cinnamaldehyde only demonstrated a slight inhibition of aflR gene expression. Taken together, our results show that these two monoterpenes can be considered to be promising preservatives in foods and feedstuffs to prevent A. flavus infection and aflatoxin contamination.
Supplementary Materials
The following are available online at https://www.mdpi.com/2076-3417/8/9/1655/s1, Table S1: Primers used in this study for RT-qPCR. They are related to the genes involved in aflatoxin and ergosterol production in Aspergillus flavus.
Author Contributions
S.-E.L. and K.K. conceived and designed all of the experiments; H.-M.K. and H.K. performed the experiments; H.-M.K. and S.-E.L. analyzed the results; and S.-E.L. wrote the paper.
Funding
This research was funded by Inje University to Kyeongsoon Kim.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lee, H.K.; Lee, S. Toxicities of active constituent isolated from Thymus vulgaris flowers and its structural derivatives against Tribolium castaneum (Herbst). Appl. Biol. Chem. 2016, 59, 821–826. [Google Scholar] [CrossRef]
- Song, J.E.; Kim, J.M.; Lee, N.H.; Yang, J.E.; Lee, H.S. Acaricidal and insecticidal activities of essential oils against a stored–food mite and stored-grain insects. J. Food Prot. 2016, 79, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.J.; Lee, S.G.; Lee, H.S. Acaricidal and insecticidal activities of essential oils of Cinnamomum zeylanicum barks cultivated from France and India against Dermatophagoides spp., Tyrophagus putrescentiae and Ricania sp. Appl. Biol. Chem. 2017, 60, 259–264. [Google Scholar] [CrossRef]
- Enan, E. Insecticidal activity of essential oils: Octopaminergic sites of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2001, 130, 325–337. [Google Scholar] [CrossRef]
- Park, B.S.; Lee, B.H.; Kim, T.W.; Ren, Y.L.; Lee, S.E. Proteomic evaluation of adults of rhyzopertha dominica resistant to phosphine. Environ. Toxicol. Pharmacol. 2008, 25, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Paster, N.; Menasherov, M.; Ravid, U.; Juven, B. Antifungal activity of oregano and thyme essential oils applied as fumigants against fungi attacking stored grain. J. Food Prot. 1995, 58, 81–85. [Google Scholar] [CrossRef]
- He, J.; Wu, D.; Zhang, Q.; Chen, H.; Li, H.; Han, Q.; Lai, X.; Wang, H.; Wu, Y.; Yuan, J.; et al. Efficacy and mechanism of cinnamon essential oil on inhibition of Colletotrichum acutatum isolated from ‘Hongyang’ kiwifruit. Front. Microbiol. 2018, 9, 1288. [Google Scholar] [CrossRef] [PubMed]
- Kalagatur, N.K.; Nirmal Ghosh, O.S.; Sundararaj, N.; Mudili, V. Antifungal activity of chitosan nanoparticles encapsulated with Cymbopogon martinii essential oil on plant pathogenic fungi Fusarium graminearum. Front. Pharmacol. 2018, 9, 610. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Lee, B.H.; Moon, Y.S.; Kim, K.; Lee, H.S.; Lee, S.E. Antifungal and antiaflatoxigenic effects of a fumigant, ethanedinitrile, on Aspergillus flavus. Appl. Biol. Chem. 2017, 60, 473–476. [Google Scholar] [CrossRef]
- Zhu, Y.; Hassan, Y.I.; Lepp, D.; Shao, S.; Zhou, T. Strategies and methodologies for developing detoxification systems to mitigate mycotoxins. Toxins 2017, 9, 130. [Google Scholar] [CrossRef] [PubMed]
- Bluma, R.; Amaiden, M.R.; Etcheverry, M. Screening of argentine plant extracts: impact on growth parameters and aflatoxin B1 accumulation by Aspergillus section Flavi. Int. J. Food Microbiol. 2008, 122, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Mateo, E.M.; Gómez, J.V.; Domínquez, I.; Gimeno-Adelantado, J.V.; Mateo-Castro, R.; Gavara, R.; Jiménez, M. Impact of bioactive packaging systems based on EVOH films and essential oils in the control of aflatoxigenic fungi and aflatoxin production in maize. Int. J. Food Microbiol. 2017, 254, 36–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, R.; Singh, P.; Prakash, B.; Dubey, N.K. Antifungal, aflatoxin inhibition and antioxidant activity of Callistemon lanceolatus (Sm.) sweet essential oil and its major component 1,8-cineole against fungal isolates from chickpea seeds. Food Control 2012, 25, 27–33. [Google Scholar] [CrossRef]
- Liang, D.; Xing, F.; Selvaraj, J.N.; Liu, X.; Wang, L.; Hua, H.; Zhou, L.; Zhao, Y.; Wang, Y.; Liu, Y. Inhibitory effect of cinnamaldehyde, citral, and eugenol on aflatoxin biosynthetic gene expression and aflatoxin B1 biosynthesis in Aspergillus flavus. J. Food Sci. 2015, 80, M2917–M2924. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.S.; Choi, W.S.; Park, E.S.; Bae, I.K.; Choi, S.D.; Paek, O.; Kim, S.H.; Chun, H.S.; Lee, S.E. Antifungal and antiaflatoxigenic methylenedioxy-containing compounds and piperine-like synthetic compounds. Toxins 2016, 8, 240. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Mahoney, N.; Campbell, B.C. Inhibition of aflatoxin B1 biosynthesis by piperlongumine isolated from Piper longum L. J. Microbiol. Biotechnol. 2002, 12, 679–682. [Google Scholar]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2ˆ(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Ma, R.; Zhang, L.; Liu, M.; Su, Y.T.; Xie, W.M.; Zhang, N.Y.; Dai, J.F.; Wang, Y.; Rajput, S.A.; Qi, D.S.; et al. Individual and combined occurrence of mycotoxins in feed ingredients and complete feeds in China. Toxins 2018, 10, 113. [Google Scholar] [CrossRef] [PubMed]
- Tola, M.; Kebede, B. Occurrence, importance and control of mycotoxins: A review. Cogent Food Agric. 2016, 2, 1191103. [Google Scholar] [CrossRef]
- Moon, Y.S.; Kim, L.; Chun, H.S.; Lee, S.E. 4-Hydroxy-7-methyl-3-phenylcoumarin suppress aflatoxin biosynthesis via downregulation of aflK expressing versicolorin B synthase in Aspergillus flavus. Molecules 2017, 22, 712. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.S.; Kim, H.M.; Chun, H.S.; Lee, S.E. Organic acids suppress aflatoxin production via lowering expression of aflatoxin biosynthesis-related genes in Aspergillus flavus. Food Control 2018, 88, 207–216. [Google Scholar] [CrossRef]
- Dehghanpour-Farashah, A.; Taheri, P. Antifungal and antiaflatoxigenic effects of Mentha longifolia essential oil against Aspergillus flavus. Int. J. New Technol. Res. 2016, 2, 30–39. [Google Scholar]
- Yu, J. Current understanding on aflatoxin biosynthesis and future perspective in reducing aflatoxin contamination. Toxins 2012, 4, 1024–1057. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Shang, B.; Wang, L.; Lu, Z.; Liu, Y. Cinnamaldehyde inhibits fungal growth and aflatoxin B1 biosynthesis by modulating the oxidative stress response of Aspergillus flavus. Appl. Microbiol. Biotechnol. 2016, 100, 1355–1364. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of (A) 1,8-cineole and (B) t-cinnamaldehyde on the growth of Aspergillus flavus. Different letters above the same column indicate significant differences among the treatments. Data are presented as the mean ± SD.
Figure 1.
Effects of (A) 1,8-cineole and (B) t-cinnamaldehyde on the growth of Aspergillus flavus. Different letters above the same column indicate significant differences among the treatments. Data are presented as the mean ± SD.

Figure 2.
Effects of 1,8-cineole on aflatoxin production by Aspergillus flavus. AFB1, aflatoxin B1; AFB2, aflatoxin B2. Different letters above the same column indicate significant differences among the treatments. Data are presented as the mean ± SD.
Figure 2.
Effects of 1,8-cineole on aflatoxin production by Aspergillus flavus. AFB1, aflatoxin B1; AFB2, aflatoxin B2. Different letters above the same column indicate significant differences among the treatments. Data are presented as the mean ± SD.

Figure 3.
Effects of t-cinnamaldehyde on aflatoxin production by Aspergillus flavus. AFB1, aflatoxin B1; AFB2, aflatoxin B2. Different letters above the same column indicate significant differences among the treatments. Data are presented as the mean ± SD.
Figure 3.
Effects of t-cinnamaldehyde on aflatoxin production by Aspergillus flavus. AFB1, aflatoxin B1; AFB2, aflatoxin B2. Different letters above the same column indicate significant differences among the treatments. Data are presented as the mean ± SD.

Figure 4.
Expression of genes (aflC, aflD, aflE, aflK, aflL, aflR, aflS, and yap) involved in aflatoxin biosynthesis in Aspergillus flavus treated with 1,8-cineole. An ergosterol biosynthetic gene, erg28, was comparatively measured after treatment with 1,8-cineole. Different letters above the same column indicate significant differences from the control group (p < 0.05).
Figure 4.
Expression of genes (aflC, aflD, aflE, aflK, aflL, aflR, aflS, and yap) involved in aflatoxin biosynthesis in Aspergillus flavus treated with 1,8-cineole. An ergosterol biosynthetic gene, erg28, was comparatively measured after treatment with 1,8-cineole. Different letters above the same column indicate significant differences from the control group (p < 0.05).

Figure 5.
Expression of genes (aflC, aflD, aflE, aflK, aflL, aflO, aflP, aflR, aflS, and yap) involved in aflatoxin biosynthesis in Aspergillus flavus treated with t-cinnamaldehyde. An ergosterol biosynthetic gene, erg28, was comparatively measured after treatment with t-cinnamaldehyde. Different letters above the same column indicate significant differences from the control group (p < 0.05).
Figure 5.
Expression of genes (aflC, aflD, aflE, aflK, aflL, aflO, aflP, aflR, aflS, and yap) involved in aflatoxin biosynthesis in Aspergillus flavus treated with t-cinnamaldehyde. An ergosterol biosynthetic gene, erg28, was comparatively measured after treatment with t-cinnamaldehyde. Different letters above the same column indicate significant differences from the control group (p < 0.05).

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, H.-M.; Kwon, H.; Kim, K.; Lee, S.-E. Antifungal and Antiaflatoxigenic Activities of 1,8-Cineole and t-Cinnamaldehyde on Aspergillus flavus. Appl. Sci. 2018, 8, 1655. https://doi.org/10.3390/app8091655
AMA Style
Kim H-M, Kwon H, Kim K, Lee S-E. Antifungal and Antiaflatoxigenic Activities of 1,8-Cineole and t-Cinnamaldehyde on Aspergillus flavus. Applied Sciences. 2018; 8(9):1655. https://doi.org/10.3390/app8091655
Chicago/Turabian StyleKim, Hyeong-Mi, Hyunwoo Kwon, Kyeongsoon Kim, and Sung-Eun Lee. 2018. "Antifungal and Antiaflatoxigenic Activities of 1,8-Cineole and t-Cinnamaldehyde on Aspergillus flavus" Applied Sciences 8, no. 9: 1655. https://doi.org/10.3390/app8091655
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.