The Construction of an Engineered Bacterial Strain and Its Application in Accumulating Mercury from Wastewater

1
Haikou Experimental Station, Chinese Academy of Tropical Agricultural Sciences, Haikou 570102, Hainan, China
2
The Key Lab of Hainan Banana Genetics and Breeding, Haikou 570102, Hainan, China
3
School of Forestry, Henan University of Science and Technology, Luoyang 471023, Henan, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Appl. Sci. 2018, 8(9), 1572; https://doi.org/10.3390/app8091572
Submission received: 7 August 2018
/
Revised: 27 August 2018
/
Accepted: 3 September 2018
/
Published: 6 September 2018
(This article belongs to the Section Applied Biosciences and Bioengineering)
Abstract
:To remove organic and inorganic mercury from wastewater, an engineered bacterial strain, BL21-7, was constructed that contained the artificial operon P16S-g10-merT-merP-merB1-merB2-ppk-rpsT. For BL21-7, the minimum inhibitory concentrations of mercuric chloride, methylmercury chloride and phenylmercury chloride in Luria-Bertani (LB) medium were 100 µmol/L, 60 µmol/L and 80 µmol/L, respectively. After being cultured in three media (liquid LB containing 80 µmol/L mercuric chloride, 40 µmol/L methylmercury chloride or 60 µmol/L phenylmercury chloride) for 72 h, the engineered bacteria accumulated up to 70.5 ± 1.5 µmol/L, 33.5 ± 3.2 µmol/L and 45.3 ± 3.7 µmol/L of mercury, respectively. In the presence of 10 µmol/L Cd2+, 10 µmol/L Pb2+ or 10 µmol/L Cu2+, the accumulation of mercurial derivatives by BL21-7 was not affected. BL21-7 could accumulate mercury well in media with pH values ranging from 5 to 8 and it could work well at temperatures from 25 °C to 37 °C. After BL21-7 was added to wastewater and cultured for 24 h, approximately 43.7% of the Hg in the wastewater was removed.
1. Introduction
Mercury (Hg) pollution threatens public health. In nature, Hg exists in three forms: elemental (Hg(0)), inorganic (Hg2+ or Hg+) and organic forms (R-Hg+ or R-Hg-X, where “R” is methyl or phenyl and “X” is acetate) [1]. Elemental Hg is volatile. Inorganic Hg commonly exists as mercuric sulfide and is used as a coloring agent in tattoos and paints. All the forms are hazardous but the extent of toxicity varies among different forms of Hg. Organic Hg is the most toxic and elemental is the least toxic. Elemental Hg can potentially become an inhalation hazard. Organic Hg can impair neurological development in fetuses and young children and can damage the central nervous system of adults [2,3]. Exposure to inorganic Hg may damage the gastrointestinal tract, the nervous system and the kidneys [4]. The Hg in agricultural soil can enter the human body through crops and Hg mainly comes from irrigated-polluted water released from factories [5]. The best resolution for this problem is removing Hg from polluted water before it is discharged from factories.
Engineered bacterial strains can be used to remove Hg from polluted water. After E. coli JM109 cells, transformed with the mouse metallothionein gene mt-1 were cultured in LB medium containing 120 µmol/L mercuric chloride (HgCl2) for 72 and 120 h, the bacteria accumulated 51.6 µmol/L Hg and 1002 µmol/L Hg, respectively [6]. However, wild-type untransformed E. coli JM109 cells (the control) could not grow at all in LB medium containing 120 µmol/L HgCl2 [6]. However, it is unclear whether the strain accumulated organic Hg, a main pollution component in wastewater [6]. Bacteria harboring pMRA17 containing the Pseudomonas K-62 genes merR, merT, merA and merB1 had a higher methylmercury tolerance than the control [7]. However, their Hg accumulation was achieved by volatilizing Hg(0) into the air, which causes public anxiety. The same problem exists in the Cupriavidus metallidurans strain MSR33, which harbors two large plasmids (pMOL28 and pMOL30) carrying genetic determinants for heavy metal resistance [8]. Although bacteria harboring pMRB01, which contains Pseudomonas K-62 genes merR, merB2 and merD, had a higher tolerance to HgCl2 than the control, they were hypersensitive to phenylmercury [7]. E. coli harboring pMKB18, which contains Pseudomonas K-62 genes merR, merT, merP and merB1 as well as the Klebsiella aerogenes polyphosphate kinase gene ppk could remove 55% of the Hg2+ and 31% of the C6H5Hg+ when it was cultured in medium containing 80 µmol/L Hg2+ or 80 µmol/L C6H5Hg+ for 20 h [9] but whether it can accumulate CH3Hg+ has not been reported. CM111 and HG3, two bacteria isolated from Hg-polluted rivers or sand, also had high minimum inhibitory concentration (MIC) values of Hg2+ [10]. Although the tolerance of CM111 and HG3 to Hg2+ was not due to Hg(0) volatilization [10], the detailed mechanisms of these two strains’ tolerance to Hg2+ are unclear. Whether these strains can remove organic Hg from a medium is also unknown.
The aim of this study was to construct an engineered bacterial strain for accumulating Hg from liquid medium. This bacterial strain should meet the following criteria: (1) it can accumulate both mercuric ions and organic mercury; (2) it has a clear genetic background; (3) its mechanisms for mercury accumulation are definite; and (4) it can keep mercury accumulated in cells. To achieve this aim, an artificial operon, P16S-g10-merT-merP-merB1-merB2-ppk-rpsT and a bacterial strain, BL21-7, were constructed. The role of the genes in the artificial operon and the mechanism by which the bacteria accumulated mercury were also studied.
2. Materials and Methods
2.1. Strains and Chemicals
A genomic DNA extraction kit and 2× Pfu MasterMix were purchased from ComWin Biotech (Beijing, China). An RNeasy Mini kit was obtained from Qiagen (Venlo, The Netherlands) and an AccuScript cDNA kit was acquired from Stratagene. An In-Fusion HD Cloning Kit, T4 DNA ligase, HgCl2, methylmercury chloride (MMC), phenylmercury chloride (PMC) and other chemicals were purchased from TAKARA (Dalian, China). Enterobacter aerogenes, E. coli DH10B, E. coli BL21 (DE3), pET28a plasmid and Tobacco NC89 seeds were stocked in our laboratory.
2.2. Constructing the Engineered Bacterial Strains
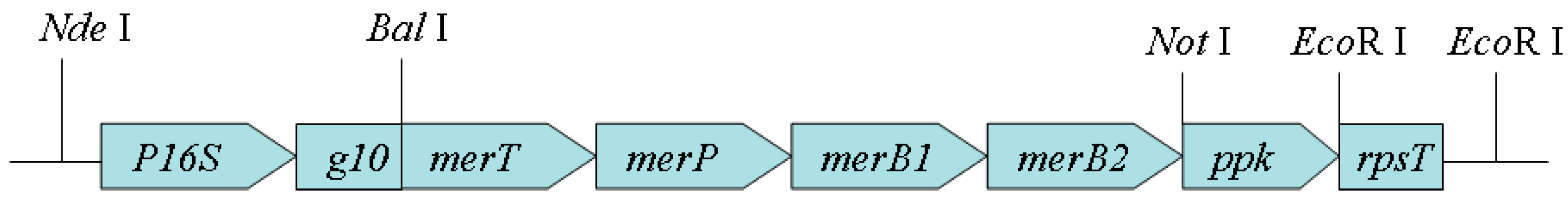
To construct the artificial operon P16S-g10-merT-merP-merB1-merB2-ppk-rpsT, a series of primers were designed and synthesized (Table 1). Based on the sequences of plasmids pMR26 (GenBank accession number: D83080.2) and pMR28 (GenBank accession number: AB013925.1), the artificial operon merT-merP-merB1-merB2 was designed and synthesized by TAKARA (Dalian, China). The synthesized DNA fragment was inserted into the region between the Nde I and Not I restriction sites of plasmid pET28a using an In-Fusion HD Cloning Kit (Clontech, code number: 639633). The resulting plasmid was named p1.
The ppk gene was amplified from Enterobacter aerogenes genomic DNA using 2× Pfu MasterMix (ComWin Biotech, code number: CW0686A). The primers used were PPPKF and PPPKR. The amplified fragment was inserted into the area between the Not I and Xho I restriction sites of plasmid p1. The resulting plasmid was named p2 (Table 1). According to published DNA sequences (GenBank accession number: Z00044.2, reference) [11], P1F and P1R were synthesized and 1 mmol/L P1F and 1 mmol/L P1R were mixed with 25 µL of 2× Pfu MasterMix. The volume of reaction was 500 µL. The mixture was annealed in a PCR amplifier. The annealed product, the strong constitutive promoter P16S [12,13,14] and the translation enhancer g10 [11], was inserted into plasmid pET28a. The resulting plasmid was named p3 (Table 1). Primers P2F and P2R were synthesized and DNA fragment merT-merP-merB1-merB2-ppk was amplified using p2 as a template. The amplified fragment was inserted into the region between the Bal I and EcoR I sites of p3. The resulting plasmid was named p4 (Table 1).
Primers P3F and P3R were synthesized according to a previously published sequence (GenBank accession number: Z00044.2). Using P3F and P3R as primers, rpsT, a strong terminator that can enhance transcript stability by forming a secondary structure at the 3′ end of the mRNA [15], was amplified from tobacco chloroplast DNA. The amplified rpsT was inserted into the EcoR I site of p4. The resulting plasmid, in which the transcription direction of rpsT was consistent with that of merT-merP-merB1-merB2-ppk, was named p5 (Table 1). The E. coli BL21 strain harboring p5 was named BL21-7. The sequence of the artificial operon P16S-g10-merT-merP-merB1-merB2-ppk-rpsT is shown in Supplementary Materials S1. To characterize the roles of the genes in the artificial operon P16S-g10-merT-merP-merB1-merB2-ppk-rpsT and the transcription of the operon, a series of artificial operons were constructed (Table 2). The detailed procedures can be found in Supplementary Materials S2.
2.3. Real-Time Reverse-Transcription PCR
The E. coli BL21 strain transformed with empty plasmid pET28a was named BL21-0. BL21-0, BL21-1, BL21-2 and BL21-7 were cultured in Luria-Bertani (LB) liquid medium for 16 h at 37 °C. Bacterial total RNA was extracted from 1 mL of these cultures using a Column Bacterial RNA Extraction kit (ABiGen, Beijing, China). Reverse transcription was performed using a PrimeScript RT Reagent Kit (TAKARA, Dalian, China). This kit contains a DNA eraser that can remove genomic DNA from RNA samples without requiring purification. The cDNA was analyzed using StepOnePlus Real-Time PCR Systems (Applied Biosystems, Foster City, CA, USA) and a two-step amplification program was used for the analysis. SYBR Premix Ex Taq II (TAKARA, Dalian, China) was used for real-time PCR. To prevent an effect of the ppk gene of the bacterium itself, ppk-rpsT was used as a candidate for detection. The sequences of the primers for ppk-rpsT overlapped both ppk and rpsT. The forward primer PPRF (5′-AACTTTATCGAAAACCCGTACCGTC-3′) was within the region of ppk. The reverse primer PPRR (5′-ATTTATTTAATCCATAATGGATTCA-3′) was within the region of rpsT. The primers were diluted in a series from 1 × 107 copies/µL to 1 × 102 copies/µL to obtain a quantitative standard curve. Experiments were repeated four times and the average values were used.
2.4. Measuring Bacterial Hg Tolerance
A series of tubes were filled with 5 mL of fresh LB liquid medium. A stock solution of HgCl2 (0.1 mol/L) was initially prepared and a series of HgCl2 stock solutions were added to the medium to final concentrations of 0, 5, 10, 20 and 40 µmol/L. Another 5 mL of fresh LB liquid medium was placed in a different series of tubes. An MMC stock solution (1 mmol/L) and a PMC stock solution (1 mmol/L) were also prepared. A series of MMC stock solutions were added to the medium to final concentrations of 0, 50, 100, 200 and 400 nmol/L. A series of PMC stock solutions were also added to the medium to final concentrations of 0, 50, 100, 200 and 400 nmol/L. After BL21-0 and BL21-7 were cultured in LB at 37 °C for 16 h, the cultures were placed in LB liquid medium containing organic Hg or HgCl2 at an OD600 = 0.01. The tubes were then placed in a shaker and cultured at 37 °C for 5 h. IPTG (Isopropyl β-d-Thiogalactoside) was then added to create a final concentration of 0.8 mmol/L. The cultures were then shaken at 37 °C for an additional 16–20 h. The light absorption values were measured at 600 nm. Each sample was measured with three replicates and the average value was used.
2.5. Measuring Minimum Inhibitory Concentration (MIC) Values
The bacterial MICs of the mercuric varieties and other metals were measured in two ways. The first was using the method described in “measuring bacterial Hg tolerance” but the final concentrations of the mercuric varieties in the medium were 3, 5, 10, 20, 40, 60, 80, 100 µmol/L. The minimum concentration of a certain mercuric variety in which bacteria cannot grow was used as the bacteria’s MIC of this mercuric variety. LB medium contains sufficient nutrients for bacterial growth; therefore, MIC data measured in LB medium may not reflect the real tolerance of the engineered bacteria. To characterize the MICs of BL21-7 when the nutrition in the medium was poor, a second method was also used to measure the bacterial MICs according to a previously published paper [8]. LB (10 g/L Bacto Tryptone, 5 g/L yeast extract, 10 g/L NaCl, 7 g/L agar, pH 7.0) plates and poor broth nutrient medium (PB) (10 g/L Bacto Tryptone, 5 g/L NaCl, 7 g/L agar, pH 7.0) plates [10] were initially prepared. One clone of BL21-7 on an LB plate was cultured in 5 mL of LB liquid medium containing 50 mg/L kanamycin for 16 h. The 10 µL of this culture grown for 16 h in LB liquid medium was plated on LB plates supplemented with a series of concentrations of each metal variety (the final concentrations of the mercuric varieties in the medium were 3, 5, 10, 20, 40, 60, 80 and 100 µmol/L). Another 10 µL of culture grown for 16 h in LB liquid medium was plated on PB plates supplemented with a series of concentrations of each metal variety (the final concentrations of the mercuric varieties in the medium were 3, 5, 10, 20, 40, 60, 80 and 100 µmol/L). The plates were incubated at 37 °C for 48 h. Then, the clones on the plates were counted and the minimum concentration of a certain mercuric variety on which the bacteria could not grow was used as the bacteria’s MIC of this mercuric variety.
2.6. Hg Content Determination
The Hg content in bacteria was measured following a previously published method [6]. Reagent blanks and spiked control samples were treated as described. The samples were analyzed in an Analyst 200 Perkin Elmer Spectrometer (Perkin Elmer, Waltham, MA, USA) with an MHS-15 Hg-Hydride System (Perkin Elmer, Waltham, MA, USA). Matrix spiked controls were produced by adding 100 ng/mL Hg to E. coli cell pellets recovered by centrifugation from 5 mL of LB cultures grown for 16 h without Hg. All samples were measured three times and the average values were used. The Hg accumulation (in μM) was calculated by multiplying the ng/mL (μg/L) value obtained from the instrument by the appropriate dilution factor used to keep the sample within the standard curve range and was then divided by the molecular mass of Hg (200.59 g) in μmol.
2.7. Hg Content Determination Inside Bacterial Cells
One clone of BL21-7 on an LB plate was cultured in 5 mL of LB liquid medium containing 50 mg/L kanamycin for 16 h. Then, 2 mL of this culture was transferred to two groups of conical flasks containing 200 mL of LB liquid medium containing 80 µmol/L HgCl2 and was cultured for 72 h. The bacteria in one group were collected by centrifugation. The Hg content was measured as described in the materials and methods and was named K1. The bacteria in the other group were also centrifuged. The supernatant was discarded and the pellet was thoroughly re-suspended in PBS (phosphate buffer saline) buffer. The mixture was centrifuged and the supernatant was discarded. The pellet was thoroughly re-suspended in PBS buffer once again. This process was repeated three times. Then, the bacteria were thoroughly ground in liquid nitrogen. The abrasive powder was fully suspended in PBS buffer and incubated at room temperature for 1 h. Then, the mixture was centrifuged and the pellet was collected. The Hg in the pellet was measured and named K2. The value of K2 was regarded as the intracellular Hg content. All of the experiments were performed four times and the average values were used.
2.8. Treating Wastewater Using the Engineered Bacteria
One clone each of BL21-7 and BL21-0 on LB plates was cultured in 5 mL of LB liquid medium containing 50 mg/L kanamycin for 16 h. Then, 2 mL of each culture was transferred to 200 mL of LB liquid medium containing 50 mg/L kanamycin and cultured at 37 °C for 5 h. IPTG was then added to the cultures to a final concentration of 0.8 mmol/L. The cultures were cultured at 37 °C for another 12 h. The bacteria were collected by centrifugation at 4 °C and 10,000 g. The supernatant was discarded and the pellet was washed with PBS three times. The bacteria suspended in PBS buffer were added 1 L of wastewater. The wastewater was placed in a shaker and cultured at 37 °C and 200 rpm for 24 h. Then, the bacteria were collected by centrifugation at 10,000 g. The Hg content in the supernatant was measured. The wastewater sample was obtained from a chlor-alkali plant in Changsha, Hunan province, China. The wastewater was filtered through medium-flow-rate filter paper. Then, the effluent was filtered again through a Millipore membrane with a pore size of 0.45 µm. The characterizations of the waste water was as the following: pH = 7.8, electric conductivity 1760 µs/cm, origin temperature 22 °C, CODCr 760 mg/L, alkalinity 106 mg/L. The content of Ca2+, Mg2+, Na+, Cl−, SO42− was 4.92 mg/L, 0.45 mg/L, 1.18 mg/L, 170 mg/L, 1.8 mg/L, respectively. The total Hg content in the waste water was 0.05 mg/L. To check the effect of the engineered strain, HgCl2 was added into the waste water to the total-Hg content was 0.97 mg/L. All of the experiments were performed four times and the average values were used.
3. Results and Discussion
3.1. Construction of Engineered Bacterial Strain BL21-7
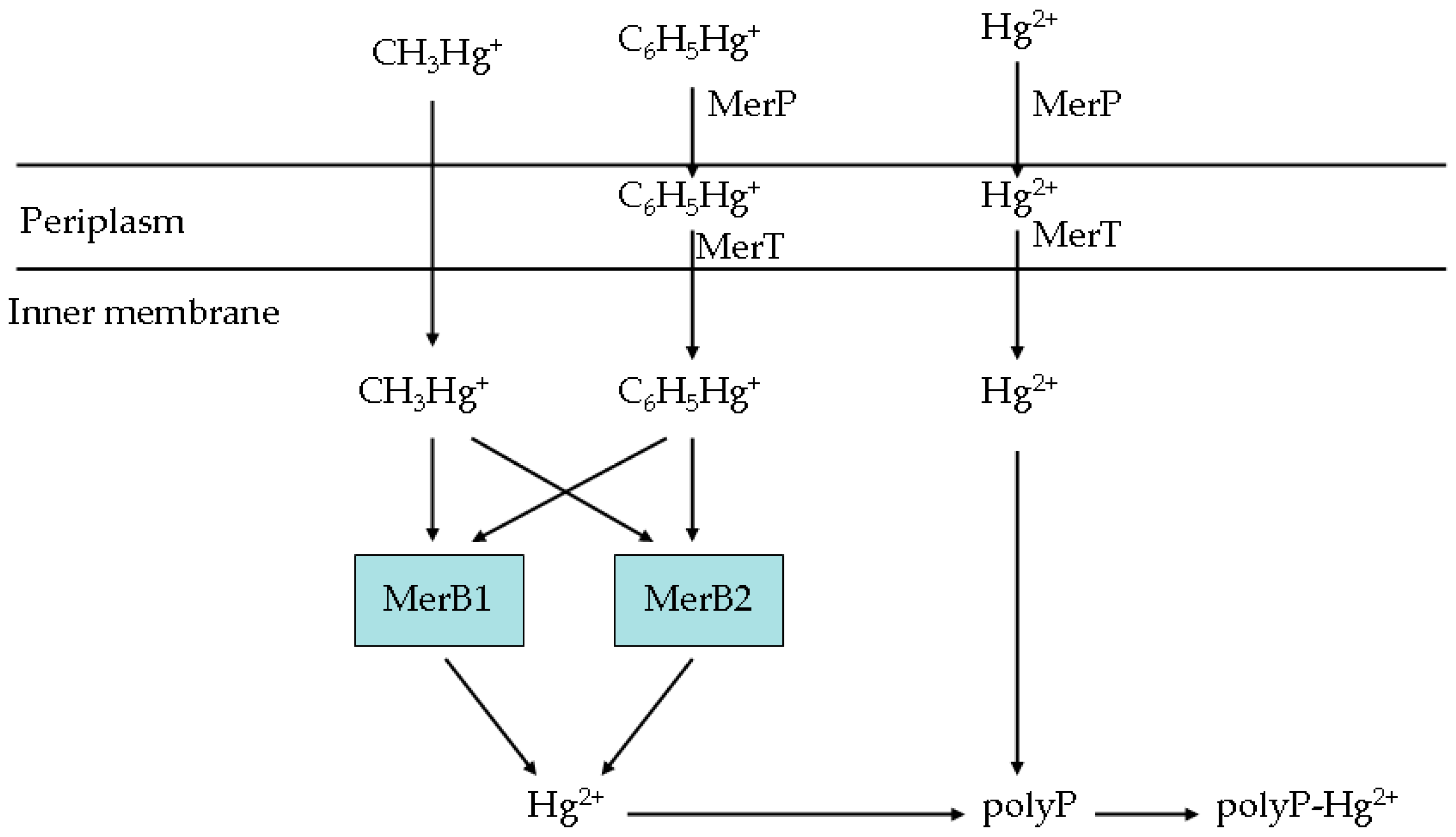
Pseudomonas K-62 is known for its high tolerance to Hg [7,16]. It can transport Hg into the cell by MerT-MerP [16]. The carbon-Hg linkages in organic Hg can be split by MerB1 and MerB2, releasing Hg2+ intracellularly [17]. An artificial operon, P16S-g10-merT-merP-merB1-merB2-ppk-rpsT, was constructed in this study (Figure 1 and Supplementary Materials S2). P16S is one of the strongest constitutive promoters. It is derived from the 5′ UTR of the plastid 16S ribosomal RNA gene [12,13,14]. The Hg tolerance of bacteria in which the transcription of mt-1 is promoted by P16S was at least six times greater than that of bacteria in which P16S was not included [6]. G10 is a well-known translation enhancer. When placed upstream of a synthetic ribosome-binding site (RBS), there was a 110-fold increase in the translation efficiency of the downstream lacZ gene [11]. As a strong terminator, rpsT can enhance transcript stability by forming a secondary structure at the 3′ end of the mRNA [15]. The ppk gene encodes a key enzyme responsible for synthesizing polyphosphate, which can respond to mercuric ions by chelating reactions and decrease their toxicity to cells [18,19]. The mercuric ions in cells were transferred from the extracellular space by MerT-MerP [16] or from organic Hg by MerB1 and MerB2 [17]. The E. coli BL21 (DE3) strain transformed with plasmid pET28a harboring the artificial operon P16S-g10-merT-merP-merB1-merB2-ppk-rpsT was named BL21-7.
3.2. Transcription of the Artificial Operon P16S-g10-merT-merP-merB1-merB2-ppk-rpsT
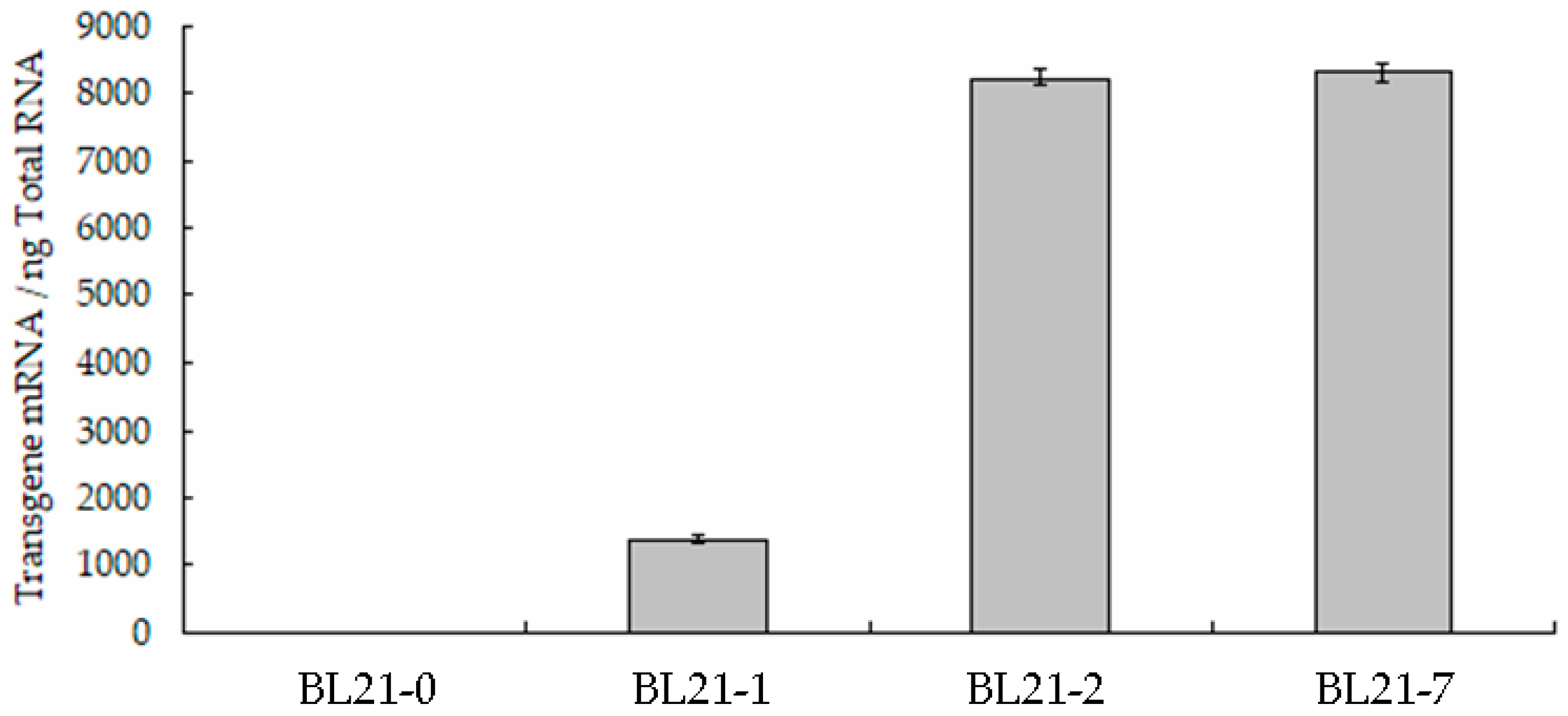
BL21-0, BL21-1, BL21-2 and BL21-7 were cultured in LB liquid medium at 37 °C for 16 h. The mRNAs of these bacteria were extracted and qPCR was performed. The cDNA of BL21-0 was used as a control. The results showed that the transcripts of ppk-rpsT in BL21-1, BL21-2 and BL21-7 were 1373 ± 62 copies/ng, 8245 ± 119 copies/ng and 8326 ± 141 copies/ng, respectively (Figure 2). The experiment was performed four times. The transcripts of ppk-rpsT in BL21-7 were approximately six-fold those in BL21-1 (Figure 2). No transcript was detected in BL21-0. These results demonstrated that the vector p5 (pET28a harboring P16S-g10-merT-merP-merB1-merB2-ppk-rpsT) could provide sufficient transcripts. Sufficient transcripts may produce high-abundance protein [6]. The Hg tolerance of BL21-2 was approximately fivefold greater than that of BL21-1 (Figure 3). In BL21-1, the artificial operon g10-ppk-rpsT was regulated by the T7 promoter, while the transcription of the same artificial operon was promoted by P16S in BL21-2. This showed that the transcription initiated by P16S was stronger than that of the T7 promoter. The Hg tolerance of BL21-7 (Figure 3) was approximately sevenfold greater than that of the strain transformed with merR-o/p-merT-merP-merB1-ppk [20]. This might be due to the strong promoter P16S, the translation enhancer g10 and the terminator rpsT in the artificial operon.
3.3. Bacterial Strain Tolerance to Hg
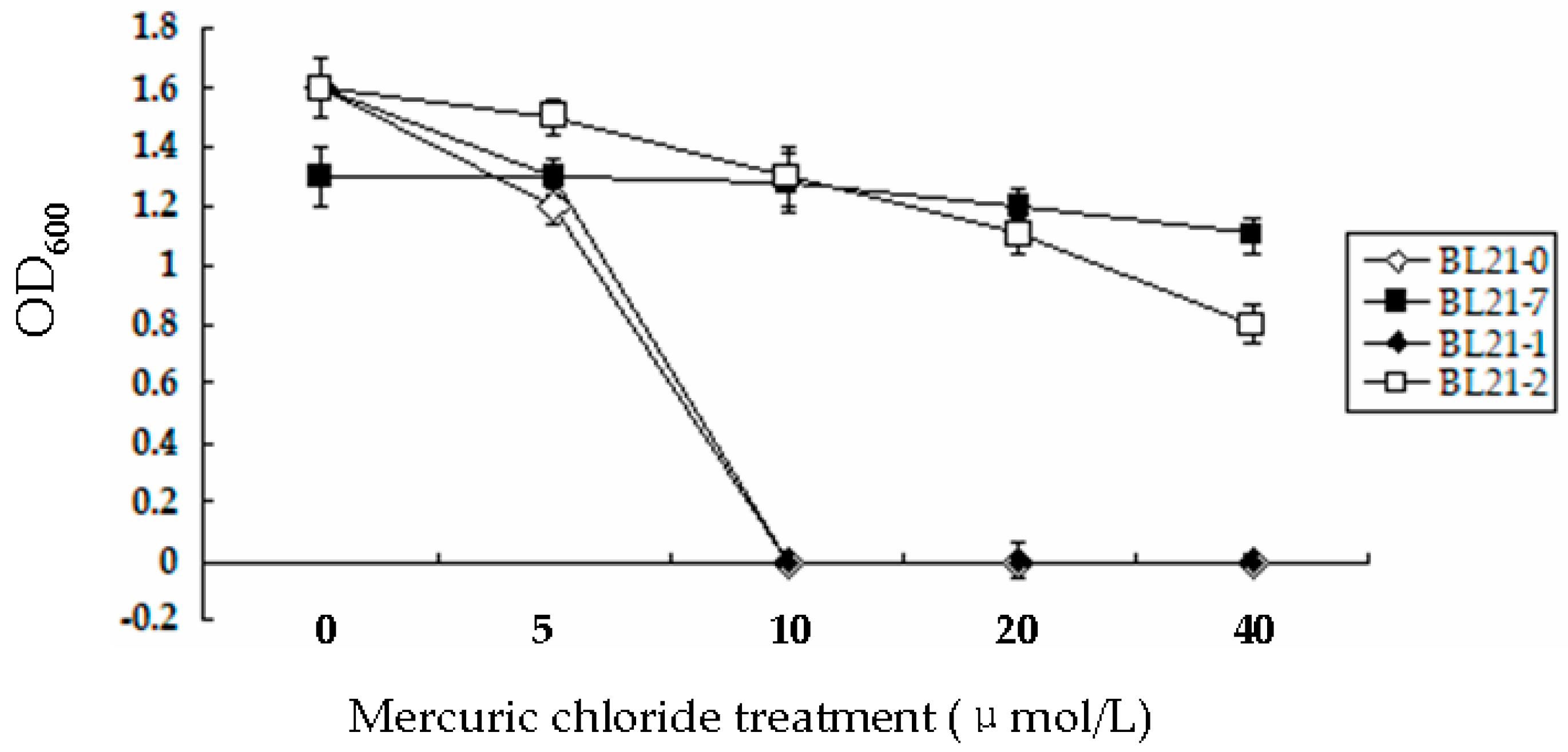
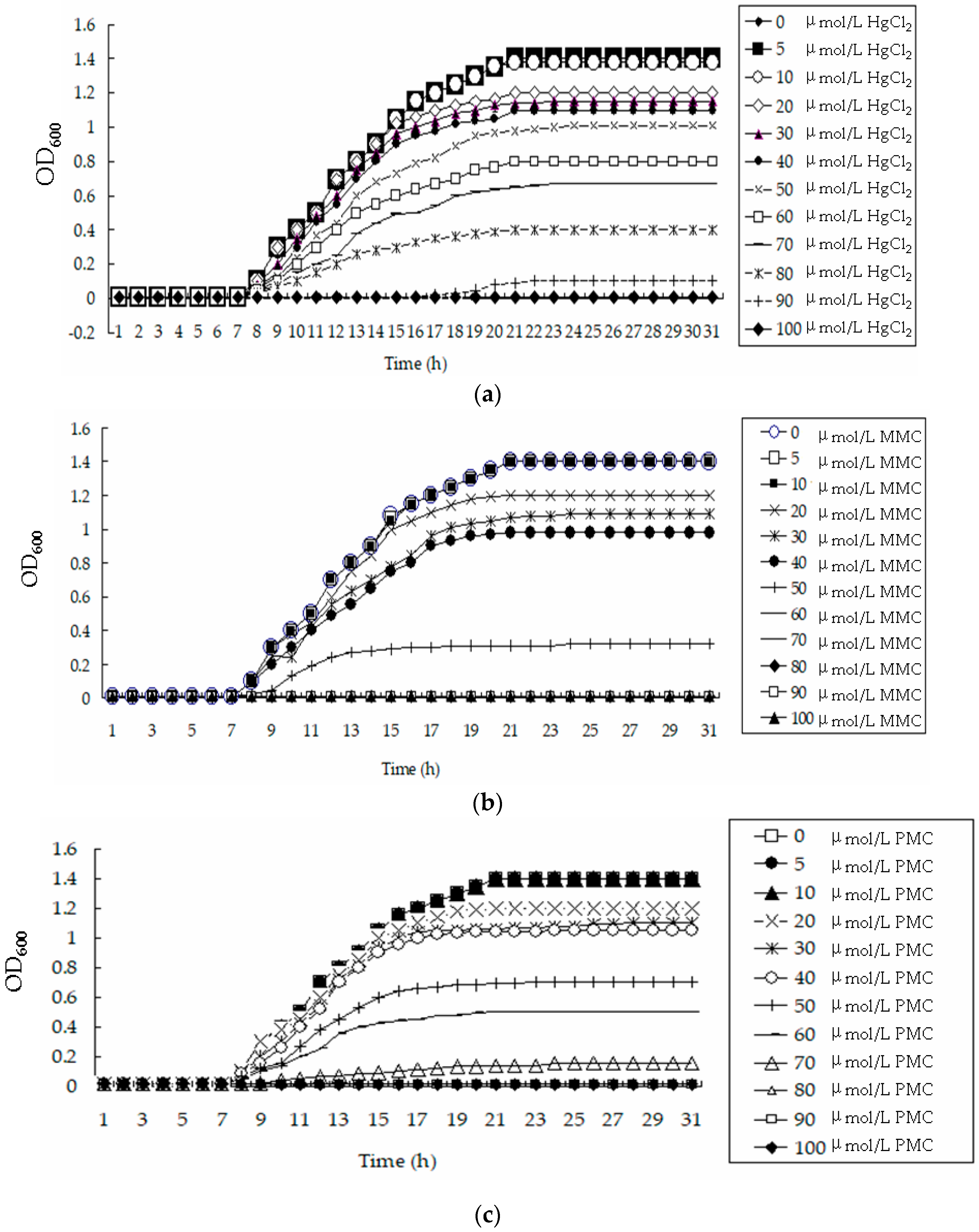
All of the bacteria were cultured for 120 h. The OD600 values were measured every 2 h. The growth curves of BL21-0, BL21-1, BL21-2 and BL21-7 cultured in liquid LB medium without Hg were S shaped. The growth of these four strains cultured in liquid LB medium containing 5 µmol/L HgCl2 also demonstrated an S shape. However, at the stationary phase, the OD600 values of BL21-0 and BL21-1 cultured in the medium containing 5 µmol/L HgCl2 were lower than those corresponding cultures without Hg. The OD600 values of BL21-7 cultured in medium containing 5 µmol/L HgCl2 were similar to those of corresponding cultures without Hg. In the media containing 10 µmol/L, 20 µmol/L and 40 µmol/L HgCl2, the growth curves of BL21-7 and BL21-2 still showed an S shape. BL21-0 cultured in LB medium without Hg grew much more quickly than that cultured in LB medium containing HgCl2. The highest concentration of HgCl2 that BL21-0 could withstand was 5 µmol/L (Figure 3). The growth of BL21-0 was inhibited completely in LB medium containing a HgCl2 concentration of 10 µmol/L or more (Figure 3). There were no significant differences between the OD600 values of BL21-0 cultured in these media for 16 h and those of BL21-0 cultured in these media for 120 h. BL21-1 was also very sensitive to HgCl2; it could not grow in LB medium containing a HgCl2 concentration of 10 µmol/L or more (Figure 3). After BL21-2 and BL21-7 were cultured in LB liquid medium containing 20 µmol/L HgCl2 for 16 h, their OD600 values were 1.1 ± 0.05 and 1.2 ± 0.05, respectively. They exhibited good tolerance to 20 µmol/L HgCl2 (Figure 3). After being cultured for 120 h, BL21-7 cultured in LB containing 20 µmol/L HgCl2 had similar OD600 values to BL21-7 cultured in LB without Hg. The bacterial strain tolerance to HgCl2 can be ordered from the strongest to the weakest as follows: BL21-7, BL21-2, BL21-1 and BL21-0.
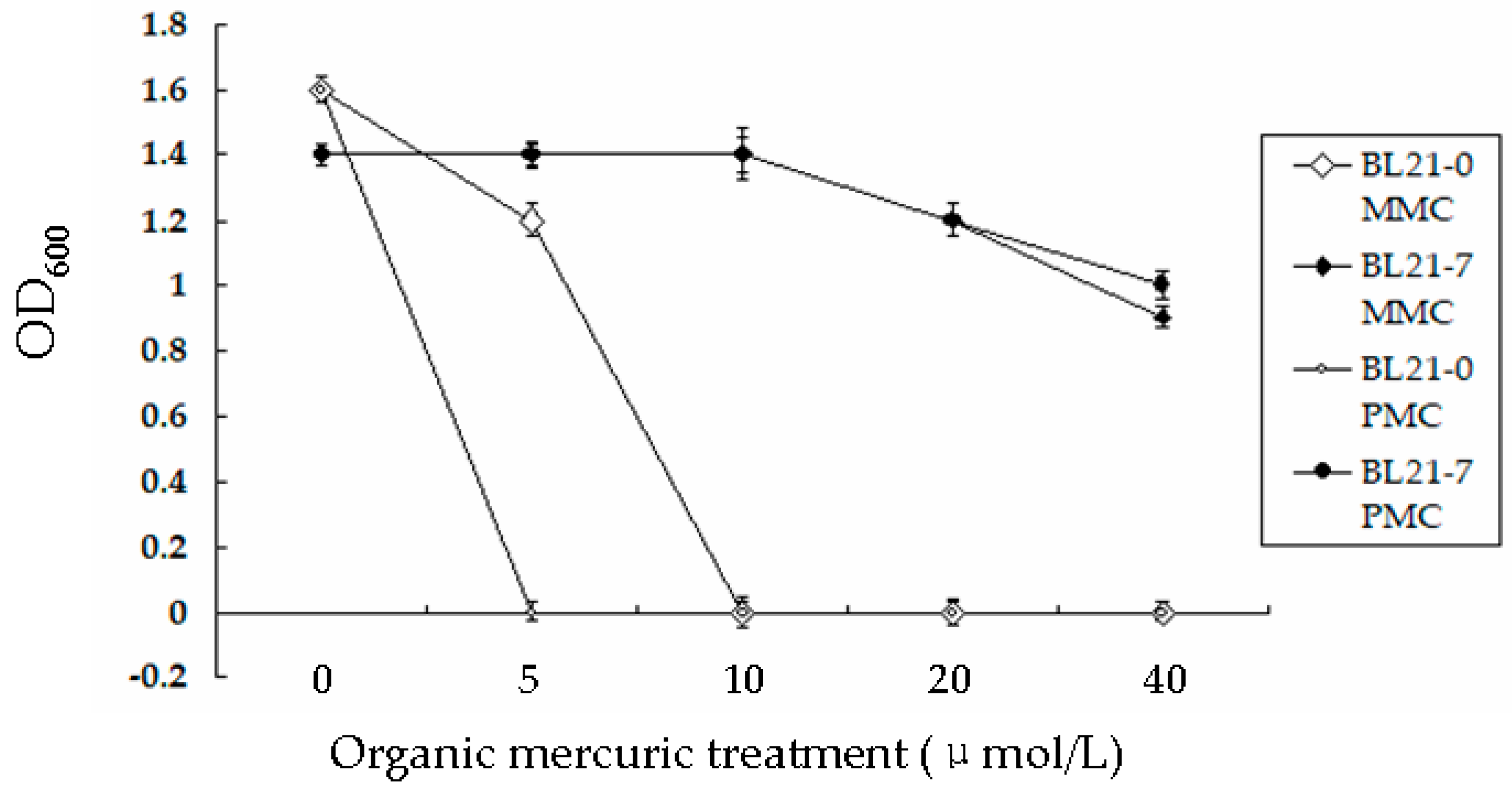
To identify whether BL21-7 displayed good tolerance to organic Hg, the engineered bacterial strain was cultured in a series of LB media containing different quantities of MMC or PMC. All of the bacteria were cultured for 120 h. The OD600 values were measured every 2 h. The growth curves of BL21-0 and BL21-7 cultured in liquid LB medium without Hg showed an S shape. BL21-0 grew well in liquid LB medium containing 5 µmol/L MMC. However, it did not grow in LB medium containing 5 µmol/L PMC or 10 µmol/L MMC. After BL21-7 was cultured in LB liquid medium containing 10 µmol/L MMC for 16 h, the OD600 value of the culture was still approximately 1.4 ± 0.08. This bacterial strain grew well in LB containing 20 µmol/L MMC, 40 µmol/L MMC, 20 µmol/L PMC or 40 µmol/L PMC (Figure 4).
To further identify the engineered strain’s tolerance to Hg, its MICs of HgCl2, PMC and MMC were measured. A series of conical flasks were filled with 200 mL of fresh LB liquid medium. A stock solution of HgCl2 (0.1 mol/L) was initially prepared and a series of HgCl2 stock solutions were added to the medium to final concentrations of 0, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90 and 100 µmol/L. The OD600 values of the cultures were measured every h. The minimum Hg concentration of LB liquid medium in which the bacteria could not grow was used as the MIC. The bacteria’s MICs for MMC and PMC were measured following the same method. All of the experiments were performed four times and the average values were used. The results showed that the growth curves of BL21-7 cultured in LB containing 0 µmol/L, 5 µmol/L and 10 µmol/L HgCl2 were very similar (Figure 5a). As shown in Figure 5, they could not be distinguished. The growth curves of BL21-7 cultured in LB containing 0 µmol/L, 5 µmol/L and 10 µmol/L MMC were also very similar (Figure 5b). For BL21-7, the MICs of Hg2+, MMC and PMC were 100 µmol/L, 60 µmol/L and 80 µmol/L, respectively (Figure 5a–c). BL21-7 could remove approximately 80% of the Hg2+ in the medium when the Hg2+ concentration was 80 µmol/L (Supplementary Materials Figure S3a). BL21-7 could effectively accumulate MMC and PMC when the concentrations of MMC and PMC were approximately 40 µmol/L and 60 µmol/L, respectively (Supplementary Materials Figure S3b,c). For BL21-7 on LB plates, the MICs of HgCl2, MMC and PMC were 100 µmol/L, 60 µmol/L and 80 µmol/L, respectively. These MICs were consistent with the measured MIC of the Hg remaining in the supernatant. To characterize the MICs of BL21-7 when the nutrition in the medium was poor, this strain was cultured on PB plates containing different Hg derivatives. The results showed that for BL21-7 on PB plates, the MICs of HgCl2, MMC and PMC were 30 µmol/L, 20 µmol/L and 30 µmol/L, respectively.
An engineered bacterial strain, MSR33 has been reported, with MICs of HgCl2 and MMC of 120 µmol/L and 80 µmol/L, respectively [8]. CM111 and HG3 also had high HgCl2 MICs [10]. Mt-1 could effectively accumulate Hg from medium containing 120 µmol/L HgCl2 [6]. These bacteria all had higher MICs than that of BL21-7. However, the mechanism of MSR33 for removing Hg involves volatilization, which releases Hg into the air and pollutes the environment. The tolerance of CM111 and HG3 to HgCl2 was thought to not be due to Hg(0) volatilization [10]. However, the detailed mechanisms by which these two strains tolerated HgCl2 remained unclear. Although mt-1 had a high tolerance to HgCl2, it remains unknown whether it can accumulate organic Hg. Comparatively, BL21-7 not only exhibited high MICs of HgCl2, MMC and PMC (Figure 5) but could also store Hg in the cell (Supplementary Materials S3). BL21-7 not only accumulated Hg2+ but also effectively accumulated organic Hg (MMC and PMC) (Supplementary Materials S3). Kiyono et al. constructed an engineered bacterial strain harboring the pMKB18 plasmid, which contained merT-merP-merB1-ppk [9]. After the engineered bacteria were cultured in LB medium containing 80 µmol/L HgCl2 for 5 h, the Hg remaining in the medium was nearly 40 µmol/L [9]. However, after BL21-7 was cultured in LB medium containing 80 µmol/L HgCl2 for 5 h, the Hg remaining in the medium was only approximately 25 ± 4 µmol/L (Supplementary Materials Figure S3a). BL21-7 could also remove more phenylmercury from the medium than E. coli harboring pMKB18 (Supplementary Materials Figure S3b) [9]. These findings demonstrated that BL21-7 had greater Hg-accumulation capacity than E. coli harboring pMKB18.
3.4. Accumulation of Hg Derivatives by BL21-7 Cultured in LB Medium
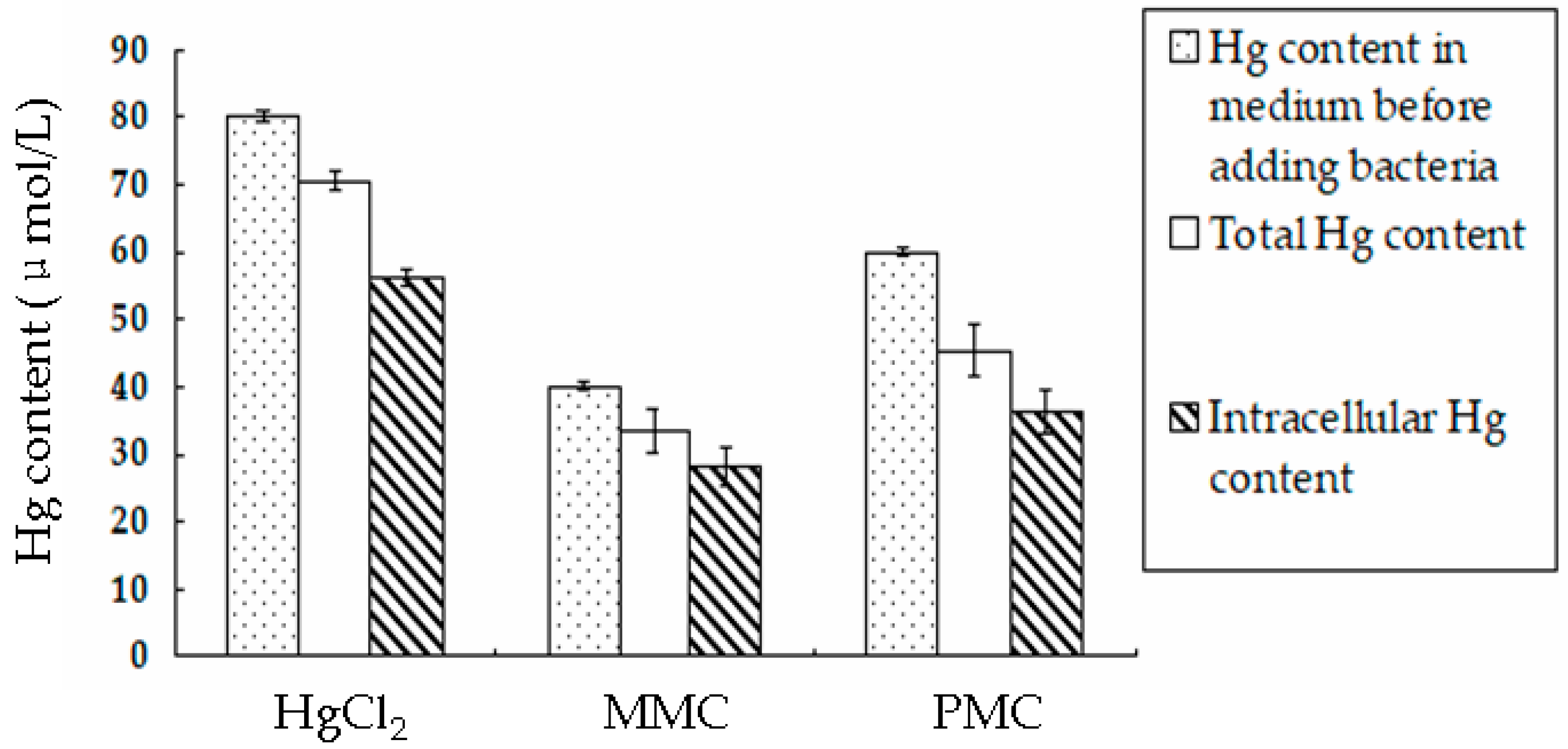
To further identify whether the bacteria could accumulate more Hg when they were cultured for a sufficient length of time, Hg in bacteria was measured after the bacteria were cultured in media containing mercuric varieties for 72 h. When bacterial BL21-7 accumulated Hg from the liquid medium, Hg could attach to the cell wall and not enter the bacterial cell. The data measured above included Hg attached to the cell wall and Hg in the bacterial cells. To measure the Hg content inside bacterial cells, we measured the total Hg content and the Hg attached to the cell wall. The difference between the total Hg content and the Hg attached to the cell wall was regarded as the intracellular Hg content. The results showed that after BL21-7 was cultured in LB medium containing 80 µmol/L HgCl2 for 72 h, the Hg content in the bacterial pellet was 70.5 ± 1.5 µmol/L (Figure 6). The intracellular Hg content was 56.2 ± 1.3 µmol/L (Figure 6). After BL21-7 was cultured in LB medium containing 40 µmol/L MMC or 60 µmol/L PMC for 72 h, the Hg content in the bacterial pellets was 33.5 ± 3.2 µmol/L and 45.3 ± 3.7 µmol/L, respectively (Figure 6). The intracellular Hg content was 28.3 ± 3.0 µmol/L and 36.5 ± 3.1 µmol/L, respectively (Figure 6). These results demonstrated that the engineered bacteria could efficiently accumulate HgCl2, MMC and PMC. This bacterial strain not only accumulated mercuric ions in the medium but also effectively accumulated organic Hg (PMC and MMC).
3.5. Effects of Other Metals on the Accumulation of Hg by BL21-7
Cd2+, Pb2+ and Cu2 are always found at Hg-polluted sites. To characterize whether these metals would affect the engineered bacterial strain when it was used to remove Hg from a medium, the MICs of BL21-7 for Cd2+, Pb2+ and Cu2+ were also measured. All of the experiments were performed four times and the average values were used. The results for BL21-7 on LB plates showed that the MICs for Cd2+, Pb2+ and Cu2+ were 13 µmol/L, 12 µmol/L and 22 µmol/L, respectively. After BL21-7 was cultured in LB liquid medium containing 10 µmol/L mercurial derivatives (HgCl2, MMC or PMC) without other metals for 5 h, the concentrations of the Hg removed from the medium were 8.80 µmol/L, 5.47 µmol/L and 7.80 µmol/L, respectively. After the engineered bacteria were cultured in LB liquid medium containing 10 µmol/L mercurial derivatives (HgCl2, MMC or PMC) in the presence of 10 µmol/L Cd2+ for 5 h, the concentrations of the Hg removed from the medium were 8.66 µmol/L, 5.53 µmol/L and 7.76 µmol/L, respectively (Table 3). No significant differences were found between the Hg removed from the medium containing Cd2+ and the Hg from the medium without Cd2+. Similar results were obtained after BL21-7 was cultured in medium containing 10 µmol/L mercurial derivatives (HgCl2, MMC or PMC) in the presence of 10 µmol/L Pb2+ or Cu2+ (Table 3). After BL21-7 was cultured in LB medium containing 10 µmol/L mercurial derivatives (HgCl2, MMC or PMC) in the presence of 10 µmol/L Cd2+, 10 µmol/L Pb2+ and 10 µmol/L Cu2+ for 5 h, the values of the Hg removed were the highest among the four treatments. However, when the standard deviations were considered, the differences between the values of Hg removed from the medium without other metals and the values of Hg removed from the medium containing 10 µmol/L Cd2+ (or the other three treatments) were not significant. These findings demonstrated that in the presence of 10 µmol/L Cd2+, Pb2+ or Cu2+, the removal of mercurial derivatives by BL21-7 was not affected (Table 3).
3.6. Effects of pH and Temperature on the Accumulation of Hg by BL21-7
To characterize the effects of pH on the ability of the engineered bacteria to remove Hg from the environment, BL21-7 was cultured in LB liquid medium overnight at 37 °C. One microliter of this culture was transferred to 100 mL of nutrition-poor PB medium (pH = 5, 6, 7, 8 or 9) containing 20 µmol/L HgCl2 and was cultured continually. The OD600 values of the cultures were measured every two h. After the cultures entered stationary phase, the Hg contents in the bacteria and medium were measured. The results showed that all of the growth curves of the cultures showed an S shape. No significant differences were found among the growth curves of cultures at pH = 5, 7 and 8 (Figure 7). For example, after BL21-7 was cultured for 30 h, the OD600 values of the cultures (pH = 5, 7 and 8) were 1.21 ± 0.03, 1.22 ± 0.03 and 1.2 ± 0.03, respectively. Among the five PB media (pH = 5, 6, 7, 8 and 9), the OD600 values of BL21-7 cultured in PB at pH = 6 were the highest in stationary phase. The engineered bacteria could also grow in PB at pH = 9. However, they grew very slowly (Figure 7). The final OD600 of the culture at pH = 9 was also the lowest among the five treatments. After BL21-7 was cultured for 40 h, the Hg contents in the supernatant of the PB media (pH = 5, 6, 7, 8 and 9) containing 20 µmol/L HgCl2 were 6.1 ± 0.10 µmol/L, 5.2 ± 0.11 µmol/L, 5.9 ± 0.12 µmol/L, 7.6 ± 0.12 µmol/L and 12.5 ± 0.11 µmol/L, respectively. The Hg content in the supernatant of the PB at pH = 6 containing 20 µmol/L HgCl2 was the lowest among the five treatments. The engineered bacterial strain accumulated the most Hg in the liquid medium at a pH of 6 and it worked well in the media with pH values from 5 to 8.
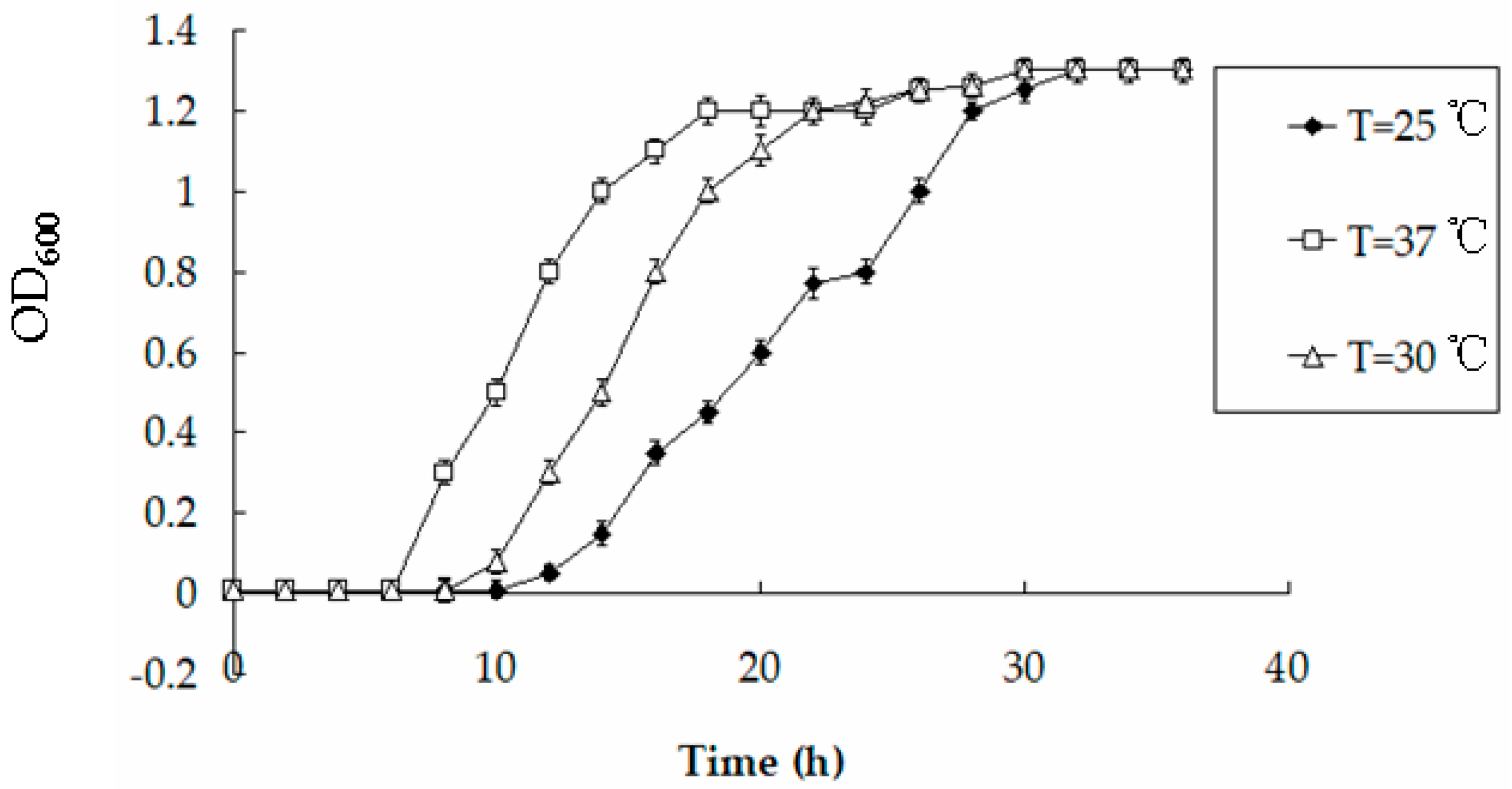
To characterize the effects of temperature on the engineered strain’s growth and accumulation of Hg, BL21-7 was cultured in LB liquid medium overnight at 37 °C. One microliter of this culture was transferred to 100 mL of nutrition-poor PB medium containing 20 µmol/L HgCl2 and was cultured at 25 °C, 30 °C and 37 °C. The OD600 values and Hg content were measured as described above. The results showed that although BL21-7 grew more slowly at 25 °C than at 30 °C or 37 °C, the OD600 values of these three treatments were similar at stationary phase (Figure 8). For example, although the OD600 values of the cultures (T = 25 °C, 30 °C and 37 °C) at 20 h were 0.6 ± 0.04, 1.1 ± 0.04 and 1.2 ± 0.03, respectively, the OD600 values of the same cultures at 34 h were all 1.3 ± 0.03. After BL21-7 was cultured for 20 h, the Hg contents in the supernatant of the PB medium (T = 25 °C, 30 °C and 37 °C) containing 20 µmol/L HgCl2 were 11.2 ± 0.12 µmol/L, 8.2 ± 0.15 µmol/L and 6.5 ± 0.11 µmol/L, respectively. When BL21-7 was cultured for 34 h, the Hg contents in the supernatant of the PB medium (T = 25 °C, 30 °C and 37 °C) containing 20 µmol/L HgCl2 were 6.1 ± 0.11 µmol/L, 6.2 ± 0.15 µmol/L and 6.0 ± 0.12 µmol/L, respectively. At stationary phase, the Hg contents in the supernatant of the PB medium (T = 25 °C, 30 °C and 37 °C) containing 20 µmol/L HgCl2 were similar. These data indicate that BL21-7 functions well at temperatures from 25 °C to 37 °C.
3.7. Hg in Wastewater Can Be Removed by BL21-7
To identify whether the engineered bacteria BL21-7 could be used to remove Hg from wastewater, 200 mL of BL21-7 bacteria and BL21-0 bacteria (E. coli BL21 harboring the empty vector pET28a) were treated as described in Methods and were added to 1 L of wastewater. The cultures were shaken at 37 °C and 200 rpm for 24 h. Then, the bacteria were collected and the Hg content in the wastewater was measured. The results showed that before the wastewater was treated using the engineered bacteria, the Hg content in the wastewater samples was 4.85 ± 0.31 µmol/L (Figure 9). After the engineered bacteria BL21-7 were added to the wastewater and shaken at 37 °C for 24 h, the Hg content in the wastewater was reduced to 2.73 ± 0.15 µmol/L (Figure 9). Approximately 43.7% of the Hg in the wastewater had been removed. However, after BL21-0 was added to the wastewater and shaken at 37 °C for 24 h, the Hg content in the wastewater was 4.8 ± 0.27 µmol/L (Figure 9). This result demonstrates that the engineered bacteria BL21-7 can accumulate Hg in wastewater. Only bacterial cells were added to the wastewater in our experiments. If substances containing sufficient nutrients were added to the wastewater with the bacterial cells, the effect on Hg accumulation might be more significant.
3.8. The Roles of the Genes in the Artificial Operon
To characterize the effects of the different genes in the artificial operon on Hg accumulation, strains harboring different mer and ppk genes were constructed. After the strains were cultured in medium containing Hg for a certain time span, the MRMs were measured. For Hg2+ accumulation, the MRMs of BL21-B1, BL21-B2 and BL21-B1B2 were similar to that of BL21-0 (Table 4). The MRM of BL21-P was lower than those of the strains above. BL21-P accumulated more Hg than the strains described above. MerP can bind to Hg2+ and plays a role as a scavenger to protect other periplasmic and membrane proteins from binding to Hg2+ [21]. BL21-T transports more Hg2+ than BL21-P. MerT is responsible for transporting Hg2+ into cells [16]. The MRM of BL21-7 was the lowest among all the strains, indicating that BL21-7 was the most effective strain for accumulating Hg2+.
For MMC accumulation, the MRM of BL21-7 was much lower than that of BL21-0 (Table 4), indicating that BL21-7 could also effectively accumulate MMC. The MRMs of BL21-T, BL21-P, BL21-TP, BL21-TPK and BL21-0 were similar, demonstrating that MerT and MerP did not participate in transporting MMC. The MRM of BL21-TPK was similar to that of BL21-TP, indicating that polyphosphate could not bind to MMC. The MRMs of BL21-B1 and BL21-B2 were lower than that of BL21-0, suggesting that MerB1 and MerB2 could also catalyze the splitting of the carbon-Hg bond in MMC. BL21-7 was the best strain for MMC removal. After BL21-7 was cultured in LB medium containing 20 µmol/L MMC for 15 h, the MRM was only approximately 1.4 µmol/L (Table 4).
For PMC accumulation, the MRMs of BL21-B1 and BL21-B2 were lower than those of BL21-T, BL21-TP and BL21-TPK, suggesting that MerB1 and MerB2 could promote PMC accumulation. This result could be due to their increasing the host cell’s tolerance to PMC, as MerB1 and MerB2 can catalyze the splitting of the carbon-Hg bond [22,23,24]. The MRM of BL21-7 was the lowest among the strains. The second lowest MRM was BL21-B1B2K.
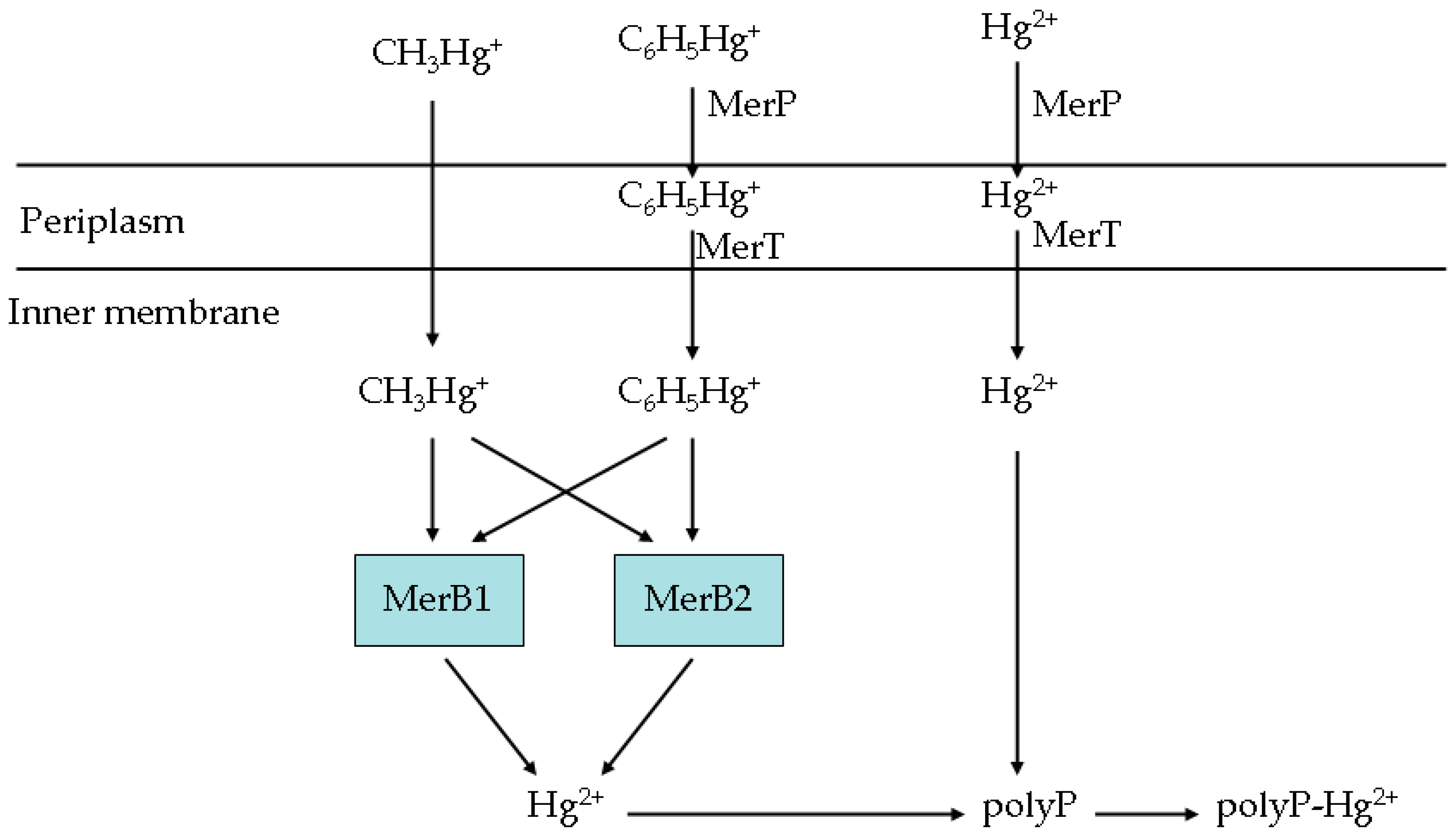
Although Pseudomonas K-62 has been shown to have approximately 1000-fold higher resistance to phenylmercury than E. coli [25], its biosafety had not been studied. E. coli BL21(DE3) was derived from normal commensals of the human gut [26] and is not harmful to humans. Therefore, we selected E. coli BL21(DE3), not Pseudomonas K-62, for transforming and constructing the engineered bacterial strain. The genome of E. coli BL21(DE3) has been sequenced and published [26]. However, although merB genes had been found in E. coli R831b and E. coli AAD3 [27], we were unable to find any mer genes in the E. coli BL21(DE3) genome. Compared to that of Pseudomonas K-62, the Hg-resistance phenotype of E. coli BL21(DE3) is negligible. To better understand the engineered bacteria’s ability to accumulate Hg, a mechanism for organic and inorganic Hg detoxification by BL21-7’s artificial operon was proposed (Figure 10). According to the proposed mechanism, in the artificial operon, both P16S and g10 can increase the expression levels of downstream genes [11,12]. RpsT can prevent the transcripts from being hydrolyzed [6]. This construct can result in sufficient transcription and protein accumulation for Hg tolerance and Hg accumulation. Phenylmercury and Hg2+ are transported into the cell periplasm by MerP [21]. The phenylmercury and Hg2+ in the periplasm are transported into the cell by MerT [8]. MerP can bind to Hg2+ and retain it in the periplasm [21]. However, high levels of phenylmercury in a cell can harm the cell. The phenylmercury tolerance of BL21-TP was lower than that of BL21-0. Methylmercury is transported into the cell by unknown means, as the MerT-MerP system does not transport methylmercury [7]. The methylmercury tolerance and methylmercury accumulation of BL21-T, BL21-P and BL21-TP were similar to those of BL21-0. Being catalyzed by MerB1 and MerB2, the carbon-Hg bond in phenylmercury or methylmercury is split and Hg2+ is released [22,23,24]. The toxicity of Hg2+ is lower than that of organic Hg. The tolerance to phenylmercury and methylmercury of BL21-TPB1B2 are higher than those of BL21-0. After ppk is overexpressed, a significant amount of polyP is produced. Hg2+ binds to polyP, forming the chelate polyP-Hg2+ [9]. The toxicity of polyP-Hg2+ is lower than that of Hg2+. Thus, the Hg tolerance and Hg accumulation of bacterial strains harboring ppk are higher than those of strains without the ppk gene.
4. Conclusions
The construction of an engineered bacterial strain, E. coli BL21-7, was reported in this study. The constructed plasmid contained a strong promoter, P16S, a translation enhancer, g10, a strong terminator, rpsT, Pseudomonas K-62 genes merT, merP, merB1 and merB2 and the Klebsiella aerogenes polyphosphate kinase gene ppk. This construct provided sufficient transcripts for Hg tolerance and Hg accumulation. A mechanism for Hg detoxification by BL21-7 was proposed. This bacterial strain did not release Hg into the environment. It could accumulate both inorganic and organic Hg. When the engineered bacterial strain BL21-7 was cultured in LB liquid medium containing 10 µmol/L mercurial derivatives (Hg2+, PMC or MMC) in the presence of 10 µmol/L Cd2+, 10 µmol/L Pb2+ or 10 µmol/L Cu2+, the removal of mercurial derivatives by BL21-7 was not affected. After BL21-7 bacterial samples were added to wastewater, the Hg content in the wastewater could be reduced significantly. These characteristics make BL21-7 a good candidate for the removal of Hg from wastewater.
Supplementary Materials
The following are available online at https://www.mdpi.com/2076-3417/8/9/1572/s1, S1: The sequence of the artificial operon P16S-g10-merT-merP-merB1-merB2-ppk-rpsT, S2: Construction of artificial operons and respective bacterial strains, S3: Hg remaining in medium after BL21-7 was cultured in LB liquid medium containing mercurial derivatives.
Author Contributions
S.C. and Z.W. did data curation. They were equal contributors. W.S. did data curation. H.S. validated the data and wrote the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China [No. 41201309].
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jan, A.T.; Murtaza, I.; Ali, A.; Mohd, Q.; Haq, R. Mercury pollution: An emerging problem and potential bacterial remediation strategies. World J. Microbiol. Biotechnol. 2009, 25, 1529–1537. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency (US EPA). EPA’s Roadmap for Mercury. July 2006. Available online: http://www.epa.gov/mercury/roadmap.htm (accessed on 26 October 2007).
- U.S. Environmental Protection Agency (US EPA) and Environment Canada. Update: Binational Toxics Strategy—Mercury Sources and Regulations. 1999. Available online: http://www.epa.gov/bns/mercury/stephg.html (accessed on 26 October 2007).
- U.S. Environmental Protection Agency (US EPA). Great Lakes Pollution Prevention and Toxics Strategy. Background Information on Mercury Sources and Regulations. 2005. Available online: http://www.epa.gov/grtlakes/bnsdocs/mercsrce/merc srce.html (accessed on 26 October 2007).
- Cachada, A.M.; Rodrigues, S.; Mieiro, C.; Ferreira da Silva, E.; Pereira, E.; Duarte, A.C. Controlling factors and environmental implications of mercury contamination in urban and agricultural soils under a long-term influence of a chlor-alkali plant in the North–West Portugal. Environ. Geol. 2009, 57, 91–98. [Google Scholar] [CrossRef]
- Ruiz, O.N.; Alvarez, D.; Gonzalez-Ruiz, G.; Torres, C. Characterization of mercury bioremediation by transgenic bacteria expressing metallothionein and polyphosphate kinase. BMC Biotechnol. 2011, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kiyono, M.; Omura, T.; Fujimori, H.; Pan-Hou, H. Lack of involvement of merT and merP in methylmercury transport in mercury resistant Pseudomonas K-62. FEMS Microbiol. Lett. 1995, 128, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Rojas, L.A.; Yanez, C.; Gonzalez, M.; Lobos, S.; Smalla, K.; Seeger, M. Characterization of the metabolically modified heavy metal-resistant Cupriavidus metallidurans strain MSR33 generated for mercury bioremediation. PLoS ONE 2011, 6, e17555. [Google Scholar] [CrossRef] [PubMed]
- Kiyono, M.; Omura, H.; Omura, T.; Murata, S.; Pan-Hou, H. Removal of inorganic and organic mercurials by immobilized bacteria having mer-ppk fusion plasmids. Appl. Microbiol. Biotechnol. 2003, 62, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Francois, F.; Lombard, C.; Guigner, J.; Soreau, P.; Brian-Jaisson, F.; Martino, G.; Vandervennet, M.; Garcia, D.; Molinier, A.; Pignol, D.; et al. Isolation and characterization of environmental bacteria capable of extracellular biosorption of mercury. Appl. Environ. Microbiol. 2011, 78, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Olins, P.O.; Rangwala, S.H. A novel sequence element derived from bacteriophage T7 mRNA acts as an enhancer of translation of the lacZ gene in Escherichia coli. J. Biol. Chem. 1989, 264, 16973–16976. [Google Scholar] [PubMed]
- Dennis, P.P.; Ehrenberg, M.; Fange, D.; Bremer, H. Varying rate of RNA chain elongation during rrn transcription in Escherichia coli. J. Bacteriol. 2009, 191, 3740–3746. [Google Scholar] [CrossRef] [PubMed]
- Newell, C.A.; Birch-Machin, I.; Hibberd, J.M.; Gray, J.C. Expression of green fluorescent protein from bacterial and plastid promoters in tobacco chloroplasts. Transgenic Res. 2003, 12, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, H.; Maliga, P. Complementarity of the 16S rRNA penultimate stem with sequences downstream of the AUG destabilizes the plastid mRNAs. Nucleic Acids Res. 2001, 29, 970–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiina, T.; Allison, L.; Maliga, P. RbcL transcript levels in tobacco plastids are independent of light: Reduced dark transcription rate is compensated by increased mRNA stability. Plant Cell 1998, 10, 1713–1722. [Google Scholar] [CrossRef] [PubMed]
- Silver, S.; Phung, L.T. Bacterial heavy metal resistance: New surprises. Annu. Rev. Microbiol. 1996, 50, 753–789. [Google Scholar] [CrossRef] [PubMed]
- Kiyono, M.; Pan-Hou, H. Genetic engineering of bacteria for environmental remediation of mercury. J. Health Sci. 2006, 52, 199–204. [Google Scholar] [CrossRef]
- Pan-Hou, H.; Kiyono, M.; Kawase, T.; Omura, T.; Endo, G. Evaluation of ppk-specified polyphosphate as a mercury remedial tool. Biol. Pharm. Bull. 2001, 24, 1423–1426. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, A. Inorganic polyphosphate: Toward making a forgotten polymer unforgettable. J. Bacteriol. 1995, 177, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Pan-Hou, H.; Kuyono, M.; Omura, H.; Omura, T.; Endo, G. Polyphosphate produced in recombinant Escherichia coli confers mercury resistance. FEMS Microbiol. Lett. 2002, 207, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Sone, Y.; Nakamura, R.; Pan-Hou, H.; Itoh, T.; Kiyono, M. Role of MerC, MerE, MerF, MerT, and/or MerP in resistance to mercurials and the transport of mercurials in Escherichia coli. Biol. Pharm. Bull. 2013, 36, 1835–1841. [Google Scholar] [CrossRef] [PubMed]
- Misra, T.K. Bacterial resistances to inorganic mercury salts and organomercurials. Plasmid 1992, 27, 1, 4–16. [Google Scholar] [CrossRef]
- Tezuka, T.; Tonomura, K. Purification and properties of an enzyme catalyzing the splitting of carbon-mercury linkages from mercury-resistant Pseudomonas K-62 strain. J. Biochem. 1976, 80, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Tezuka, T.; Tonomura, K. Purification and properties of a second enzyme catalyzing the splitting of carbon-mercury linkages from mercury-resistant Pseudomonas K-62. J. Bacteriol. 1978, 135, 138–143. [Google Scholar] [PubMed]
- Tonomura, K.; Maeda, K.; Futai, F.; Nakagami, T.; Yamada, M. Stimulative vaporization of phenylmercuric acetate by mercury-resistant bacteria. Nature 1968, 217, 644–646. [Google Scholar] [CrossRef] [PubMed]
- Studier, F.W.; Daegelen, P.; Lenski, R.E.; Maslov, S.; Kim, J.F. Understanding the differences between genome sequences of Escherichia coli B strains REL606 and BL21(DE3) and comparision of the E. coli B and K-12 genomes. J. Mol. Biol. 2009, 394, 653–680. [Google Scholar] [CrossRef] [PubMed]
- Barkay, T.; Wagner-Dobler, I. Microbial transformations of mercury: Potentials, challenges and achievements in controlling mercury toxicity in the environment. Adv. Appl. Microbiol. 2005, 57, 1–52. [Google Scholar] [PubMed]
Figure 1.
Schematic diagram of the designed artificial operon.

Figure 2.
The expression levels of ppk-rpsT in different E. coli strains. Columns 1-4 represent BL21-0, BL21-1, BL21-2 and BL21-7, respectively.
Figure 2.
The expression levels of ppk-rpsT in different E. coli strains. Columns 1-4 represent BL21-0, BL21-1, BL21-2 and BL21-7, respectively.

Figure 3.
E. coli strains’ tolerance to HgCl2. The OD600 values were measured after the strains were cultured in LB liquid medium containing HgCl2 for 120 h.
Figure 3.
E. coli strains’ tolerance to HgCl2. The OD600 values were measured after the strains were cultured in LB liquid medium containing HgCl2 for 120 h.
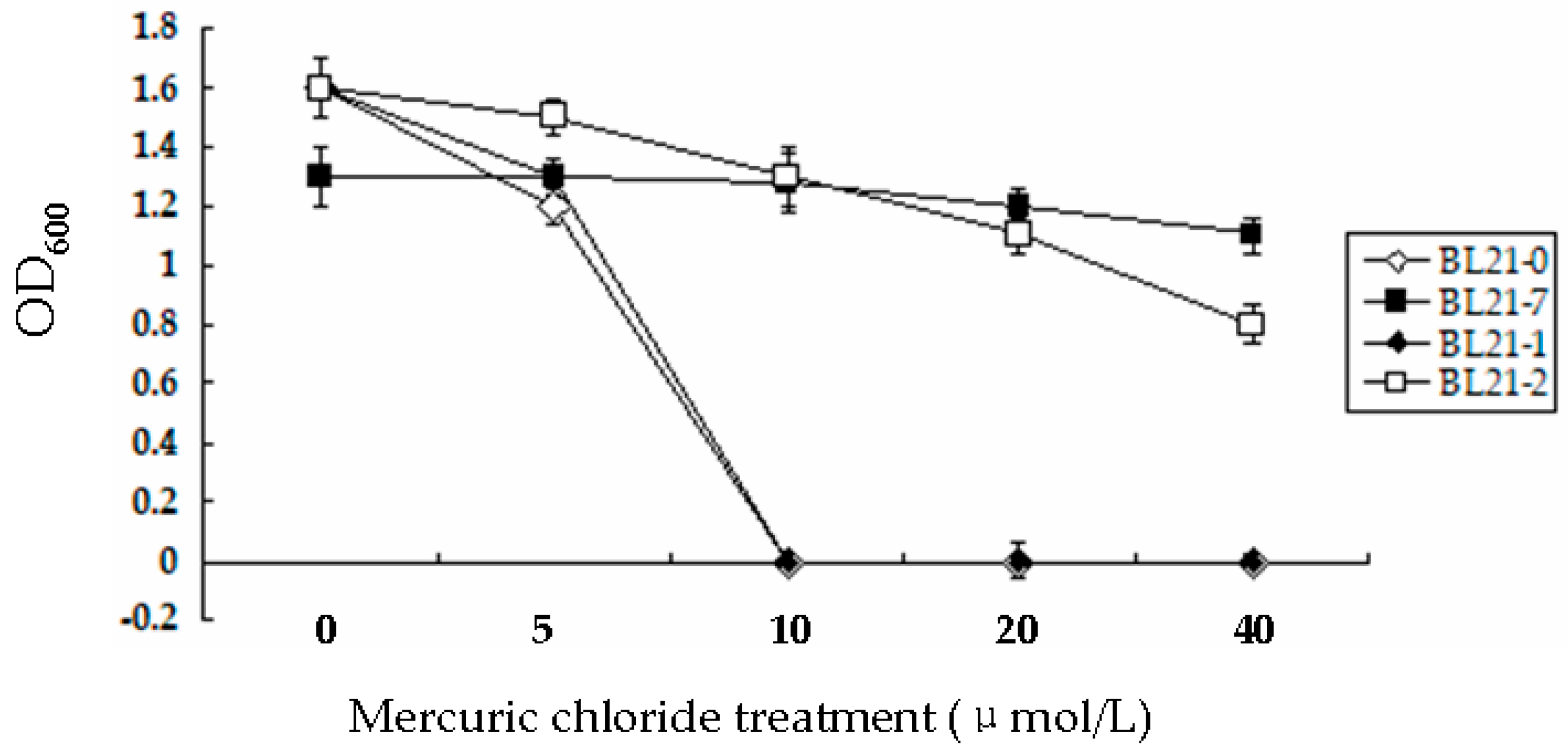
Figure 4.
E. coli strains’ tolerance to organic Hg. The OD600 values were measured after the strains were cultured in LB liquid medium containing MMC or PMC for 120 h.
Figure 4.
E. coli strains’ tolerance to organic Hg. The OD600 values were measured after the strains were cultured in LB liquid medium containing MMC or PMC for 120 h.

Figure 5.
Growth curves of BL21-7 cultured in LB liquid medium containing mercurial derivatives. (a) shows BL21-7 cultured in LB liquid medium containing HgCl2. (b) represents BL21-7 cultured in the presence of MMC. (c) shows BL21-7 cultured in the presence of PMC.
Figure 5.
Growth curves of BL21-7 cultured in LB liquid medium containing mercurial derivatives. (a) shows BL21-7 cultured in LB liquid medium containing HgCl2. (b) represents BL21-7 cultured in the presence of MMC. (c) shows BL21-7 cultured in the presence of PMC.
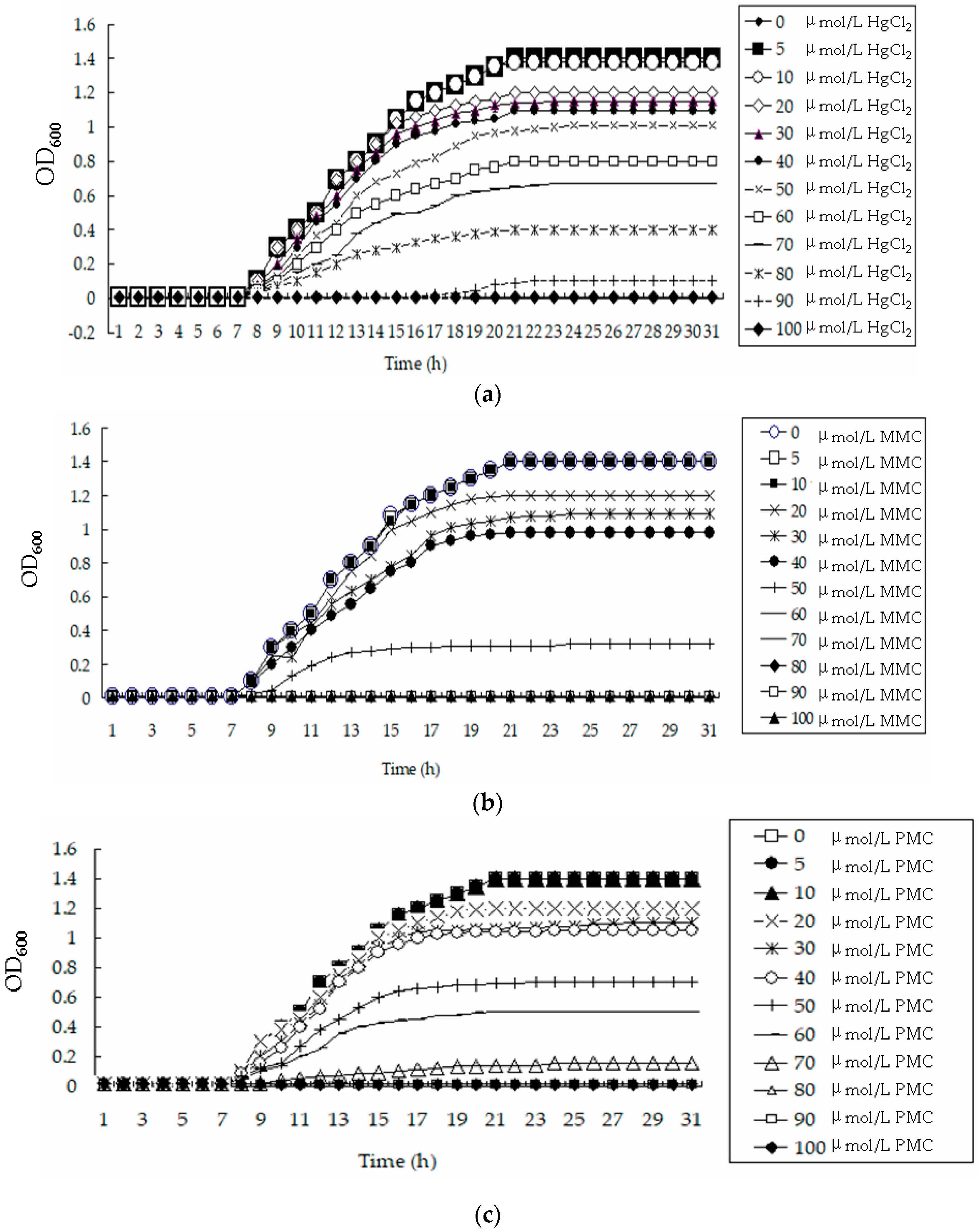
Figure 6.
Hg content in BL21-7 bacteria cultured in LB liquid medium containing mercurial derivatives.
Figure 6.
Hg content in BL21-7 bacteria cultured in LB liquid medium containing mercurial derivatives.
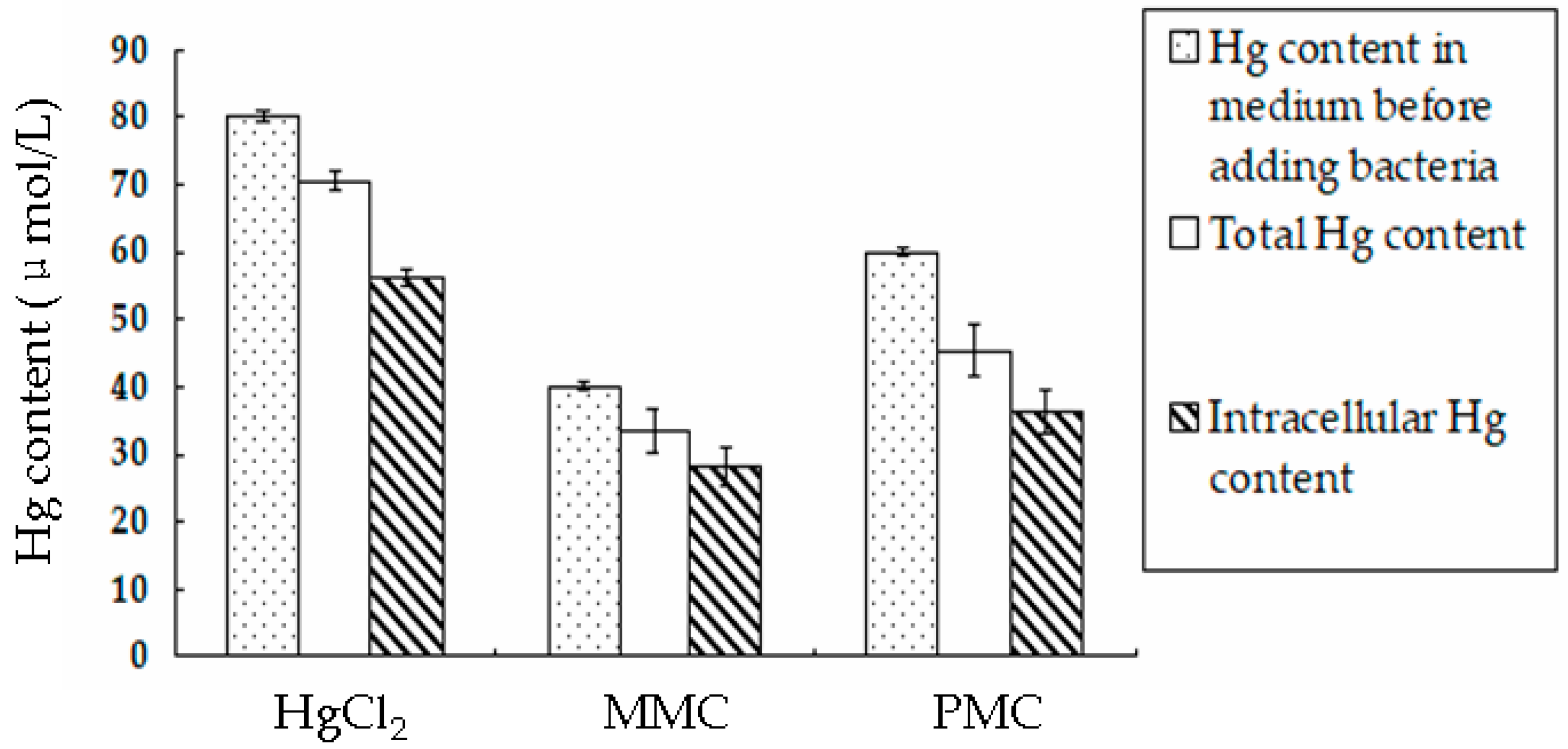
Figure 7.
Growth curves of BL21-7 cultured in PB media at different pH values.
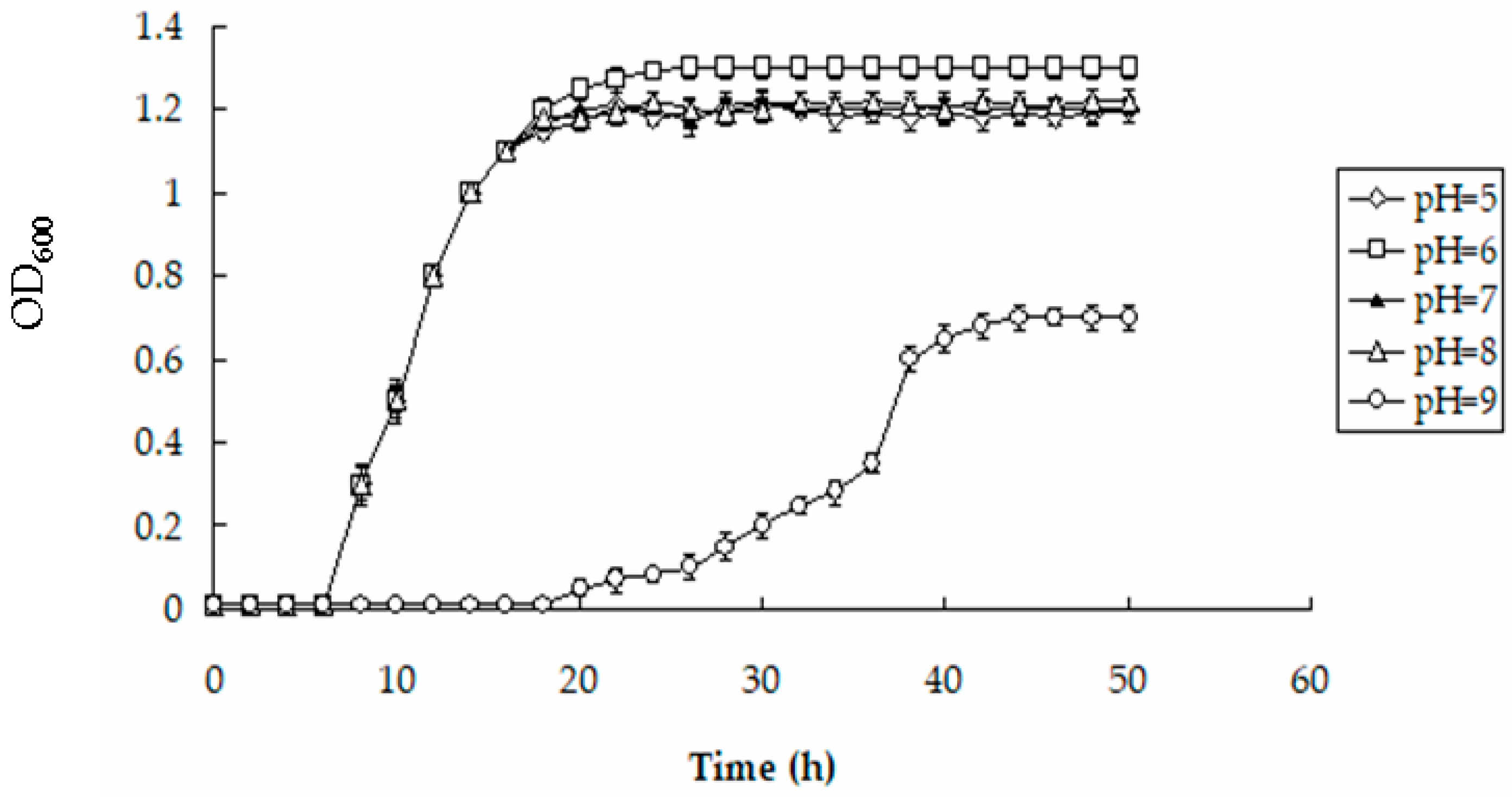
Figure 8.
Growth curves of BL21-7 cultured at different temperatures.

Figure 9.
Hg content in wastewater. CK indicates the Hg content in the wastewater sample without engineered bacteria. BL21-7 indicates the Hg content in the wastewater treated with BL21-7. BL21-0 indicates the Hg content in the wastewater treated with BL21-0.
Figure 9.
Hg content in wastewater. CK indicates the Hg content in the wastewater sample without engineered bacteria. BL21-7 indicates the Hg content in the wastewater treated with BL21-7. BL21-0 indicates the Hg content in the wastewater treated with BL21-0.

Figure 10.
Schematic mechanism proposed for organic and inorganic Hg detoxification by the constructed operon of BL21-7.
Figure 10.
Schematic mechanism proposed for organic and inorganic Hg detoxification by the constructed operon of BL21-7.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The primers used in this study.
| Primer | Sequence | Plasmid |
|---|---|---|
| PPPKF | 5′-ATAAGAATGCGGCCGCATTTACCACGTCCTGTGATT-3′; Not I restriction site is underlined. | p2 |
| PPPKR | 5′-CCGATCCGCTCGAGGGTTAATCGGGTTGCTCGAG-3′; Xho I restriction site is underlined. | p2 |
| P1F | 5′-CGACATATGGCTCCCCCGCCGTCGTTCAATGAGAATGGATAAGAGGCTCGTGGGATTGACGTGAGGGGGCAGGGATGGCTATATTTCTGGGAGCGAACTCCGGGCGAATACGAAGCGCTTGGATACAGTTGTAGGGAGGGATTTATCTTTTAACTTTAAGAAGGAGTGGCCAAGCGCTATTCGATCGACATATGGGAC-3′ Nde I and Bal I restriction sites are underlined. | p3 |
| P1R | 5′-GTCCCATATGTCGATCGAATAGCGCTTGGCCACTCCTTCTTAAAGTTAAAAGATAAATCCCTCCCTACAACTGTATCCAAGCGCTTCGTATTCGCCCGGAGTTCGCTCCCAGAAATATAGCCATCCCTGCCCCCTCACGTCAATCCCACGAGCCTCTTATCCATTCTCATTGAACGACGGCGGGGGAGCCATATGTCG-3′ Nde I and Bal I restriction sites are underlined. | p3 |
| P2F | 5′-CGGTGGCCAGTATGTCTGAACCACAAAAACG-3′; Bal I restriction site is underlined. | p4 |
| P2R | 5′-CGCGAATTCTTATCATTTATCGGCATAGGT-3′; EcoR I restriction site is underlined. | p4 |
| P3F | 5′-CCGGAATTCCGTGTTCTATAACGACAAAA-3′; EcoR I restriction site is underlined. | p5 |
| P3R | 5’-CCGGAATTCAAGAAAATATCGAAGAAAAAT-3′; EcoR I restriction site is underlined. | p5 |
Table 2.
The constructs of the artificial operons and the corresponding bacterial strains.
| Bacterial Strains | Plasmids Harboring Artificial Operons |
|---|---|
| BL21-0 | pET28a |
| BL21-1 | pET28a (g10-ppk-rpsT) |
| BL21-2 | pET28a (P16S-g10-ppk-rpsT) |
| BL21-T | pET28a (P16S-g10-merT-rpsT) |
| BL21-P | pET28a (P16S-g10-merP-rpsT) |
| BL21-B1 | pET28a (P16S-g10-merB1-rpsT) |
| BL21-B2 | pET28a (P16S-g10-merB2-rpsT) |
| BL21-TP | pET28a (P16S-g10-merT-merP-rpsT) |
| BL21-B1B2 | pET28a (P16S-g10-merB1-merB2-rpsT) |
| BL21-TPK | pET28a (P16S-g10-merT-merP-ppk-rpsT) |
| BL21-B1B2K | pET28a (P16S-g10-merB1-merB2-ppk-rpsT) |
| BL21-TPB1B2 | pET28a (P16S-g10-merT-merP-merB1-merB2-rpsT) |
| BL21-7 | pET28a (P16S-g10-merT-merP-merB1-merB2-ppk-rpsT) |
Table 3.
Mercury removed from LB medium in which BL21-7 was cultured for 5 h in the presence of other metals.
Table 3.
Mercury removed from LB medium in which BL21-7 was cultured for 5 h in the presence of other metals.
| Metals | HgCl2 Removed (µmol/L) | MMC Removed (µmol/L) | PMC Removed (µmol/L) |
|---|---|---|---|
| None | 8.80 ± 0.16 | 5.47 ± 0.28 | 7.80 ± 0.17 |
| Cd2+ | 8.66 ± 0.15 | 5.53 ± 0.23 | 7.76 ± 0.15 |
| Pb2+ | 8.85 ± 0.09 | 5.41 ± 0.25 | 7.73 ± 0.18 |
| Cu2+ | 8.95 ± 0.12 | 5.44 ± 0.26 | 7.83 ± 0.17 |
| Cd2+ + Pb2+ + Cu2+ | 8.98 ± 0.19 | 5.59 ± 0.27 | 7.85 ± 0.19 |
Table 4.
Mercury remaining in the medium after strains were cultured in LB medium.
| Strains | HgCl2 (µmol/L) | MMC (µmol/L) | PMC (µmol/L) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 5 h | 10 h | 15 h | 20 h | 0 h | 5 h | 10 h | 15 h | 20 h | 0 h | 5 h | 10 h | 15 h | 20 h | |
| BL21-T | 20.0 | 17.1 | 16.3 | 15.2 | 15.0 | 20.0 | 19.8 | 19.5 | 19.3 | 19.0 | 20.0 | 19.9 | 19.9 | 19.8 | 19.6 |
| BL21-B1 | 20.0 | 19.8 | 19.8 | 19.8 | 19.4 | 20.0 | 19.8 | 19.2 | 18.5 | 18.1 | 20.0 | 18.2 | 17.5 | 16.3 | 15.5 |
| BL21-B2 | 20.0 | 19.8 | 19.8 | 19.7 | 19.6 | 20.0 | 19.3 | 18.9 | 18.8 | 18.8 | 20.0 | 18.2 | 17.3 | 16.5 | 16.2 |
| BL21-B1B2 | 20.0 | 19.9 | 19.8 | 19.5 | 19.2 | 20.0 | 19.5 | 18.5 | 18.0 | 17.5 | 20.0 | 17.4 | 16.5 | 15.3 | 15.1 |
| BL21-TP | 20.0 | 14.8 | 13.2 | 11.5 | 11.1 | 20.0 | 19.9 | 19.7 | 19.5 | 19.5 | 20.0 | 19.9 | 19.9 | 19.8 | 19.5 |
| BL21-P | 20.0 | 19.6 | 18.5 | 18.3 | 18.0 | 20.0 | 19.9 | 19.8 | 19.8 | 19.5 | 20.0 | 19.8 | 19.7 | 19.5 | 19.5 |
| BL21-0 | 20.0 | 19.9 | 19.8 | 19.7 | 19.2 | 20.0 | 19.8 | 19.9 | 19.6 | 19.6 | 20.0 | 19.8 | 19.7 | 19.5 | 19.3 |
| BL21-1 | 20.0 | 19.8 | 19.7 | 19.9 | 19.5 | 20.0 | 19.9 | 19.5 | 19.4 | 19.4 | 20.0 | 19.9 | 19.8 | 19.5 | 19.2 |
| BL21-2 | 20.0 | 12.5 | 11.2 | 10.0 | 9.5 | 20.0 | 19.8 | 19.9 | 19.6 | 19.6 | 20.0 | 19.8 | 19.8 | 19.6 | 19.5 |
| BL21-B1B2K | 20.0 | 13.1 | 12.5 | 11.3 | 10.0 | 20.0 | 16.1 | 15.3 | 14.2 | 14.1 | 20.0 | 10.2 | 8.5 | 6.8 | 6.1 |
| BL21-TPK | 20.0 | 7.9 | 4.5 | 3.8 | 2.2 | 20.0 | 19.8 | 19.6 | 19.5 | 19.3 | 20.0 | 19.9 | 19.8 | 19.8 | 19.5 |
| BL21-7 | 20.0 | 6.8 | 3.7 | 2.5 | 1.1 | 20.0 | 6.7 | 3.2 | 1.4 | 1.1 | 20.0 | 5.2 | 2.3 | 1.5 | 1.0 |
| BL21-TPB1B2 | 20.0 | 14.6 | 13.2 | 12.3 | 12.2 | 20.0 | 19.5 | 19.2 | 18.3 | 17.2 | 20.0 | 17.5 | 17.1 | 15.2 | 15.1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chang, S.; Wu, Z.; Sun, W.; Shu, H. The Construction of an Engineered Bacterial Strain and Its Application in Accumulating Mercury from Wastewater. Appl. Sci. 2018, 8, 1572. https://doi.org/10.3390/app8091572
AMA Style
Chang S, Wu Z, Sun W, Shu H. The Construction of an Engineered Bacterial Strain and Its Application in Accumulating Mercury from Wastewater. Applied Sciences. 2018; 8(9):1572. https://doi.org/10.3390/app8091572
Chicago/Turabian StyleChang, Shenghe, Zhengjing Wu, Wei Sun, and Haiyan Shu. 2018. "The Construction of an Engineered Bacterial Strain and Its Application in Accumulating Mercury from Wastewater" Applied Sciences 8, no. 9: 1572. https://doi.org/10.3390/app8091572
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.