Social Interactions in Two Groups of Zoo-Housed Adult Female Asian Elephants (Elephas maximus) that Differ in Relatedness


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Elephant Management
2.2.1. Zoo A
2.2.2. Zoo B
2.2.3. Piloting
2.2.4. Behavioural Observations—Full Study
2.2.5. Elephant Identification and Observer Training
2.2.6. Social Behaviour Analysis
3. Results
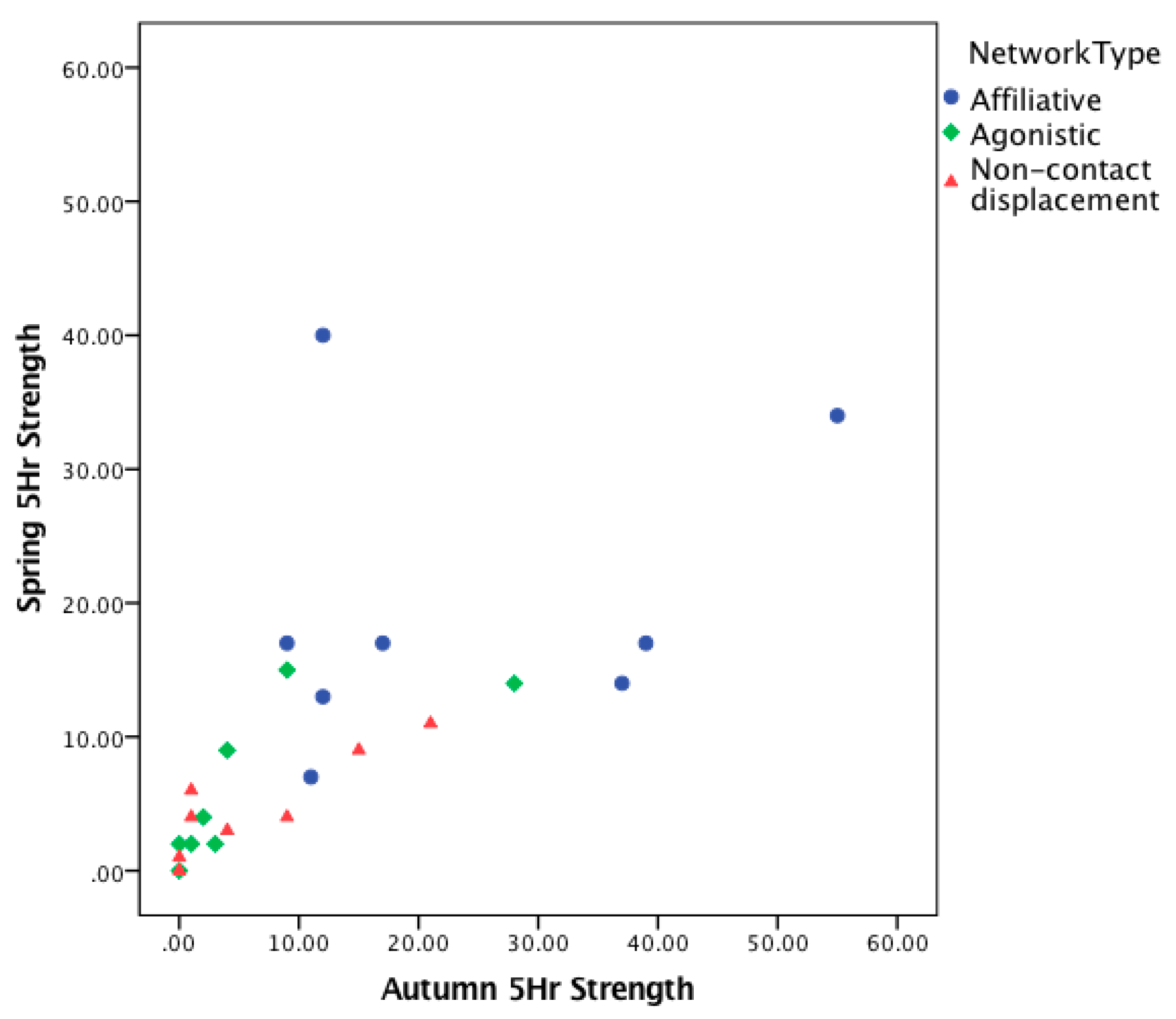
3.1. Consistency over Time
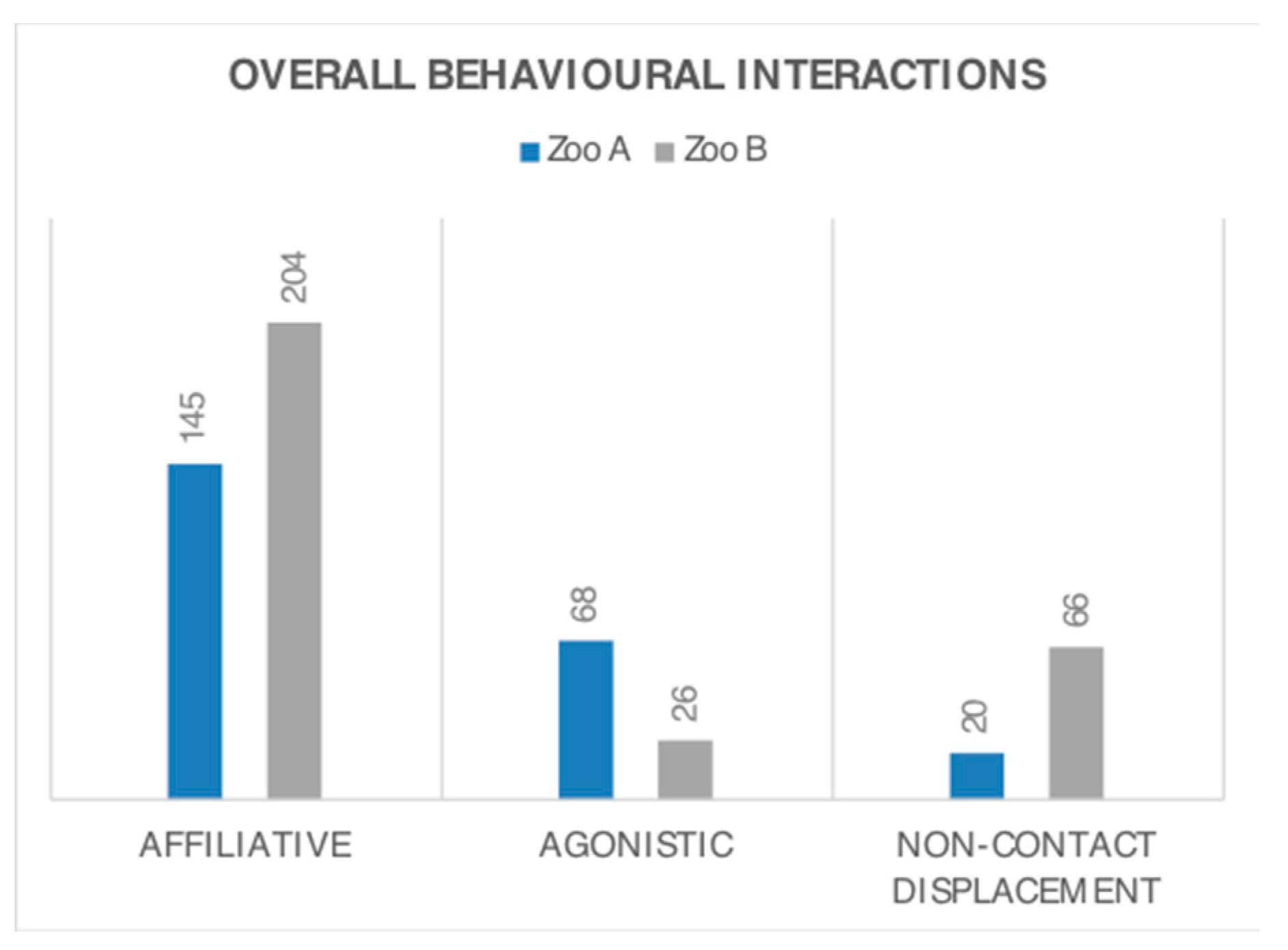
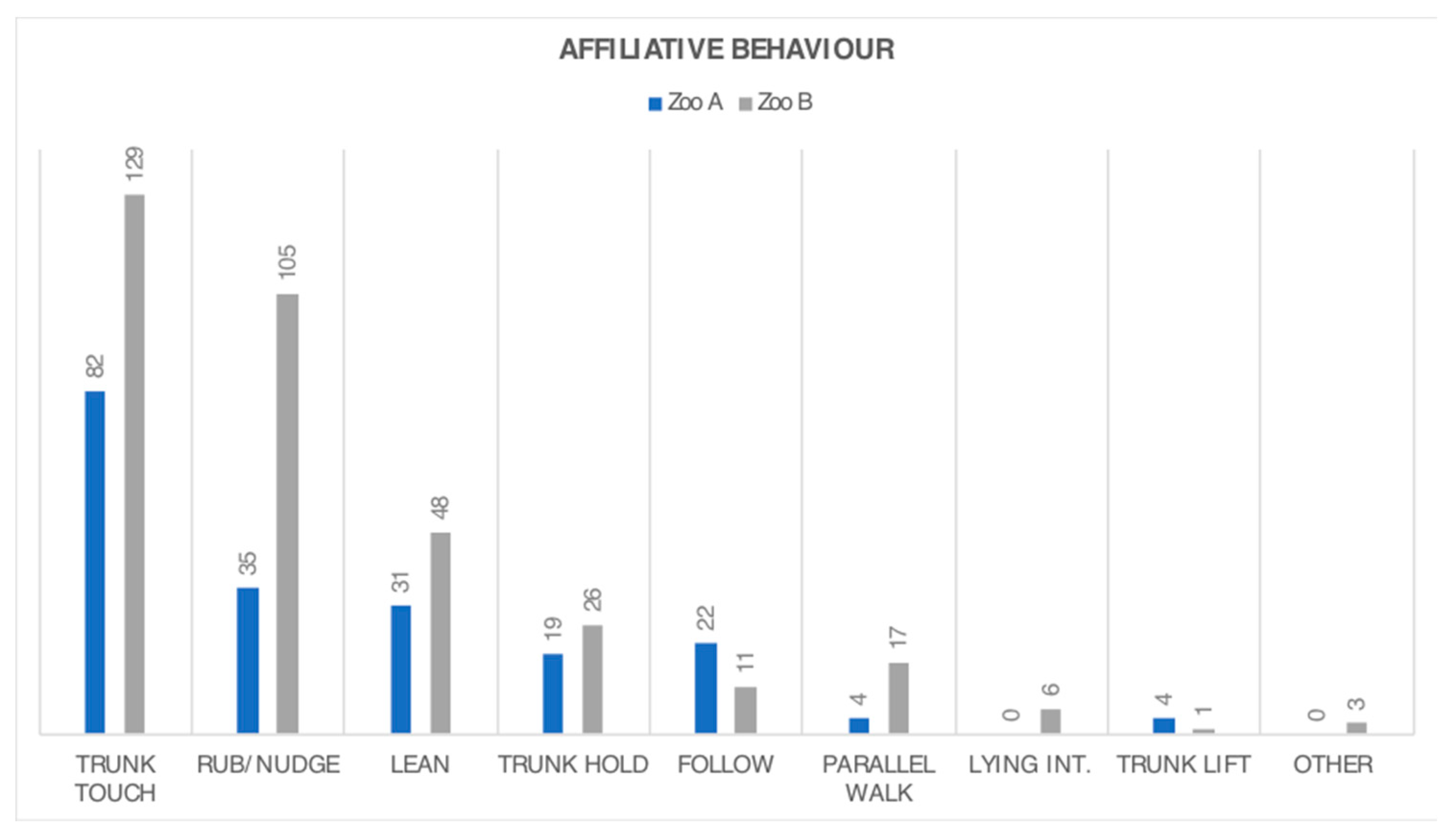
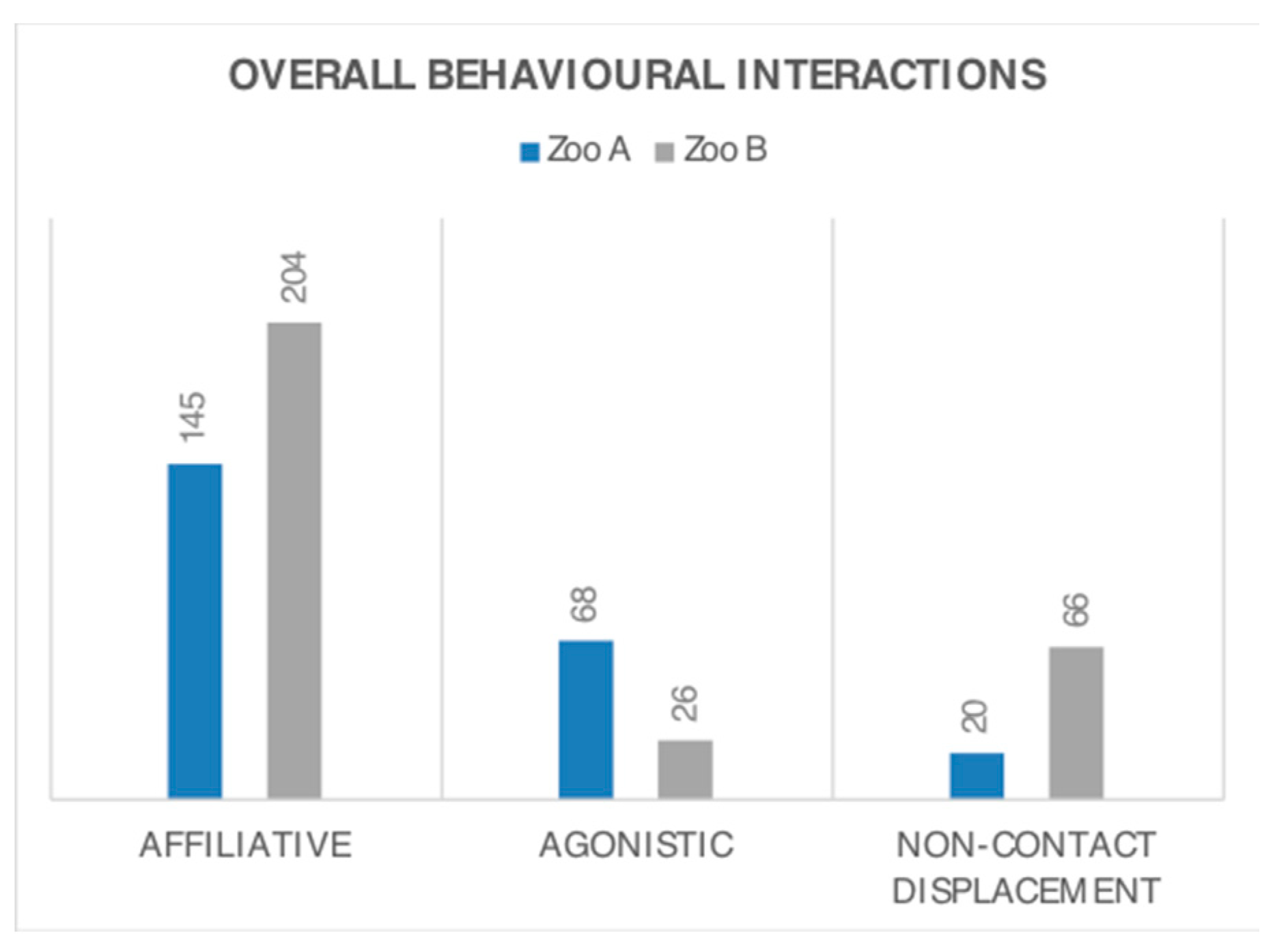
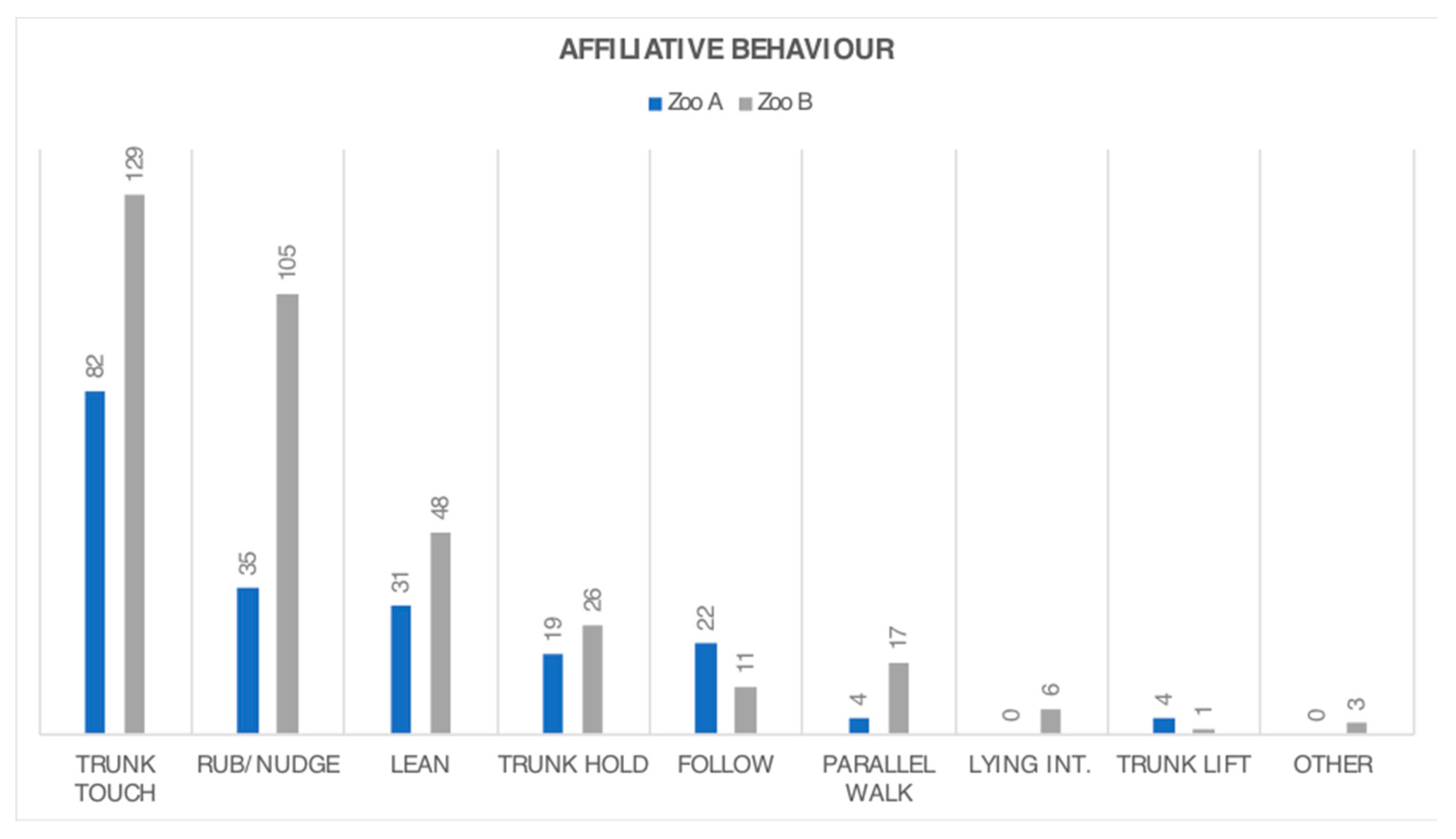
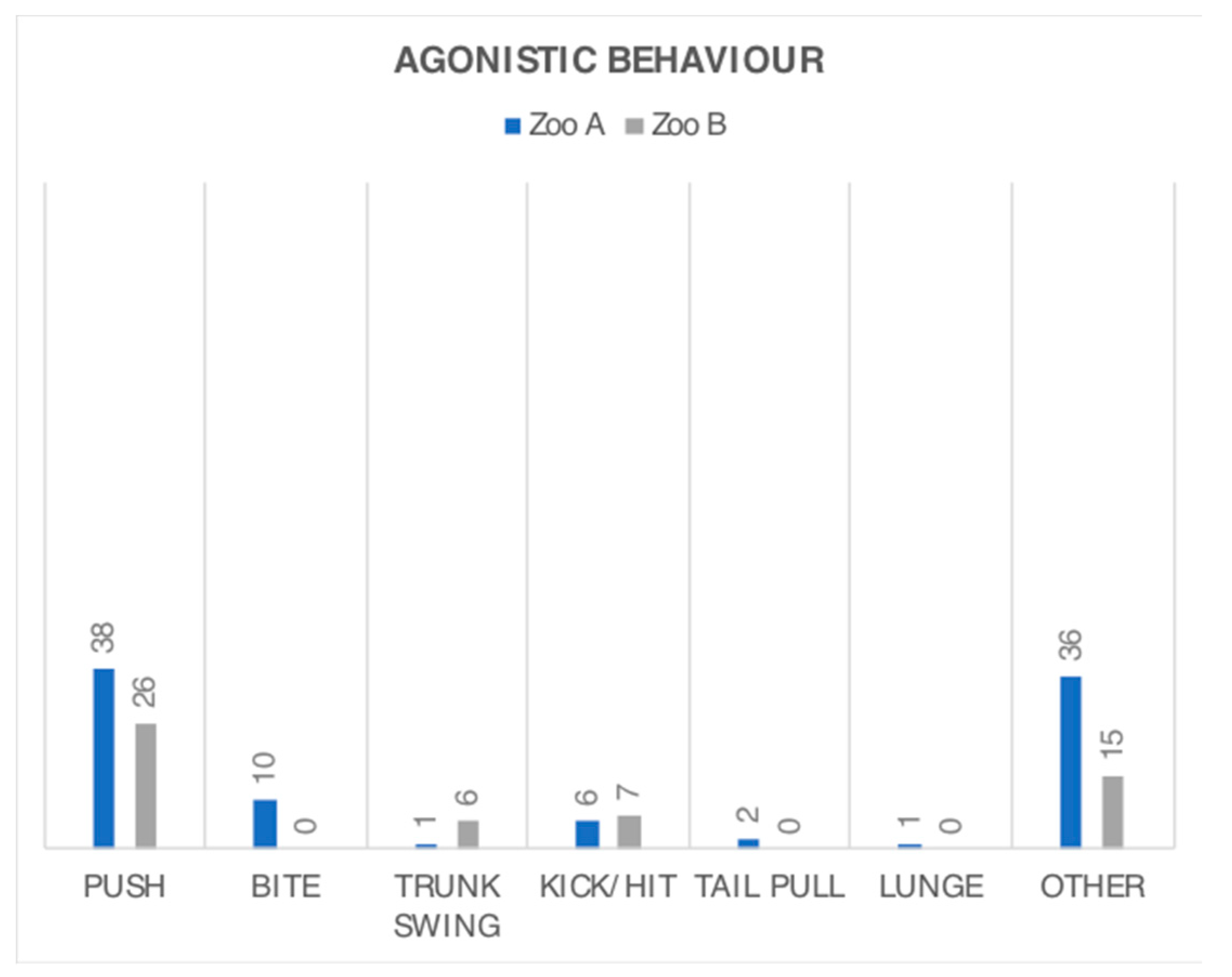
3.2. Behaviour Frequency
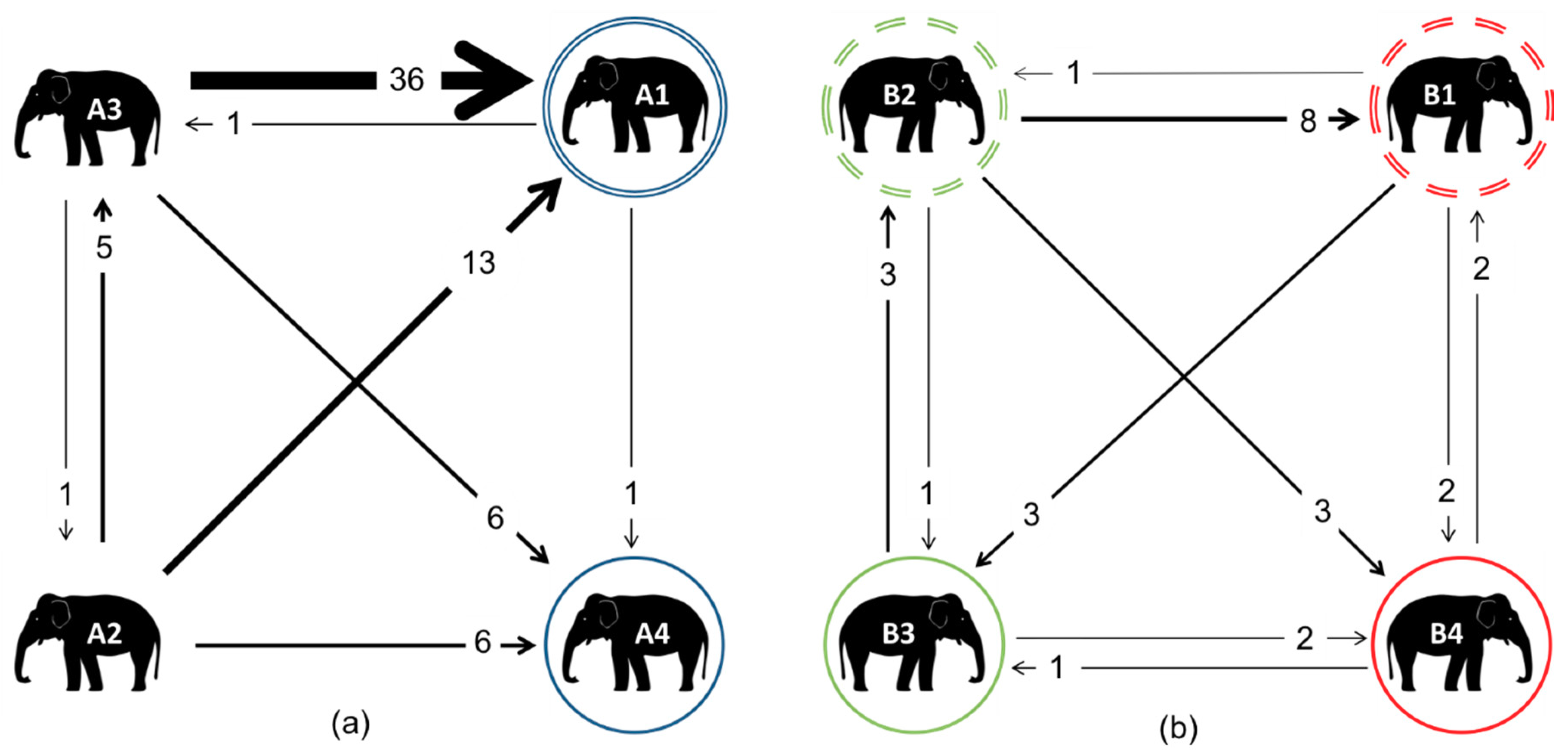
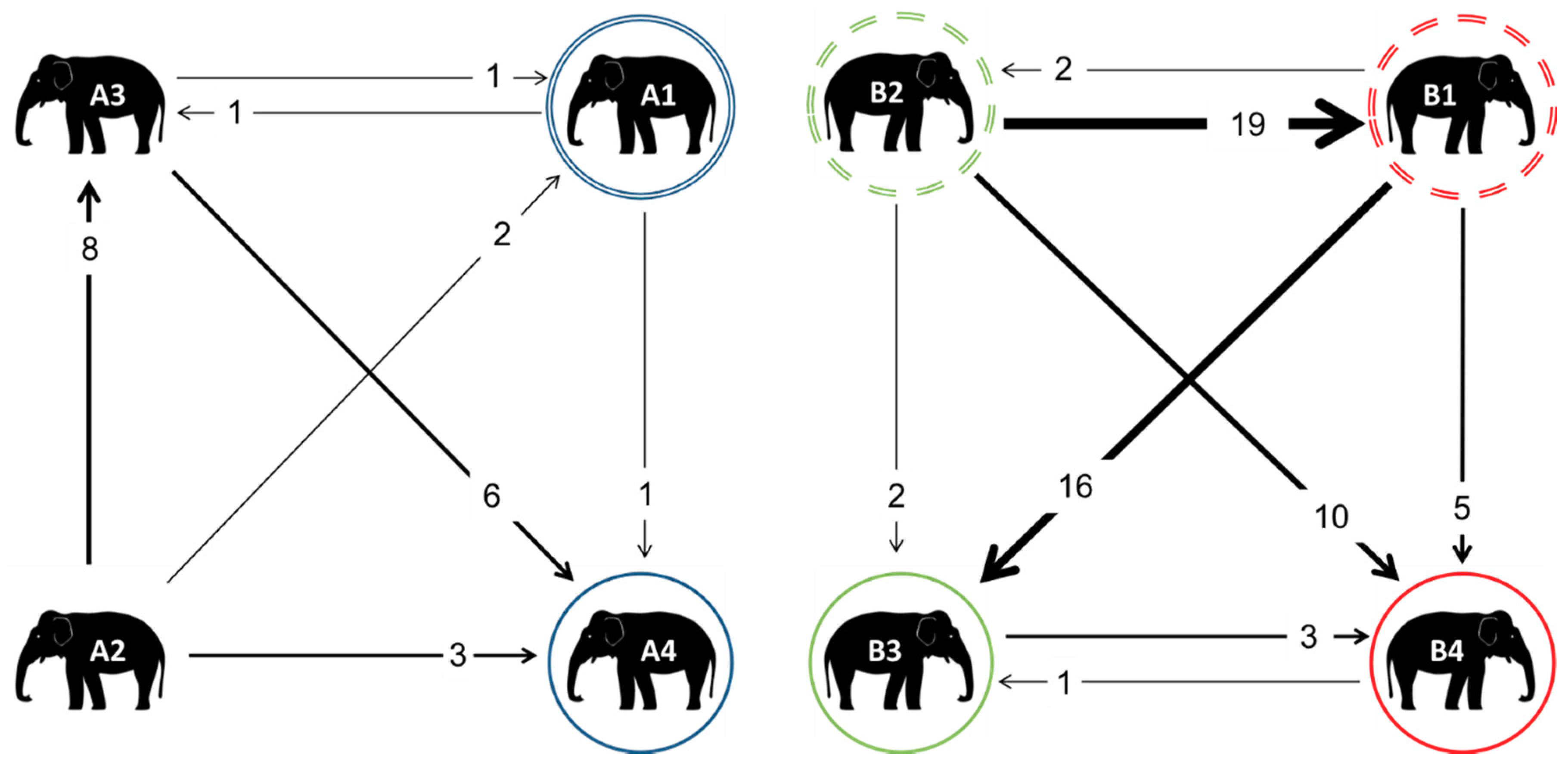
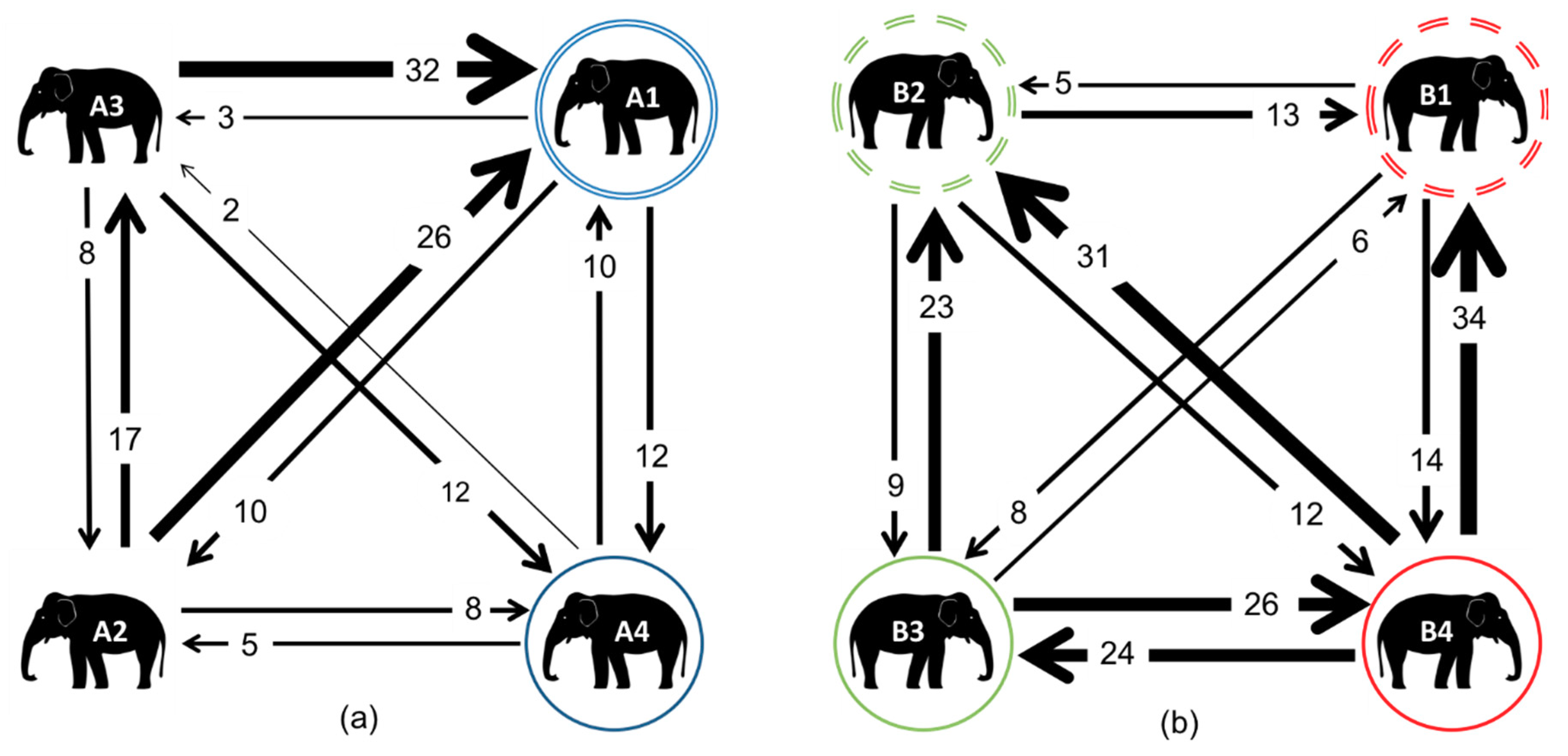
3.3. Affiliative Network
3.4. Agonistic Network
3.5. Non-Contact Displacement Network
4. Discussion
Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moss, C.J.; Poole, J.H. Relationships and social structure of African elephants. In Primate Social Relationships: An Integrated Approach; Hinde, R.A., Ed.; Blackwell Scientific Publications: Oxford, UK, 1983. [Google Scholar]
- Poole, J.H.; Moss, C.J. Elephant Sociality and Complexity: The Scientific Evidence; Johns Hopkins University Press: Baltimore, MD, USA, 2008; ISBN 9780801888182. [Google Scholar]
- Schulte, B.A. Social structure and helping behavior in captive elephants. Zoo Biol. 2000, 19, 447–459. [Google Scholar] [CrossRef]
- McKay, G.M. Behavior and ecology of the Asiatic elephant in Southeastern Ceylon. Smithson. Contrib. Zool. 1973. [Google Scholar] [CrossRef]
- Poole, J.H.; Granli, P. Signals, Gestures, and Behavior of African Elephants. In The Amboseli Elephants; University of Chicago Press: Chicago, IL, USA; pp. 109–124.
- Vidya, T.N.C.; Sukumar, R. Social and reproductive behaviour in elephants. Curr. Sci. 2005, 89, 1200–1207. [Google Scholar]
- Laursen, L.; Bekoff, M. Loxodonta africana. Mamm. Species 1978, 1. [Google Scholar] [CrossRef]
- Greco, B.J.; Meehan, C.L.; Heinsius, J.L.; Mench, J.A. Why pace? The influence of social, housing, management, life history, and demographic characteristics on locomotor stereotypy in zoo elephants. Appl. Anim. Behav. Sci. 2017, 194, 104–111. [Google Scholar] [CrossRef]
- Fernando, P.; Lande, R. Molecular genetic and behavioral analysis of social organization in the Asian elephant (Elephas maximus). Behav. Ecol. Sociobiol. 2000, 48, 84–91. [Google Scholar] [CrossRef]
- Vidya, T.N.C.; Sukumar, R. Social organization of the Asian elephant (Elephas maximus) in southern India inferred from microsatellite DNA. J. Ethol. 2005, 23, 205–210. [Google Scholar] [CrossRef]
- McComb, K.; Moss, C.; Durant, S.M.; Baker, L.; Sayialel, S. Matriarchs as repositories of social knowledge in African elephants. Science 2001, 292, 491–494. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.C. Allomothering among African elephants. Anim. Behav. 1987, 35, 278–291. [Google Scholar] [CrossRef]
- Poole, J.H.; Moss, C. Elephant mate searching: Group dynamics and vocal and olfactory communication. Symp. Zool. Soc. Lond. 1989, 61, 111–125. [Google Scholar]
- Gobush, K.S.; Wasser, S.K. Behavioural correlates of low relatedness in African elephant core groups of a poached population. Anim. Behav. 2009, 78, 1079–1086. [Google Scholar] [CrossRef]
- De Silva, S.; Wittemyer, G. A Comparison of Social Organization in Asian Elephants and African Savannah Elephants. Int. J. Primatol. 2012, 33, 1125–1141. [Google Scholar] [CrossRef]
- De Silva, S.; Ranjeewa, A.D.G.; Kryazhimskiy, S. The dynamics of social networks among female Asian elephants. BMC Ecol. 2011, 11, 17. [Google Scholar] [CrossRef] [PubMed]
- Wittemyer, G.; Getz, W.M. Hierarchical dominance structure and social organization in African elephants, Loxodonta africana. Anim. Behav. 2007, 73, 671–681. [Google Scholar] [CrossRef]
- Archie, E.A.; Moss, C.J.; Alberts, S.C. The ties that bind: Genetic relatedness predicts the fission and fusion of social groups in wild African elephants. Proc. R. Soc. B Biol. Sci. 2006, 273, 513–522. [Google Scholar] [CrossRef] [PubMed]
- De Silva, S.; Schmid, V.; Wittemyer, G. Fission–fusion processes weaken dominance networks of female Asian elephants in a productive habitat. Behav. Ecol. 2017, 28, 243–252. [Google Scholar] [CrossRef]
- Wittemyer, G.; Okello, J.B.A.; Rasmussen, H.B.; Arctander, P.; Nyakaana, S.; Douglas-Hamilton, I.; Siegismund, H.R. Where sociality and relatedness diverge: The genetic basis for hierarchical social organization in African elephants. Proc. Biol. Sci. 2009, 276, 3513–3521. [Google Scholar] [CrossRef] [PubMed]
- Garaï, M.E. Special Relationships between Female Asian Elephants (Elephas maximus) in Zoological Gardens. Ethology 2010, 90, 187–205. [Google Scholar] [CrossRef]
- Hutchins, M. Variation in nature: Its implications for zoo elephant management. Zoo Biol. 2006, 25, 161–171. [Google Scholar] [CrossRef]
- Harris, M.; Sherwin, C.; Harris, S. The Welfare, Housing and Husbandry of Elephants in UK Zoos; Final Report; University of Bristol: Bristol, UK, 2008. [Google Scholar]
- Clubb, R.; Mason, G. A Review of the Welfare of Zoo Elephants in Europe; University of Oxford: Oxford, UK, 2002. [Google Scholar]
- Asher, L.; Williams, E.; Yon, L. Developing Behavioural Indicators, as Part of a Wider Set of Indicators, to Assess the Welfare of Elephants in UK Zoos; University of Nottingham: Nottingham, UK, 2015. [Google Scholar]
- Rees, P.A. The sizes of elephant groups in zoos: Implications for elephant welfare. J. Appl. Anim. Welf. Sci. 2009, 12, 44–60. [Google Scholar] [CrossRef] [PubMed]
- Livet, J.; Jahn, T. Database of Asian Elephants. Available online: http://www.asianelephant.net/database.htm (accessed on 3 November 2015).
- Walter, O. Management Guidelines for the Welfare of Zoo Animals Elephants (Loxodonta Africana and Elephas Maximus), 3rd ed.; British and Irish Association of Zoos and Aquariums: London, UK, 2010. [Google Scholar]
- Coleing, A. The application of social network theory to animal behaviour. Biosci. Horiz. 2009, 2, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Poole, T.B.; Taylor, V.J.; Fernando, S.B.U.; Ratnasooriya, W.D.; Ratnayeke, A.; Lincoln, G.; McNeilly, A.; Manatunga, A.M.V.R. Social behaviour and breeding physiology of a group of Asian elephants Elephas maximus at the Pinnawala Elephant Orphanage, Sri Lanka. Int. Zoo Yearb. 1997, 35, 297–310. [Google Scholar] [CrossRef]
- Riley, J. The Social Relationships between Female African Elephants (Loxodonta Africana) at Knowsley Safari Park; Nottingham Trent University: Nottingham, UK, 2014. [Google Scholar]
- Makecha, R.; Fad, O.; Gardens, B.; Stan, U.S.A.; Ii, A.K. The Role of Touch in the Social Interactions of Asian Elephants (Elephas maximus). Int. J. Comp. Psychol. 2012, 25, 60–82. [Google Scholar]
- Veasey, J. Concepts in the care and welfare of captive elephants. Int. Zoo Yearb. 2006, 40, 63–79. [Google Scholar] [CrossRef]
- Martin, P.; Bateson, P. Measuring Behaviour: An Introductory Guide; Cambridge University Press: Cambridge, UK, 2007; ISBN 0521446147. [Google Scholar]
- Wasserman, S.; Faust, K. Social Network Analysis: Methods and Applications; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar] [CrossRef]
- Wey, T.; Blumstein, D.T.; Shen, W.; Jordán, F. Social network analysis of animal behaviour: A promising tool for the study of sociality. Anim. Behav. 2008, 75, 333–344. [Google Scholar] [CrossRef]
- Jacoby, D.M.P.; Fear, L.N.; Sims, D.W.; Croft, D.P. Shark personalities? Repeatability of social network traits in a widely distributed predatory fish. Behav. Ecol. Sociobiol. 2014, 68, 1995–2003. [Google Scholar] [CrossRef]
- Farine, D.R.; Whitehead, H. Constructing, conducting and interpreting animal social network analysis. J. Anim. Ecol. 2015, 84, 1144–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCowan, B.; Anderson, K.; Heagarty, A.; Cameron, A. Utility of social network analysis for primate behavioral management and well-being. Appl. Anim. Behav. Sci. 2008, 109, 396–405. [Google Scholar] [CrossRef]
- Drews, C. The Concept and Definition of Dominance in Animal Behaviour. Behaviour 1993, 125, 283–313. [Google Scholar] [CrossRef]
- Horová, E.; Brandlová, K.; Gloneková, M. The First Description of Dominance Hierarchy in Captive Giraffe: Not Loose and Egalitarian, but Clear and Linear. PLoS ONE 2015, 10, e0124570. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, J.W.S.; Blackwell, E.J.; Casey, R.A. Dominance in domestic dogs-useful construct or bad habit? J. Vet. Behav. Clin. Appl. Res. 2009, 4, 135–144. [Google Scholar] [CrossRef]
- Rowell, T.E. The concept of social dominance. Behav. Biol. 1974, 11, 131–154. [Google Scholar] [CrossRef]
- Bernstein, I.S. Dominance relationships and ranks: Explanations, correlations, and empirical challenges. Behav. Brain Sci. 1981, 4, 449–457. [Google Scholar] [CrossRef]
- Archie, E.A.; Morrison, T.A.; Foley, C.A.H.; Moss, C.J.; Alberts, S.C. Dominance rank relationships among wild female African elephants, Loxodonta africana. Anim. Behav. 2006, 71, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Appleby, M.C. The probability of linearity in hierarchies. Anim. Behav. 1983, 31, 600–608. [Google Scholar] [CrossRef]
- Rodrigues, M.A.; Boeving, E.R. Comparative social grooming networks in captive chimpanzees and bonobos. Primates 2018. [Google Scholar] [CrossRef] [PubMed]
- Stagni, E.; Normando, S.; de Mori, B. Distances between individuals in an artificial herd of African elephants (Loxodonta africana africana) during resource utilisation in a semi-captive environment. Res. Vet. Sci. 2017, 113, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Bonaparte-Saller, M.; Mench, J.A. Assessing the dyadic social relationships of female african (Loxodonta africana) and asian (Elephas maximus) zoo elephants using proximity, tactile contact, and keeper surveys. Appl. Anim. Behav. Sci. 2018, 199, 45–51. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zoo | Individual ID | Origin | Age |
|---|---|---|---|
| A | A1 | Wild caught | 31 (est) |
| A | A2 | Wild caught | 31(est) |
| A | A3 | Captive bred | 19 |
| A | A4 | Captive bred | 17 |
| B | B1 | Captive bred | 31 |
| B | B2 | Captive bred | 24 |
| B | B3 | Captive bred | 12 |
| B | B4 | Captive bred | 8 |
| Type | Behaviour | Descriptions |
|---|---|---|
| Affiliative | Leaning | Leaning on another elephant |
| Affiliative | Lying interaction | Standing over another elephant—usually one that is lying down or young; Placing at least one foot on top of another elephant—usually one that is lying down; Sitting in a crouched position on top of another elephant that is in lying rest |
| Affiliative | Trunk touch | Putting the trunk in the mouth of another elephant; Touching another elephant, not the mouth or genitals, with the trunk in a non-aggressive manner; Touching the genital area of another elephant with the trunk; |
| Affiliative | Trunk hold | Intertwining of trunks between two elephants |
| Affiliative | Body rub or Nudge | Gentle physical contact between two elephants, which may be head–head, head–body, or body–body (not including touching with trunk); Rubbing the body against another elephant |
| Affiliative | Parallel walk | Two elephants walking side by side in a non-aggressive manner, for three or more steps |
| Affiliative | Follow | One elephant walks closely behind (within two elephant body lengths) of another elephant |
| Affiliative | Offer food | One elephant pushes a pile of food towards another elephant, looks like an offering of the resource |
| Affiliative | Trunk Lift | Trunk is outstretched and raised towards an approaching individual |
| Affiliative | Play | Engaging in active play with another elephant, including head-to-head sparring, trunk wrestling, mounting, chasing, and rolling on one another. Does not include behaviour observed following an antagonistic encounter or as part of courtship |
| Agonistic | Kick or hit | Strike out or hit an elephant with a foot in a seemingly aggressive manner—may include kicking of sand towards another elephant; Hitting another elephant with the trunk or tail |
| Agonistic | Charge | Move towards another elephant with the head held high, pace usually quickens as individual gets closer to the target elephant |
| Agonistic | Chase | Charge leading to pursuit of another elephant |
| Agonistic | Push | One elephant forces or pushes against the body (usually the rump) of another elephant, resulting in the elephant that is being pushed moving at least two steps |
| Agonistic | Stand off | Two elephants standing facing in opposite directions with foreheads pushing against each other |
| Agonistic | Floor smack | Hitting the trunk on the floor in an aggressive manner, may be accompanied by a ‘snort’ |
| Agonistic | Lunge | A lunging motion followed by physical contact, used to prevent another elephant standing up |
| Agonistic | Tusking | Poking or jabbing at another elephant with the tusk |
| Agonistic | Tail Pulling | Sharply pulling the tail of another elephant |
| Agonistic | Directed trunk swing | Head oriented towards another elephant, human, or change in the environment, violently swinging the trunk around in an aggressive display |
| Agonistic | Contact displacement | Movement of one elephant resulting in another elephant leaving its location (within 10 s) caused by physical contact between individuals such as a push or nudge |
| Agonistic | Aggressive display: standing | Facing another elephant in an aggressive posture; Head held high, Ears wide or flapping |
| Agonistic | Aggressive display: walking | Facing another elephant while walking with head bobbing up and down or side-to-side, ears wide or flapping |
| Agonistic | Size up | Two elephants directly facing each other, standing as tall as possible, heads raised, and ears spread wide |
| Agonistic | Bite | Biting of the body, trunk or tail of another elephant |
| Non-contact displacement | NCD | Movement of one elephant towards the other, resulting in another elephant leaving its location (within 10 s)—no physical contact occurs between elephants |
| Other | Describe |
| Term | Definition |
|---|---|
| Dyad | A pair of individuals |
| Node | Represents an individual or study subject |
| Tie | Represents the social interactions between two nodes |
| Network | A collection of ties between nodes |
| Asymmetric matrix | A grid containing each node along the horizontal and vertical axes. The horizontal axes represent the initiating elephant, and the vertical axes represent the receiving elephant. The frequency of an interaction from the initiator to the receiver is recorded in the intersecting square for that dyad. |
| Digraph | A graph in which nodes are connected by social ties. An arrow is used to show who initiated and received the social interaction within the dyad. |
| Weighted digraph | A graph in which the arrows are weighted with the frequency or strength of the social interaction. Each weighted tie has a direction to indicate who initiated and received the social interaction within a dyad. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harvey, N.D.; Daly, C.; Clark, N.; Ransford, E.; Wallace, S.; Yon, L. Social Interactions in Two Groups of Zoo-Housed Adult Female Asian Elephants (Elephas maximus) that Differ in Relatedness. Animals 2018, 8, 132. https://doi.org/10.3390/ani8080132
Harvey ND, Daly C, Clark N, Ransford E, Wallace S, Yon L. Social Interactions in Two Groups of Zoo-Housed Adult Female Asian Elephants (Elephas maximus) that Differ in Relatedness. Animals. 2018; 8(8):132. https://doi.org/10.3390/ani8080132
Chicago/Turabian StyleHarvey, Naomi D., Carolyn Daly, Natasha Clark, Eleanor Ransford, Stefanie Wallace, and Lisa Yon. 2018. "Social Interactions in Two Groups of Zoo-Housed Adult Female Asian Elephants (Elephas maximus) that Differ in Relatedness" Animals 8, no. 8: 132. https://doi.org/10.3390/ani8080132