
Social Licking in Pregnant Dairy Heifers

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Experimental Section
2.1. Data Collection

{kind=link}
| Variables | Description |
|---|---|
| Groomer | The animal that licks the body of another animal |
| Recipient | The animal that has its body licked |
| Body part licked | Head, neck, front part of the body (i.e. all body parts from thoracic vertebrae to chest) or back part of the body (from lumbar, sacral and coccygeal vertebrae to abdomen) as described by Val-Laillet et al. [11] |
| Solicited social licking | Animal has head lowered, chin stretched under the head of another animal, sometimes accompanied by head butts a |
| Forced termination | Groomer stops licking the recipient after receiving a physical interaction from the recipient or another animal (butt or push) |
| Duration | Length (in time) of a licking event that was initiated when the tongue touches the body surface of the other animal and ended when the tongue stops touching it. When breaks were <30 s events were compiled into the same bout as suggested by Val-Laillet et al. [11] |
2.2. Data Analysis
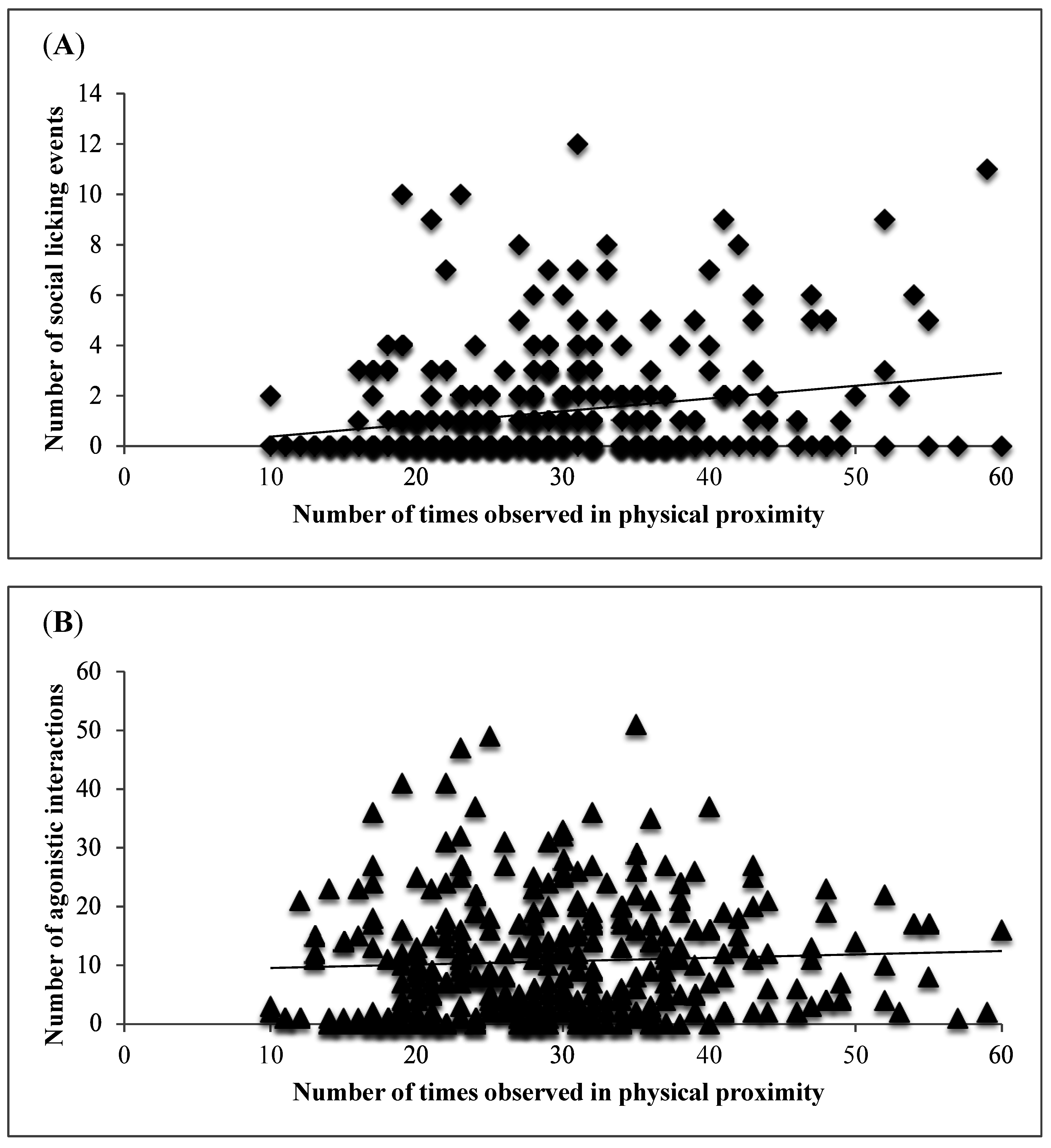
3. Results and Discussion
| Variables | Pasture | Free-stall | P |
|---|---|---|---|
| Total number of social licking events per group | 10 ± 10 | 67 ± 10 | 0.01 |
| Social licking (% of all social interactions) | 8 ± 2 | 12 ± 2 | 0.14 |
| Cows acting as (% of animals): | |||
| Groomer | 54 ± 7 | 81 ± 7 | 0.03 |
| Recipient | 56 ± 9 | 87 ± 9 | 0.01 |
| Both | 60 ± 14 | 70 ± 14 | 0.58 |
| Duration (s) | 39 ± 14 | 37 ± 13 | 0.91 |
| Solicited social licking (% of licking events) | 36 | 29 | 0.14 |
| Body part licked (% of licking events) | 0.09 | ||
| Head | 46 | 47 | |
| Neck | 25 | 35 | |
| Front | 3 | 7 | |
| Back | 25 | 10 | |
| Behavior pre-licking (% of licking events) | 0.19 | ||
| Grazing or feeding | 71 | 60 | |
| Idle | 21 | 33 | |
| Other | 8 | 7 | |
| Forced termination (%) | 30 | 13 | <0.01 |
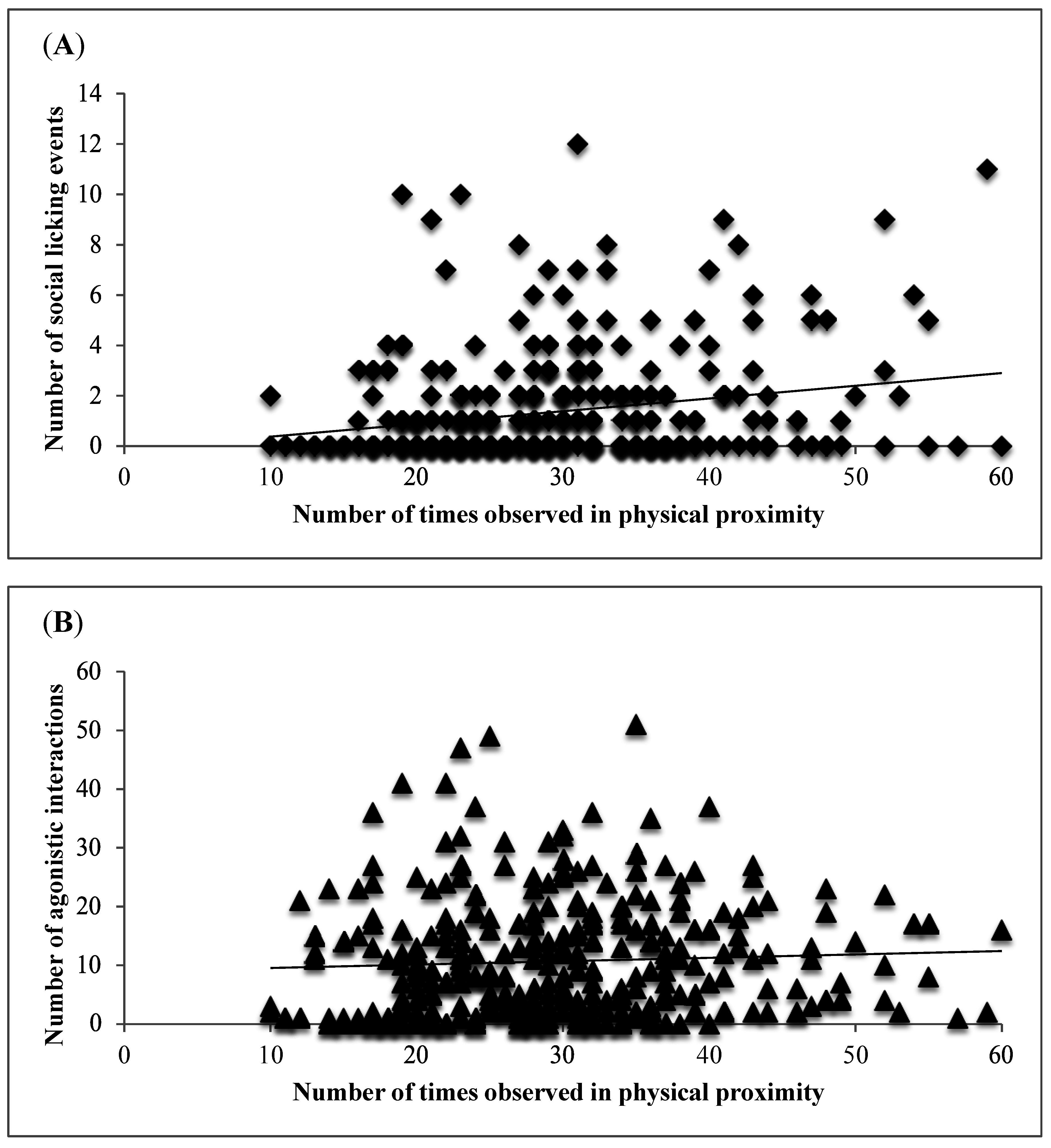
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Boissy, A.; Manteuffel, G.; Jensen, M.B.; Moe, R.O.; Spruijt, B.; Keeling, L.J.; Winckler, C.; Forkman, B.; Dimitrov, I.; Langbein, J.; et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 2007, 92, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Newby, N.C.; Duffield, T.F.; Pearl, D.L.; Leslie, K.E.; Leblanc, S.J.; von Keyserlingk, M.A. Short communication: Use of a mechanical brush by holstein dairy cattle around parturition. J. Dairy Sci. 2013, 96, 2339–2344. [Google Scholar] [CrossRef] [PubMed]
- Bouissou, M.F.; Boissy, A.; Le Neindre, P.; Veissier, I. The Social Behaviour of Cattle; CABI: Wellingford, UK, 2001; p. 424. [Google Scholar]
- Reinhardt, V.; Reinhardt, A. Cohesive relationships in a cattle herd (Bos indicus). Behaviour 1981, 77, 121–151. [Google Scholar] [CrossRef]
- Dunbar, R.I.M.; Schultz, S. Bondedness and sociality. Behaviour 2010, 147, 775–853. [Google Scholar] [CrossRef]
- Sato, S.; Tarumizu, K.; Hatae, K. The influence of social factors on allogrooming in cows. Appl. Anim. Behav. Sci. 1993, 38, 235–244. [Google Scholar] [CrossRef]
- Dunbar, R.I.M. The social role of touch in humans and primates: Behavioural functions and neurobiological mechanisms. Neurosci. Biobehav. Rev. 2010, 34, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro Machado, T.M. Comportamentos Afiliativos em Vacas Leiteiras a Pasto, o Papel da Lambida; Universidade Federal de Santa Catarina: Florianopolis, Brazil, 2009. [Google Scholar]
- Laister, S.; Stockinger, B.; Regner, A.-M.; Zenger, K.; Knierim, U.; Winckler, C. Social licking in dairy cattle—Effects on heart rate in performers and receivers. Appl. Anim. Behav. Sci. 2011, 130, 81–90. [Google Scholar] [CrossRef]
- Sato, S. Social licking pattern and its relationships to social dominance and live weight gain in weaned calves. Appl. Anim. Behav. Sci. 1984, 32, 3–12. [Google Scholar] [CrossRef]
- Val-Laillet, D.; Guesdon, V.; von Keyserlingk, M.A.G.; de Passillé, A.M.; Rushen, J. Allogrooming in cattle: Relationships between social preferences, feeding displacements and social dominance. Appl. Anim. Behav. Sci. 2009, 116, 141–149. [Google Scholar] [CrossRef]
- Sato, S.; Tarumizu, K. Heart rates before, during and after allogrooming in cattle (Bos taurus). J. Ethol. 1993, 11, 149–150. [Google Scholar] [CrossRef]
- Gygax, L.; Neisen, G.; Wechsler, B. Socio-spatial relationships in dairy cows. Ethology 2010, 116, 10–23. [Google Scholar] [CrossRef]
- Takeda, K.; Sato, S.; Sugawara, K. The number of farm mates influences social and maintenance behaviours of Japanese black cows in a communal pasture. Appl. Anim. Behav. Sci. 2000, 67, 181–192. [Google Scholar] [CrossRef]
- Huber, R.; Baumung, R.; Wurzinger, M.; Semambo, D.; Mwai, O.; Winckler, C. Grazing, social and comfort behaviour of Ankole and crossbred (Ankole × Holstein) heifers on pasture in south western Uganda. Appl. Anim. Behav. Sci. 2008, 112, 223–234. [Google Scholar] [CrossRef]
- Lim, M.M.; Young, L.J. Neuropeptidergic regulation of affiliative behavior and social bonding in animals. Hormon. Behav. 2006, 50, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Berscheid, E.; Snyder, M.; Omoto, A.M. The relationship closeness inventory—Assessing the closeness of interpersonal relationships. J. Personal. Soc. Psychol. 1989, 57, 792–807. [Google Scholar] [CrossRef]
- Miller, K.; Wood-Gush, D.G.M. Some effects of housing on the social behaviour of dairy cows. Anim. Sci. 1991, 53, 271–278. [Google Scholar] [CrossRef]
- Proudfoot, K.L.; Vieira, D.M.; Weary, D.M.; von Keyserlingk, M.A.G. Competition at the feed bunk changes the feeding, standing, and social behavior of transition dairy cows. J. Dairy Sci. 2009, 92, 3116–3123. [Google Scholar] [CrossRef] [PubMed]
- Huzzey, J.M.; DeVries, T.J.; Valois, P.; von Keyserlingk, M.A.G. Stocking density and feed barrier design affect the feeding and social behavior of dairy cattle. J. Dairy Sci. 2006, 89, 126–133. [Google Scholar] [CrossRef]
- Fregonesi, J.A.; Veira, D.M.; von Keyserlingk, M.A.; Weary, D.M. Effects of bedding quality on lying behavior of dairy cows. J. Dairy Sci. 2007, 90, 5468–5472. [Google Scholar] [CrossRef] [PubMed]
- Krohn, C.C. Behaviour of dairy cows kept in extensive (loose housing/pasture) or intensive (tie stall) environments. III. Grooming, exploration and abnormal behaviour. Appl. Anim. Behav. Sci. 1994, 42, 73–86. [Google Scholar] [CrossRef]
- NAHMS. Dairy 2007: Facility Characteristics and Cow Comfort on U.S. Dairy Operations, 2007; Centers for Epidemiology and Animal Health, USDA: Fort Collins, CO, USA, 2010; p. 169. [Google Scholar]
- Schuppli, C.A.; Von Keyserlingk, M.A.; Weary, D.M. Access to pasture for dairy cows: Responses from an on-line engagement. J. Anim. Sci. 2014, 92, 5185–5192. [Google Scholar] [CrossRef] [PubMed]
- Canadian Council of Animal Care (CCAC). Canadian Council of Animal Care Guidelines on the Care and Use of Farm Animals in Research, Teaching And Testing; CCAC: Ottawa, ON, Canada, 2009. [Google Scholar]
- NRC. Nutrient Requirements of Dairy Cattle, 7th ed.; National Academy Press: Washington, DC, USA, 2001. [Google Scholar]
- Altmann, J. Observational study of behavior sampling methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef] [PubMed]
- Lehner, P.N. Handbook of Ethological Methods, 2nd ed.; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Fraser, A. The Behaviour of the Horse; CABI: Wallingford, UK, 1992; p. 288. [Google Scholar]
- Hurnik, J.F.; Lewis, N.J.; Taylor, A.; Pinheiro Machado Filho, L.C. Farm Animal Behavior Laboratory Manual; Department of Animal and Poultry Science, University of Guelph: Guelph, ON, Canada, 1995. [Google Scholar]
- Kondo, S.; Hurnik, J.F. Stabilization of social hierarchy in dairy cows. Appl. Anim. Behav. Sci. 1990, 27, 287–297. [Google Scholar] [CrossRef]
- Coimbra, P.A.D.; Machado Filho, L.C.P.; Hötzel, M.J. Effects of social dominance, water trough location and shade availability on drinking behaviour of cows on pasture. Appl. Anim. Behav. Sci. 2012, 139, 175–182. [Google Scholar] [CrossRef]
- Sato, S.; Sako, S.; Maeda, A. Social licking patterns in cattle (Bos taurus)—Influence of environmental and social factors. Appl. Anim. Behav. Sci. 1991, 32, 3–12. [Google Scholar] [CrossRef]
- Schütz, K.E.; Rogers, A.R.; Poulouin, Y.A.; Cox, N.R.; Tucker, C.B. The amount of shade influences the behavior and physiology of dairy cattle. J. Dairy Sci. 2010, 93, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Pérez, A.P.; Veà Baró, J.J. Does allogrooming serve a hygienic function in cercocebus torquatus lunulatus? Am. J. Primatol. 1999, 49, 223–242. [Google Scholar] [CrossRef]
- Spruijt, B.M.; van Hooff, J.A.; Gispen, W.H. Ethology and neurobiology of grooming behavior. Physiol. Rev. 1992, 72, 825–852. [Google Scholar] [PubMed]
- Wood, M.T. Social grooming patterns in two herds of monozygotic twin dairy cows. Anim. Behav. 1977, 25, 635–642. [Google Scholar] [CrossRef]
- Aureli, F.; Preston, S.D.; de Waal, F.B.M. Heart rate responses to social interactions in free-moving Rhesus macaques (Macaca mulatto)—A pilot study. J. Comp. Psychol. 1999, 113, 59–65. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tresoldi, G.; Weary, D.M.; Pinheiro Machado Filho, L.C.; Von Keyserlingk, M.A.G. Social Licking in Pregnant Dairy Heifers. Animals 2015, 5, 1169-1179. https://doi.org/10.3390/ani5040404
Tresoldi G, Weary DM, Pinheiro Machado Filho LC, Von Keyserlingk MAG. Social Licking in Pregnant Dairy Heifers. Animals. 2015; 5(4):1169-1179. https://doi.org/10.3390/ani5040404
Chicago/Turabian StyleTresoldi, Grazyne, Daniel M. Weary, Luiz Carlos Pinheiro Machado Filho, and Marina A. G. Von Keyserlingk. 2015. "Social Licking in Pregnant Dairy Heifers" Animals 5, no. 4: 1169-1179. https://doi.org/10.3390/ani5040404