The Varied Role of Efflux Pumps of the MFS Family in the Interplay of Bacteria with Animal and Plant Cells

 ,
,
Abstract
:1. Introduction
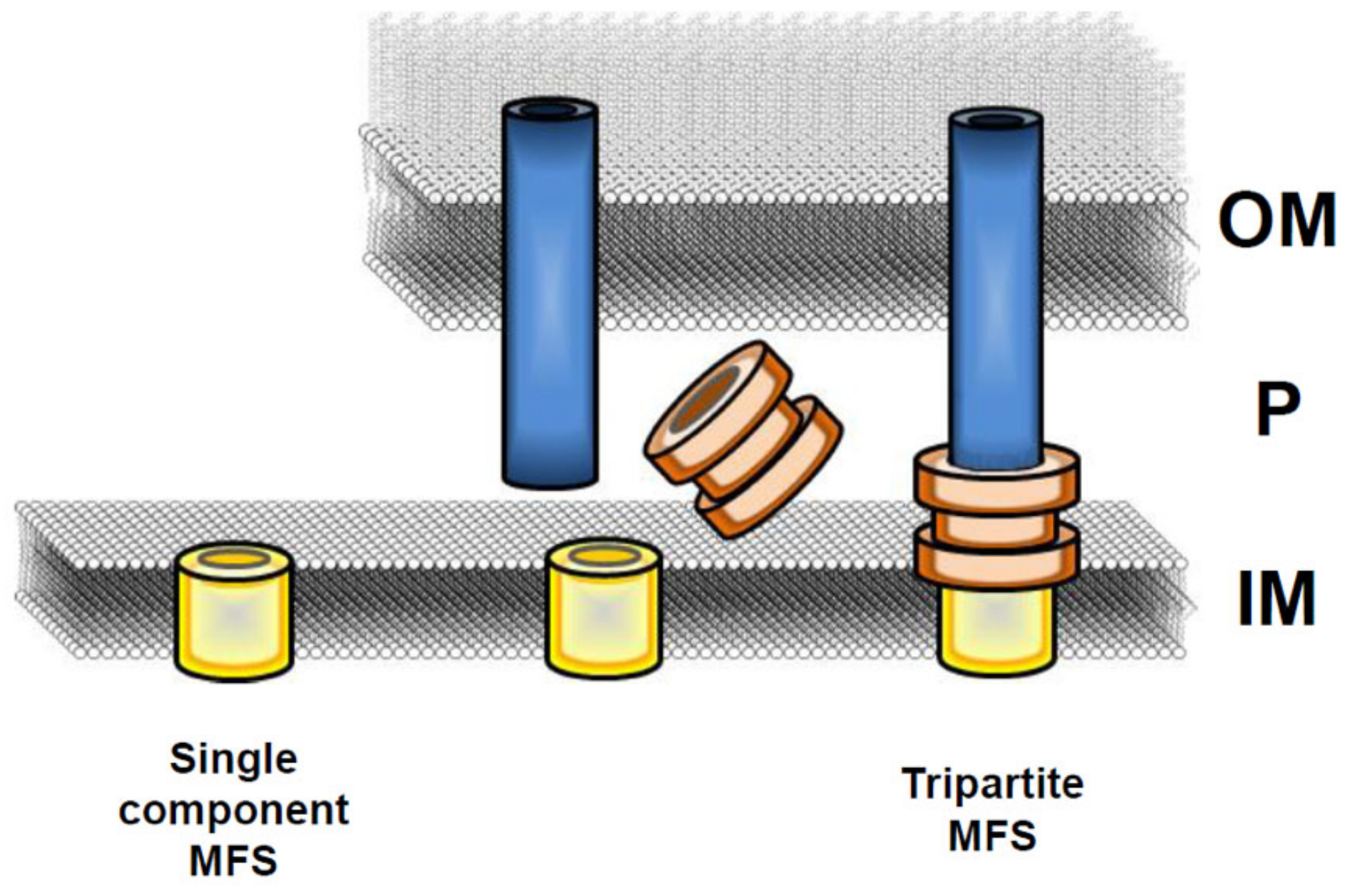
2. Organization and Evolution of the MFS EPs
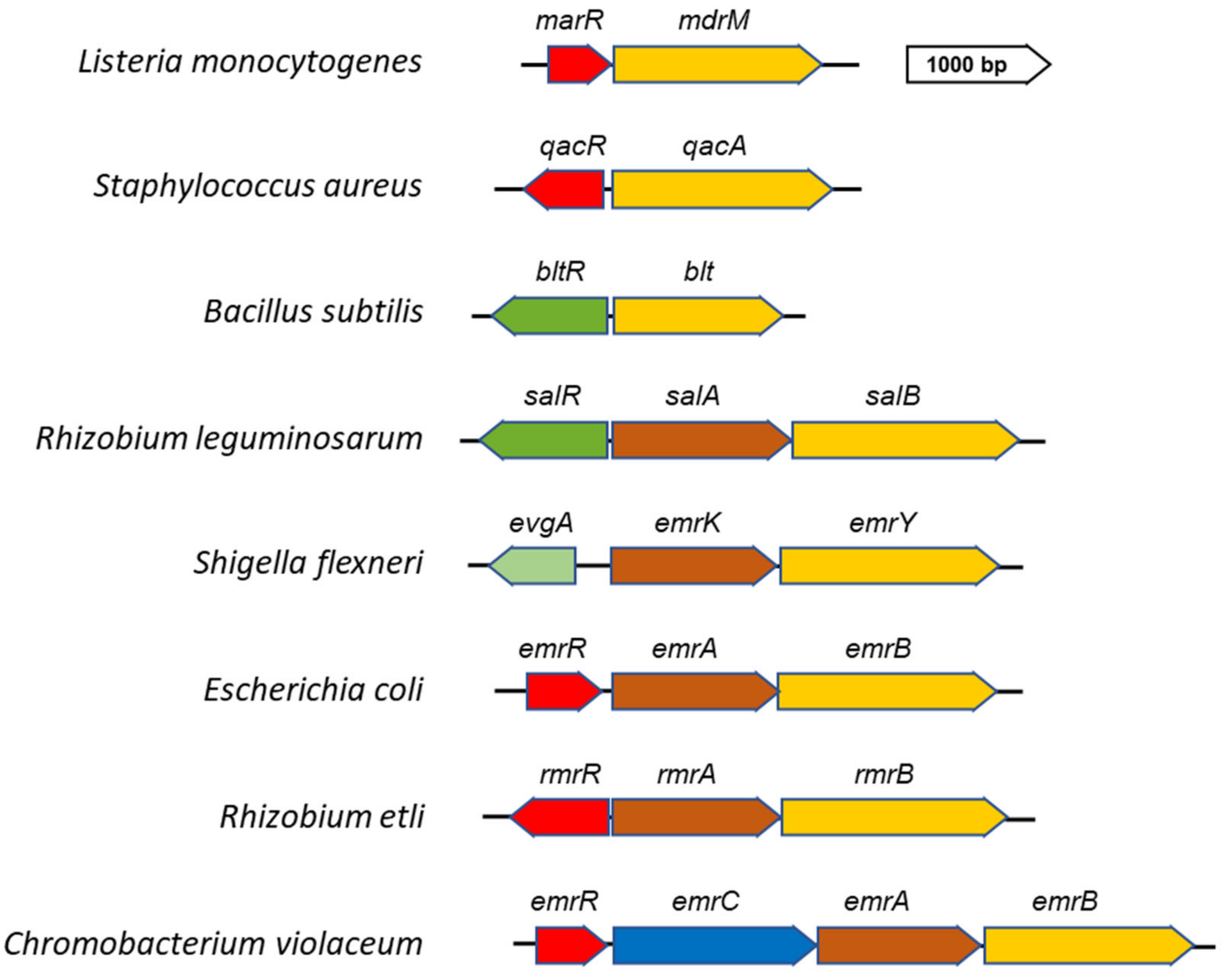
3. Transcriptional Strategies for the Regulation of MFS EPs
4. Role of MFS EPs in Interactions Between Bacteria and Plant Cells
5. Role of the MFS EPs in the Virulence of Bacterial Pathogens in Animal Hosts
6. Role of the MFS EPs in Bacterial Communication and Biofilm Formation
7. Conclusions
Funding
Conflicts of Interest
References
- Saier, M.H.; Paulsen, I.T.; Sliwinski, M.K.; Pao, S.S.; Skurray, R.A.; Nikaido, H. Evolutionary origins of multidrug and drug-specific efflux pumps in bacteria. FASEB J. 1998, 12, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Multidrug Resistance in Bacteria. Annu. Rev. Biochem. 2009, 78, 119–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.L.; Sánchez, M.B.; Martínez-Solano, L.; Hernandez, A.; Garmendia, L.; Fajardo, A.; Alvarez-Ortega, C. Functional role of bacterial multidrug efflux pumps in microbial natural ecosystems. FEMS Microbiol. Rev. 2009, 33, 430–449. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L.J. Clinically Relevant chromosomally encoded nultidrug resistance efflux pumps in bacteria. Clin. Microbiol. Rev. 2006, 19, 382–402. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H.; Pagès, J.M. Broad-specificity efflux pumps and their role in multidrug resistance of Gram-negative bacteria. FEMS Microbiol. Rev. 2012, 36, 340–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.Z.; Plésiat, P.; Nikaido, H. The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef] [PubMed]
- Du, D.; Wang-Kan, X.; Neuberger, A.; van Veen, H.W.; Pos, K.M.; Piddock, L.J.V.; Luisi, B.F. Multidrug efflux pumps: Structure, function and regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Neyfakh, A.A. Mystery of multidrug transporters: The answer can be simple. Mol. Microbiol. 2002, 44, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, I.T. Multidrug efflux pumps and resistance: Regulation and evolution. Curr. Opin. Microbiol. 2003, 6, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L.J. Multidrug—Resistance efflux pumps—Not just for resistance. Nat. Rev. Microbiol. 2006, 4, 629–636. [Google Scholar] [CrossRef]
- Alvarez-Ortega, C.; Olivares, J.; Martínez, J.L. RND multidrug efflux pumps: What are they good for? Front. Microbiol. 2013, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Blanco, P.; Hernando-Amado, S.; Reales-Calderon, J.A.; Corona, F.; Lira, F.; Alcalde-Rico, M.; Bernardini, A.; Sanchez, M.B.; Martinez, J.L. Bacterial multidrug efflux pumps: Much more than antibiotic resistance determinants. Microorganisms 2016, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Deng, Z.; Yan, A. Bacterial multidrug efflux pumps: Mechanisms, physiology and pharmacological exploitations. Biochem. Biophys. Res. Commun. 2014, 453, 254–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leuzzi, A.; Di Martino, M.L.; Campilongo, R.; Falconi, M.; Barbagallo, M.; Marcocci, L.; Pietrangeli, P.; Casalino, M.; Grossi, M.; Micheli, G.; et al. Multifactor regulation of the MdtJI polyamine transporter in Shigella. PLoS ONE 2015, 10, e0136744. [Google Scholar] [CrossRef] [PubMed]
- Alcalde-Rico, M.; Hernando-Amado, S.; Blanco, P.; Martínez, J.L. Multidrug Efflux Pumps at the Crossroad between Antibiotic Resistance and Bacterial Virulence. Front. Microbiol. 2016, 7, 1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Q.; Paulsen, I.T. Comparative analyses of fundamental differences in membrane transport capabilities in prokaryotes and eukaryotes. PLoS Comput. Biol. 2005, 1, e27. [Google Scholar] [CrossRef]
- Konstantinidis, K.T.; Tiedje, J.M. Trends between gene content and genome size in prokaryotic species with larger genomes. Proc. Natl. Acad. Sci. USA 2004, 101, 3160–3165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCutcheon, J.P.; Moran, N.A. Extreme genome reduction in symbiotic bacteria. Nat. Rev. Microbiol. 2011, 10, 13–26. [Google Scholar] [CrossRef]
- Prosseda, G.; Di Martino, M.L.; Campilongo, R.; Fioravanti, R.; Micheli, G.; Casalino, M.; Colonna, B. Shedding of genes that interfere with the pathogenic lifestyle: The Shigella model. Res. Microbiol. 2012, 163, 399–406. [Google Scholar] [CrossRef]
- Hassan, K.A.; Liu, Q.; Elbourne, L.D.H.; Ahmad, I.; Sharples, D.; Naidu, V.; Chan, C.L.; Li, L.; Harborne, S.P.D.; Pokhrel, A.; et al. Pacing across the membrane: The novel PACE family of efflux pumps is widespread in Gram-negative pathogens. Res. Microbiol. 2018, 169, 450–454. [Google Scholar] [CrossRef]
- Delmar, J.A.; Yu, E.W. The AbgT family: A novel class of antimetabolite transporters. Protein Sci. 2016, 25, 322–337. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.M.; Richmond, G.E.; Piddock, L.J. Multidrug efflux pumps in Gram-negative bacteria and their role in antibiotic resistance. Future Microbiol. 2014, 9, 1165–1177. [Google Scholar] [CrossRef] [PubMed]
- Hinchliffe, P.; Greene, N.P.; Paterson, N.G.; Crow, A.; Hughes, C.; Koronakis, V. Structure of the periplasmic adaptor protein from a major facilitator superfamily (MFS) multidrug efflux pump. FEBS Lett. 2014, 588, 3147–3153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, D.; van Veen, H.W.; Murakami, S.; Pos, K.M.; Luisi, B.F. Structure, mechanism and cooperation of bacterial multidrug transporters. Curr. Opin. Struct. Biol. 2015, 33, 76–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symmons, M.F.; Marshall, R.L.; Bavro, V.N. Architecture and roles of periplasmic adaptor proteins in tripartite efflux assemblies. Front. Microbiol. 2015, 6, 513. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Lee, J.H. Antibiofilm agents: A new perspective for antimicrobial strategy. J. Microbiol. 2017, 55, 753–766. [Google Scholar] [CrossRef]
- Quistgaard, E.M.; Löw, C.; Guettou, F.; Nordlund, P. Understanding transport by the major facilitator superfamily (MFS): Structures pave the way. Nat. Rev. Mol. Cell Biol. 2016, 17, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Neuberger, A.; Du, D.; Luisi, B.F. Structure and mechanism of bacterial tripartite efflux pumps. Res. Microbiol. 2018, 169, 401–413. [Google Scholar] [CrossRef]
- Hinchliffe, P.; Symmons, M.F.; Hughes, C.; Koronakis, V. Structure and operation of bacterial tripartite pumps. Annu. Rev. Microbiol. 2013, 67, 221–242. [Google Scholar] [CrossRef]
- Pao, S.S.; Paulsen, I.T.; Saier, M.H., Jr. Major facilitator superfamily. Microbiol. Mol. Biol. Rev. 1998, 62, 1–34. [Google Scholar]
- Fluman, N.; Bibi, E. Bacterial multidrug transport through the lens of the major facilitator superfamily. Biochim. Biophys. Acta 2009, 1794, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.S.; Shlykov, M.A.; Castillo, R.; Sun, E.I.; Saier, M.H., Jr. The major facilitator superfamily (MFS) revisited. FEBS J. 2012, 279, 2022–2035. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; He, X.; Szewczyk, P.; Nguyen, T.; Chang, G. Structure of the multidrug transporter EmrD from Escherichia coli. Science 2006, 312, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Law, C.J.; Maloney, P.C.; Wang, D.N. Ins and outs of major facilitator superfamily antiporters. Annu. Rev. Microbiol. 2008, 62, 289–305. [Google Scholar] [CrossRef] [PubMed]
- Koronakis, V.; Eswaran, J.; Hughes, C. Structure and function of TolC: The bacterial exit duct for proteins and drugs. Annu. Rev. Biochem. 2004, 73, 467–489. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.W.; Hu, R.M.; Chu, F.Y.; Lin, H.R.; Yang, T.C. Characterization of a major facilitator superfamily (MFS) tripartite efflux pump EmrCABsm from Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 2013, 68, 2498–2505. [Google Scholar] [CrossRef] [PubMed]
- Barroso, K.C.M.; Previato-Mello, M.; Batista, B.B.; Batista, J.H.; da Silva Neto, J.F. EmrR-Dependent Upregulation of the Efflux Pump EmrCAB Contributes to Antibiotic Resistance in Chromobacterium violaceum. Front. Microbiol. 2018, 9, 2756. [Google Scholar] [CrossRef]
- Grkovic, S.; Brown, M.H.; Skurray, R.A. Regulation of bacterial drug export systems. Microbiol. Mol. Biol. Rev. 2002, 66, 671–701. [Google Scholar] [CrossRef]
- Xu, Z.; Yan, A. Multidrug efflux systems in microaerobic and anaerobic bacteria. Antibiotics 2015, 4, 379–396. [Google Scholar] [CrossRef] [PubMed]
- Lomovskaya, O.; Lewis, K.; Matin, A. EmrR is a negative regulator of the Escherichia coli multidrug resistance pump EmrAB. J. Bacteriol. 1995, 177, 2328–2334. [Google Scholar] [CrossRef]
- Xiong, A.; Gottman, A.; Park, C.; Baetens, M.; Pandza, S.; Matin, A. The EmrR protein represses the Escherichia coli emrRAB multidrug resistance operon by directly binding to its promoter region. Antimicrob. Agents Chemother. 2000, 44, 2905–2907. [Google Scholar] [CrossRef]
- Woolley, R.C.; Vediyappan, G.; Anderson, M.; Lackey, M.; Ramasubramanian, B.; Jiangping, B.; Borisova, T.; Colmer, J.A.; Hamood, A.N.; McVay, C.S.; et al. Characterization of the Vibrio cholerae vceCAB multiple-drug resistance efflux operon in Escherichia coli. J. Bacteriol. 2005, 187, 5500–5503. [Google Scholar] [CrossRef] [PubMed]
- Federici, L.; Du, D.; Walas, F.; Matsumura, H.; Fernandez-Recio, J.; McKeegan, K.S.; Borges-Walmsley, M.I.; Luisi, B.F.; Walmsley, A.R. The crystal structure of the outer membrane protein VceC from the bacterial pathogen Vibrio cholerae at 1.8 Å Resolution. J. Biol. Chem. 2005, 280, 15307–15314. [Google Scholar] [CrossRef] [PubMed]
- Borges-Walmsley, M.I.; Du, D.; McKeegan, K.S.; Sharples, G.J.; Walmsley, A.R. VceR regulates the vceCAB drug efflux pump operon of Vibrio cholerae by alternating between mutually exclusive conformations that bind either drugs or promoter DNA. J. Mol. Biol. 2005, 349, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Shafer, W.M. The farAB-encoded efflux pump mediates resistance of gonococci to long-chained antibacterial fatty acids. Mol. Microbiol. 1999, 33, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Rouquette-Loughlin, C.; Folster, J.P.; Shafer, W.M. FarR regulates the farAB-encoded efflux pump of Neisseria gonorrhoeae via an MtrR regulatory mechanism. J. Bacteriol. 2003, 185, 7145–7152. [Google Scholar] [CrossRef]
- Lee, E.H.; Hill, S.A.; Napier, R.; Shafer, W.M. Integration Host Factor is required for FarR repression of the farAB-encoded efflux pump of Neisseria gonorrhoeae. Mol. Microbiol. 2006, 60, 1381–1400. [Google Scholar] [CrossRef] [PubMed]
- Prosseda, G.; Falconi, M.; Giangrossi, M.; Gualerzi, C.O.; Micheli, G.; Colonna, B. The virF promoter in Shigella: More than just a curved DNA stretch. Mol. Microbiol. 2004, 51, 523–537. [Google Scholar] [CrossRef]
- Prosseda, G.; Mazzola, A.; Di Martino, M.L.; Tielker, D.; Micheli, G.; Colonna, B. A temperature-induced narrow DNA curvature range sustains the maximum activity of a bacterial promoter in vitro. Biochemistry 2010, 49, 2778–2785. [Google Scholar] [CrossRef]
- Quillin, S.J.; Schwartz, K.T.; Leber, J.H. The novel Listeria monocytogenes bile sensor BrtA controls expression of the cholic acid efflux pump MdrT. Mol. Microbiol. 2011, 81, 129–142. [Google Scholar] [CrossRef]
- Crimmins, G.T.; Herskovits, A.A.; Rehder, K.; Sivick, K.E.; Lauer, P.; Dubensky, T.W., Jr.; Portnoy, D.A. Listeria monocytogenes multidrug resistance transporters activate a cytosolic surveillance pathway of innate immunity. Proc. Natl. Acad. Sci. USA 2008, 105, 10191–10196. [Google Scholar] [CrossRef] [PubMed]
- Tennent, J.M.; Lyon, B.R.; Gillespie, M.T.; May, J.W.; Skurray, R.A. Cloning and expression of Staphylococcus aureus plasmid-mediated quaternary ammonium resistance in Escherichia coli. Antimicrob. Agents Chemother. 1985, 27, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.C.; Villet, R.A.; Estabrooks, Z.A.; Hooper, D.C. Native efflux pumps contribute resistance to antimicrobials of skin and the ability of Staphylococcus aureus to colonize skin. J. Infect. Dis. 2014, 209, 1485–1493. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.C.; Bolduc, G.R.; Medeiros, H.; Vyas, J.M.; Wang, Y.; Hooper, D.C. Role of the Tet38 Efflux Pump in Staphylococcus aureus Internalization and Survival in Epithelial Cells. Infect. Immun. 2015, 83, 4362–4372. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.C.; Hooper, D.C. Phosphorylation of MgrA and its effect on expression of the NorA and NorB efflux pumps of Staphylococcus aureus. J. Bacteriol. 2010, 192, 2525–2534. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Onodera, Y.; Lee, J.C.; Hooper, D.C. NorB, an efflux pump in Staphylococcus aureus strain MW2, contributes to bacterial fitness in abscesses. J. Bacteriol. 2008, 190, 7123–7129. [Google Scholar] [CrossRef]
- He, X.; Ahn, J. Differential gene expression in planktonic and biofilm cells of multiple antibiotic-resistant Salmonella Typhimurium and Staphylococcus aureus. FEMS Microbiol. Lett. 2011, 325, 180–188. [Google Scholar] [CrossRef]
- Ding, Y.; Fu, Y.; Lee, J.C.; Hooper, D.C. Staphylococcus aureus NorD, a putative efflux pump coregulated with the Opp1 oligopeptide permease, contributes selectively to fitness in vivo. J. Bacteriol. 2012, 194, 6586–6593. [Google Scholar] [CrossRef]
- Ahmed, M.; Lyass, L.; Markham, P.N.; Taylor, S.S.; Vázquez-Laslop, N.; Neyfakh, A.A.; Va, N. Two highly similar multidrug transporters of Bacillus subtilis whose expression is differentially regulated. J. Bacteriology 1995, 177, 3904–3910. [Google Scholar] [CrossRef]
- Woolridge, D.P.; Vazquez-Laslop, N.; Markham, P.N.; Chevalier, M.S.; Gerner, E.W.; Neyfakh, A.A. Efflux of the natural polyamine spermidine facilitated by the Bacillus subtilis multidrug transporter Blt. J. Biol. Chem. 1997, 272, 8864–8866. [Google Scholar] [CrossRef]
- Di Martino, M.L.; Campilongo, R.; Casalino, M.; Micheli, G.; Colonna, B.; Prosseda, G. Polyamines: Emerging players in bacteria-host interactions. Int. J. Med. Microbiol. 2013, 303, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Campilongo, R.; Di Martino, M.L.; Marcocci, L.; Pietrangeli, P.; Leuzzi, A.; Grossi, M.; Casalino, M.; Nicoletti, M.; Micheli, G.; Colonna, B.; et al. Molecular and functional profiling of the polyamine content in enteroinvasive E. coli: Looking into the gap between commensal E. coli and harmful Shigella. PLoS ONE 2014, 9, e106589. [Google Scholar] [CrossRef] [PubMed]
- Tett, A.J.; Karunakaran, R.; Poole, P.S. Characterisation of SalRAB a salicylic acid inducible positively regulated efflux system of Rhizobium leguminosarum bv viciae 3841. PLoS ONE 2014, 9, e103647. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, H.; Nishino, K.; Hirata, T.; Yamaguchi, A. Comprehensive studies of drug resistance mediated by overexpression of response regulators of two-component signal transduction systems in Escherichia coli. J. Bacteriol. 2003, 185, 1851–1856. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, Y.; Utsumi, R. Alkali metals in addition to acidic pH activate the EvgS histidine kinase sensor in Escherichia coli. J. Bacteriol. 2014, 196, 3140–3149. [Google Scholar] [CrossRef] [PubMed]
- Pasqua, M.; Grossi, M.; Scinicariello, S.; Aussel, L.; Barras, F.; Colonna, B.; Prosseda, G. The MFS efflux pump EmrKY contributes to the survival of Shigella within macrophages. Sci. Rep. 2019, 9, 2906. [Google Scholar] [CrossRef] [PubMed]
- Njamkepo, E.; Fawal, N.; Tran-Dien, A.; Hawkey, J.; Strockbine, N.; Jenkins, C.; Talukder, K.A.; Bercion, R.; Kuleshov, K.; Kolínská, R.; et al. Global phylogeography and evolutionary history of Shigella dysenteriae type 1. Nat. Microbiol. 2016, 1, 16027. [Google Scholar] [CrossRef]
- Pasqua, M.; Michelacci, V.; Di Martino, M.L.; Tozzoli, R.; Grossi, M.; Colonna, B.; Morabito, S.; Prosseda, G. The Intriguing Evolutionary Journey of Enteroinvasive E. coli (EIEC) toward Pathogenicity. Front. Microbiol. 2017, 8, 2390. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.A. Natural products and plant disease resistance. Nature 2001, 411, 843–847. [Google Scholar] [CrossRef]
- Maggiorani Valecillos, A.; Rodríguez Palenzuela, P.; López-Solanilla, E. The role of several multidrug resistance systems in Erwinia chrysanthemi pathogenesis. Mol. Plant-Microbe Interact. 2006, 19, 607–613. [Google Scholar] [CrossRef]
- Barras, F.; van Gijsegem, F.; Chatterjee, A.K. Extracellular enzymes and pathogenesis of soft-rot Erwinia. Annu. Rev. Phytopathol. 1994, 32, 201–234. [Google Scholar] [CrossRef]
- Ravirala, R.S.; Barabote, R.D.; Wheeler, D.M.; Reverchon, S.; Tatum, O.; Malouf, J.; Liu, H.; Pritchard, L.; Hedley, P.E.; Birch, P.R.; et al. Efflux pump gene expression in Erwinia chrysanthemi is induced by exposure to phenolic acids. Mol. Plant-Microbe Interact. 2007, 20, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.R.; Marques, A.T.; Becker, J.D.; Moreira, L.M. The Sinorhizobium meliloti EmrR regulator is required for efficient colonization of Medicago sativa root nodules. Mol. Plant-Microbe Interact. 2014, 27, 388–399. [Google Scholar] [CrossRef] [PubMed]
- González-Pasayo, R.; Martínez-Romero, E. Multiresistance genes of Rhizobium etli CFN42. Mol. Plant. Microbe. Interact. 2000, 13, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Capela, D.; Carrere, S.; Batut, J. Transcriptome-based identification of the Sinorhizobium meliloti NodD1 regulon. Appl. Environ. Microbiol. 2005, 71, 4910–4913. [Google Scholar] [CrossRef]
- Eda, S.; Mitsui, H.; Minamisawa, K. Involvement of the SmeAB multidrug efflux pump in resistance to plant antimicrobials and contribution to nodulation competitiveness in Sinorhizobium meliloti. Appl. Environ. Microbiol. 2011, 77, 2855–2862. [Google Scholar] [CrossRef]
- Rossbach, S.; Kunze, K.; Albert, S.; Zehner, S.; Göttfert, M. The Sinorhizobium meliloti EmrAB efflux system is regulated by flavonoids through a TetR-like regulator (EmrR). Mol. Plant-Microbe Interact. 2014, 27, 379–387. [Google Scholar] [CrossRef]
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic Acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef]
- Woodward, J.J.; Iavarone, A.T.; Portnoy, D.A. c-di-AMP secreted by intracellular Listeria monocytogenes activates a host type I interferon response. Science 2010, 328, 1703–1705. [Google Scholar] [CrossRef]
- Truong-Bolduc, Q.C.; Khan, N.S.; Vyas, J.M.; Hooper, D.C. Tet38 efflux pump affects Staphylococcus aureus internalization by epithelial cells through interaction with CD36 and contributes to bacterial escape from acidic and nonacidic phagolysosomes. Infect. Immun. 2017, 85, e00862–e00916. [Google Scholar] [CrossRef]
- Bianco, M.V.; Blanco, F.C.; Forrellad, M.A.; Aguilar, D.; Campos, E.; Klepp, L.I.; Hernández-Pando, R.; Cataldi, A.A.; Bigi, F. Knockout mutation of p27-p55 operon severely reduces replication of Mycobacterium bovis in a macrophagic cell line and survival in a mouse model of infection. Virulence 2011, 2, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Bigi, F.; Gioffré, A.; Klepp, L.; Santangelo, M.P.; Alito, A.; Caimi, K.; Meikle, V.; Zumárraga, M.; Taboga, O.; Romano, M.I.; et al. The knockout of the lprG-Rv1410 operon produces strong attenuation of Mycobacterium tuberculosis. Microbes Infect. 2004, 6, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.N.; Takaki, K.; Connolly, L.E.; Wiedenhoft, H.; Winglee, K.; Humbert, O.; Edelstein, P.H.; Cosma, C.L.; Ramakrishnan, L. Drug tolerance in replicating mycobacteria mediated by a macrophage-induced efflux mechanism. Cell 2011, 145, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.P.; Nguyen, L.; Gatfield, J.; Visconti, K.; Nguyen, K.; Schnappinger, D.; Ehrt, S.; Liu, Y.; Heifets, L.; Pieters, J.; et al. Ancestral antibiotic resistance in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2005, 102, 12200–12205. [Google Scholar] [CrossRef] [PubMed]
- Schnappinger, D.; Ehrt, S.; Voskuil, M.I.; Liu, Y.; Mangan, J.A.; Monahan, I.M.; Dolganov, G.; Efron, B.; Butcher, P.D.; Nathan, C.; et al. Transcriptional adaptation of Mycobacterium tuberculosis within macrophages: Insights into the phagosomal environment. J. Exp. Med. 2003, 198, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sharma, R.; Bhattacharyya, T.; Bhando, T.; Pathania, R. Fosfomycin resistance in Acinetobacter baumannii is mediated by efflux through a major facilitator superfamily (MFS) transporter-AbaF. J. Antimicrob. Chemother. 2017, 72, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Varela, M.; Corral, J.; Vallejo, J.A.; Rumbo-Feal, S.; Bou, G.; Aranda, J.; Barbé, J. Mutations in the β-Subunit of the RNA Polymerase Impair the Surface-Associated Motility and Virulence of Acinetobacter baumannii. Infect. Immun. 2017, 85, e00327–e00417. [Google Scholar] [CrossRef]
- Sahu, P.K.; Iyer, P.S.; Gaikwad, M.B.; Talreja, S.C.; Pardesi, K.R.; Chopade, B.A. An MFS transporter-like ORF from MDR Acinetobacter baumannii AIIMS 7 is associated with adherence and biofilm formation on biotic/abiotic surface. Int. J. Microbiol. 2012, 2012, 490647. [Google Scholar] [CrossRef]
- Liu, J.Y.; Miller, P.F.; Willard, J.; Olson, E.R. Functional and biochemical characterization of Escherichia coli sugar efflux transporters. J. Biol. Chem. 1999, 274, 22977–22984. [Google Scholar] [CrossRef]
- May, T.; Ito, A.; Okabe, S. Induction of multidrug resistance mechanism in Escherichia coli biofilms by interplay between tetracycline and ampicillin resistance genes. Antimicrob. Agents Chemother. 2009, 53, 4628–4639. [Google Scholar] [CrossRef]
- Koita, K.; Rao, C.V. Identification and analysis of the putative pentose sugar efflux transporters in Escherichia coli. PLoS ONE 2012, 7, e43700. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Cai, Y.; Chen, Z.; Gao, S.; Geng, X.; Li, Y.; Li, Y.; Jia, J.; Sun, Y. Bifunctional enzyme SpoT is involved in biofilm formation of Helicobacter pylori with multidrug resistance by upregulating efflux pump Hp1174 (gluP). Antimicrob. Agents Chemother. 2018, 62, e00957–e01018. [Google Scholar] [CrossRef] [PubMed]
- Resch, A.; Rosenstein, R.; Nerz, C.; Götz, F. Differential gene expression profiling of Staphylococcus aureus cultivated under biofilm and planktonic conditions. Appl. Environ. Microbiol. 2005, 71, 2663–2676. [Google Scholar] [CrossRef] [PubMed]
- Hagman, K.E.; Pan, W.; Spratt, B.G.; Balthazar, J.T.; Judd, R.C.; Shafer, W.M. Resistance of Neisseria gonorrhoeae to antimicrobial hydrophobic agents is modulated by the mtrRCDE efflux system. Microbiology 1995, 141, 611–622. [Google Scholar] [CrossRef]
- Lucas, C.E.; Balthazar, J.T.; Hagman, K.E.; Shafer, W.M. The MtrR repressor binds the DNA sequence between the mtrR and mtrC genes of Neisseria gonorrhoeae. J. Bacteriol. 1997, 179, 4123–4128. [Google Scholar] [CrossRef] [PubMed]
- Guymon, L.F.; Walstad, D.L.; Sparling, P.F. Cell envelope alterations in antibiotic-sensitive and-resistant strains of Neisseria gonorrhoeae. J. Bacteriol. 1978, 136, 391–401. [Google Scholar] [PubMed]
- McFarland, L.; Mietzner, T.A.; Knapp, J.S.; Sandstrom, E.; Holmes, K.K.; Morse, S.A. Gonococcal sensitivity to fecal lipids can be mediated by an Mtr-independent mechanism. J. Clin. Microbiol. 1983, 18, 121–127. [Google Scholar] [Green Version]
- Urdaneta, V.; Casadesús, J. Adaptation of Salmonella enterica to bile: Essential role of AcrAB-mediated efflux. Environ. Microbiol. 2018, 20, 1405–1418. [Google Scholar] [CrossRef]
- Gahan, C.G.; Hill, C. Listeria monocytogenes: Survival and adaptation in the gastrointestinal tract. Front. Cell. Infect. Microbiol. 2014, 4, 9. [Google Scholar] [CrossRef]
- Kaplan Zeevi, M.; Shafir, N.S.; Shaham, S.; Friedman, S.; Sigal, N.; Nir Paz, R.; Boneca, I.G.; Herskovits, A.A. Listeria monocytogenes multidrug resistance transporters and cyclic di-AMP, which contribute to type I interferon induction, play a role in cell wall stress. J. Bacteriol. 2013, 195, 5250–5261. [Google Scholar] [CrossRef]
- McFarland, A.P.; Luo, S.; Ahmed-Qadri, F.; Zuck, M.; Thayer, E.F.; Goo, Y.A.; Hybiske, K.; Tong, L.; Woodward, J.J. Sensing of Bacterial Cyclic Dinucleotides by the Oxidoreductase RECON Promotes NF-κB Activation and Shapes a Proinflammatory Antibacterial State. Immunity 2017, 46, 433–445. [Google Scholar] [CrossRef] [PubMed]
- McFarland, A.P.; Burke, T.P.; Carletti, A.A.; Glover, R.C.; Tabakh, H.; Welch, M.D.; Woodward, J.J. RECON-dependent inflammation in hepatocytes enhances Listeria monocytogenes cell-to-cell spread. MBio 2018, 9, e00526–e00618. [Google Scholar] [CrossRef]
- Tadmor, K.; Pozniak, Y.; Burg Golani, T.; Lobel, L.; Brenner, M.; Sigal, N.; Herskovits, A.A. Listeria monocytogenes MDR transporters are involved in LTA synthesis and triggering of innate immunity during infection. Front. Cell. Infect. Microbiol. 2014, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Bigi, F.; Alito, A.; Romano, M.I.; Zumarraga, M.; Caimi, K.; Cataldi, A. The gene encoding P27 lipoprotein and a putative antibiotic-resistance gene form an operon in Mycobacterium tuberculosis and Mycobacterium bovis. Microbiology 2000, 146, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Viale, M.N.; Park, K.T.; Imperiale, B.; Gioffre, A.K.; Colombatti Olivieri, M.A.; Moyano, R.D.; Morcillo, N.; Santangelo, M.; de Santangelo, M.L.; Davis, W.; et al. Characterization of a Mycobacterium avium subsp. avium operon associated with virulence and drug detoxification. Biomed Res. Int. 2014, 2014, 809585. [Google Scholar] [CrossRef] [PubMed]
- Martinot, A.J.; Farrow, M.; Bai, L.; Layre, E.; Cheng, T.Y.; Tsai, J.H.; Iqbal, J.; Annand, J.W.; Sullivan, Z.A.; Hussain, M.M.; et al. Mycobacterial metabolic syndrome: LprG and Rv1410 regulate triacylglyceride levels, growth rate and virulence in Mycobacterium tuberculosis. PLoS Pathog. 2016, 12, e1005351. [Google Scholar] [CrossRef] [PubMed]
- Sassetti, C.M.; Rubin, E.J. Genetic requirements for mycobacterial survival during infection. Proc. Natl. Acad. Sci. USA 2003, 100, 12989–12994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rengarajan, J.; Bloom, B.R.; Rubin, E.J. Genome-wide requirements for Mycobacterium tuberculosis adaptation and survival in macrophages. Proc. Natl. Acad. Sci. USA 2005, 102, 8327–8332. [Google Scholar] [CrossRef] [PubMed]
- Ramón-García, S.; Stewart, G.R.; Hui, Z.K.; Mohn, W.W.; Thompson, C.J. The mycobacterial P55 efflux pump is required for optimal growth on cholesterol. Virulence 2015, 6, 444–448. [Google Scholar] [CrossRef] [Green Version]
- De Rossi, E.; Aínsa, J.A.; Riccardi, G. Role of mycobacterial efflux transporters in drug resistance: An unresolved question. FEMS Microbiol. Rev. 2006, 30, 36–52. [Google Scholar] [CrossRef]
- Lin, W.; de Sessions, P.F.; Teoh, G.H.; Mohamed, A.N.; Zhu, Y.O.; Koh, V.H.; Ang, M.L.; Dedon, P.C.; Hibberd, M.L.; Alonso, S. Transcriptional profiling of Mycobacterium tuberculosis exposed to in vitro lysosomal stress. Infect. Immun. 2016, 84, 2505–2523. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, M.L.; Fioravanti, R.; Barbabella, G.; Prosseda, G.; Colonna, B.; Casalino, M. Molecular evolution of the nicotinic acid requirement within the Shigella/EIEC pathotype. Int. J. Med. Microbiol. 2013, 303, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Dorsey-Oresto, A.; Malik, M.; Wang, J.Y.; Drlica, K.; Zhao, X.; Lu, T. Escherichia coli genes that reduce the lethal effects of stress. BMC Microbiol. 2010, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Stauff, D.L.; Bassler, B.L. Quorum sensing in Chromobacterium violaceum: DNA recognition and gene regulation by the CviR receptor. J. Bacteriol. 2011, 193, 3871–3878. [Google Scholar] [CrossRef] [PubMed]
- Roscetto, E.; Rocco, F.; Carlomagno, M.S.; Casalino, M.; Colonna, B.; Zarrilli, R.; Di Nocera, P.P. PCR-based rapid genotyping of Stenotrophomonas maltophilia isolates. BMC Microbiol. 2008, 8, 202. [Google Scholar] [CrossRef] [PubMed]
- De Carolis, E.; Posteraro, B.; Florio, A.R.; Colonna, B.; Prosseda, G.; Bugli, F.; Lorenzetti, S.R.; Fiscarelli, E.; Inzitari, R.; Iavarone, F.; et al. Analysis of heat-induced changes in protein expression of Stenotrophomonas maltophilia K279a reveals a role for GroEL in the host-temperature adaptation. Int. J. Med. Microbiol. 2011, 301, 273–281. [Google Scholar] [CrossRef]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Domenico, E.G.; Cavallo, I.; Pontone, M.; Toma, L.; Ensoli, F. Biofilm producing Salmonella Typhi: Chronic colonization and development of gallbladder cancer. Int. J. Mol. Sci. 2017, 18, 1887. [Google Scholar] [CrossRef]
- Baugh, S.; Ekanayaka, A.S.; Piddock, L.J.; Webber, M.A. Loss of or inhibition of all multidrug resistance efflux pumps of Salmonella enterica serovar Typhimurium results in impaired ability to form a biofilm. J. Antimicrob. Chemother. 2012, 67, 2409–2417. [Google Scholar] [CrossRef]
- Matsumura, K.; Furukawa, S.; Ogihara, H.; Morinaga, Y. Roles of multidrug efflux pumps on the biofilm formation of Escherichia coli K-12. Biocontrol Sci. 2011, 16, 69–72. [Google Scholar] [CrossRef]
- Psakis, G.; Saidijam, M.; Shibayama, K.; Polaczek, J.; Bettaney, K.E.; Baldwin, J.M.; Baldwin, S.A.; Hope, R.; Essen, L.O.; Essenberg, R.C.; et al. The sodium-dependent d-glucose transport protein of Helicobacter pylori. Mol. Microbiol. 2009, 71, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, M.; Ohta, T.; Uchiyama, I.; Baba, T.; Yuzawa, H.; Kobayashi, I.; Cui, L.; Oguchi, A.; Aoki, K.; Nagai, Y.; et al. Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet 2001, 357, 1225–1240. [Google Scholar] [CrossRef]
- Sahu, P.K.; Iyer, P.S.; Oak, A.M.; Pardesi, K.R.; Chopade, B.A. Characterization of eDNA from the clinical strain Acinetobacter baumannii AIIMS 7 and its role in biofilm formation. Sci. World J. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Whitchurch, C.B.; Tolker-Nielsen, T.; Ragas, P.C.; Mattick, J.S. Extracellular DNA required for bacterial biofilm formation. Science 2002, 295, 1487. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Effux Pump * | Microorganism | Main Functions ** | Reference | |
|---|---|---|---|---|
| Interaction with plants | EmrAB | Erwinia chrysanthemi | Virulence in potato, resistance to phenolic acids | [70,72] |
| EmrAB | Sinorhizobium melitoti | Response to plant induced luteolin | [73,77] | |
| RmrAB | Rhizobium etli | nodule formation, resistance to plant flavonoid | [74] | |
| SalRAB | Rhizobium leguminosarum | Resistance to salicylic acid | [63] | |
| Interaction with animal cells | EmrKY | Shigella flexneri | Survival in macrophages | [66] |
| EmrAB | Escherichia coli | Resistance to bile salts | [40] | |
| FarAB | Neisseria gonorrhoeae | Resistance to long-chained fatty acid | [45] | |
| MdrT | Listeria monocytogenes | Export of cholic acid and c-di-AMP | [51,79] | |
| MdrM | L. monocytogenes | Export of c-di-AMP | [79] | |
| Tet38 | Staphylococcus aureus | Internalization and survival in epithelial cells | [53,54,80] | |
| NorB | S. aureus | Survival in abscesses and in biofilms | [56,57] | |
| NorD | S. aureus | Survival in abscesses | [58] | |
| P55 | Mycobacterium tuberculosi Mycobacterium bovis | Survival in macrophages and in mouse model | [81,82] | |
| Tap | M. tuberculosis | Survival in macrophages | [83,84,85] | |
| VceCAB | Vibrio cholerae | Resistance to bile salts | [42] | |
| Bacterial communication and Biofilm formation | EmrCAB | Chromobacterium violaceum | Quorum-sensing signaling | [37] |
| AbaF | Acinetobacter baumannii | Biofilm formation, virulence | [86] | |
| AbaQ | A. baumannii | Surface—associated motility, virulence | [87] | |
| Pmt | A. baumannii | Nucleic acid transporter—biofilm formation | [88] | |
| SetB | E. coli | Biofilm formation | [89] | |
| TetAC | E. coli | Biofilm maturation | [90] | |
| EmrD | E. coli | Biofilm formation | [91] | |
| GluP | Helicobacter pylori | Biofilm formation | [92] | |
| ProP | S. aureus | Protection from osmotic stress | [93] | |
| EmrCABsm | Stenotrophomonas maltophilia | Extrusion of environmental molecules | [36] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pasqua, M.; Grossi, M.; Zennaro, A.; Fanelli, G.; Micheli, G.; Barras, F.; Colonna, B.; Prosseda, G. The Varied Role of Efflux Pumps of the MFS Family in the Interplay of Bacteria with Animal and Plant Cells. Microorganisms 2019, 7, 285. https://doi.org/10.3390/microorganisms7090285
Pasqua M, Grossi M, Zennaro A, Fanelli G, Micheli G, Barras F, Colonna B, Prosseda G. The Varied Role of Efflux Pumps of the MFS Family in the Interplay of Bacteria with Animal and Plant Cells. Microorganisms. 2019; 7(9):285. https://doi.org/10.3390/microorganisms7090285
Chicago/Turabian StylePasqua, Martina, Milena Grossi, Alessandro Zennaro, Giulia Fanelli, Gioacchino Micheli, Frederic Barras, Bianca Colonna, and Gianni Prosseda. 2019. "The Varied Role of Efflux Pumps of the MFS Family in the Interplay of Bacteria with Animal and Plant Cells" Microorganisms 7, no. 9: 285. https://doi.org/10.3390/microorganisms7090285