
Lactobacillus gasseri PA-3 Uses the Purines IMP, Inosine and Hypoxanthine and Reduces their Absorption in Rats

Abstract
:1. Introduction
2. Experimental Section
2.1. Reagents, Radioactive Compounds, Media, and Lactic Acid Bacteria
2.2. Assays of Purine Nucleoside and Nucleotide Degradation
2.3. Measurement of Purines by HPLC
2.4. Uptake of Purines in DM-py
2.5. Proliferation Assay
2.6. Viable Cell Counts
2.7. Animal Experiments
2.8. Statistical Analyses
3. Results
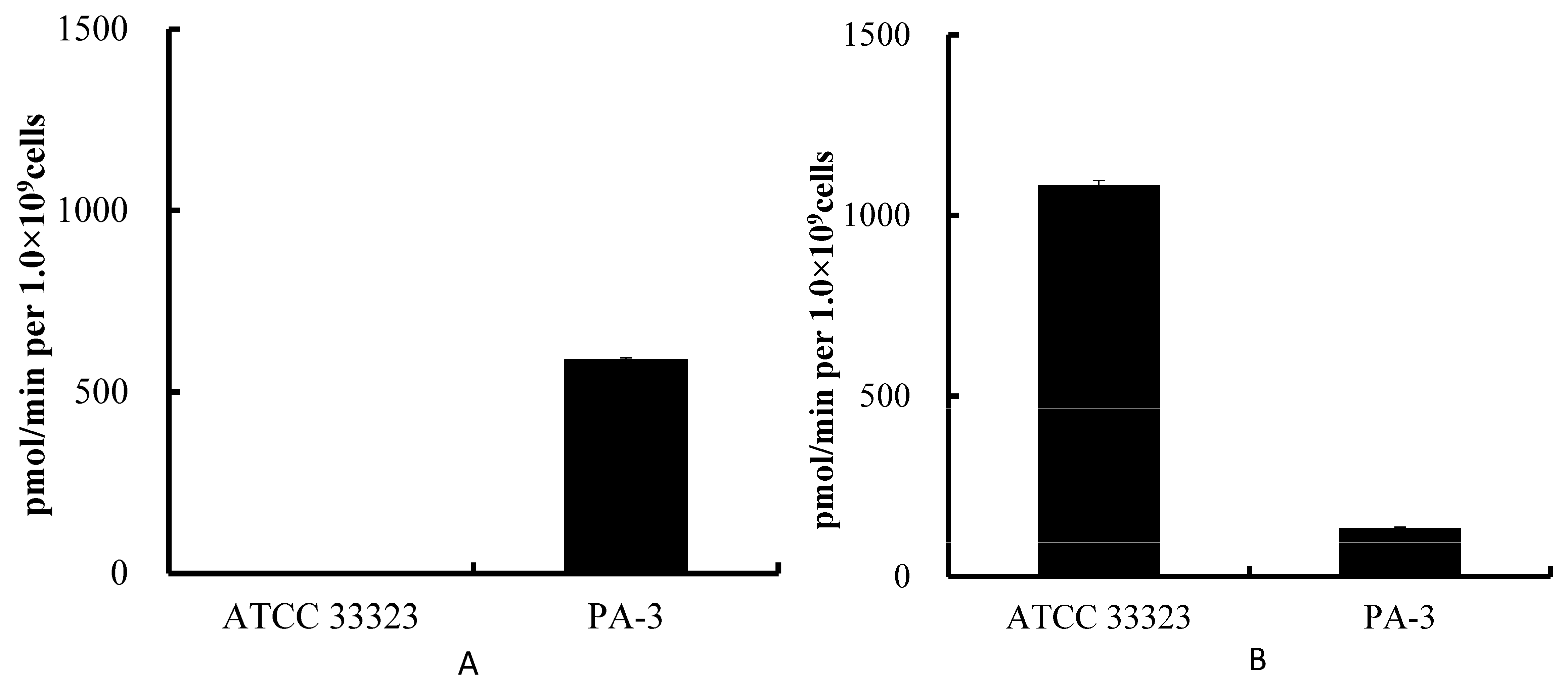
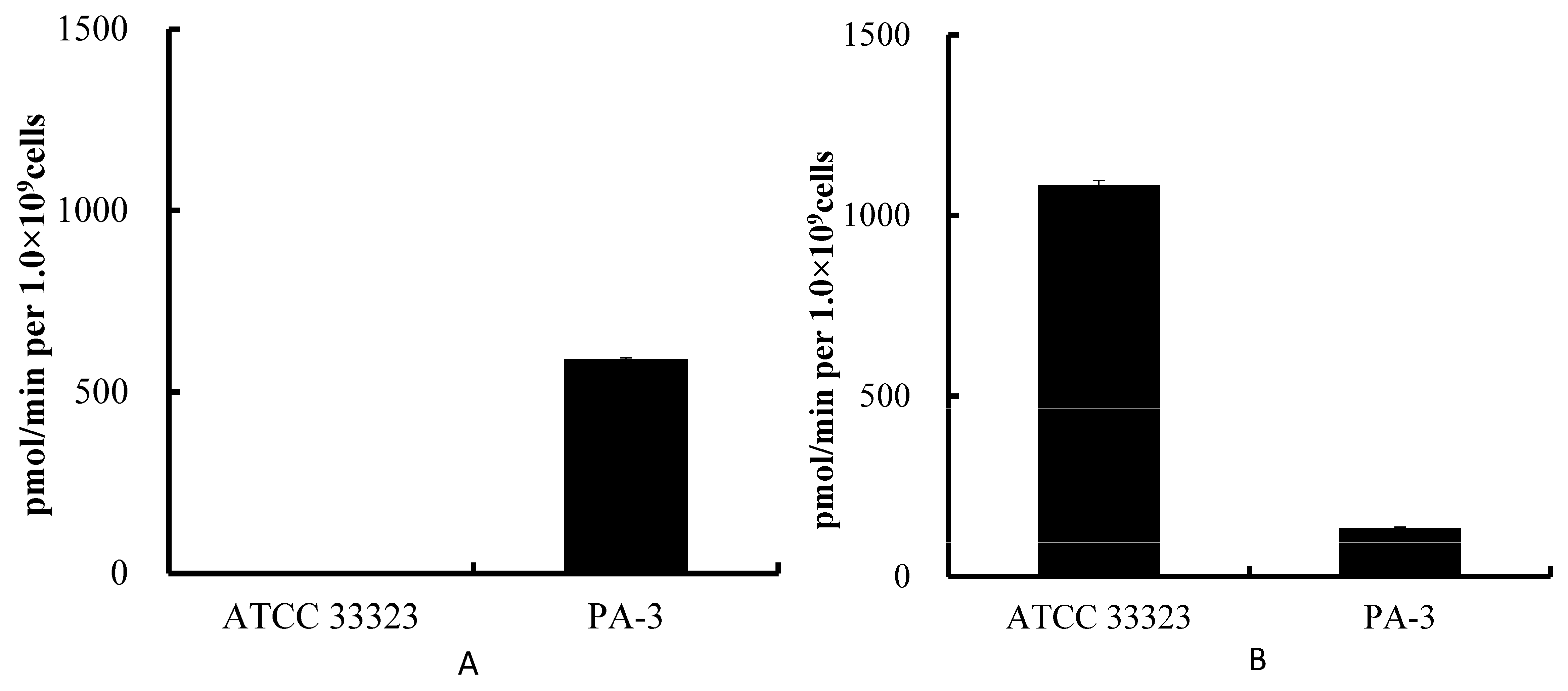
3.1. Degradation of Purines by ATCC 33323 and PA-3
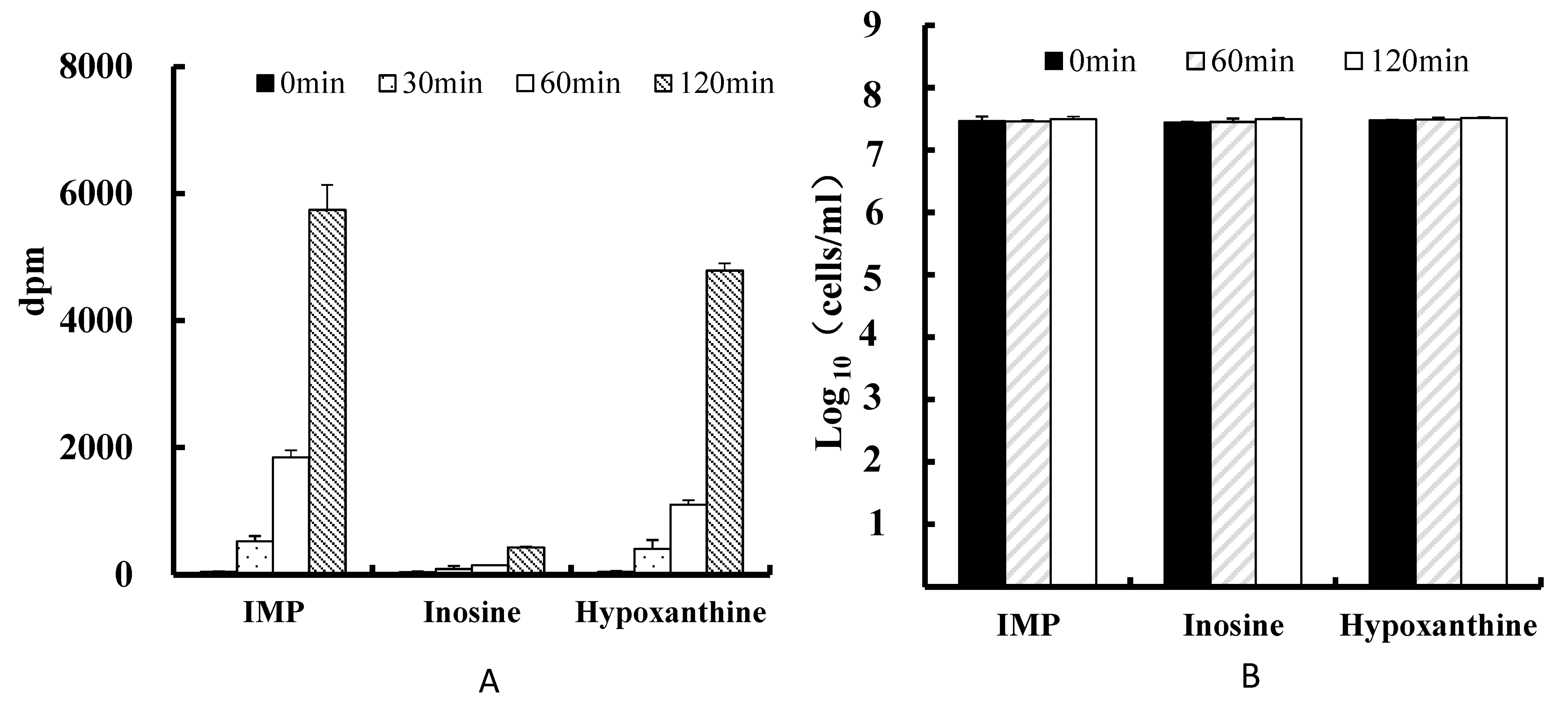
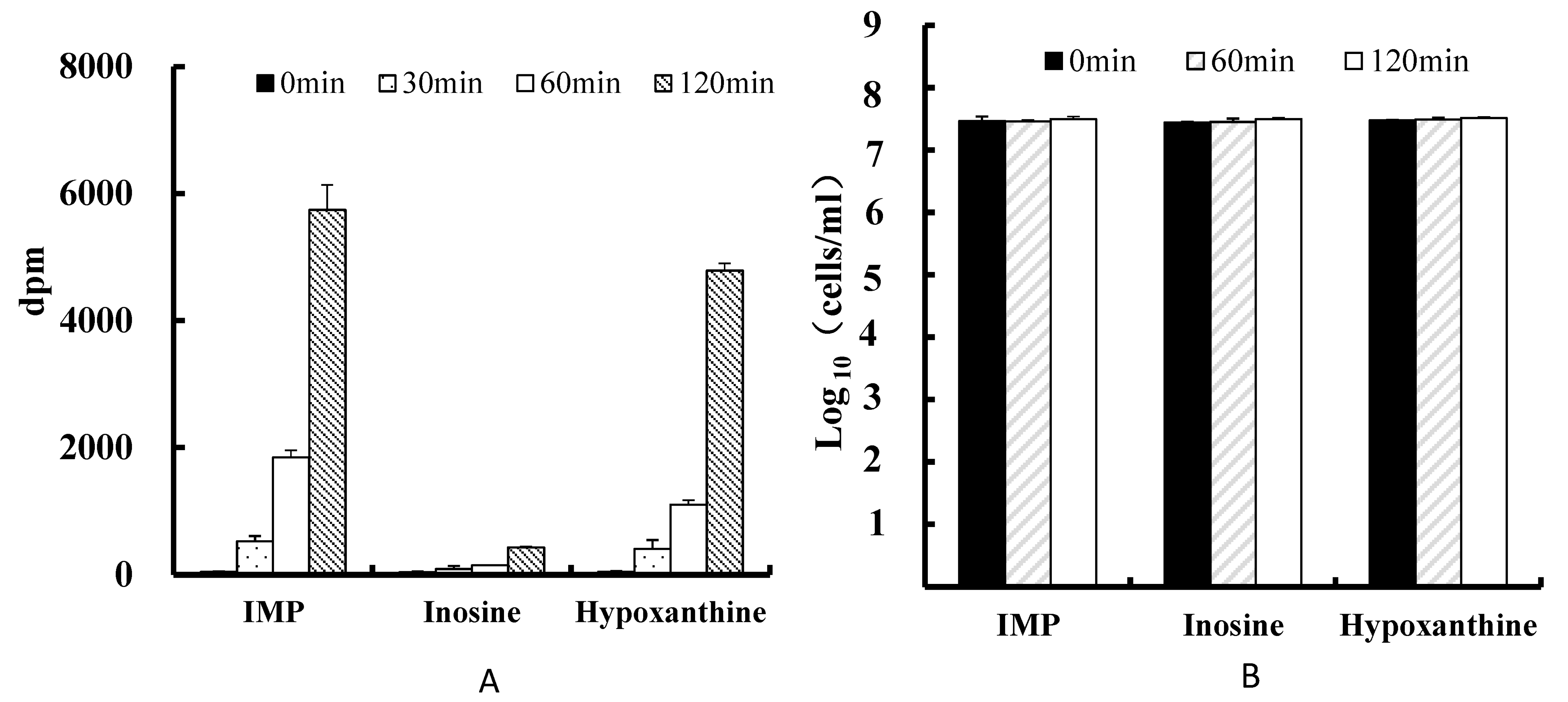
3.2. Uptake of Purines
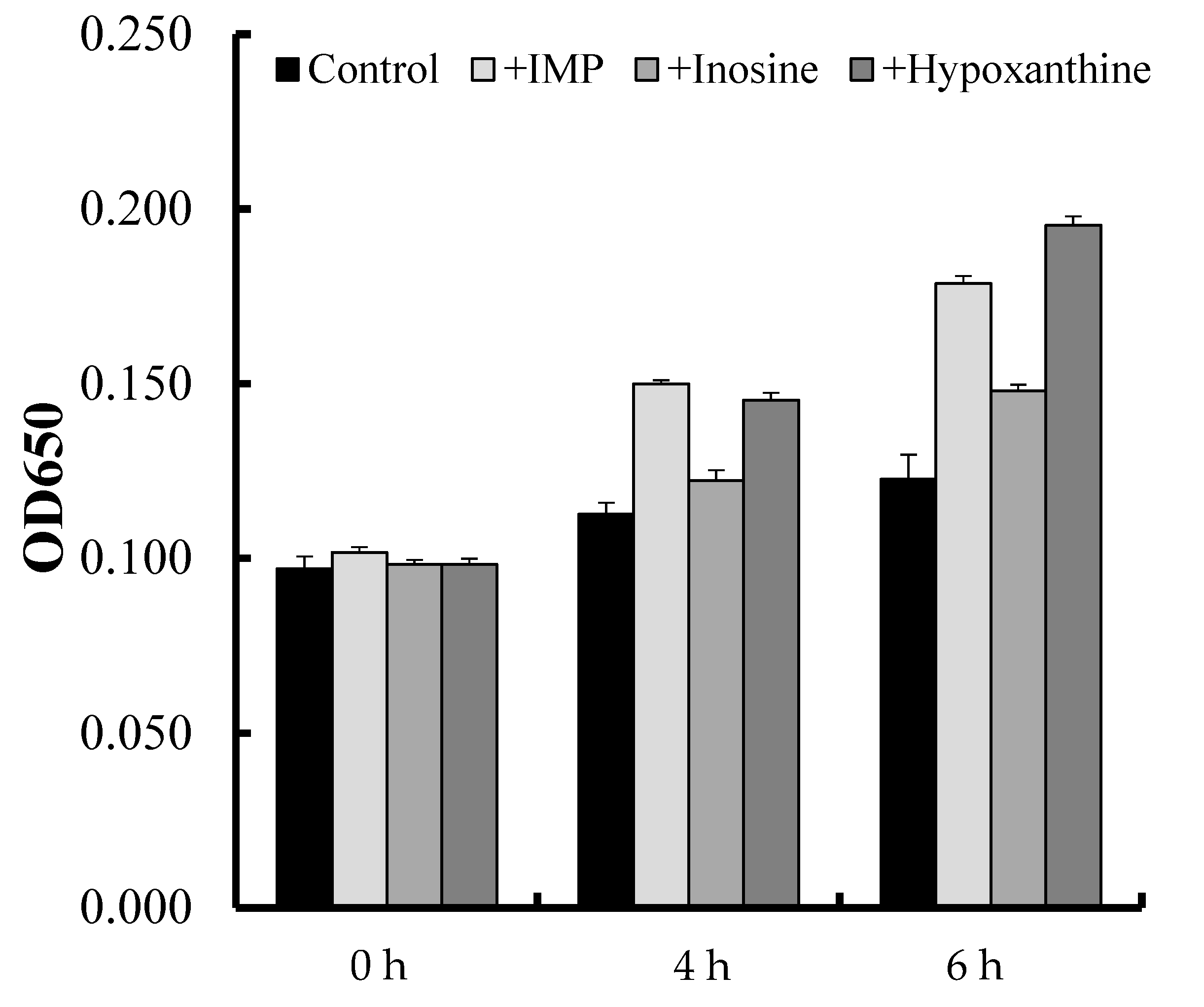
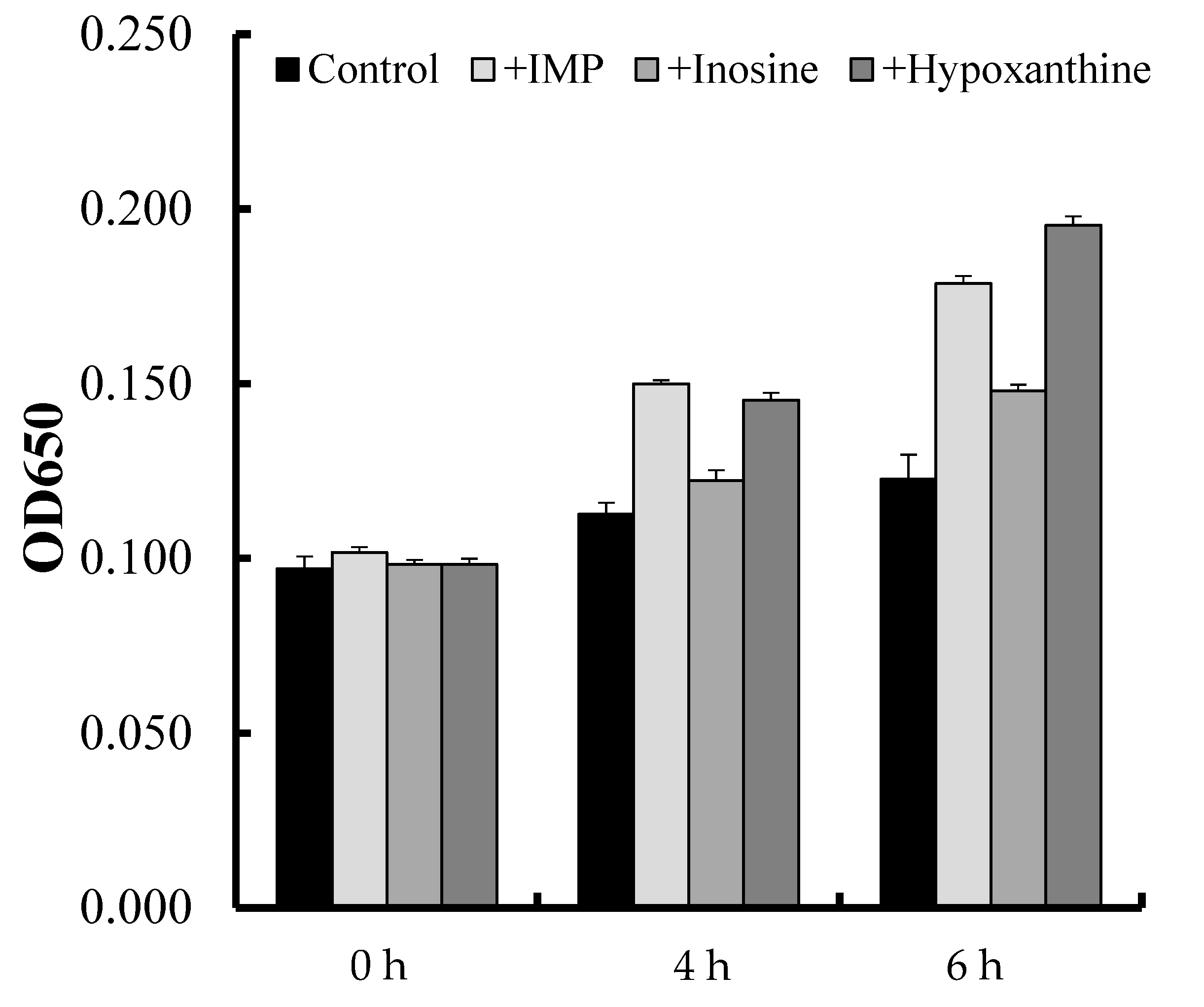
3.3. Proliferation Assay
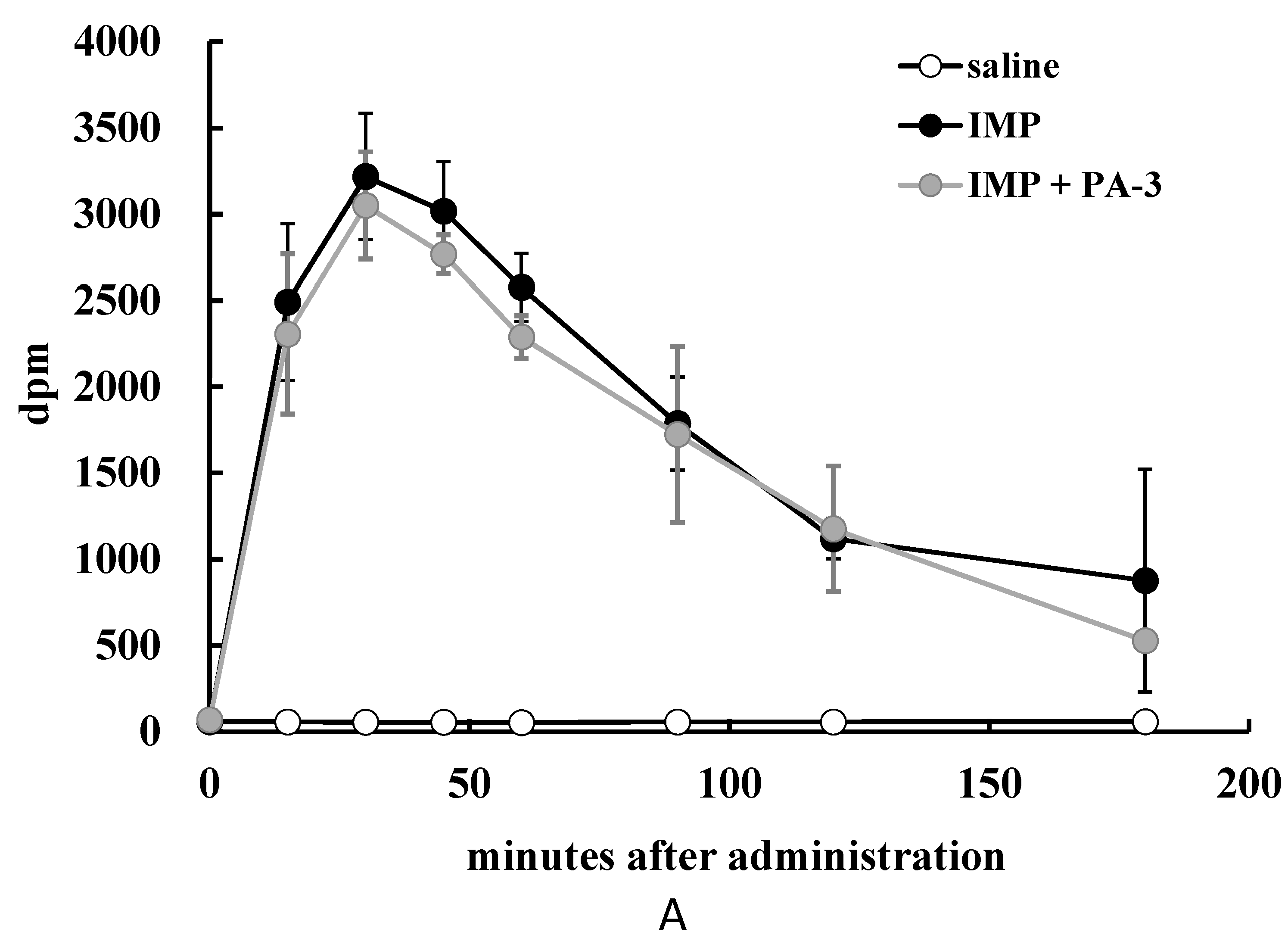
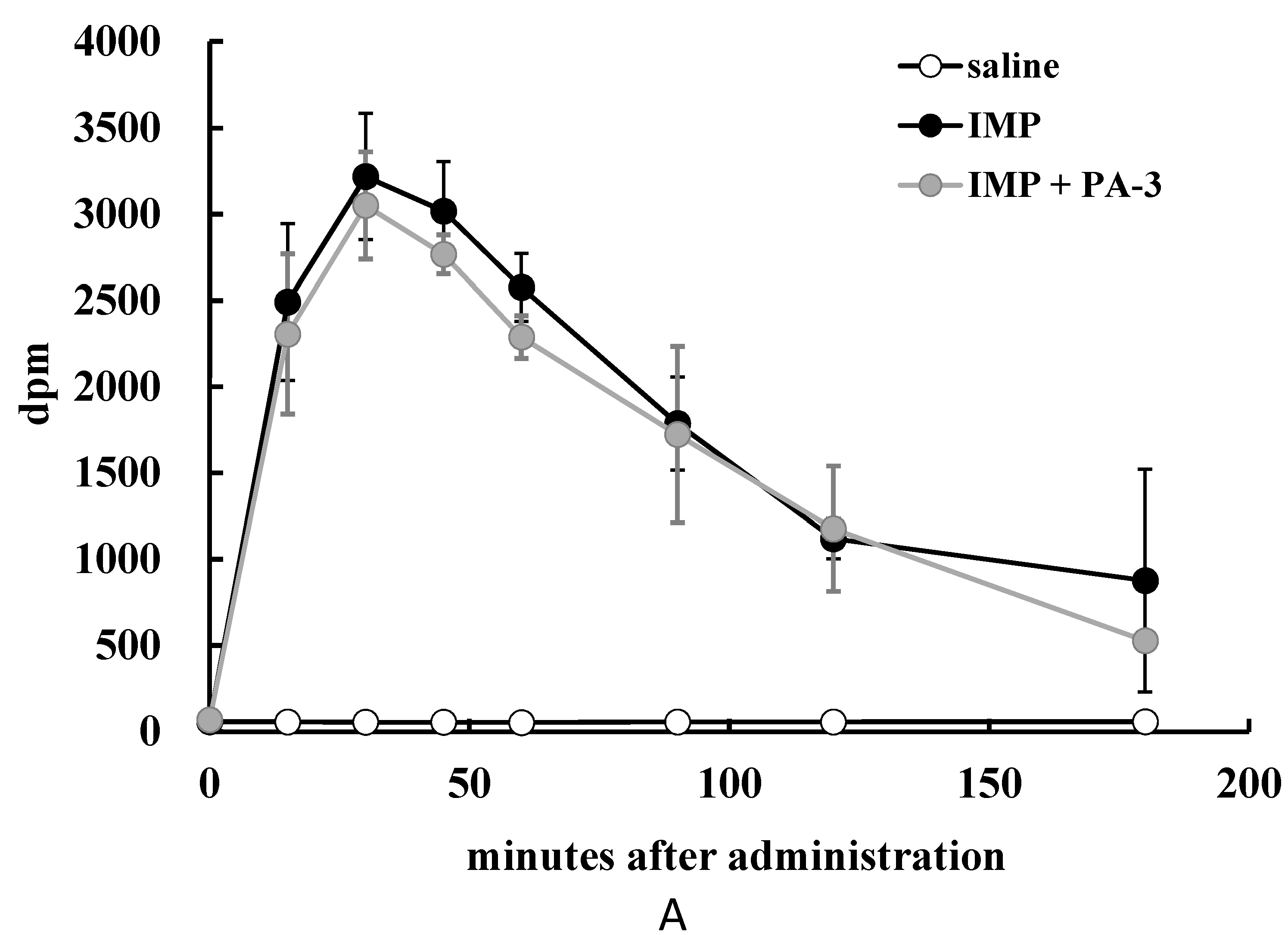
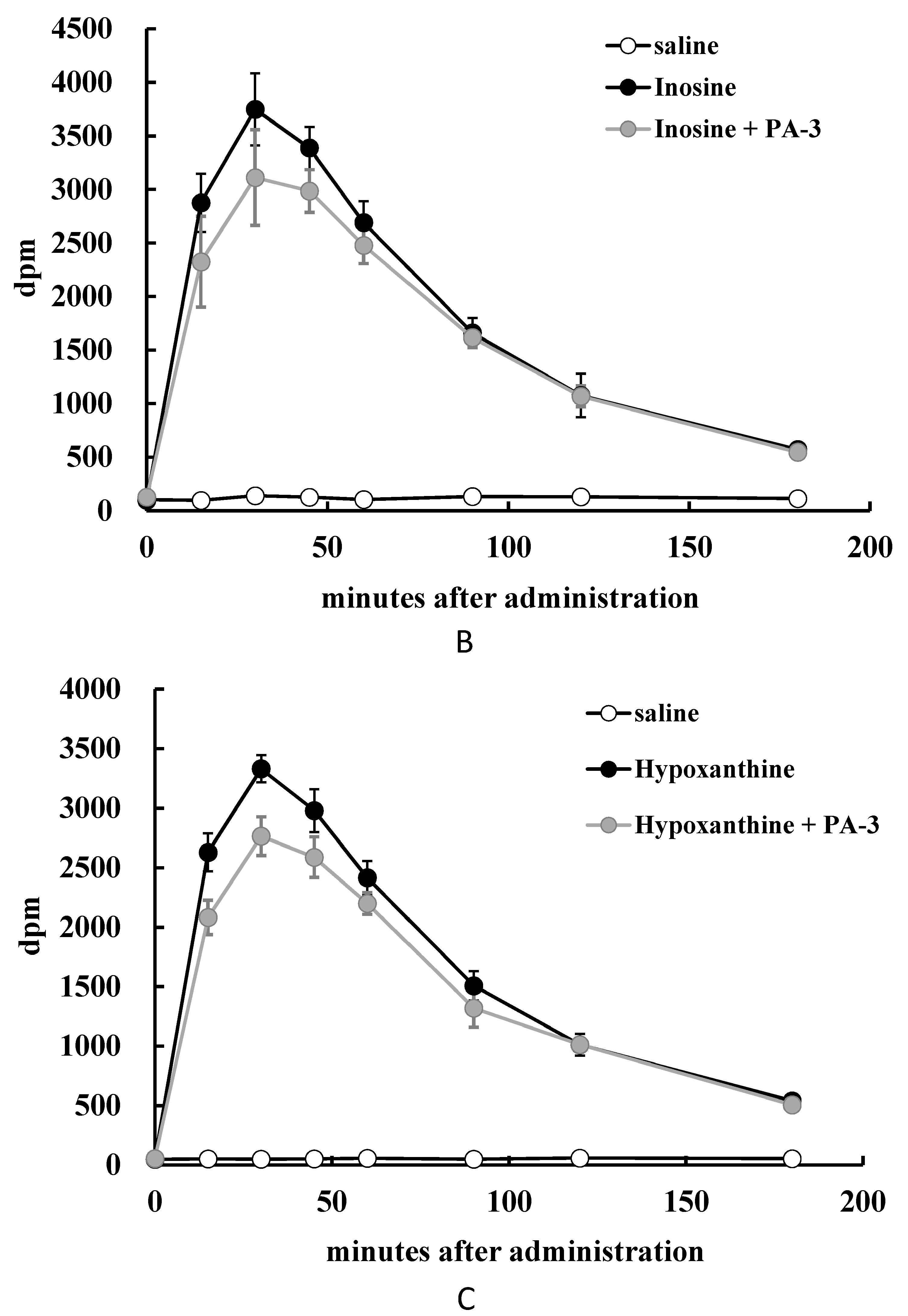
3.4. Animal Experiment
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Benedict, J.D.; Forsham, P.H.; Stetten, D., Jr. The metabolism of uric acid in the normal and gouty human studied with the aid of isotopic uric acid. J. Biol. Chem. 1949, 181, 183–193. [Google Scholar] [PubMed]
- Garrel, D.R.; Verdy, M.; PetitClerc, C.; Martin, C.; Brule, D.; Hamet, P. Milk- and soy-protein ingestion: Acute effect on serum uric acid concentration. Am. J. Clin. Nutr. 1991, 53, 665–669. [Google Scholar] [PubMed]
- Choi, H.K.; Atkinson, K.; Karlson, E.W.; Willett, W.; Curhan, G. Purine-rich foods, dairy and protein intake, and the risk of gout in men. N. Engl. J. Med. 2004, 350, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.K.; Liu, S.; Curhan, G. Intake of purine-rich foods, protein, and dairy products and relationship to serum levels of uric acid: The Third National Health and Nutrition Examination Survey. Arthritis Rheum. 2005, 52, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Villegas, R.; Xiang, Y.B.; Elasy, T.; Xu, W.H.; Cai, H.; Cai, Q.; Linton, M.F.; Fazio, S.; Zheng, W.; Shu, X.O. Purine-rich foods, protein intake, and the prevalence of hyperuricemia: The Shanghai Men’s Health Study. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.A.; Chohan, S. We can make gout management more successful now. Curr. Opin. Rheumatol. 2008, 20, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, H. Japanese guideline for the management of hyperuricemia and gout: Second edition. Nucleosides Nucleotides Nucleic Acids 2011, 30, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, K.; Aoyagi, Y.; Fukuuchi, T.; Inazawa, K.; Yamaoka, N. Total purine and purine base content of common foodstuffs for facilitating nutritional therapy for gout and hyperuricemia. Biol. Pharm. Bull. 2014, 37, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Rundles, R.W.; Metz, E.N.; Silberman, H.R. Alloprinol in the treatment of gout. Ann. Intern. Med. 1966, 64, 229–258. [Google Scholar] [PubMed]
- Kilstrup, M.; Hammer, K.; Ruhdal Jensen, P.; Martinussen, J. Nucleotide metabolism and its control in lactic acid bacteria. FEMS Microbiol. Rev. 2005, 29, 555–590. [Google Scholar] [CrossRef] [PubMed]
- Bolotin, A.; Wincker, P.; Mauger, S.; Jaillon, O.; Malarme, K.; Weissenbach, J.; Ehrlich, S.D.; Sorokin, A. The complete genome sequence of the lactic acid bacterium Lactococcus lactis ssp. lactis IL1403. Genome Res. 2001, 11, 731–753. [Google Scholar] [CrossRef] [PubMed]
- Yamada, N.; Iwamoto, C.; Nakamura, M.; Soeda, M.; Tsuboi, H.; Kano, H.; Asami, Y. Reducing effect of Lactobacillus gasseri PA-3 on the absorption of food-derived purines. Milk Sci. 2016, 65, 25–31. [Google Scholar]
- Yamada, N.; Iwamoto, C.; Kano, H.; Yamaoka, N.; Fukuuchi, T.; Kaneko, K.; Asami, Y. Evaluation of purine utilization by Lactobacillus gasseri strains with potential to decrease the absorption of food-derived purines in the human intestine. Nucleosides Nucleotides Nucleic Acids 2016, 35, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Elli, M.; Zink, R.; Rytz, A.; Reniero, R.; Morelli, L. Iron requirement of Lactobacillus spp. in completely chemically defined growth media. J. Appl. Microbiol. 2000, 88, 695–703. [Google Scholar] [PubMed]
- Lin, Y.; Zhang, W.; Zhu, F.; Su, J.; Fang, D.; Yang, Y.; Zhang, G.; Xie, L.; Zhang, R.; Wang, H. Subcellular localization of N-deoxyribosyltransferase in Lactobacillus fermentum: Cell surface association of an intracellular nucleotide metabolic enzyme. FEMS Microbiol. Lett. 2011, 323, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Martinussen, J.; Sørensen, C.; Jendresen, C.B.; Kilstrup, M. Two nucleoside transporters in Lactococcus lactis with different substrate specificities. Microbiology 2010, 156, 3148–3157. [Google Scholar] [CrossRef] [PubMed]
- Martinussen, J.; Wadskov-Hansen, S.L.; Hammer, K. Two nucleoside uptake system in Lactococcus lactis: Competition between purine nucleosides and cytidine allows for modulation of intracellular nucleotide pools. J. Bacteriol. 2003, 185, 1503–1508. [Google Scholar] [CrossRef] [PubMed]
- Jendresen, C.B.; Martinussen, J.; Kilstrup, M. The PurR regulon in Lactococcus lactis—Transcriptional regulation of the purine nucleotide metabolism and translational machinery. Microbiology 2012, 158, 2026–2038. [Google Scholar] [CrossRef] [PubMed]
- Stow, R.A.; Bronk, J.R. Purine nucleoside transport and metabolism in isolated rat jejunum. J. Physiol. 1993, 468, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Salati, L.M.; Gross, C.J.; Henderson, L.M.; Savaiano, D.A. Absorption and metabolism of adenine, adenosine-5′-monophosphate, adenosine and hypoxanthine by the isolated vascularly perfused rat small intestine. J. Nutr. 1984, 114, 753–760. [Google Scholar] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Constituent | Final Concentration(/L) |
|---|---|
| D(+) Glucose | 10 g |
| Potassium hydrogen phosphate | 3.1 g |
| di-ammonium hydrogen citrate | 2 g |
| Potassium dihydrogen phosphate | 1.5 g |
| Potassium acetate | 10 g |
| Calcium lactate 5H2O | 1 g |
| Tween-80 | 1 g |
| Heptahydrated magnesium | 500 mg |
| Sodium chloride | 20 mg |
| Hydrated manganese sulphate | 20 mg |
| Cobalt sulphate 7H2O | 500 mg |
| Biotin | 1 mg |
| Cyanocobalamin | 0.02 mg |
| Folic acid | 0.2 mg |
| para-aminobenzoic acid | 0.2 mg |
| Nicotinic acid | 10 mg |
| Calcium pantothenate | 10 mg |
| Riboflavin | 10 mg |
| Pyridoxal HCl | 10 mg |
| Myo-inositol | 10 mg |
| Ascorbic acid | 0.5 mg |
| dl-aminobutyric acid | 100 mg |
| l-phenylalanine | 100 mg |
| l-serine | 100 mg |
| l-threonine | 100 mg |
| l-cycteine Hydrochloride Mnohydrate | 100 mg |
| l-asparagine Mnohydrate | 100 mg |
| l-isoleucine | 100 mg |
| l-tyrosine | 100 mg |
| l-tryptophan | 100 mg |
| l-valine | 100 mg |
| dl-alanine | 200 mg |
| glycine | 200 mg |
| l-histidine Hydrochloride Mnohydrate | 200 mg |
| l-lysine Hydrochloride | 200 mg |
| l-proline | 200 mg |
| l-arginine | 200 mg |
| l-leocine | 200 mg |
| l-aspartic acid | 300 mg |
| l-glutamic acid | 300 mg |
| thymine | 100 mg |
| cytidine | 100 mg |
| 2′-deoxyuridine | 100 mg |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamada, N.; Saito-Iwamoto, C.; Nakamura, M.; Soeda, M.; Chiba, Y.; Kano, H.; Asami, Y. Lactobacillus gasseri PA-3 Uses the Purines IMP, Inosine and Hypoxanthine and Reduces their Absorption in Rats. Microorganisms 2017, 5, 10. https://doi.org/10.3390/microorganisms5010010
Yamada N, Saito-Iwamoto C, Nakamura M, Soeda M, Chiba Y, Kano H, Asami Y. Lactobacillus gasseri PA-3 Uses the Purines IMP, Inosine and Hypoxanthine and Reduces their Absorption in Rats. Microorganisms. 2017; 5(1):10. https://doi.org/10.3390/microorganisms5010010
Chicago/Turabian StyleYamada, Naruomi, Chizuru Saito-Iwamoto, Marie Nakamura, Misato Soeda, Yoshika Chiba, Hiroshi Kano, and Yukio Asami. 2017. "Lactobacillus gasseri PA-3 Uses the Purines IMP, Inosine and Hypoxanthine and Reduces their Absorption in Rats" Microorganisms 5, no. 1: 10. https://doi.org/10.3390/microorganisms5010010