
Microbial Biofilm Community Variation in Flowing Habitats: Potential Utility as Bioindicators of Postmortem Submersion Intervals

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Study Site Descriptions
2.2. Environmental Water Parameters
2.3. Epinecrotic Biofilm Study Design and Sampling
2.4. Epilithic Biofilm Study Design and Processing
2.5. DNA Extraction
2.6. ARISA
2.7. Statistical Analyses
3. Results
3.1. Environmental Water Quality Characteristics
3.2. Microbial Biofilm Communities
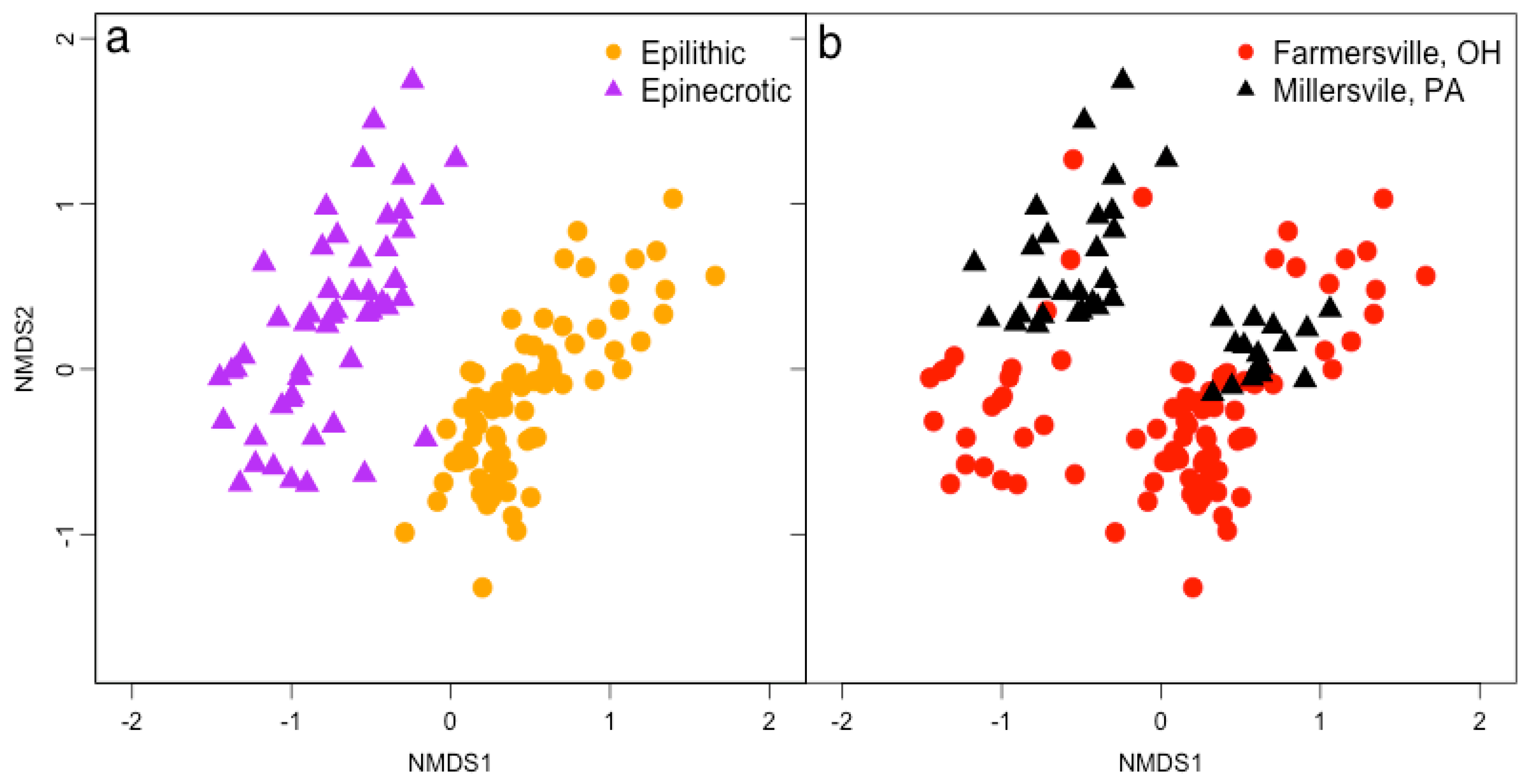

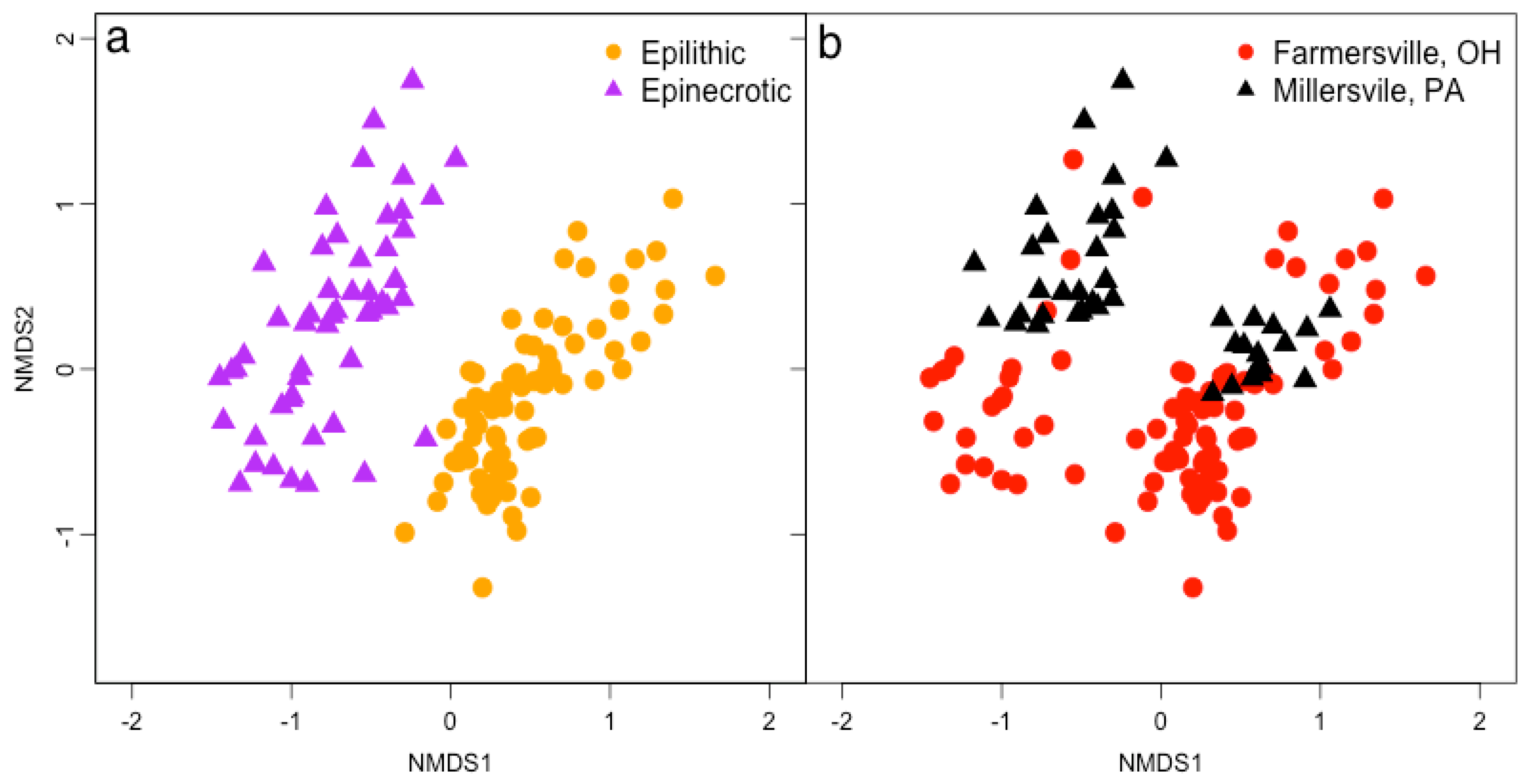{kind=link}
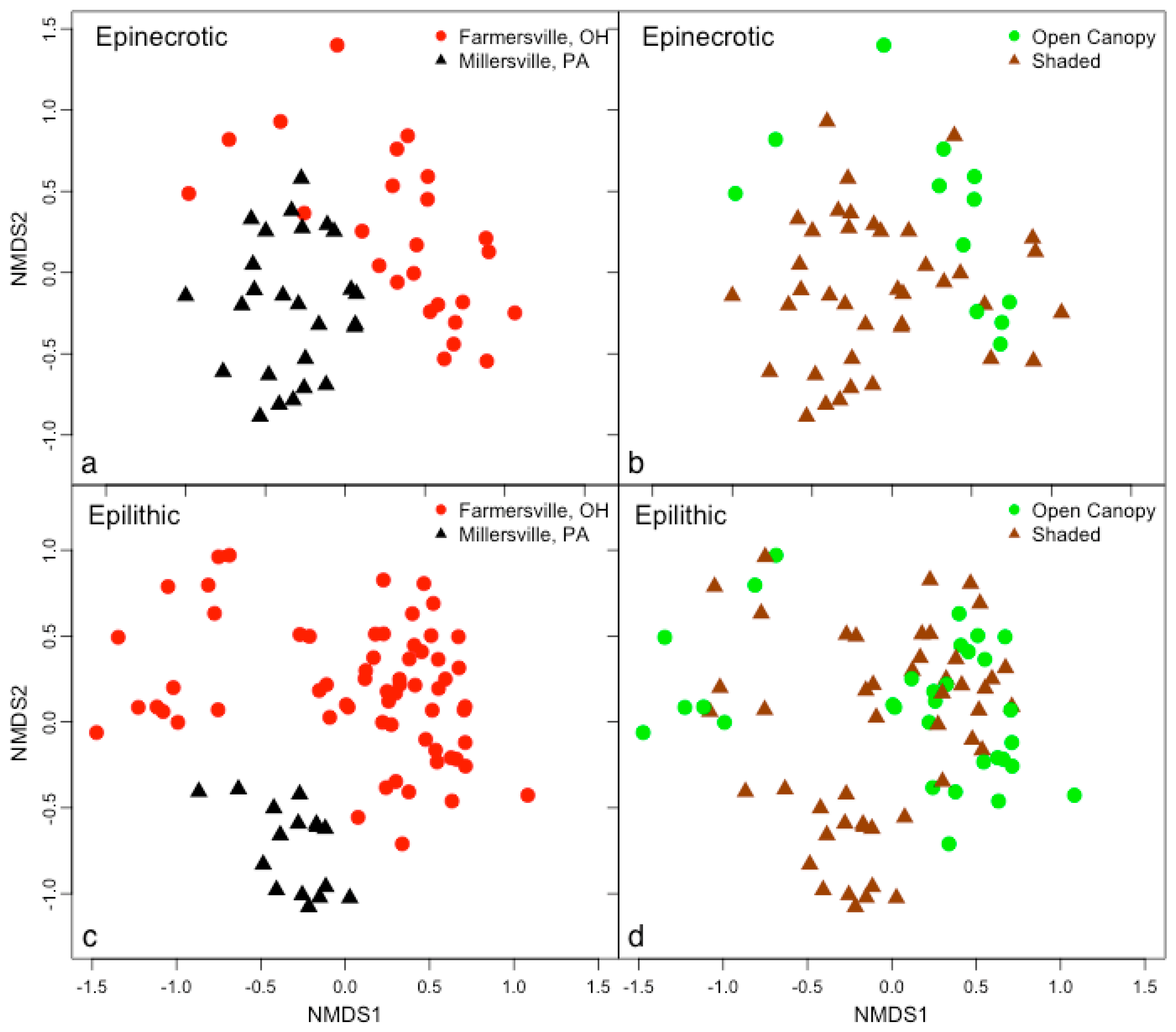{kind=link}
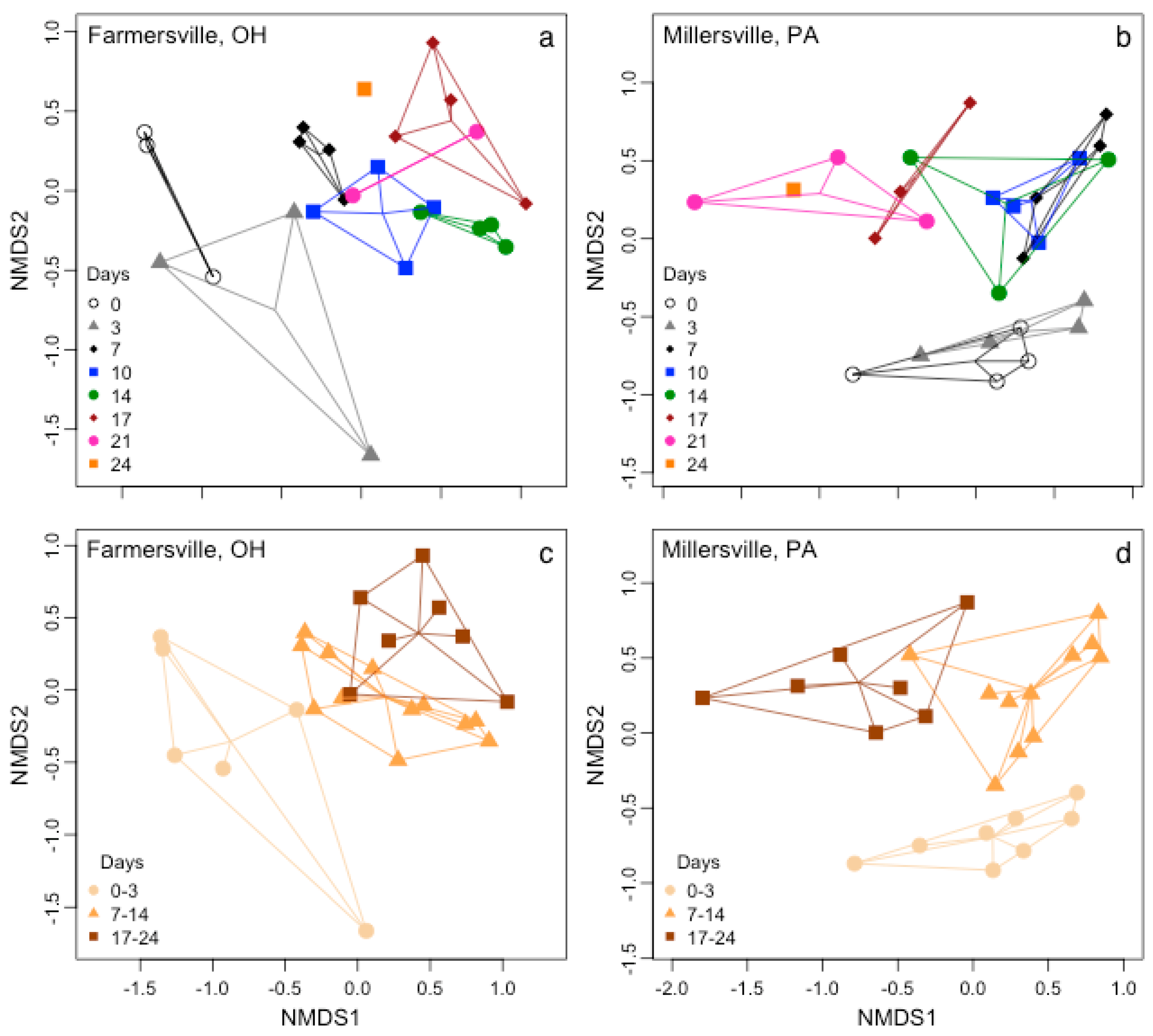{kind=link}
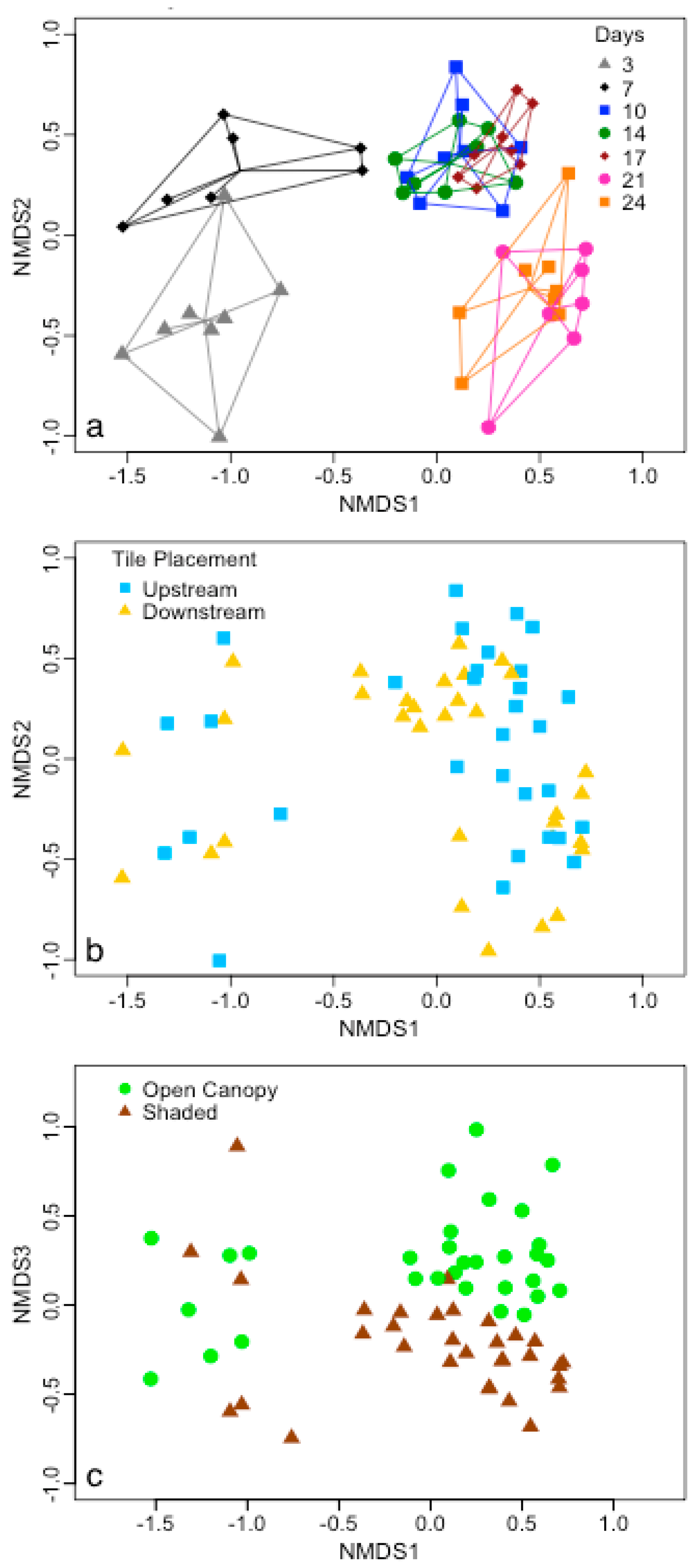{kind=link}
| Date | Time (Days) | NO3− | NO2 | SO42− | NH3 | CaCO3 | TSS | SpCond | TDS | DO | pH | Temp |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 29 June | 0 | 2.35 | 0.054 | 21.0 | 0.10 | 291 | 26 | 617 | 0.40 | 3.78 | 7.92 | 20.6 |
| 02 July | 3 | 2.95 | 0.025 | 20.5 | 0.09 | 295 | 10 | 636 | 0.41 | 5.56 | 7.99 | 21.3 |
| 06 July | 7 | 2.40 | 0.049 | 22.0 | 0.22 | 332 | 20 | 652 | 0.42 | 3.21 | 7.91 | 21.5 |
| 09 July | 10 | 2.55 | 0.016 | 23.0 | 0.13 | 294 | 10 | 652 | 0.42 | 5.28 | 8.02 | 21.8 |
| 13 July | 14 | 3.40 | 0.034 | 23.0 | 0.26 | 307 | - | 650 | 0.42 | 4.57 | 8.17 | 18.6 |
| 16 July | 17 | 2.30 | 0.020 | 23.5 | 0.18 | 335 | 27 | 644 | 0.42 | 3.65 | 8.01 | 21.6 |
| 20 July | 21 | 1.95 | 0.023 | 27.0 | 0.15 | 308 | 20 | 644 | 0.42 | 2.84 | 7.96 | 21.5 |
| 23 July | 24 | 1.95 | 0.019 | 27.0 | 0.14 | 295 | 28 | 653 | 0.43 | 3.68 | 8.04 | 21.3 |
| Mean | - | 2.48 ± 0.49 | 0.030 ± 0.014 | 23 ± 2 | 0.16 ± 0.06 | 307 ± 17 | 20 ± 7 | 643 ± 12 | 0.42 ± 0.01 | 4.07 ± 0.97 | 8.00 ± 0.08 | 21.0 ± 1.1 |
| Upstream/Shaded | - | 2.39 ± 0.66 | 0.034 ± 0.024 | 24 ± 4 | 0.14 ± 0.05 | 313 ± 22 | 15 ± 6 | 666 ± 25 | 0.43 ± 0.01 | 2.87 ± 0.96 | 7.85 ± 0.10 | 19.7 ± 1.4 |
| Downstream/Open | - | 2.58 ± 0.67 | 0.025 ± 0.020 | 22 ± 1 | 0.17 ± 0.11 | 302 ± 18 | 34 ± 19 | 621 ± 13 | 0.40 ± 0.01 | 5.27 ± 1.36 | 8.16 ± 0.08 | 22.3 ± 0.7 |
| Date | Time (Days) | DO | SpCond | ORP | pH | Salinity | TDS | Temperature |
|---|---|---|---|---|---|---|---|---|
| 26 June | 0 | 6.94 | 966 | 93.0 | 8.2 | 0.48 | 0.483 | 18.3 |
| 29 June | 3 | 4.00 | 928 | 64.5 | 8.1 | 0.46 | 0.464 | 20.5 |
| 03 July | 7 | 6.80 | 1038 | 66.8 | 8.5 | 0.52 | 0.519 | 20.1 |
| 06 July | 10 | 6.55 | 950 | 47.8 | 8.2 | 0.47 | 0.475 | 20.7 |
| 10 July | 14 | 4.05 | 908 | 37.2 | 8.1 | 0.45 | 0.454 | 22.2 |
| 13 July | 17 | 4.34 | 1130 | 37.9 | 8.1 | 0.57 | 0.565 | 20.1 |
| 17 July | 21 | 4.78 | 963 | 36.7 | 8.1 | 0.48 | 0.459 | 23.5 |
| 20 July | 24 | 5.59 | 779 | 29.6 | 8.1 | 0.38 | 0.389 | 21.8 |
| Mean | - | 5.38 ± 2.25 | 957 ± 101 | 51.7 ± 21.5 | 8.2 ± 0.1 | 0.47 ± 0.05 | 0.476 ± 0.051 | 20.9 ± 1.6 |
| Day | Dayton | Millersville | ||
|---|---|---|---|---|
| Pseudo-F | p-Value | Pseudo-F | p-Value | |
| 0 vs. 3 | 1.7683 | 0.0292 | 1.5209 | 0.8925 |
| 0 vs. 7 | 2.0327 | 0.0042 | 2.1454 | 0.0441 |
| 0 vs. 10 | 2.7234 | 0.6000 | 2.2779 | 0.0336 |
| 0 vs. 14 | 4.2148 | 0.6000 | 3.6802 | 0.1000 |
| 0 vs. 17 | 3.4430 | 0.6000 | 2.6717 | 0.1500 |
| 0 vs. 21 | 0.0286 | 1.0000 | 1.8198 | 0.5042 |
| 3 vs. 7 | 1.3976 | 1.0000 | 1.7396 | 0.0021 |
| 3 vs. 10 | 1.6904 | 1.0000 | 1.9337 | 0.0357 |
| 3 vs. 14 | 2.8101 | 0.6000 | 3.2167 | 0.0333 |
| 3 vs. 17 | 2.2252 | 0.0042 | 2.1349 | 0.1000 |
| 3 vs. 21 | 1.2239 | 1.0000 | 1.4336 | 1.0000 |
| 7 vs. 10 | 1.2510 | 1.0000 | 1.6097 | 1.0000 |
| 7 vs. 14 | 2.5020 | 0.0021 | 2.9106 | 0.1333 |
| 7 vs. 17 | 2.0530 | 0.0021 | 2.1186 | 0.0042 |
| 7 vs. 21 | 1.1205 | 1.0000 | 1.4207 | 0.2375 |
| 10 vs. 14 | 0.9974 | 1.0000 | 2.8144 | 0.2000 |
| 10 vs. 17 | 1.8117 | 1.0000 | 1.8826 | 0.6999 |
| 10 vs. 21 | 1.0865 | 1.0000 | 1.1429 | 1.0000 |
| 14 vs. 17 | 2.2648 | 0.6468 | 2.1364 | 1.0000 |
| 14 vs. 21 | 1.6193 | 1.0000 | 2.4703 | 0.3793 |
| 17 vs. 21 | 0.7587 | 1.0000 | 0.7478 | 1.0000 |
| 0–3 vs. 7–14 | 2.9224 | 0.0018 | 4.4026 | 0.0003 |
| 0–3 vs. 17–24 | 3.0783 | 0.0012 | 4.2985 | 0.0006 |
| 7–14 vs. 17–24 | 2.4225 | 0.0087 | 3.2061 | 0.0006 |
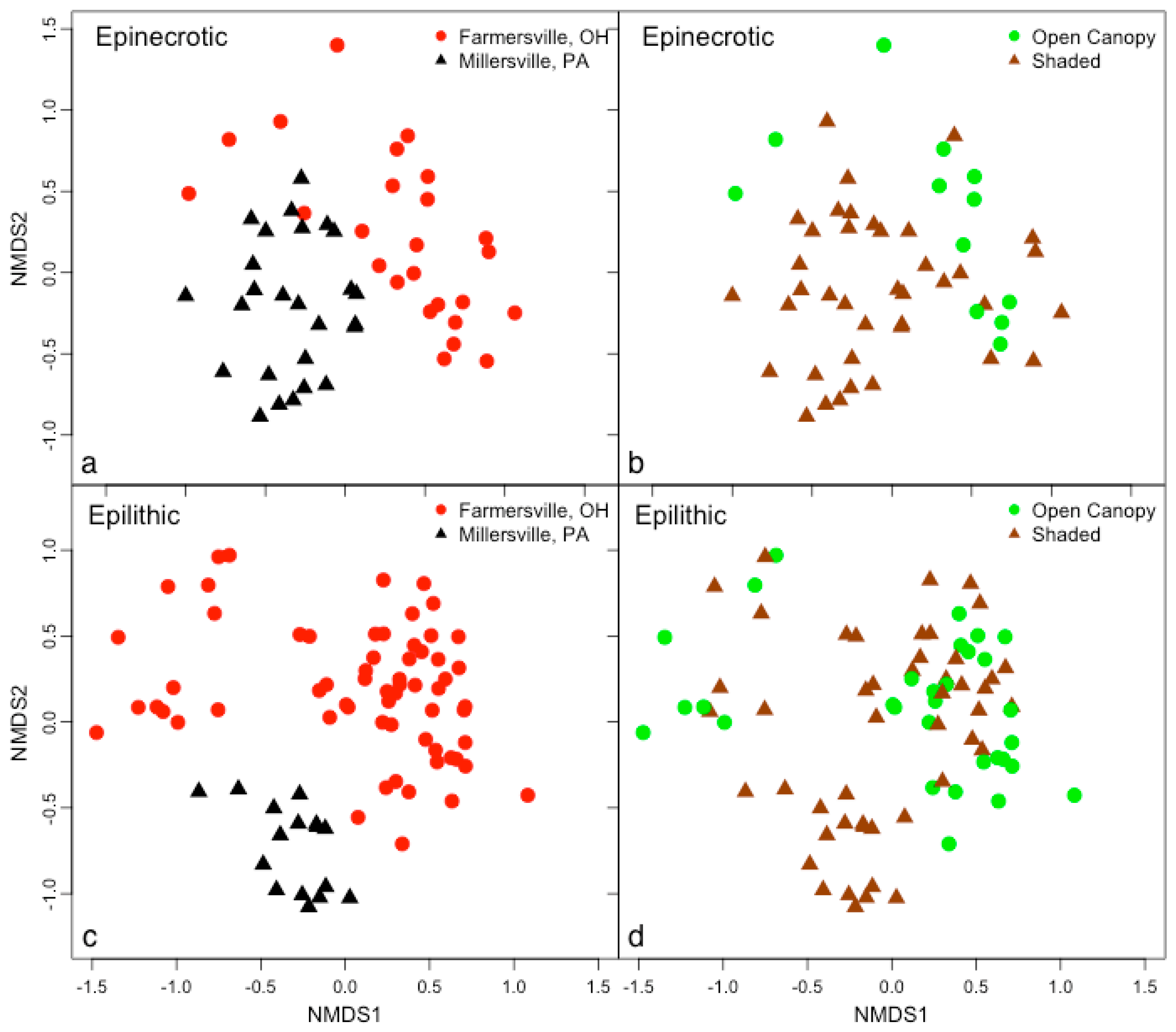
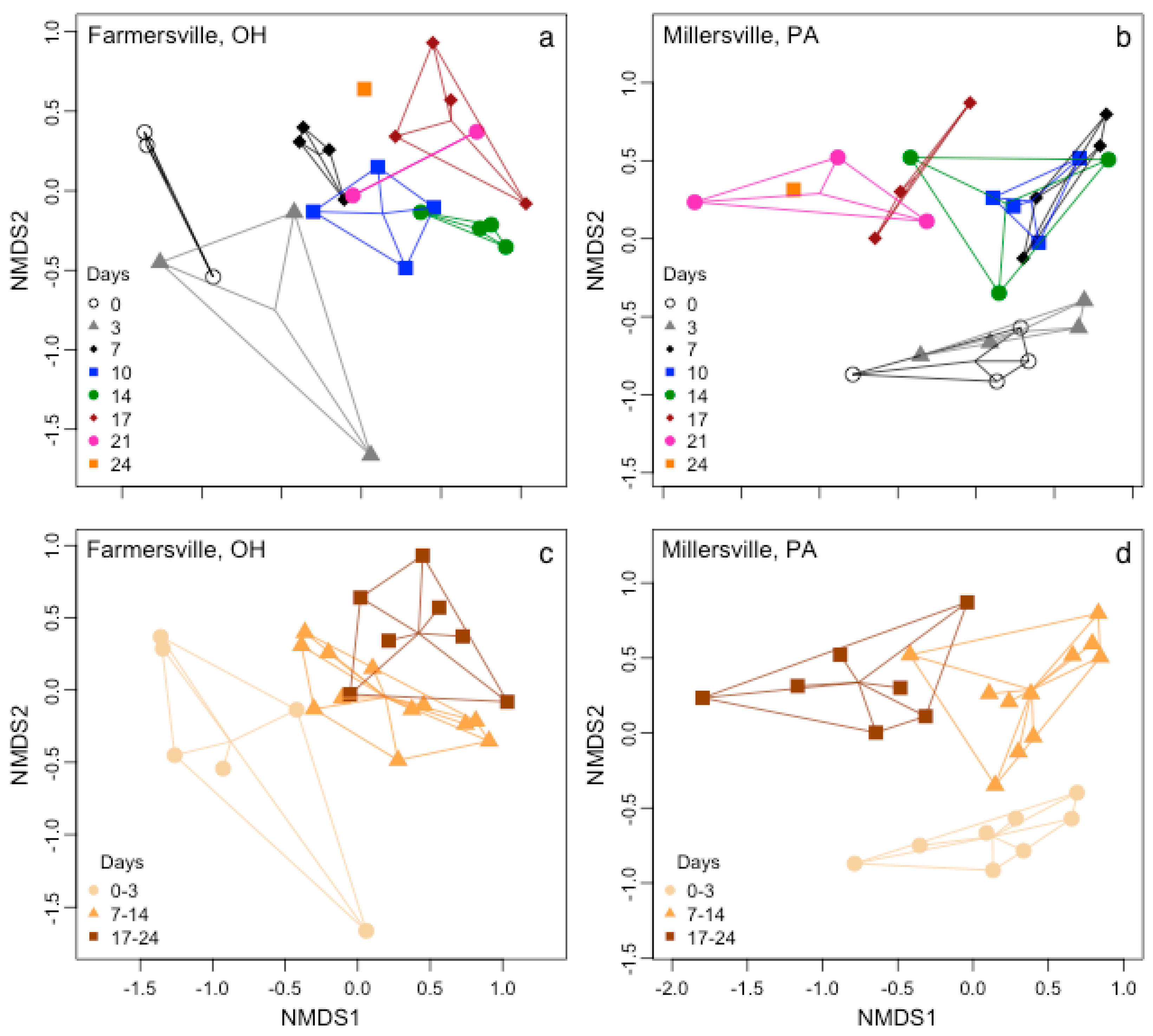
4. Discussion
4.1. Substrate Type Affects Microbial Biofilm Communities
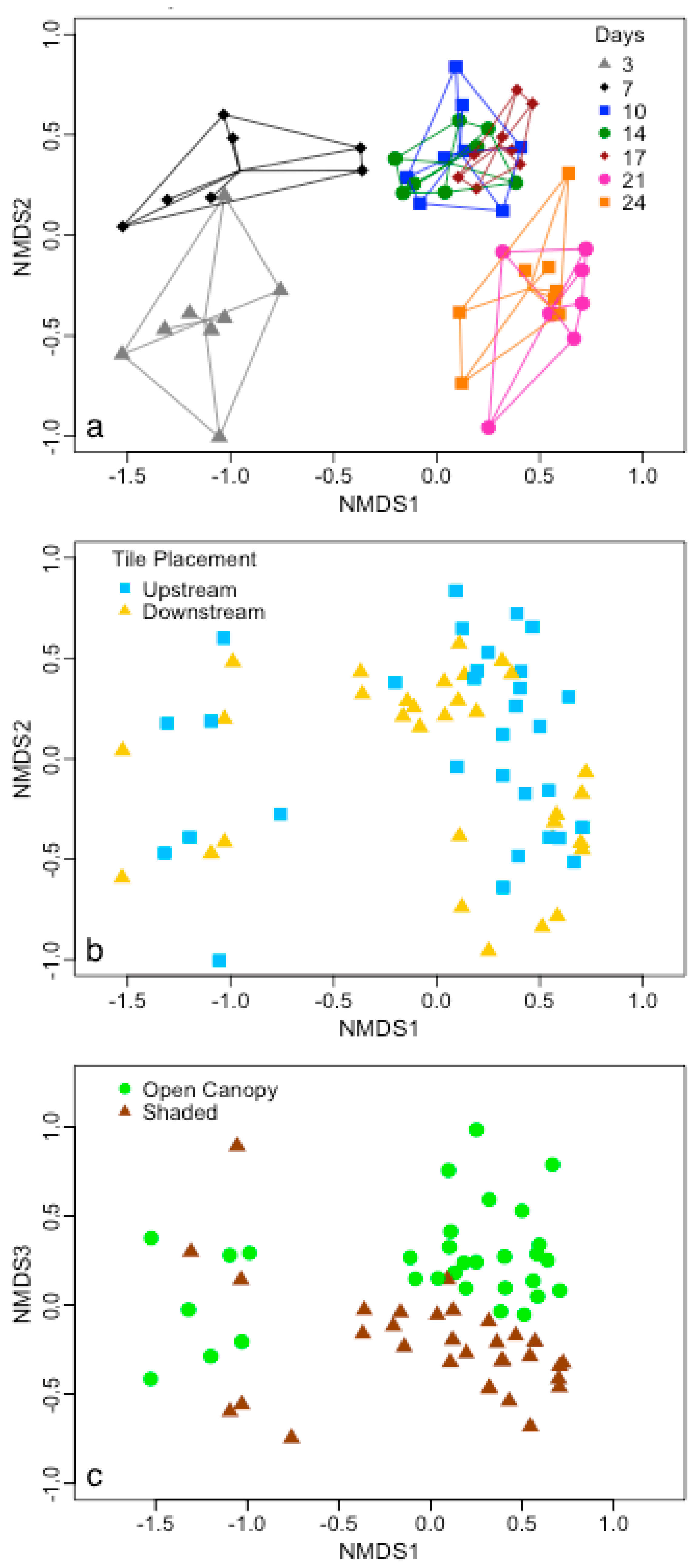
4.2. Epinecrotic Community Response to Environmental Conditions
4.3. Biofilm Community Succession
5. Conclusions and Potential for Applications and Management of Biofilms
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gleason, H.A. The individualistic concept of the plant association. Bull. Torrey Bot. Club 1926, 53, 7–26. [Google Scholar] [CrossRef]
- Clements, F.E. Nature and structure of the climax. Ecology 1936, 24, 252–284. [Google Scholar] [CrossRef]
- Bormann, F.; Likens, G. Patterns and Process in a Forested Ecosystem; Springer-Verlag: New York, NY, USA, 1979. [Google Scholar]
- Weiher, E.; Keddy, P. Ecological Assembly Rules; Perspectives, Advances, Retreats; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Fierer, N.; Nemergut, D.; Knight, R.; Craine, J.M. Changes through time: Integrating microorganisms into the study of succession. Res. Microbiol. 2010, 161, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Barrios, M.; Wolff, M. Initial study of arthropods succession and pig carrion decomposition in two freshwater ecosystems in the Colombian Andes. Forensic Sci. Int. 2011, 212, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H. Microbial biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef] [PubMed]
- Giller, P.S.; Malmqvist, B. The Biology of Streams and Rivers; Crawley, M.J., Little, C., Southwood, T.R.E., Ulfstrand, S., Eds.; Oxford University Press Inc.: New York, NY, USA, 1998. [Google Scholar]
- Hoagland, K.D.; Roemer, S.C.; Rosowski, J.R.; Rosowski, R. Colonization and community structure of two periphyton assemblages, with emphasis on the diatoms (Bacillariophyceae). Am. J. Bot. 1982, 69, 188–213. [Google Scholar] [CrossRef]
- Freeman, C.; Lock, M.A. The biofilm polysaccharide matrix: A buffer against changing organic substrate supply? Limnol. Oceanogr. 1995, 40, 273–278. [Google Scholar] [CrossRef]
- Sutherland, I.W. The biofilm matrix—An immobilized but dynamic microbial environment. Trends Microbiol. 2001, 9, 222–227. [Google Scholar] [CrossRef]
- Freeman, C.; Chapman, P.J.; Gilman, K.; Lock, M.A.; Reynolds, B.; Wheater, H.S. Ion exchange mechanisms and the entrapment of nutrients by river biofilms. Hydrobiologia 1995, 297, 61–65. [Google Scholar] [CrossRef]
- Battin, T.J.; Kaplan, L.A.; Newbold, J.D.; Hansen, C.M.E. Contributions of microbial biofilms to ecosystem processes in stream mesocosms. Nature 2003, 426, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Augspurger, C.; Karwautz, C.; MuBmann, M.; Daims, H.; Battin, T.J. Drivers of bacterial colonization patterns in stream biofilms. FEMS Microbiol. Ecol. 2010, 72, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Sinsabaugh, R.L.; Repert, D.; Weiland, T.; Golladay, S.W.; Linkins, A.E. Exoenzyme accumulation in epilithic biofilms. Hydrobiologia 1991, 222, 29–37. [Google Scholar] [CrossRef]
- Romaní, A.M.; Fischer, H.; Mille-lindblom, C.; Tranvik, L.J.; Tranvik, J.; Romani, A.M. Interactions of bacteria and fungi on decomposing litter: Differential extracellular enzyme activities. Ecology 2006, 87, 2559–2569. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Golladay, S.W.; Linkins, A.E. Comparison of epilithic and epixylic biofilm development in a boreal river. Freshw. Biol. 1991, 25, 179–187. [Google Scholar] [CrossRef]
- Tank, J.L.; Dodds, W.K. Nutrient limitation of epilithic and epixylic biofilms in ten North American streams. Freshw. Biol. 2003, 48, 1031–1049. [Google Scholar] [CrossRef]
- Hladyz, S.; Cook, R.A.; Petrie, R.; Nielsen, D.L. Influence of substratum on the variability of benthic biofilm stable isotope signatures: Implications for energy flow to a primary consumer. Hydrobiologia 2011, 664, 135–146. [Google Scholar] [CrossRef]
- Haefner, J.N.; Wallace, J.R.; Merritt, R.W. Pig decomposition in lotic aquatic systems: The potential use of algal growth in establishing a postmortem submersion interval (PMSI). J. Forensic Sci. 2004, 49, 1–7. [Google Scholar] [CrossRef]
- Zimmerman, K.A.; Wallace, J.R. The potential to determine a postmortem submersion interval based on algal/diatom diversity on decomposing mammalian carcasses in brackish ponds in Delaware. J. Forensic Sci. 2008, 53, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Docherty, K.M.; Young, K.C.; Maurice, P.A.; Bridgham, S.D. Dissolved organic matter concentration and quality influences upon structure and function of freshwater microbial communities. Microb. Ecol. 2006, 52, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Singer, G.; Besemer, K.; Schmitt-Kopplin, P.; Hodl, I.; Battin, T.J. Physical heterogeneity increases biofilm resource use and its molecular diversity in stream mesocosms. PLoS ONE 2010, 5, e9988. [Google Scholar] [CrossRef] [PubMed]
- Smucker, N.J.; Vis, M.L. Acid mine drainage affects the development and function of epilithic biofilms in streams. J. N. Am. Benthol. Soc. 2011, 30, 728–738. [Google Scholar] [CrossRef]
- Lang, J.; McEwan, R.; Benbow, M. Abiotic autumnal organic matter deposition and grazing disturbance effects on epilithic biofilm succession. FEMS Microbiol. Ecol. 2015, 91. [Google Scholar] [CrossRef] [PubMed]
- Casamatta, D.A.; Verb, R.G. Algal colonization of submerged carcasses in a mid-order woodland stream. J. Forensic Sci. 2000, 45, 1280–1285. [Google Scholar] [CrossRef] [PubMed]
- Dickson, G.C.; Poulter, R.T.M.; Maas, E.W.; Probert, P.K.; Kieser, J.A. Marine bacterial succession as a potential indicator of postmortem submersion interval. Forensic Sci. Int. 2011, 209, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Benbow, M.; Pechal, J.; Lang, J.; Erb, R.; Wallace, J. The potential of high-throughput metagenomic sequencing of aquatic bacterial communities to estimate the postmortem submersion interval. J. Forensic Sci. 2015, 60, 1500–1510. [Google Scholar] [CrossRef] [PubMed]
- Tomberlin, J.; Benbow, M. Forensic Entomology: International Dimensions and Frontiers; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Vance, G.M.; VanDyk, J.K.; Rowley, W.A. A device for sampling aquatic insects associated with carrion in water. J. Forensic Sci. 1995, 40, 479–482. [Google Scholar] [CrossRef]
- Keiper, J.B.; Chapman, E.G.; Foote, B.A. Midge larvae (Diptera: Chironomidae) as indicators of postmortem submersion interval of carcasses in a woodland stream: A preliminary report. J. Forensic Sci. 1997, 42, 1074–1079. [Google Scholar] [CrossRef] [PubMed]
- Hobischak, N.R.; Anderson, G.S. Time of submergence using aquatic invertebrate succession and decompositional changes. J. Forensic Sci. 2002, 47, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.R.; Merritt, R.W.; Kimbirauskas, R.; Benbow, M.E.; McIntosh, M. Caddisflies assist with homicide case: Determining a postmortem submersion interval using aquatic insects. J. Forensic Sci. 2008, 53, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Claeson, S.M.; Li, J.L.; Compton, J.E.; Bisson, P.A. Response of nutrients, biofilm, and benthic insects to salmon carcass addition. Can. J. Fish. Aquat. Sci. 2006, 63, 1230–1241. [Google Scholar] [CrossRef]
- Parmenter, R.R.; Lamarra, V.A. Nutrient cycling in a freshwater marsh: The decomposition of fish and waterfowl carrion. Limnol. Oceanogr. 1991, 36, 976–987. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Adler, P.H. Seasonal colonization and decomposition of rat carrion in water and on land in an open field in South Carolina. J. Med. Entomol. 1998, 35, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Dick, I.P.; Scott, R.C. Pig ear skin: As an in-vitro model for human skin permeability. J. Pharm. Pharmacol. 1992, 44, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Sekkat, N.; Kalia, Y.N.; Guy, R.H. Biophysical study of porcine ear skin in vitro and its comparison to human skin in vivo. J. Pharm. Sci. 2002, 91, 2376–2381. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, U.; Kaiser, M.; Toll, R.; Mangelsdorf, S.; Audring, H.; Otberg, N.; Sterry, W.; Lademann, J. Porcine ear skin: An in vitro model for human skin. Skin Res. Technol. 2007, 13, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Schoenly, K.G.; Haskell, N.H.; Hall, R.D.; Gbur, J.R. Comparative performance and complementarity of four sampling methods and arthropod preference tests from human and porcine remains at the Forensic Anthropology Center in Knoxville, Tennessee. J. Med. Entomol. 2007, 44, 881–894. [Google Scholar] [CrossRef] [PubMed]
- Pechal, J.L.; Benbow, M.E.; Crippen, T.L.; Tarone, A.M.; Tomberlin, J.K. Delayed insect access alters carrion decomposition and necrophagous insect community assembly. Ecosphere 2014, 5. [Google Scholar] [CrossRef]
- Pechal, J.L.J.; Crippen, T.L.T.; Benbow, M.E.; Tarone, A.M.; Dowd, S.; Tomberlin, J.K. The potential use of bacterial community succession in forensics as described by high throughput metagenomic sequencing. Int. J. Leg. Med. 2014, 128, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Steinman, A.D.; Lamberti, G.A.; Leavitt, P.R. Biomass and pigments of benthic algae. In Methods in Stream Ecology; Hauer, F.R., Lamberti, G.A., Eds.; Academic Press: New York, NY, USA, 2007; pp. 357–380. [Google Scholar]
- Zhou, J.; Bruns, M.A.; Tiedje, J.M. DNA recovery from soils of diverse composition. Appl. Environ. Microbiol. 1996, 62, 316–322. [Google Scholar] [PubMed]
- Miller, D.N.; Bryant, J.E.; Madsen, E.L.; Ghiorse, W.C. Evaluation and optimization of DNA extraction and purification procedures for soil and sediment samples. Appl. Environ. Microbiol. 1999, 65, 4715–4724. [Google Scholar] [PubMed]
- Lear, G.; Dong, Y.; Lewis, G. Comparison of methods for the extraction of DNA from stream epilithic biofilms. Antonie Van Leeuwenhoek 2010, 98, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.M.; Triplett, E.W. Automated approach for ribosomal intergenic spacer analysis of microbial diversity and its application to freshwater bacterial communities. Appl. Environ. Microbiol. 1999, 65, 4630–4636. [Google Scholar] [PubMed]
- Bienhold, C.; Boetius, A.; Ramette, A. The energy-diversity relationship of complex bacterial communities in Arctic deep-sea sediments. ISME J. 2012, 6, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Van Dorst, J.; Bissett, A.; Palmer, A.S.; Brown, M.; Snape, I.; Stark, J.S.; Raymond, B.; McKinlay, J.; Ji, M.; Winsley, T.; et al. Community fingerprinting in a sequencing world. FEMS Microbiol. Ecol. 2014, 89, 316–330. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, M.; Brusetti, L.; Quatrini, P.; Borin, S.; Puglia, A.M.; Rizzi, A.; Zanardini, E.; Sorlini, C.; Corselli, C.; Daffonchio, D. Comparison of different primer sets for use in automated ribosomal intergenic spacer analysis of complex bacterial communities. Appl. Environ. Microbiol. 2004, 70, 6147–6156. [Google Scholar] [CrossRef] [PubMed]
- Fechner, L.C.; Vincent-Hubert, F.; Gaubert, P.; Bouchez, T.; Gourlay-Francé, C.; Tusseau-Vuillemin, M.H. Combined eukaryotic and bacterial community fingerprinting of natural freshwater biofilms using automated ribosomal intergenic spacer analysis. FEMS Microbiol. Ecol. 2010, 74, 542–553. [Google Scholar] [CrossRef] [PubMed]
- Rees, G.N.; Baldwin, D.S.; Watson, G.O.; Perryman, S.; Nielsen, D.L. Ordination and significance testing of microbial community composition derived from terminal restriction fragment length polymorphisms: Application of multivariate statistics. Antonie Van Leeuwenhoek 2005, 86, 339–347. [Google Scholar] [CrossRef]
- Walsh, C.J. The TRFLP Peak Sorting Function for EXCEL. Available online: http://www.sci.monash.edu.au/wsc/staff/walsh/treeflap.xls (accessed on 17 December 2015).
- Ramette, A. Quantitative community fingerprinting methods for estimating the abundance of operational taxonomic units in natural microbial communities. Appl. Environ. Microbiol. 2009, 75, 2495–2505. [Google Scholar] [CrossRef] [PubMed]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austr. Ecol. 2001, 26, 32–46. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. Vegan: Community Ecology Package. Available online: http://cran.r-project.org/package=vegan (accessed on 20 June 2015).
- Das, M.; Royer, T.V.; Leff, L.G. Diversity of fungi, bacteria, and actinomycetes on leaves decomposing in a stream. Appl. Environ. Microbiol. 2007, 73, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Sabater, S.; Gregory, S.V.; Sedell, J.R. Community dynamics and metabolism of benthic algae colonizing wood and rock substrata in a forest stream. J. Phycol. 1998, 34, 561–567. [Google Scholar] [CrossRef]
- Golladay, S.W.; Sinsabaugh, R.L. Biofilm development on leaf and wood surfaces in a boreal river. Freshw. Biol. 1991, 25, 437–450. [Google Scholar] [CrossRef]
- Van der Grinten, E.; Janssen, M.; Simis, S.G.H.; Barranguet, C.; Admiraal, W. Phosphate regime structures species composition in cultured phototrophic biofilms. Freshw. Biol. 2004, 49, 369–381. [Google Scholar] [CrossRef]
- Lock, M.; Hynes, H. The fate of “dissolved” organic carbon derived from autumn-shed maple leaves (Acer saccharurn) in a temperate hard-water stream. Limnol. Oceanogr. 1976, 21, 436–443. [Google Scholar] [CrossRef]
- McNamara, C.J.; Leff, L.G. Response of biofilm bacteria to dissolved organic matter from decomposing maple leaves. Microb. Ecol. 2004, 48, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Besemer, K.; Singer, G.; Hödl, I.; Battin, T.J. Bacterial community composition of stream biofilms in spatially variable-flow environments. Appl. Environ. Microbiol. 2009, 75, 7189–7195. [Google Scholar] [CrossRef] [PubMed]
- Ledger, M.E.; Hildrew, A.G. Temporal and spatial variation in the epilithic biofilm of an acid stream. Freshw. Biol. 1998, 40, 655–670. [Google Scholar] [CrossRef]
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lang, J.M.; Erb, R.; Pechal, J.L.; Wallace, J.R.; McEwan, R.W.; Benbow, M.E. Microbial Biofilm Community Variation in Flowing Habitats: Potential Utility as Bioindicators of Postmortem Submersion Intervals. Microorganisms 2016, 4, 1. https://doi.org/10.3390/microorganisms4010001
Lang JM, Erb R, Pechal JL, Wallace JR, McEwan RW, Benbow ME. Microbial Biofilm Community Variation in Flowing Habitats: Potential Utility as Bioindicators of Postmortem Submersion Intervals. Microorganisms. 2016; 4(1):1. https://doi.org/10.3390/microorganisms4010001
Chicago/Turabian StyleLang, Jennifer M., Racheal Erb, Jennifer L. Pechal, John R. Wallace, Ryan W. McEwan, and Mark Eric Benbow. 2016. "Microbial Biofilm Community Variation in Flowing Habitats: Potential Utility as Bioindicators of Postmortem Submersion Intervals" Microorganisms 4, no. 1: 1. https://doi.org/10.3390/microorganisms4010001