Emergence of Algal Blooms: The Effects of Short-Term Variability in Water Quality on Phytoplankton Abundance, Diversity, and Community Composition in a Tidal Estuary

Abstract
:1. Introduction
2. Experimental Section
2.1. Study Site
2.2. Methods and Materials
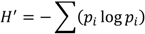
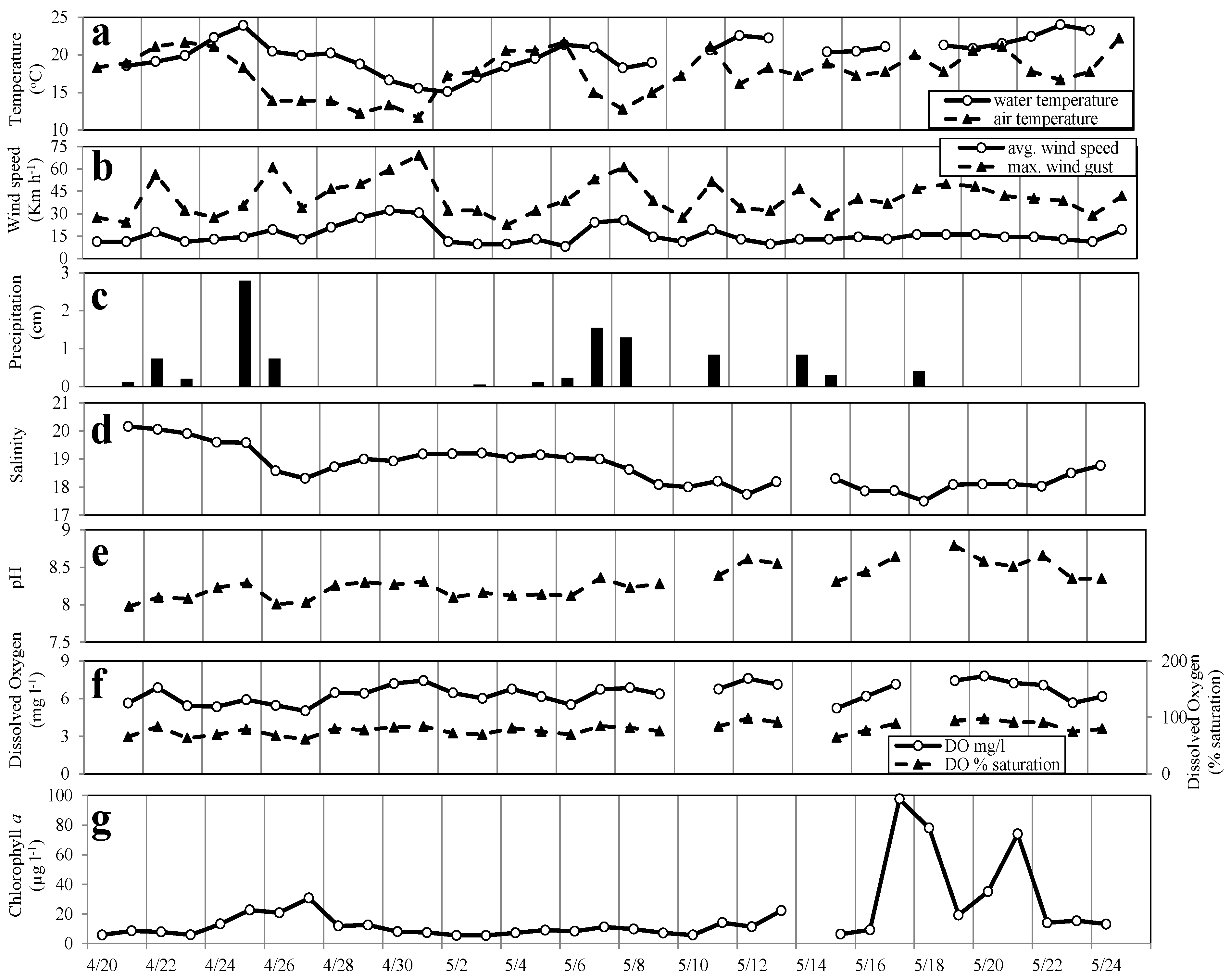
3. Results and Discussion
3.1. Meteorological and Physical Parameters
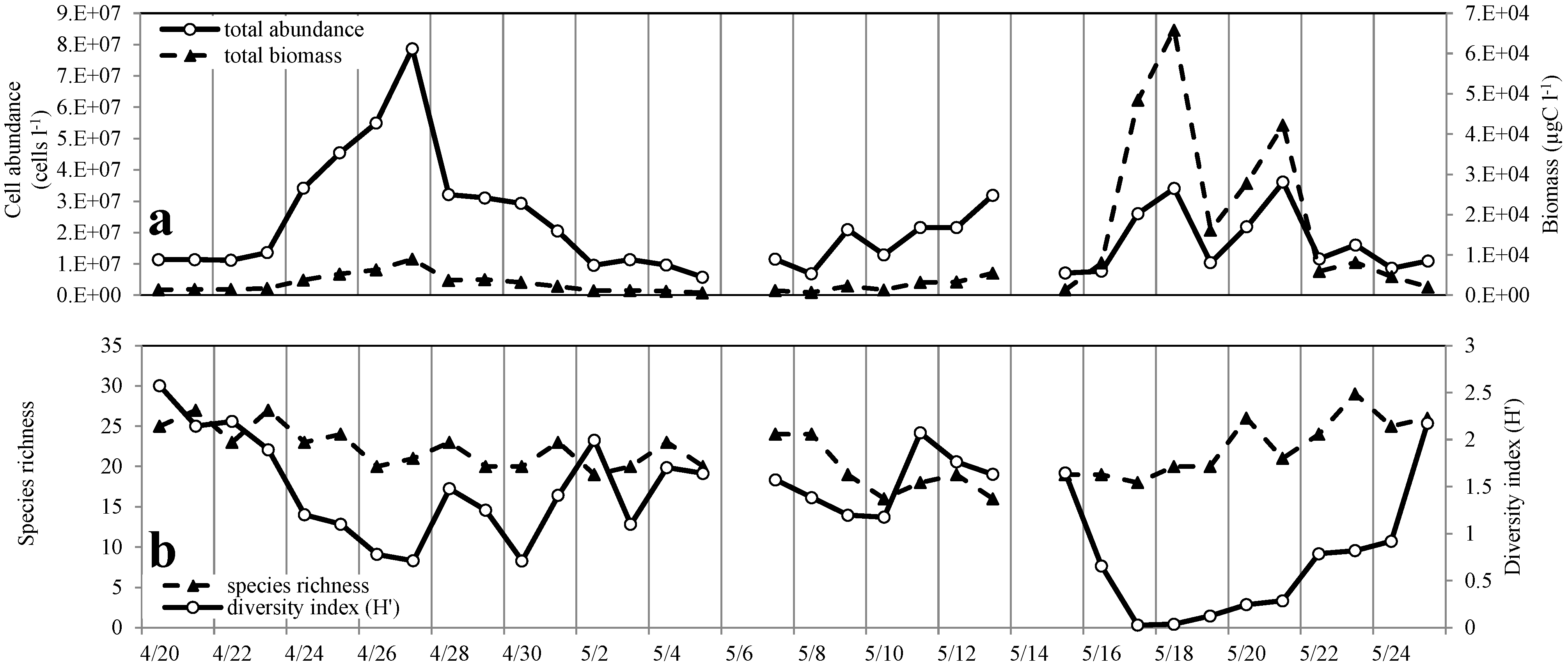
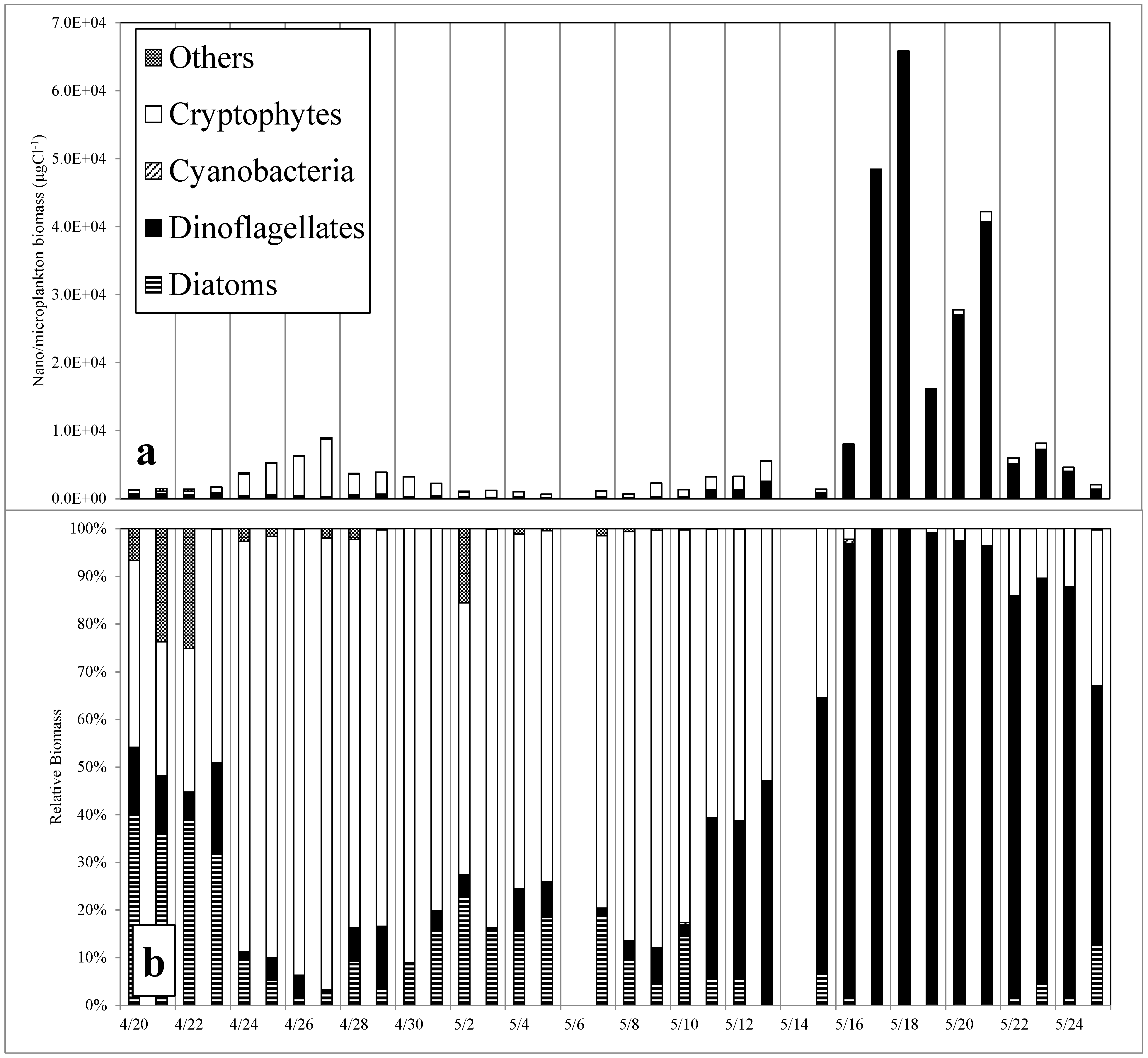
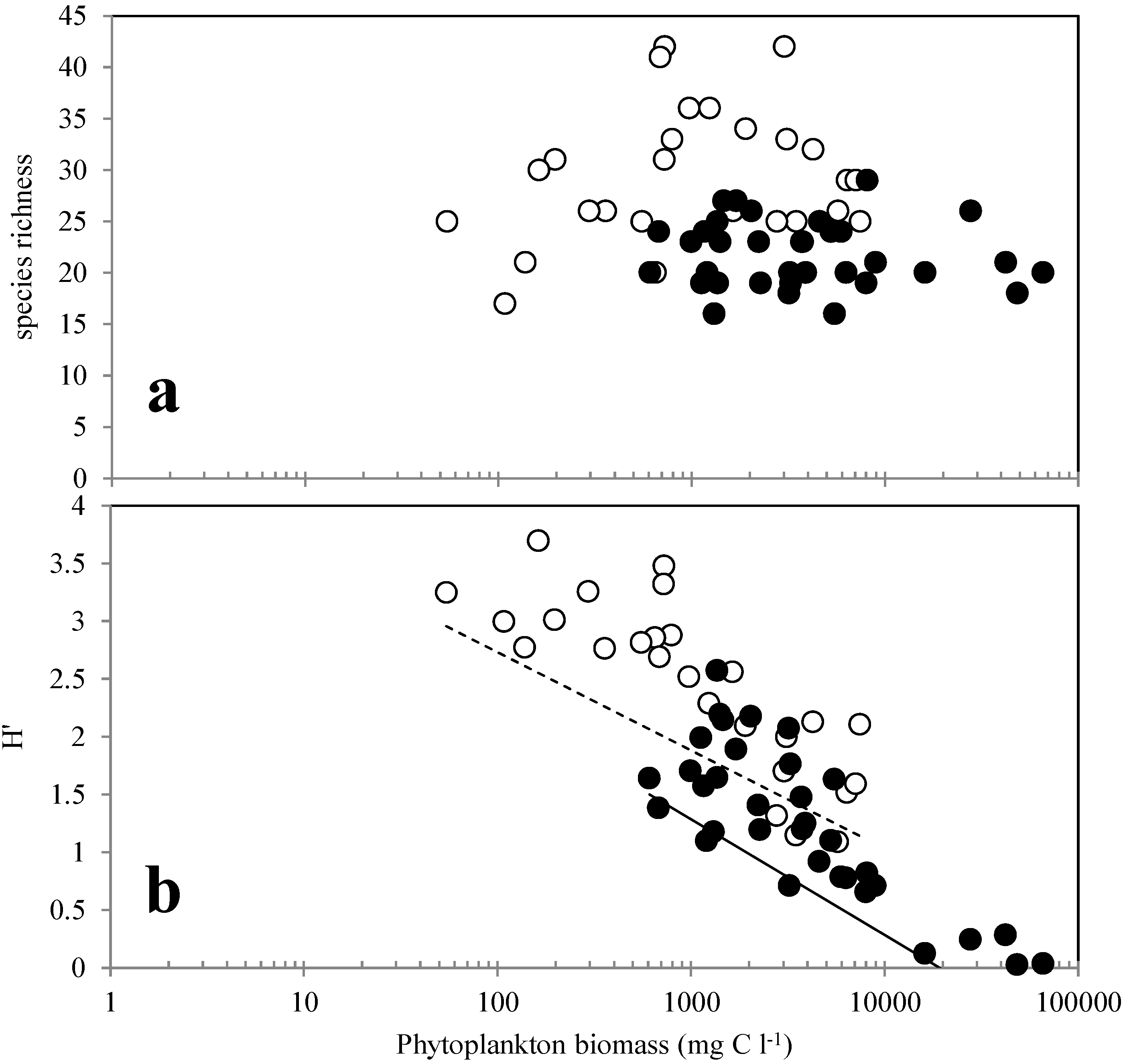
3.2. Phytoplankton Abundance, Composition and Diversity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytoplankton Taxa | Abundance | ||
|---|---|---|---|
| Mean Value | Minimum Value | Maximum Value | |
| Diatoms | |||
| unidentified Centrales 10–30 µm | 2.0 × 106 | 1.0 × 103 | 5.3 × 106 |
| unidentified Centrales 30–60 µm | 1.0 × 104 | 2.6 × 102 | 1.1 × 105 |
| Chaetoceros pendulus | 6.7 × 102 | 2.6 × 102 | 1.0 × 103 |
| Chaetoceros sp. | 1.2 × 105 | 7.7 × 102 | 4.3 × 105 |
| Cocconeis sp. | 2.8 × 102 | 2.6 × 102 | 5.1 × 102 |
| Coscinodiscus sp. | 5.7 × 102 | 2.6 × 102 | 1.3 × 103 |
| Cyclotella sp. | 1.1 × 105 | 5.1 × 102 | 4.3 × 105 |
| Cylindrotheca closterium | 5.3 × 102 | 2.6 × 102 | 1.5 × 103 |
| Dactyliosolen fragilissimus | 2.9 × 104 | 5.1 × 102 | 3.2 × 105 |
| Gyrosigma fasciola | 3.1 × 102 | 2.6 × 102 | 5.1 × 102 |
| Leptocylindrus minimus | 7.7 × 104 | 7.7 × 102 | 5.4 × 105 |
| Navicula sp. | 7.3 × 102 | 2.6 × 102 | 3.8 × 103 |
| Nitzchia sp. | 2.6 × 102 | 2.6 × 102 | 2.6 × 102 |
| unidentified Pennales 10–30 µm | 2.6 × 105 | 2.6 × 102 | 8.7 × 105 |
| unidentified Pennales30–60 µm | 8.9 × 103 | 2.6 × 102 | 1.1 × 105 |
| unidentified Pennales > 60 µm | 9.0 × 102 | 2.6 × 102 | 2.8 × 103 |
| Pleurosigma sp. | 2.6 × 102 | 2.6 × 102 | 2.6 × 102 |
| Rhizosolenia setigera | 1.2 × 103 | 2.6 × 102 | 3.8 × 103 |
| Skeletonema costatum | 1.0 × 105 | 1.0 × 103 | 9.7 × 105 |
| Thalassiosira sp. | 6.7 × 102 | 2.6 × 102 | 1.3 × 103 |
| Dinoflagellates | |||
| Akashwio sanguinea | 3.0 × 103 | 2.6 × 102 | 1.8 × 104 |
| Cochlodinium polykrikoides | 1.1 × 104 | 5.1 × 102 | 3.7 × 104 |
| unidentified dinoflagellate | 1.8 × 105 | 5.1 × 102 | 5.4 × 105 |
| Dinophysis punctata | 5.4 × 102 | 2.6 × 102 | 2.0 × 103 |
| Diplopsalis lenticula | 3.1 × 102 | 2.6 × 102 | 5.1 × 102 |
| Gymnodinium sp. | 8.9 × 104 | 2.6 × 102 | 8.7 × 105 |
| Gymnodinium instriatum | 3.7 × 106 | 2.6 × 102 | 3.1 × 107 |
| Heterocapsaro tundata | 6.6 × 105 | 1.1 × 105 | 3.9 × 106 |
| Heterocapsa triquetra | 1.0 × 104 | 2.6 × 102 | 1.1 × 105 |
| Polykrikos kofoidii | 5.9 × 103 | 1.0 × 103 | 4.5 × 104 |
| Prorocentrum micans | 9.3 × 102 | 2.6 × 102 | 5.9 × 103 |
| Prorocentrum minimum | 2.2 × 104 | 2.6 × 102 | 4.3 × 105 |
| Protoperidinium sp. | 7.5 × 102 | 2.6 × 102 | 1.8 × 103 |
| Scrippsiella trochoidea | 7.3 × 102 | 2.6 × 102 | 2.3 × 103 |
| Cryptomonads | |||
| Cryptomonas sp. | 1.5 × 107 | 5.4 × 105 | 7.6 × 107 |
| Cyanobacteria | |||
| Lyngbya sp. | 5.3 × 105 | 5.1 × 102 | 2.3 × 106 |
| Chlorophytes | |||
| Ankistrodesmus falcatus | 8.7 × 103 | 2.6 × 102 | 1.1 × 105 |
| Chlamydomonas sp. | 3.6 × 105 | 1.1 × 105 | 8.7 × 105 |
| Euglenoids | |||
| Euglena sp. | 7.6 × 104 | 2.6 × 102 | 4.3 × 105 |
| Eutreptia lanowii | 2.0 × 103 | 5.1 × 102 | 5.6 × 103 |
| Prasinophytes | |||
| Pyramimonas sp. | 7.2 × 104 | 2.6 × 102 | 3.2 × 105 |
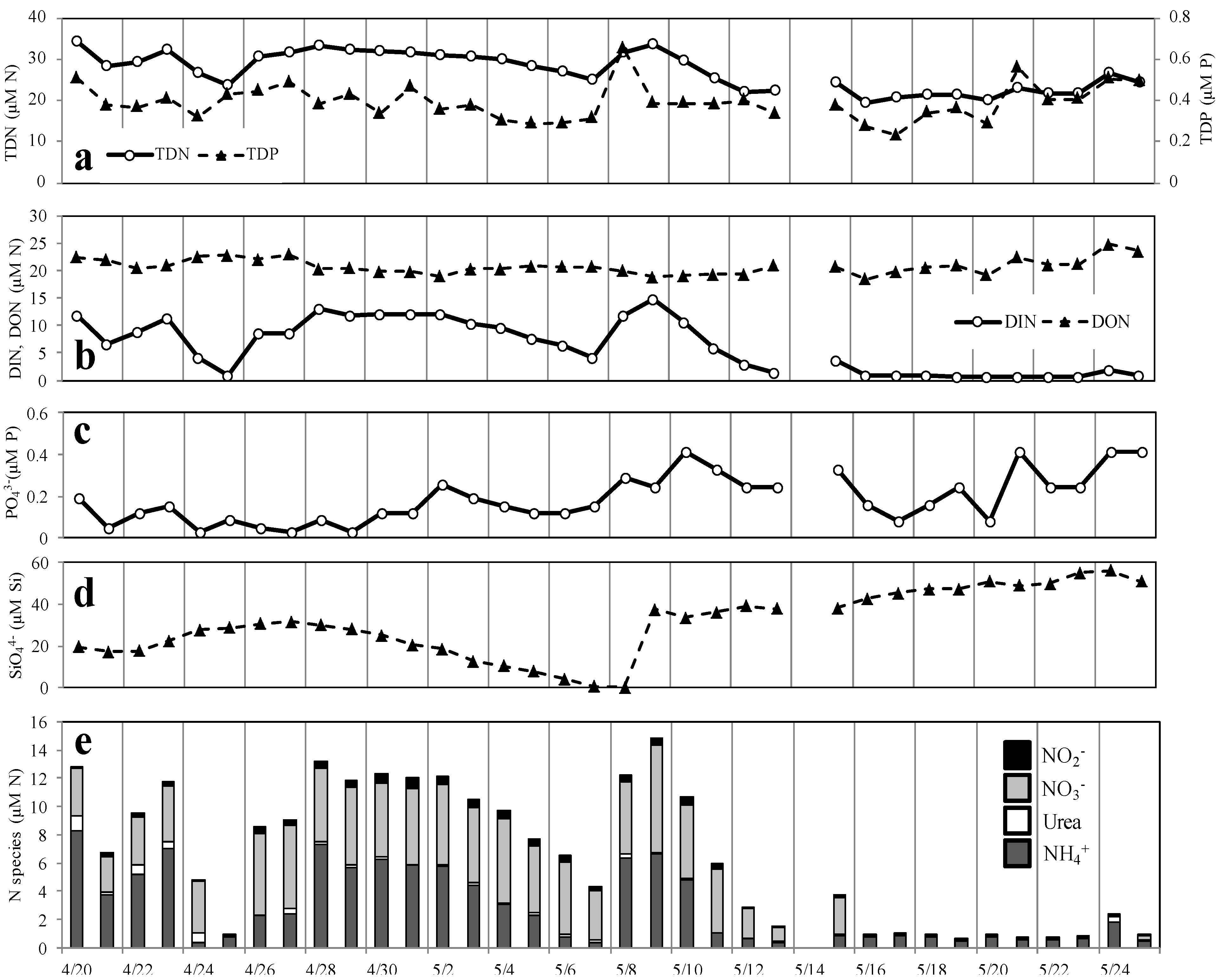
3.3. Nutrient Concentrations

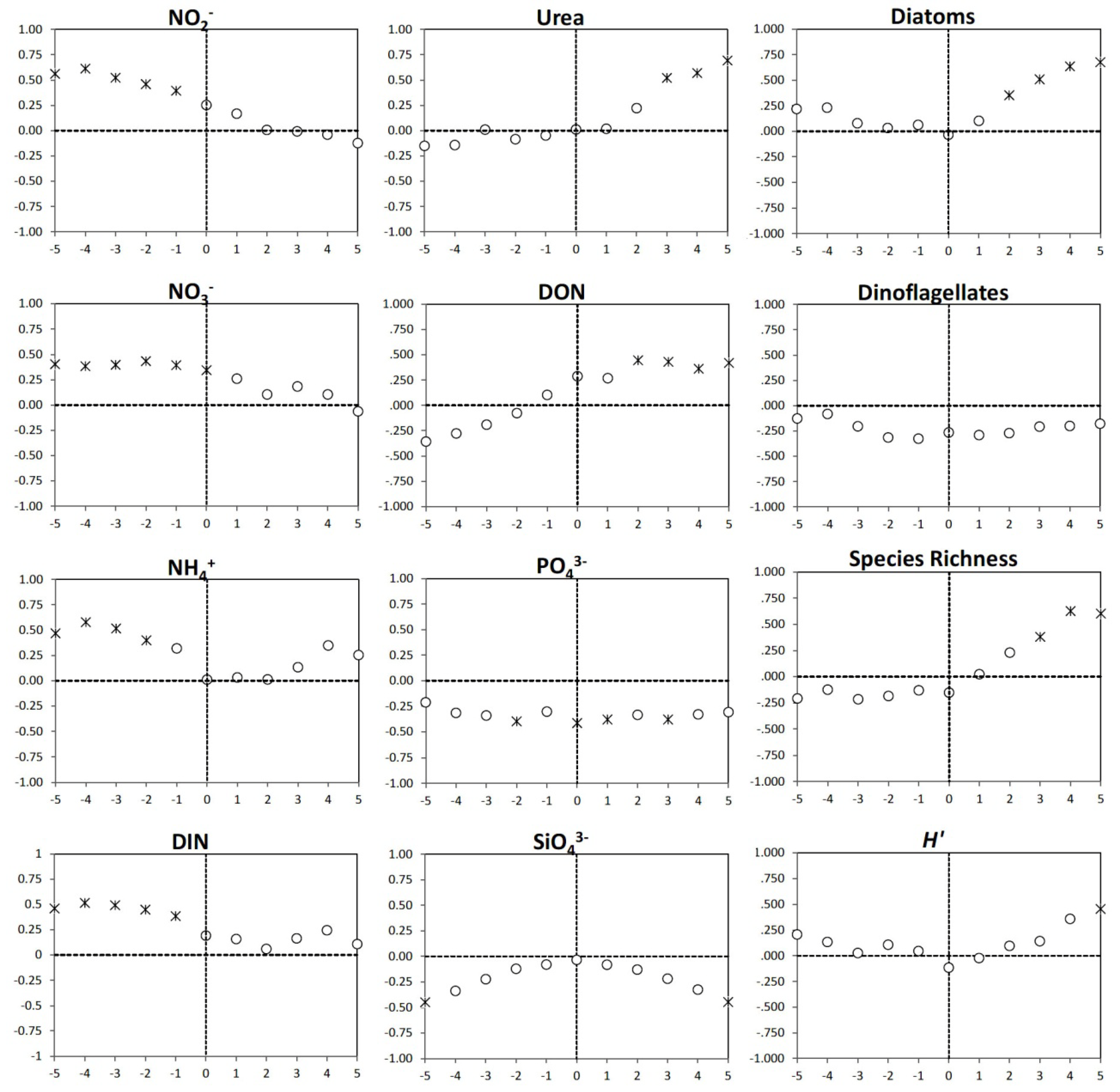
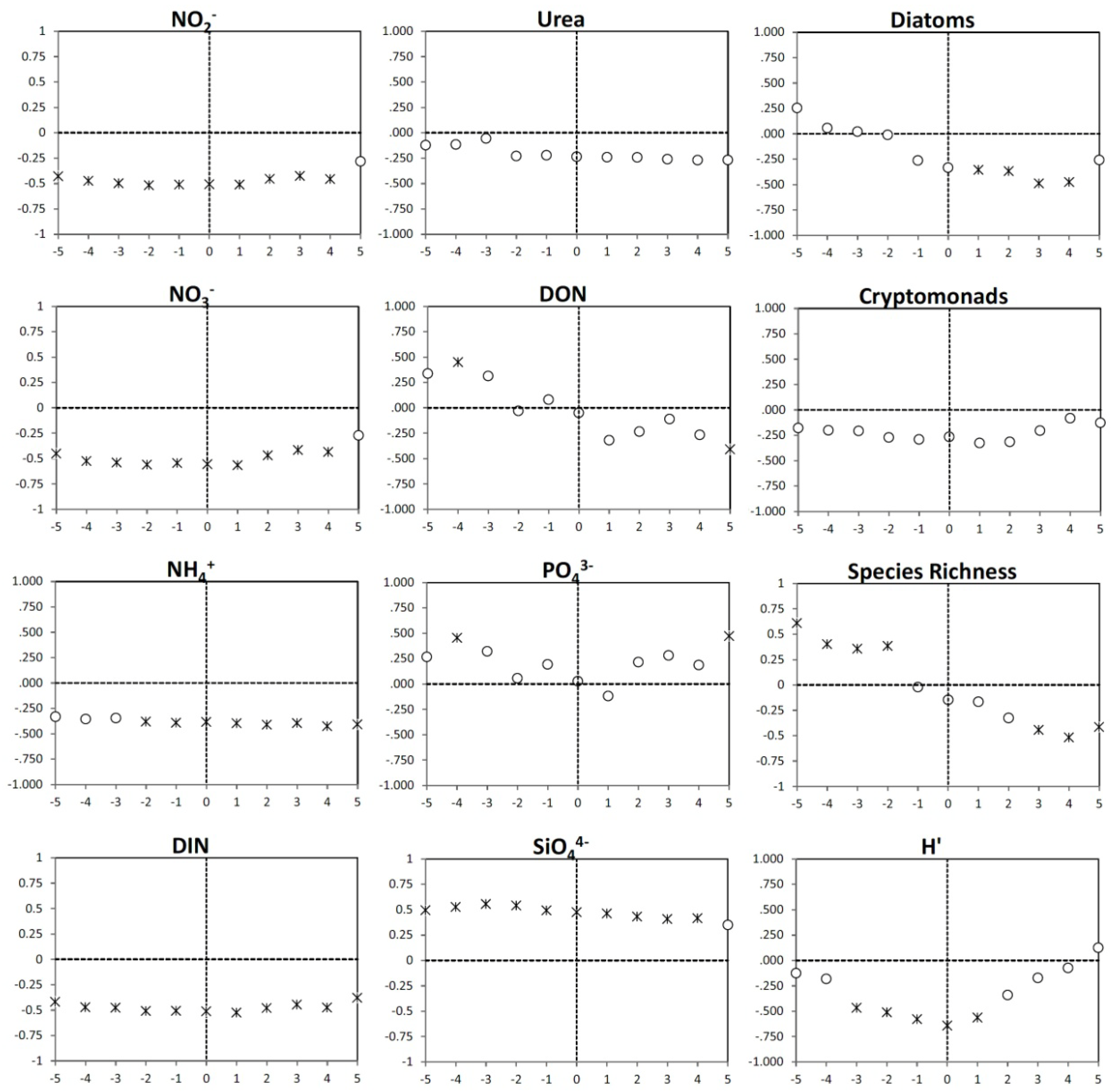
3.4. Time Lag Correlations

3.5. Discussion
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Hutchinson, G.E. The paradox of the plankton. Am. Nat. 1961, 95, 137–145. [Google Scholar]
- Grover, J.P.; Chrzanowski, T.H. Limiting resources, disturbance, and diversity in phytoplankton communities. Ecol. Monogr. 2004, 74, 533–551. [Google Scholar] [CrossRef]
- Cloern, J.E.; Dufford, R. Phytoplankton community ecology: Principles applied in San Francisco Bay. Mar. Ecol. Prog. Ser. 2005, 285, 11–28. [Google Scholar] [CrossRef]
- Spatharis, S.; Tsirtsis, G.; Danielidis, D.B.; Chi, T.D.; Mouillot, D. Effects of pulsed nutrient inputs on phytoplankton assemblage structure and blooms in an enclosed coastal area. Estuar. Coast. Shelf Sci. 2007, 73, 807–815. [Google Scholar] [CrossRef]
- Hubertz, E.D.; Cahoon, L.B. Short-term variability of water quality parameters in two shallow estuaries of North Carolina. Estuaries 1999, 22, 814–823. [Google Scholar] [CrossRef]
- Filippino, K.C.; Egerton, T.A.; Morse, R.E.; Hunley, W.S.; Mulholland, M.R. Nutrient dynamics in the tidal James River before, during, and after Hurricane Irene and Tropical Storm Lee. Estuar. Coast. Shelf Sci. 2013. submitted for publication. [Google Scholar]
- Morse, R.E.; Mulholland, M.R.; Egerton, T.A.; Marshall, H.G. Daily variability in phytoplankton abundance and nutrient concentrations in a tidally dominated eutrophic estuary. Mar. Ecol. Prog. Ser. 2013. submitted for publication. [Google Scholar]
- Morse, R.E.; Mulholland, M.R.; Hunley, W.S.; Fentress, S.; Wiggins, M.; Blanco-Garcia, J.L. Controls on the initiation and development of blooms of the dinoflagellate Cochlodinium polykrikoides Margalef in lower Chesapeake Bay and its tributaries. Harmful Algae 2013, 28, 71–82. [Google Scholar] [CrossRef]
- McCormick, P.V.; Cairns, J., Jr. Algae as indicators of environmental change. J. Appl. Phycol. 1994, 6, 509–526. [Google Scholar] [CrossRef]
- Buchanan, C.; Lacouture, R.V.; Marshall, H.G.; Olson, M.; Johnson, J.M. Phytoplankton reference communities for Chesapeake Bay and its tidal tributaries. Estuaries 2005, 28, 138–159. [Google Scholar] [CrossRef]
- Paerl, H.W.; Valdes-Weaver, L.M.; Joyner, A.R.; Winkelmann, V. Phytoplankton indicators of ecological change in the eutrophying Pamlico Sound system, North Carolina. Ecol. Appl. 2007, 17, S88–S101. [Google Scholar] [CrossRef]
- Litaker, W.; Duke, C.S.; Kenny, B.E.; Ramus, J. Short-term environmental variability and phytoplankton abundance in a shallow tidal estuary. II. Spring and fall. Mar. Ecol. Prog. Ser. 1993, 94, 141–154. [Google Scholar] [CrossRef]
- Malone, T.C.; Conley, D.J.; Fisher, T.R.; Glibert, P.M.; Harding, L.W.; Sellner, K.G. Scales of nutrient-limited phytoplankton productivity in Chesapeake Bay. Estuaries 1996, 19, 371–385. [Google Scholar] [CrossRef]
- Paerl, H.W.; Rossignol, K.L.; Hall, S.N.; Peierls, B.L.; Wetz, M.S. Phytoplankton community indicators of short-and long-term ecological change in the anthropogenically and climatically impacted Neuse River Estuary, North Carolina, USA. Estuar. Coasts 2010, 33, 485–497. [Google Scholar] [CrossRef]
- Duarte, P.; Macedo, M.F.; da Fonseca, L.C. The relationship between phytoplankton diversity and community function in a coastal lagoon. Hydrobiologia 2006, 555, 3–18. [Google Scholar] [CrossRef]
- Jouenne, F.; Lefebvre, S.; Véron, B.; Lagadeuc, Y. Phytoplankton community structure and primary production in small intertidal estuarine-bay ecosystem (eastern English Channel, France). Mar. Biol. 2007, 151, 805–825. [Google Scholar] [CrossRef]
- Glibert, P.M.; Magnien, R.; Lomas, M.W.; Alexander, J.; Tan, C.; Haramoto, E.; Trice, M.; Kana, T.M. Harmful algal blooms in the Chesapeake and coastal bays of Maryland, USA: Comparison of 1997, 1998, and 1999 events. Estuaries 2001, 24, 875–883. [Google Scholar] [CrossRef]
- Gobler, C.J.; Burson, A.; Koch, F.; Tang, Y.; Mulholland, M.R. The role of nitrogenous nutrients in the occurrence of harmful algal blooms caused by Cochlodinium polykrikoides in New York estuaries (USA). Harmful Algae 2012, 17, 64–74. [Google Scholar] [CrossRef]
- Anderson, D.M.; Glibert, P.M.; Burkholder, J.M. Harmful algal blooms and eutrophication: Nutrient sources, composition, and consequences. Estuaries 2002, 25, 704–726. [Google Scholar] [CrossRef]
- Heisler, J.; Glibert, P.M.; Burkholder, J.M.; Anderson, D.M.; Cochlan, W.; Dennison, W.C.; Dortch, Q.; Gobler, C.J.; Heil, C.A.; Humphries, E.; et al. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef]
- Morse, R.E.; Shen, J.; Blanco-Garcia, J.L.; Hunley, W.S.; Fentress, S.; Wiggins, M.; Mulholland, M.R. Environmental and physical controls on the formation and transport of blooms of the dinoflagellate Cochlodinium polykrikoides Margalef in the lower Chesapeake Bay and its tributaries. Estuar. Coasts 2011, 34, 1006–1025. [Google Scholar] [CrossRef]
- Mulholland, M.R.; Morse, R.E.; Boneillo, G.E.; Bernhardt, P.W.; Filippino, K.C.; Procise, L.A.; Blanco-Garcia, J.L.; Marshall, H.G.; Egerton, T.A.; Hunley, W.S.; et al. Understanding causes and impacts of the dinoflagellate, Cochlodinium polykrikoides, blooms in the Chesapeake Bay. Estuar. Coasts 2009, 32, 734–747. [Google Scholar] [CrossRef]
- Mulholland, M.R.; Boneillo, G.E.; Bernhardt, P.W.; Minor, E.C. Comparison of nutrient and microbial dynamics over a seasonal cycle in a mid-Atlantic coastal lagoon prone to Aureococcus anophagefferens (brown tide) blooms. Estuar. Coasts 2009, 32, 1176–1194. [Google Scholar] [CrossRef]
- Glibert, P.M.; Kelly, V.; Alexander, J.; Codispoti, L.A.; Boicourt, W.C.; Trice, T.M.; Michael, B. In situ nutrient monitoring: A tool for capturing nutrient variability and the antecedent conditions that support algal blooms. Harmful Algae 2008, 8, 175–181. [Google Scholar] [CrossRef]
- Mitchell-Innes, B.A.; Walker, D.R. Short-term variability during an anchor station study in the southern Benguela upwelling system: Phytoplankton production and biomass in relation to specie changes. Prog. Oceanogr. 1991, 28, 65–89. [Google Scholar] [CrossRef]
- Blair, C.H.; Cox, J.H.; Kuo, C.Y. Investigation of Flushing Time in the Lafayette River, Norfolk, Virginia; School of Engineering Technical Report No. 76-C4; Old Dominion University: Norfolk, VA, USA, 1976. [Google Scholar]
- White, E.G. A Physical Hydrographic Study of the Lafayette River. M.S. Thesis, Old Dominion University, Norfolk, VA, USA, 1972. [Google Scholar]
- Owen, D.W.; Rogers, L.M.; Peoples, M.H. Shoreline Situation Report; Cities of Chesapeake, Norfolk, and Portsmouth; Special Report in Applied Marine Science and Oceanic Engineering Number 136; Virginia Institute of Marine Science: Gloucester Point, VA, USA, 1976. [Google Scholar]
- Berman, M.; Berquist, H.; Hershner, C.; Killeen, S.; Rudnicky, T.; Schatt, D.; Weiss, D.; Woods, H. City of Norfolk Shoreline Situation Report; Special Report in Applied Marine Science and Ocean Engineering Number 378; Virginia Institute of Marine Science: Gloucester Point, VA, USA, 2002. [Google Scholar]
- Purcell, T.W. Phytoplankton Succession in the Lafayette River Estuary, Norfolk, Virginia. M.S. Thesis, Old Dominion University, Norfolk, VA, USA, 1973. [Google Scholar]
- Marshall, H.G. Plankton in James River Estuary, Virginia III. Phytoplankton in the Lafayette and Elizabeth Rivers (Western and Eastern Branches). Castanea 1968, 33, 255–258. [Google Scholar]
- Kalenak, L.A. Factors Effecting Phytoplankton Assemblages in the Lafayette River Estuary. M.S. Thesis, Old Dominion University, Norfolk, Virginia, USA, 1982. [Google Scholar]
- Egerton, T.A.; Marshall, H.G.; Hunley, W.S. Integration of Microscopy and Underway Chlorophyll Mapping for Monitoring Algal Bloom Development. Oceans 2012, 2012. [Google Scholar] [CrossRef]
- Welschmeyer, N.A. Fluorometric analysis of chlorophyll a in the presence of chlorophyll b and pheopigments. Limnol. Oceanogr. 1994, 39, 1985–1992. [Google Scholar] [CrossRef]
- Solarzano, L. Determination of ammonia in natural waters by the phenol hypochlorite method. Limnol. Oceanogr. 1969, 14, 16–23. [Google Scholar] [CrossRef]
- Marshall, H.G.; Alden, R.W. A comparison of phytoplankton assemblages and environmental relationships in three estuarine rivers of the lower Chesapeake Bay. Estuaries 1990, 13, 287–300. [Google Scholar] [CrossRef]
- Affronti, L.F.; Marshall, H.G. Using frequency of dividing cells in estimating autotrophic picoplankton growth and productivity in the Chesapeake Bay. Hydrobiologia 1994, 284, 193–203. [Google Scholar] [CrossRef]
- Smayda, T.J. From Phytoplankters to Biomass. In Phytoplankton Manual; UNESCO: Paris, France, 1978; pp. 273–279. [Google Scholar]
- Tang, Y.Z.; Egerton, T.A.; Kong, L.; Marshall, H.G. Morphological variation and phylogenetic analysis of the dinoflagellate Gymnodinium aureolum from a tributary of Chesapeake Bay. J. Eukaryot. Microbiol. 2008, 55, 91–99. [Google Scholar] [CrossRef]
- Shannon, C.; Weaver, W. The Mathematical Theory Information; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Waide, R.B.; Willig, M.R.; Steiner, C.F.; Mittelbach, G.; Gough, L.; Dodson, S.I.; Juday, P.; Parmenter, R. The relationship between productivity and species richness. Annu. Rev. Ecol. Syst. 1999, 30, 257–300. [Google Scholar]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Witman, J.D.; Cusson, M.; Archambault, P.; Pershing, A.J.; Mieszkowska, N. The relation between productivity and species diversity in temperate-arctic marine ecosystems. Ecology 2008, 89, S66–S80. [Google Scholar] [CrossRef]
- Klaveness, D. Ecology of the Cryptomonadida: A First Review. In Growth and Reproductive Strategies of Freshwater Phytoplankton; Sandgren, C.D., Ed.; Cambridge University Press: Cambridge, UK, 1988; pp. 105–134. [Google Scholar]
- Menezes, M.; Novarino, G. How diverse are planktonic cryptomonads in Brazil? Advantages and difficulties of a taxonomic-biogeographical approach. Hydrobiologia 2003, 502, 297–306. [Google Scholar] [CrossRef]
- Steidinger, K.A.; Tangen, K. Dinoflagellates. In Identifying Marine Phytoplankton; Tomas, C.R., Ed.; Academic Press: San Diego, CA, USA, 1997; pp. 387–598. [Google Scholar]
- Coats, D.W.; Park, M.G. Parasitism of photosynthetic dinoflagellates by three strains of Amoebophrya (Dinophyta): Parasite survival, infectivity, generation time, and host specificity. J. Phycol. 2002, 38, 520–528. [Google Scholar]
- Conley, D.J.; Malone, T.C. Annual cycle of dissolved silicate in Chesapeake Bay: Implications for the production and fate of phytoplankton biomass. Mar. Ecol. Prog. Ser. 1992, 81, 121–128. [Google Scholar] [CrossRef]
- Marshall, H.G.; Lane, M.F.; Nesius, K.K.; Burchardt, L. Assessment and significance of phytoplankton species composition within Chesapeake Bay and Virginia tributaries through a long-term monitoring program. Environ. Monit. Assess. 2009, 150, 143–155. [Google Scholar] [CrossRef]
- Williams, M.R.; Filoso, S.; Longstaff, B.J.; Dennison, W.C. Long-term trends of water quality and biotic metrics in Chesapeake Bay: 1986 to 2008. Estuar. Coasts 2010, 33, 1279–1299. [Google Scholar] [CrossRef]
- Roberts, D.A.; Poore, A.G.; Johnston, E.L. MBACI sampling of an episodic disturbance: Storm water effects on algal epifauna. Mar. Environ. Res. 2007, 64, 514–523. [Google Scholar] [CrossRef]
- Najjar, R.G.; Pyke, C.R.; Adams, M.B.; Breitburg, D.; Hershner, C.; Kemp, M.; Howarth, R.; Mulhollad, M.R.; Paolisso, M.; Secor, D.; et al. Potential climate-change impacts on the Chesapeake Bay. Estuar. Coast. Shelf Sci. 2010, 86, 1–20. [Google Scholar] [CrossRef]
- Orth, R.J.; Williams, M.R.; Marion, S.R.; Wilcox, D.J.; Carruthers, T.J.; Moore, K.A.; Kemp, W.M.; Dennison, W.C.; Rybicki, N.; Bergstrom, P.; et al. Long-term trends in submersed aquatic vegetation (SAV) in Chesapeake Bay, USA, related to water quality. Estuar. Coasts 2010, 33, 1144–1163. [Google Scholar] [CrossRef]
- Cho, K.H.; Wang, H.V.; Shen, J.; Valle-Levinson, A.; Teng, Y.C. A modeling study on the response of Chesapeake Bay to hurricane events of Floyd and Isabel. Ocean Model. 2012, 49, 22–46. [Google Scholar]
- Marshall, H.G.; Lacouture, R.V.; Buchanan, C.; Johnson, J.M. Phytoplankton assemblages associated with water quality and salinity regions in Chesapeake Bay, USA. Estuar. Coast. Shelf Sci. 2006, 69, 10–18. [Google Scholar] [CrossRef]
- Mallin, M.A.; Paerl, H.W.; Rudek, J. Seasonal phytoplankton composition, productivity and biomass in the Neuse River estuary, North Carolina. Estuar. Coast. Shelf Sci. 1991, 32, 609–623. [Google Scholar] [CrossRef]
- Weisse, T.; Kirchhoff, B. Feeding of the heterotrophic freshwater dinoflagellate. Peridiniopsis berolinense on cryptophytes: Analysis by flow cytometry and electronic particle counting. Aquat. Microb. Ecol. 1997, 12, 153–164. [Google Scholar] [CrossRef]
- Adolf, J.E.; Bachvaroff, T.; Place, A.R. Can cryptophyte abundance trigger toxic Karlodinium veneficum blooms in eutrophic estuaries? Harmful Algae 2008, 8, 119–128. [Google Scholar] [CrossRef]
- Jimenéz, R. Ecological Factors Related to Gyrodinium instriatum Bloom in the Inner Estuary of the Gulf of Guayaquil. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 257–262. [Google Scholar]
- Kim, H.G.; Park, J.S.; Lee, S.G.; An, K.H. Population Cell Volume and Carbon Content in Monospecific Dinoflagellate Blooms. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 769–773. [Google Scholar]
- Uchida, T.; Kamiyama, T.; Matsuyama, Y. Predation by a photosynthetic dinoflagellate Gyrodinium instriatum on loricated ciliates. J. Plankton Res. 1997, 19, 603–608. [Google Scholar] [CrossRef]
- Li, A.; Stoecker, D.K.; Coats, D.W. Use of the ‘food vacuole content’ method to estimate grazing by the mixotrophic dinoflagellate Gyrodinium galatheanum on cryptophytes. J. Plankton Res. 2001, 23, 303–318. [Google Scholar] [CrossRef]
- Hansen, P.J. The role of photosynthesis and food uptake for the growth of marine mixotrophic dinoflagellates1. J. Eukaryot. Microbiol. 2001, 58, 203–214. [Google Scholar] [CrossRef]
- Shikata, T.; Nagasoe, S.; Matsubara, T.; Yamasaki, Y.; Shimasaki, Y.; Oshima, Y.; Uchida, T.; Jenkinson, I.R.; Honjo, T. Encystment and excystment of Gyrodinium instriatum Freudenthal et Lee. J. Oceanogr. 2008, 64, 355–365. [Google Scholar] [CrossRef]
- Nagasoe, S.; Kim, D.I.; Shimasaki, Y.; Oshima, Y.; Yamaguchi, M.; Honjo, T. Effects of temperature, salinity and irradiance on the growth of the red tide dinoflagellate Gyrodinium instriatum Freudenthal et Lee. Harmful Algae 2006, 5, 20–25. [Google Scholar] [CrossRef]
- Malmquist, D. VIMS researchers monitor harmful algal bloom. Available online: http://www.vims.edu/newsandevents/topstories/alexandrium_bloom.php (accessed on 20 April 2013).
- Marshall, H.G. Seasonal phytoplankton composition in the lower Chesapeake Bay and Old Plantation Creek, Cape Charles, Virginia. Estuaries 1980, 3, 207–216. [Google Scholar] [CrossRef]
- Marshall, H.G.; Lacouture, R. Seasonal patterns of growth and composition of phytoplankton in the lower Chesapeake Bay and vicinity. Estuar. Coast. Shelf Sci. 1986, 23, 115–130. [Google Scholar] [CrossRef]
- Adolf, J.E.; Yeager, C.L.; Miller, W.D.; Mallonee, M.E.; Harding, L.W. Environmental forcing of phytoplankton floral composition, biomass, and primary productivity in Chesapeake Bay, USA. Estuar. Coast. Shelf Sci. 2006, 67, 108–122. [Google Scholar] [CrossRef]
- Jordan, T.E.; Correll, D.L.; Weller, D.E. Relating nutrient discharges from watersheds to land use and stream flow variability. Water Resour. Res. 1997, 33, 2579–2590. [Google Scholar] [CrossRef]
- Langland, M.J.; Phillips, S.W.; Raffensperger, J.P.; Moyer, D.L. Changes in Stream Flow and Water Quality in Selected Nontidal Sites in the Chesapeake Bay Basin, 1985–2003; U.S. Geological Survey Scientific Investigations Report 2004–5259; U.S. Geological Survey: Baltimore, MD, USA, 2004; p. 50. [Google Scholar]
- Vought, L.B.M.; Pinay, G.; Fuglsang, A.; Ruffinoni, C. Structure and function of buffer strips from a water quality perspective in agricultural landscapes. Landsc. Urban Plan. 1995, 31, 323–331. [Google Scholar] [CrossRef]
- Laws, E.A.; Ziemann, D.; Schulman, D. Coastal water quality in Hawaii: The importance of buffer zones and dilution. Mar. Environ. Res. 1999, 48, 1–21. [Google Scholar] [CrossRef]
- Syversen, N.; Haarstad, K. Retention of pesticides and nutrients in a vegetated buffer root zone compared to soil with low biological activity. Int. J. Environ. Anal. Chem. 2005, 85, 1175–1187. [Google Scholar] [CrossRef]
- Nichols, F.H.; Cloern, J.E.; Luoma, S.N.; Peterson, D.H. The modification of an estuary. Science 1986, 231, 567–573. [Google Scholar]
- Sciandra, A.; Lazzara, L.; Claustre, H.; Babin, M. Responses of growth rate, pigment composition and optical properties of Cryptomonas sp. to light and nitrogen stresses. Mar. Ecol. Prog. Ser. 2000, 201, 107–120. [Google Scholar] [CrossRef]
- Cloern, J.E. Effects of light intensity and temperature on Cryptomonas ovata (Cryptophyceae) growth and nutrient uptake rates. J. Phycol. 1977, 13, 389–395. [Google Scholar]
- Bradley, P.B.; Lomas, M.W.; Bronk, D.A. Inorganic and organic nitrogen use by phytoplankton along Chesapeake Bay, measured using a flow cytometric sorting approach. Estuar. Coasts 2010, 33, 971–984. [Google Scholar] [CrossRef]
- Kemp, W.M.; Boynton, W.R.; Adolf, J.E.; Boesch, D.F.; Boicourt, W.C.; Brush, G.; Cornwell, J.C.; Fisher, T.R.; Glibert, P.M.; Hagy, J.D.; et al. Eutrophication of Chesapeake Bay: Historical trends and ecological interactions. Mar. Ecol. Prog. Ser. 2005, 303, 1–29. [Google Scholar] [CrossRef]
- Furnas, M.J. In situ growth rates of marine phytoplankton: Approaches to measurement, community and species growth rates. J. Plankton Res. 1990, 12, 1117–1151. [Google Scholar] [CrossRef]
- Anderson, D.M.; Wall, D. Potential importance of benthic cysts of Gonyaulax tamarensis and G. excavates in initiating toxic dinoflagellate blooms. J. Phycol. 1978, 20, 224–234. [Google Scholar] [CrossRef]
- Anderson, D.M.; Rengefors, K. Community assembly and seasonal succession of marine dinoflagellates in a temperate estuary: The importance of life cycle events. Limnol. Oceanogr. 2006, 51, 860–873. [Google Scholar] [CrossRef]
- Seaborn, D.W.; Marshall, H.G. Dinoflagellate cysts within sediment collections from the southern Chesapeake Bay and tidal regions of the James, York, and Rappahannock rivers. Va. J. Sci. 2008, 59, 135–141. [Google Scholar]
- Tomas, C.R.; Smayda, T.J. Red tide blooms of Cochlodinium polykrikoides in a coastal cove. Harmful Algae 2008, 7, 308–317. [Google Scholar] [CrossRef]
- Sunda, W.G.; Graneli, E.; Gobler, C.J. Positive feedback and the development and persistence of ecosystem disruptive algal blooms. J. Phycol. 2006, 42, 963–974. [Google Scholar] [CrossRef]
- Berman, T.; Bronk, D.A. Dissolved organic nitrogen: A dynamic participant in aquatic ecosystems. Aquat. Microb. Ecol. 2003, 31, 279–305. [Google Scholar] [CrossRef]
- Li, A.; Stoecker, D.K.; Coats, D.W. Spatial and temporal aspects of Gyrodinium galatheanum in Chesapeake Bay: Distribution and mixotrophy. J. Plankton Res. 2000, 22, 2105–2124. [Google Scholar] [CrossRef]
- Tyler, M.A.; Seliger, H.H. Annual subsurface transport of a red tide dinoflagellate to its bloom area: Water circulation patterns and organism distributions in the Chesapeake Bay. Limnol. Oceanogr. 1978, 23, 227–246. [Google Scholar] [CrossRef]
- Nehring, S. Mechanisms for Recurrent Nuisance Algal Blooms in Coastal Zones: Resting Cyst Formation as Life-Strategy of Dinoflagellates. In Proceedings of the International Coastal Congress, ICC-Kiel 92: Interdisciplinary Discussion of Coastal Research and Coastal Management Issues and Problems; Sterr, H., Hofstade, J., Plag, H.-P., Eds.; P. Lang: Frankfurt, Germany, 1993; pp. 454–467. [Google Scholar]
- Sellner, K.G.; Sellner, S.G.; Lacouture, R.V.; Magnien, R.E. Excessive nutrients select for dinoflagellates in the stratified Patapsco River estuary: Margalef reigns. Mar. Ecol. Prog. Ser. 2001, 220, 93–102. [Google Scholar] [CrossRef]
- Peierls, B.L.; Hall, N.S.; Paerl, H.W. Non-monotonic responses of phytoplankton biomass accumulation to hydrologic variability: A comparison of two coastal plain North Carolina estuaries. Estuar. Coasts 2012, 35, 1376–1392. [Google Scholar] [CrossRef]
- Leibold, M.A. Biodiversity and nutrient enrichment in pond plankton communities. Evolut. Ecol. Res. 1999, 1, 73–95. [Google Scholar]
- Irigoien, X.; Huisman, J.; Harris, R.P. Global biodiversity patterns of marine phytoplankton and zooplankton. Nature 2004, 429, 863–867. [Google Scholar] [CrossRef]
- Chesapeake Bay Program Plankton Database. Available online: http://www.chesapeakebay.net/data/downloads/baywide_cbp_plankton_database (accessed on 20 April 2013).
- Glibert, P.M.; Alexander, J.; Meritt, D.W.; North, E.W.; Stoecker, D.K. Harmful algae pose additional challenges for oyster restoration: Impacts of the harmful algae Karlodinium veneficum, and Prorocentrum minimum on early life stages of the oysters Crassostrea virginica and Crassostrea ariakensis. J. Shellfish Res. 2007, 26, 919–925. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Egerton, T.A.; Morse, R.E.; Marshall, H.G.; Mulholland, M.R. Emergence of Algal Blooms: The Effects of Short-Term Variability in Water Quality on Phytoplankton Abundance, Diversity, and Community Composition in a Tidal Estuary. Microorganisms 2014, 2, 33-57. https://doi.org/10.3390/microorganisms2010033
Egerton TA, Morse RE, Marshall HG, Mulholland MR. Emergence of Algal Blooms: The Effects of Short-Term Variability in Water Quality on Phytoplankton Abundance, Diversity, and Community Composition in a Tidal Estuary. Microorganisms. 2014; 2(1):33-57. https://doi.org/10.3390/microorganisms2010033
Chicago/Turabian StyleEgerton, Todd A., Ryan E. Morse, Harold G. Marshall, and Margaret R. Mulholland. 2014. "Emergence of Algal Blooms: The Effects of Short-Term Variability in Water Quality on Phytoplankton Abundance, Diversity, and Community Composition in a Tidal Estuary" Microorganisms 2, no. 1: 33-57. https://doi.org/10.3390/microorganisms2010033