
Nest-Gallery Development and Caste Composition of Isolated Foraging Groups of the Drywood Termite, Incisitermes minor (Isoptera: Kalotermitidae)

 ,
,
Abstract
:1. Introduction
2. Materials and Method
2.1. Wood Specimen and Experimental Setup
2.2. Monitoring
2.3. Analysis
2.3.1. X-Ray CT Scanning and Data Analysis
2.3.2. Colony Examination
3. Results
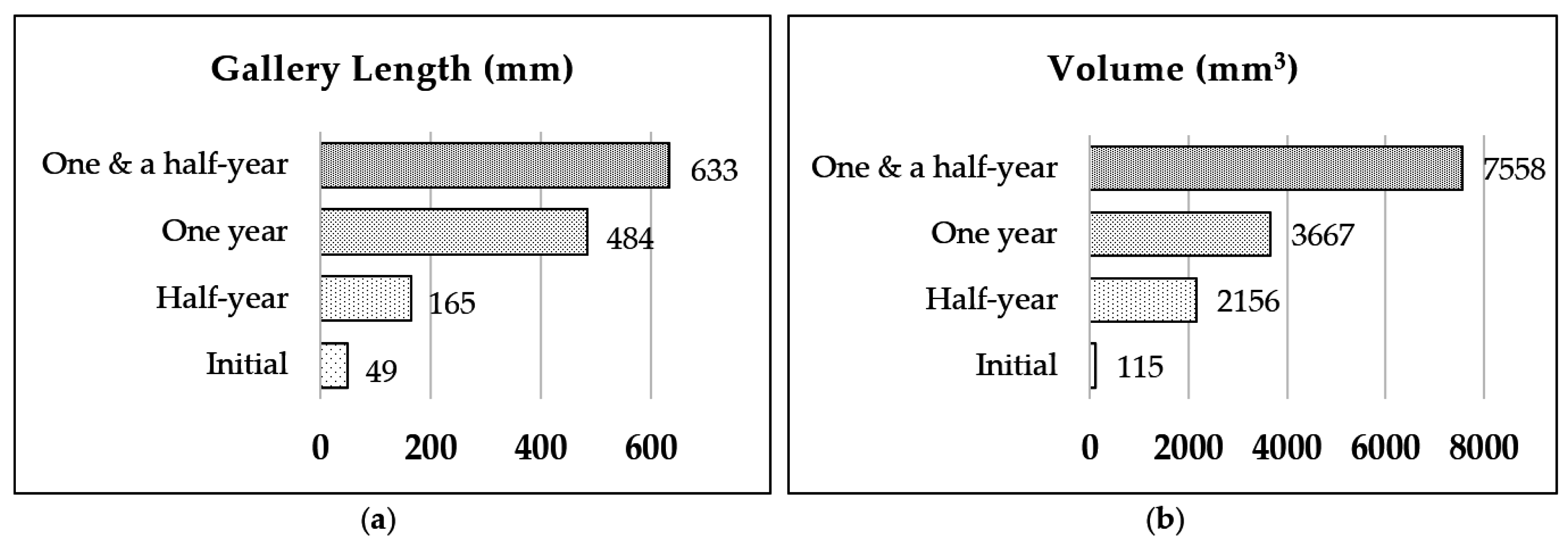
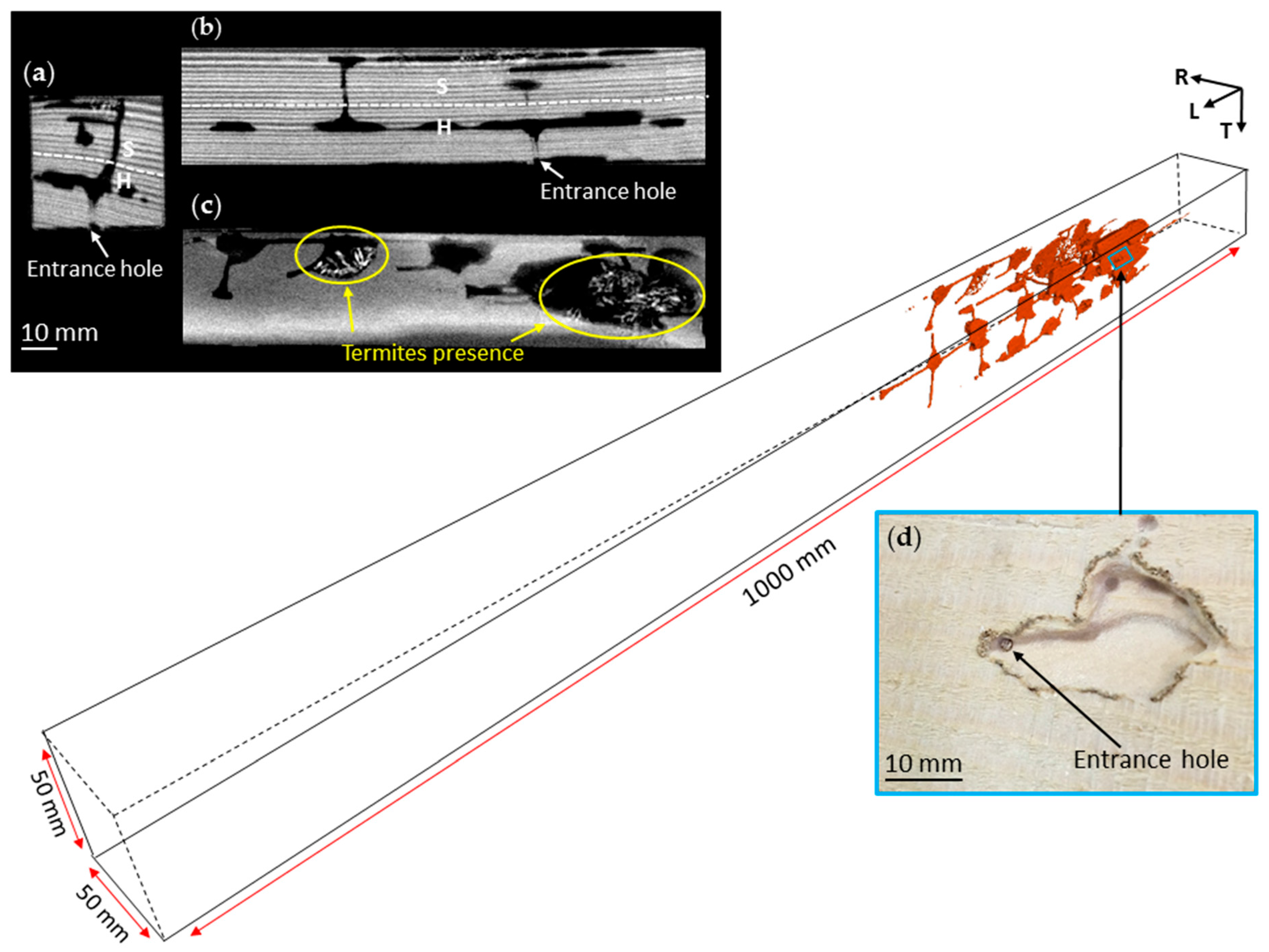
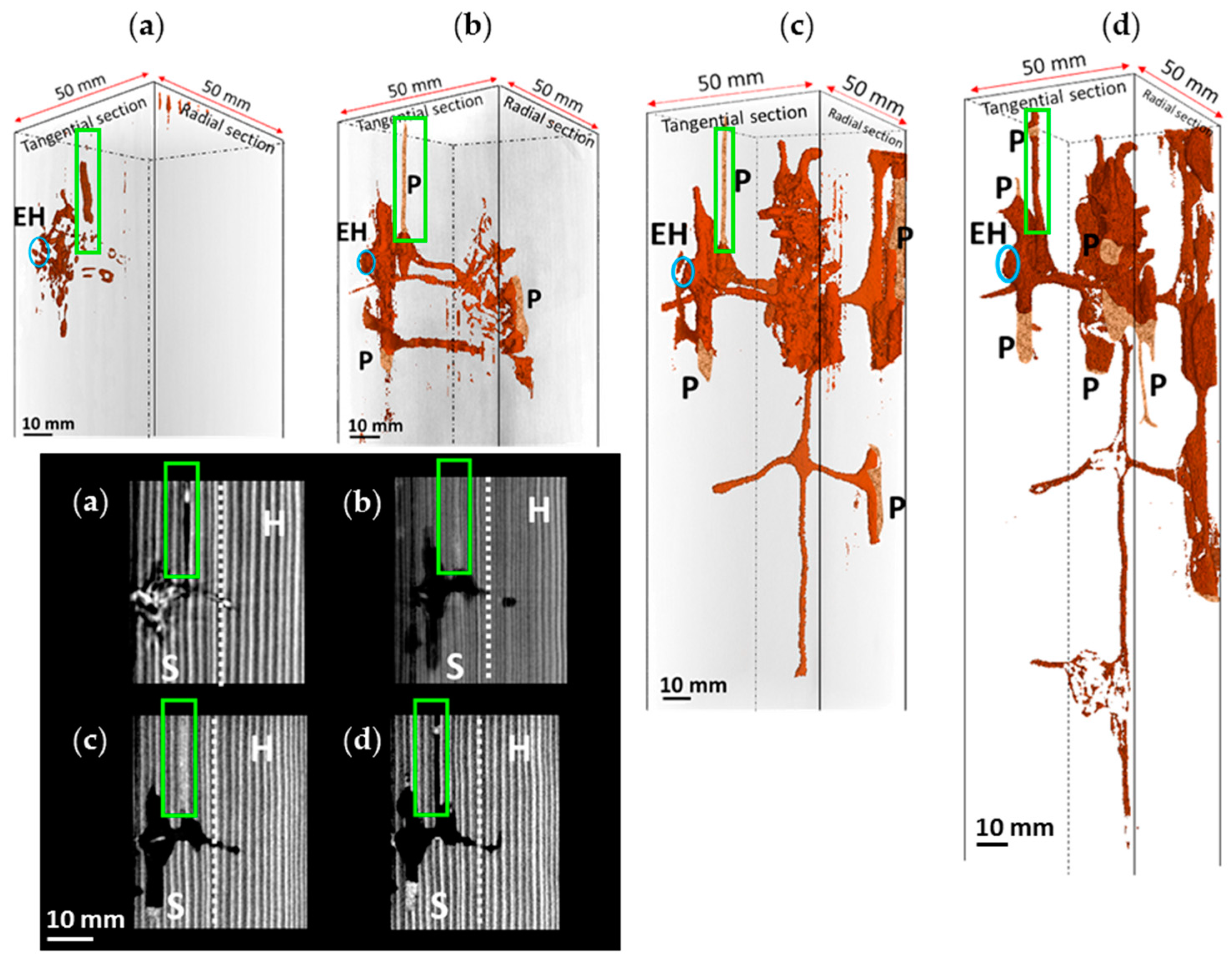
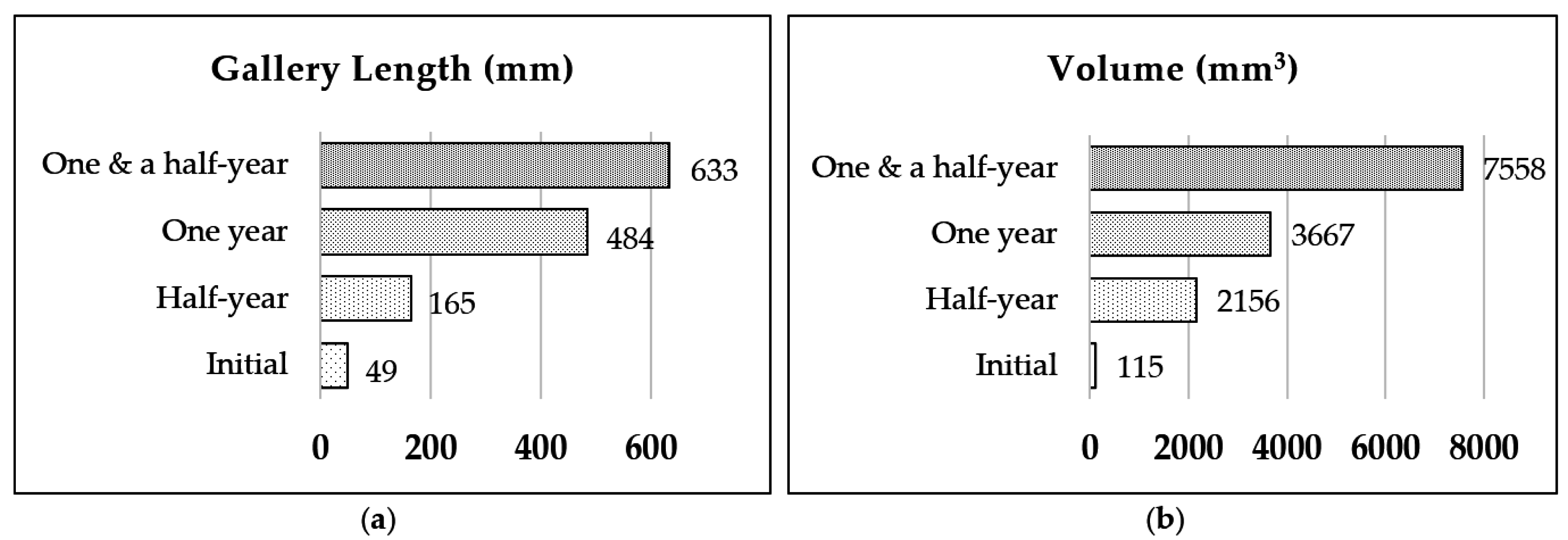
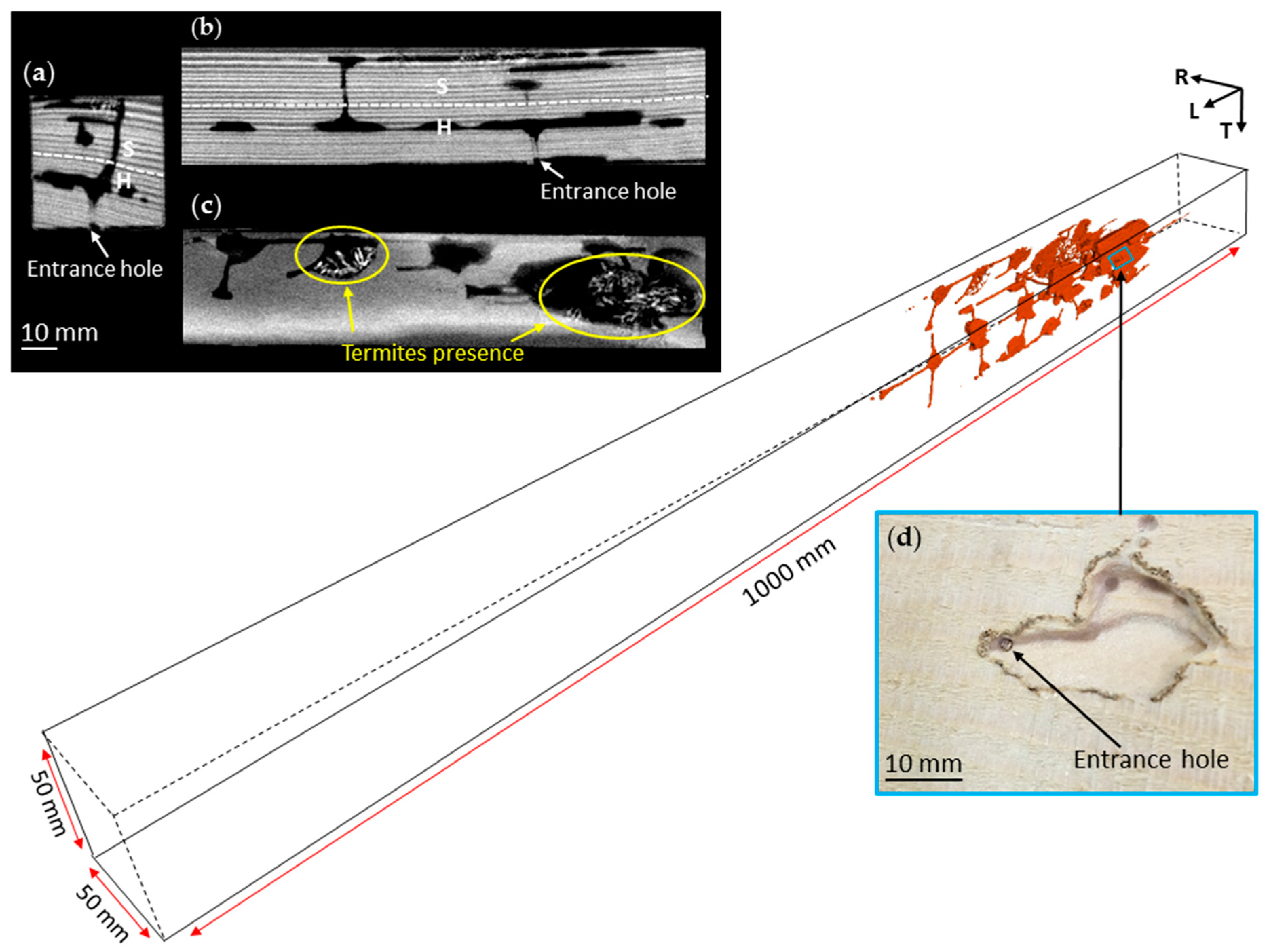
3.1. Nest-Gallery Development in Spruce A Timber
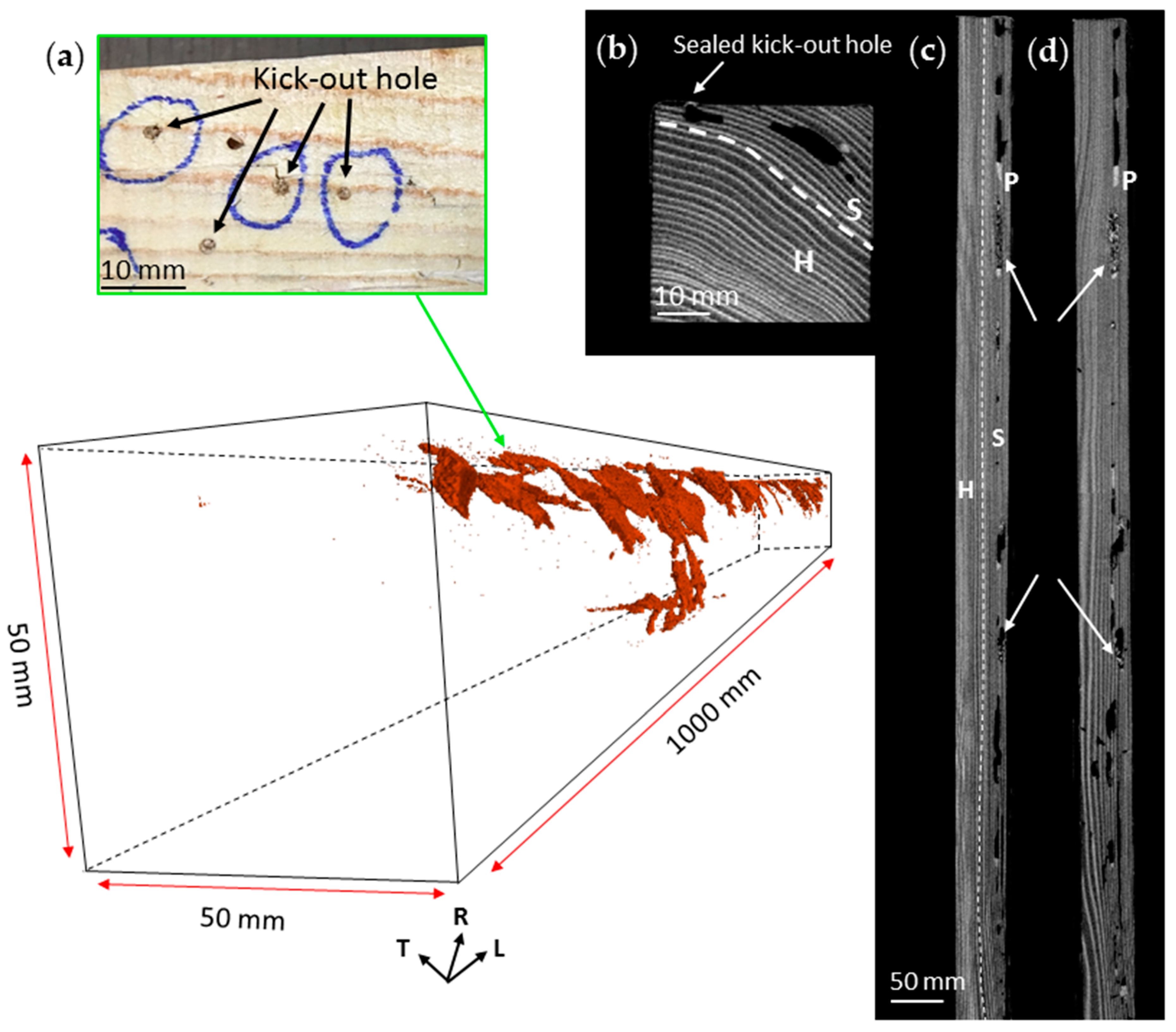
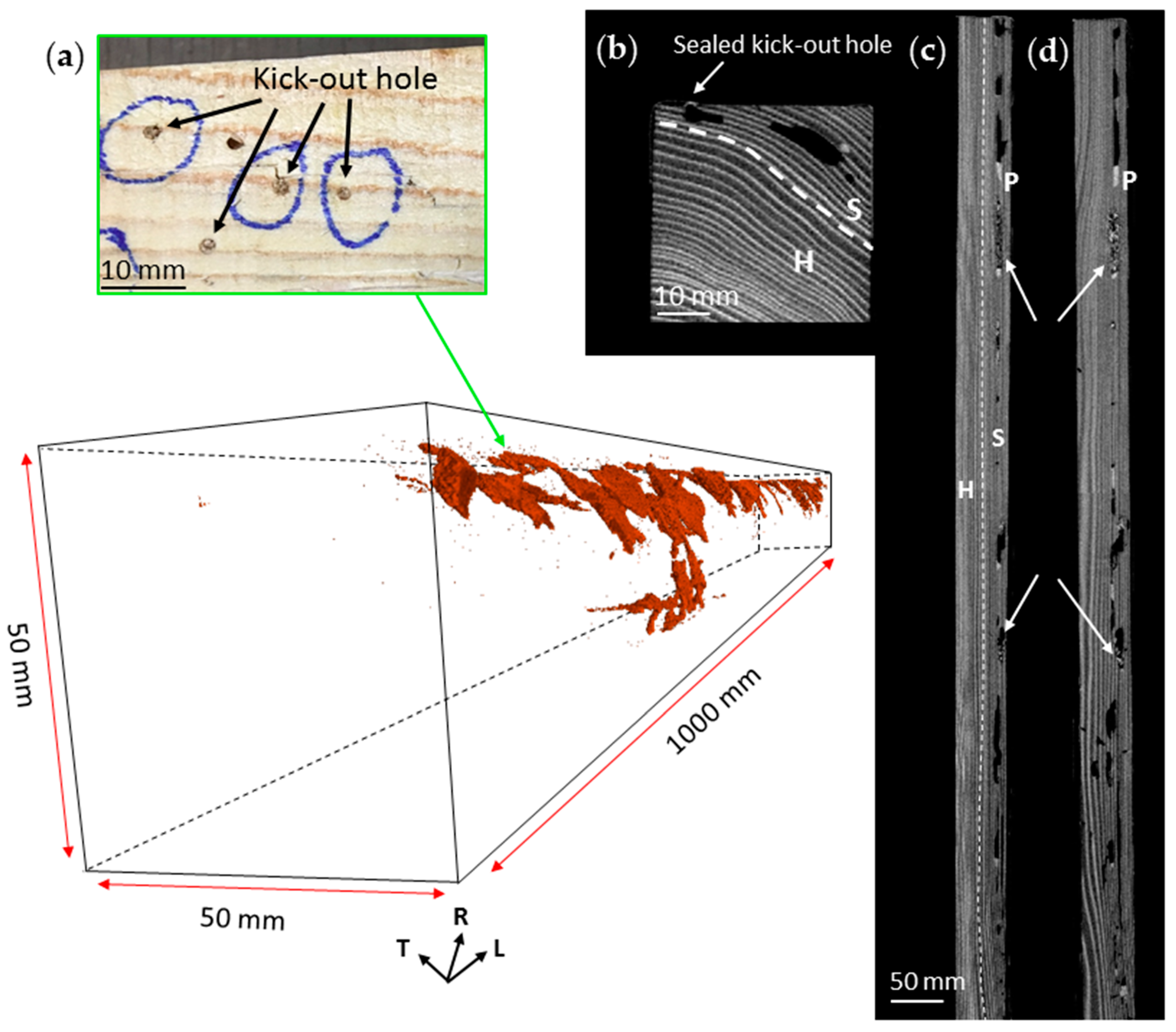
3.2. Nest-Gallery System in Spruce B Timber
3.3. Nest-Gallery System in Sugi Timber
3.4. Colony Composition
- “False” worker (Figure 6g). The term “false” worker follows Korb et al. [13], and defines a majority of individuals in one-piece nesters [6]. They are not doing tasks and work [14] to the same extent as that of the (true) worker in intermediate and separate nesters [6], i.e., they are less involved in truly altruistic working tasks, such as foraging, brood care, or nest duties. Luykx [15] defined this caste as “pseudergates”, since the worker is not a true sterile worker caste; but a totipotent individual [12] that can molt into a secondary reproductive, soldier, or, most commonly, into nymph and alate. They may rather be regarded as large immatures that delay reproductive maturity (‘hopeful reproductives’) [13,16].
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fuchs, A.; Schreyer, A.; Feuerbach, S.; Korb, J. A new technique for termite monitoring using computer tomography and endoscopy. Int. J. Pest Manag. 2004, 50, 63–66. [Google Scholar] [CrossRef]
- Perna, A.; Jost, C.; Couturier, E.; Valverde, S.; Douady, S.; Theraulaz, G. The structure of gallery networks in the nests of termite cubitermes spp. Revealed by X-ray tomography. Naturwissenschaften 2008, 95, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Perna, A.; Jost, C.; Valverde, S.; Gautrais, J.; Theraulaz, G.; Kuntz, P. The topological fortress of termites. In Bio-Inspired Computing and Communication; Lio, P., Yoneki, E., Verma, D.C., Eds.; Springer: Heidelberg, Germany, 2008; Volume 5151, pp. 165–173. [Google Scholar]
- Perna, A.; Valverde, S.; Gautrais, J.; Jost, C.; Sole, R.; Kuntz, P.; Theraulaz, G. Topological efficiency in three-dimensional gallery networks of termite nests. Phys. Stat. Mech. Appl. 2008, 387, 6235–6244. [Google Scholar] [CrossRef]
- Himmi, S.K.; Yoshimura, T.; Yanase, Y.; Oya, M.; Torigoe, T.; Imazu, S. X-ray tomographic analysis of the initial structure of the royal chamber and the nest-founding behavior of the drywood termite incisitermes minor. J. Wood Sci. 2014, 60, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Abe, T. Evolution of life type in termites. In Evolution and Coadaptation in Biotic Communities; Kawano, S., Connell, J.H., Hidaka, T., Eds.; University of Tokyo Press: Tokyo, Japan, 1987; pp. 126–148. [Google Scholar]
- Tschinkel, W.R. The nest architecture of the florida harvester ant, pogonomyrmex badius. J. Insect Sci. 2004, 4, PMC528881. [Google Scholar] [CrossRef]
- Grace, J.K.; Woodrow, R.J.; Oshiro, R.J. Expansive gallery systems of one-piece termites (isoptera: Kalotermitidae). Sociobiology 2009, 54, 37–44. [Google Scholar]
- Lewis, V.R.; Forschler, B.T. Management of drywood termites: Past practices, present situation and future prospects. In Urban Insect Pests: Sustainable Management Strategies; Dhang, P., Ed.; CABI: Boston, MA, USA, 2014; pp. 130–153. [Google Scholar]
- Harvey, P.A. The distribution and biology of the common drywood termite, kalotermes minor: Life history of kalotermes minor. In Termites and Termite Control; Kofoid, C.A., Ed.; University of California Press: Berkeley, CA, USA, 1934; pp. 217–233. [Google Scholar]
- Himmi, S.K.; Yoshimura, T.; Yanase, Y.; Mori, T.; Torigoe, T.; Imazu, S. Wood anatomical selectivity of drywood termite in the nest-gallery establishment revealed by X-ray tomography. Wood Sci. Technol. 2016, 50, 631–643. [Google Scholar] [CrossRef]
- Roisin, Y. Diversity and evolution of caste patterns. In Termites: Evolution, Sociality, Symbioses, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 95–119. [Google Scholar]
- Korb, J.; Hartfelder, K. Life history and development—A framework for understanding developmental plasticity in lower termites. Biol. Rev. 2008, 83, 295–313. [Google Scholar] [CrossRef] [PubMed]
- Korb, J. Workers of a drywood termite do not work. Front. Zool. 2007. [Google Scholar] [CrossRef] [PubMed]
- Luykx, P. Termite colony dynamics as revealed by the sex- and caste-ratios of whole colonies of incisitermes schwarzi banks (isoptera: Kalotermitidae). Insect Soc. 1986, 33, 221–248. [Google Scholar] [CrossRef]
- Myles, T.G. Review of secondary reproduction in termites (insecta: Isoptera) with comments on its role in termite ecology and social evolution. Sociobiology 1999, 33, 1–43. [Google Scholar]
- Morimoto, K. Termite. In Termite and Its Management (in Japanese), 1st ed.; Association, J.T.C., Ed.; Japan Termite Control Association: Tokyo, Japan, 2000; pp. 1–119. (In Japanese) [Google Scholar]
- Weesner, F.M. External anatomy. In Biology of Termites; Krishna, K., Weesner, F.M., Eds.; Academic Press: New York, NY, USA, 1969; Volume 1, pp. 19–47. [Google Scholar]
- Heidecker, J.L.; Leuthold, R.H. The organisation of collective foraging in the harvester termite hodotermes mossambicus (isoptera). Behav. Ecol. Sociobiol. 1984, 14, 195–202. [Google Scholar] [CrossRef]
- Lee, S.-H.; Su, N.-Y. Simulation study on the tunnel networks of subterranean termites and the foraging behavior. J. Asia Pac. Entomol. 2010, 13, 83–90. [Google Scholar] [CrossRef]
- Traniello, J.F.A.; Leuthold, R.H. Behavior and ecology of foraging in termites. In Termites: Evolution, Sociality, Symbioses, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 141–168. [Google Scholar]
- Kano, H.; Shibutani, S.; Hayashi, K.; Iijima, Y.; Doi, S. Effect of high-temperature drying processes on termite resistance of sugi (cryptomeria japonica) heartwood. Mokuzai Gakkaishi 2004, 50, 91–98. (In Japanese) [Google Scholar]
- Arihara, S.; Umeyama, A.; Bando, S.; Kobuke, S.; Imoto, S.; Ono, M.; Yoshikawa, K.; Amita, K.; Hashimoto, S. Termiticidal constituents of the black-heart of cryptomeria japonica. Mokuzai Gakkaishi 2004, 50, 413–421. (In Japanese) [Google Scholar]
- Sogabe, A.; Kinjo, K.; Abe, F.; Yamauchi, T.; Yaga, S. Termiticidal substances from the heartwood of cryptomeria japonica d.Don. Mokuzai Gakkaishi 2000, 46, 124–131. (In Japanese) [Google Scholar]
- Shibutani, S.; Takata, K.; Doi, S. Quantitative comparisons of antitermite extractives in heartwood from the same clones of cryptomeria japonica planted at two different sites. J. Wood Sci. 2007, 53, 285–290. [Google Scholar] [CrossRef]
- Caron, A.; Altaner, C.M.; Gardiner, B.; Jarvis, M.C. Distribution of extractives in sitka spruce (picea sitchensis) grown in the Northern UK. Eur. J. Wood Prod. 2013, 71, 697–704. [Google Scholar] [CrossRef]
- Bonabeau, E. Social insect colonies as complex adaptive systems. Ecosystems 1998, 1, 437–443. [Google Scholar] [CrossRef]
- Levin, S.A. Ecosystems and the biosphere as complex adaptive systems. Ecosystems 1998, 1, 431–436. [Google Scholar] [CrossRef]
- Indrayani, Y.; Yoshimura, T.; Imamura, Y. A novel control strategy for dry-wood termite incisitermes minor infestation using a bait system. J. Wood Sci. 2008, 54, 220–224. [Google Scholar] [CrossRef]
- Indrayani, Y.; Yoshimura, T.; Yanase, Y.; Fujii, Y.; Matsuoka, H.; Imamura, Y. Observation of feeding behavior of three termite (isoptera) species: Incisitermes minor, coptotermes formosanus, and reticulitermes speratus. Sociobiology 2007, 49, 121–134. [Google Scholar]
- Harvey, P.A. Colonization of the common dry-wood termite in wooden structures. In Termites and Termite Control; Kofoid, C.A., Ed.; University of California Press: Berkeley, CA, USA, 1934; pp. 239–265. [Google Scholar]
- Nagin, R. Caste determination in neotermes jouteli (banks). Insect. Soc. 1972, 19, 39–61. [Google Scholar] [CrossRef]
- Atkinson, T.H. Seasonal changes in composition of colonies of the western drywood termite, incisitermes minor (hagen) (isoptera: Kalotermitidae). Available online: http://www.pestboard.ca.gov/howdoi/research/1994.pdf (accessed on 25 May 2016).
- Korb, J.; Lenz, M. Reproductive decision-making in the termite, cryptotermes secundus (kalotermitidae), under variable food conditions. Behav. Ecol. 2004, 15, 390–395. [Google Scholar] [CrossRef]
- Hoffmann, K.; Foster, K.R.; Korb, J. Nest value mediates reproductive decision making within termite societies. Behav. Ecol. 2012, 23, 1203–1208. [Google Scholar] [CrossRef]
- Lenz, M; Yoshimura, T.; Lewis, V.R.; The Commonwealth Scientific and Industrial Research Organization (CSIRO), Canberra, Australia (L.M.); Kyoto University, Kyoto, Japan (Y.T.); University of California, Berkeley, CA, USA (L.V.R.). Unpublished observations. 2012.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Timber Specimens | New Nest-Founding Activity | |||
|---|---|---|---|---|
| Nuptial Flight (Dealate Reproductives) | Colony Activity | Total | ||
| Successful Nest-Founding * | Attempt ** | |||
| Spruce (n = 60) | 29 | 21 | 2 | 52 |
| Sugi (n = 40) | 10 | 8 | 1 | 19 |
| Timber | Detail Location of Infestation on Timber | ||||||
|---|---|---|---|---|---|---|---|
| Specimens | Collected | Opened | Sapwood | Heartwood | Tangential Section | Radial Section | Cross Section |
| Spruce A | November 2012 | March 2016 | 〇 | 〇 | |||
| Spruce B | November 2014 | March 2016 | 〇 | 〇 | |||
| Sugi | November 2014 | March 2016 | 〇 | 〇 | |||
| Timber | Caste Composition | Total | ||||
|---|---|---|---|---|---|---|
| Primary Reproductive | “False” Worker | Soldier | Nymph | Neotenic Reproductive | ||
| Spruce A | - | 38 | 1 | 10 | 1 * | 50 |
| Spruce B | - | 89 | 6 | 21 | 1 ** | 117 |
| Sugi | 1 *** | 102 | 2 | 48 | - | 153 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Himmi, S.K.; Yoshimura, T.; Yanase, Y.; Oya, M.; Torigoe, T.; Akada, M.; Imadzu, S. Nest-Gallery Development and Caste Composition of Isolated Foraging Groups of the Drywood Termite, Incisitermes minor (Isoptera: Kalotermitidae). Insects 2016, 7, 38. https://doi.org/10.3390/insects7030038
Himmi SK, Yoshimura T, Yanase Y, Oya M, Torigoe T, Akada M, Imadzu S. Nest-Gallery Development and Caste Composition of Isolated Foraging Groups of the Drywood Termite, Incisitermes minor (Isoptera: Kalotermitidae). Insects. 2016; 7(3):38. https://doi.org/10.3390/insects7030038
Chicago/Turabian StyleHimmi, S. Khoirul, Tsuyoshi Yoshimura, Yoshiyuki Yanase, Masao Oya, Toshiyuki Torigoe, Masanori Akada, and Setsuo Imadzu. 2016. "Nest-Gallery Development and Caste Composition of Isolated Foraging Groups of the Drywood Termite, Incisitermes minor (Isoptera: Kalotermitidae)" Insects 7, no. 3: 38. https://doi.org/10.3390/insects7030038