Performance of Three Isolates of Metarhizium anisopliae and Their Virulence against Zeugodacus cucurbitae under Different Temperature Regimes, with Global Extrapolation of Their Efficiency

 , and
, and
Abstract
:
1. Introduction
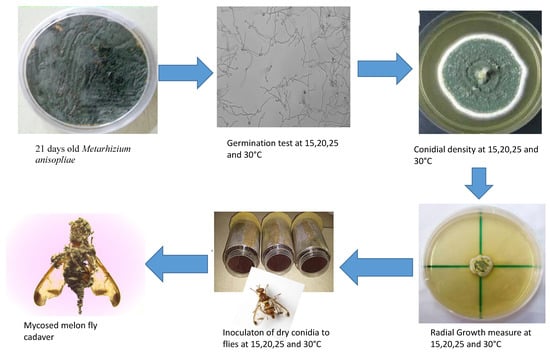
2. Materials and Methods
2.1. Rearing of Z. cucurbitae
2.2. Fungal Isolates
2.3. Effect of Temperature on Conidial Germination
2.4. Effect of Temperatures on Sporulation
2.5. Effect of Temperature on Radial Growth
2.6. Effect of Temperature on Fungal Virulence
2.7. Data Analysis
3. Results
3.1. Effect of Temperature on Conidial Germination
3.2. Effect of Temperature on Radial Growth
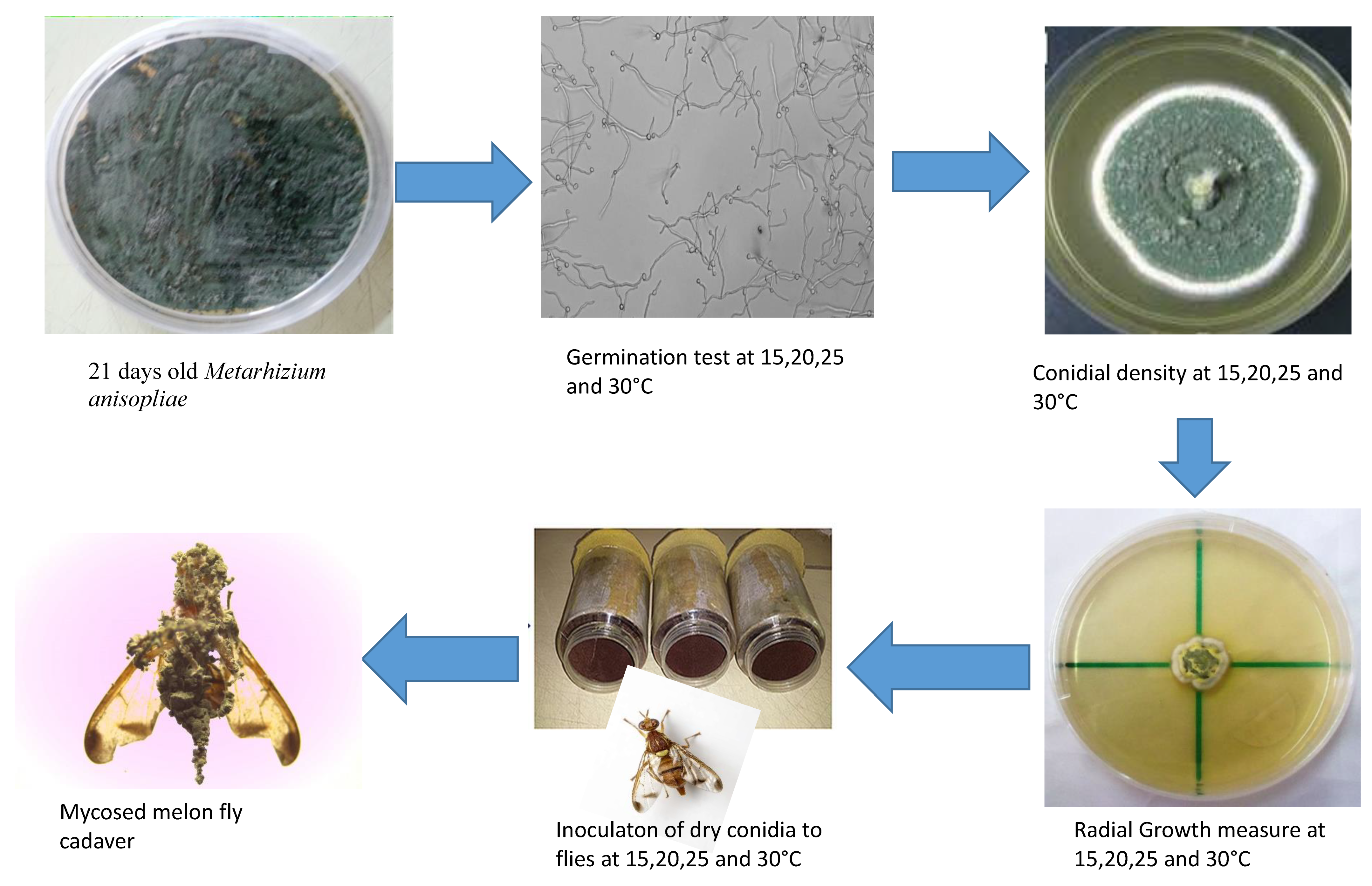
3.3. Effect of Temperature on Conidial Density
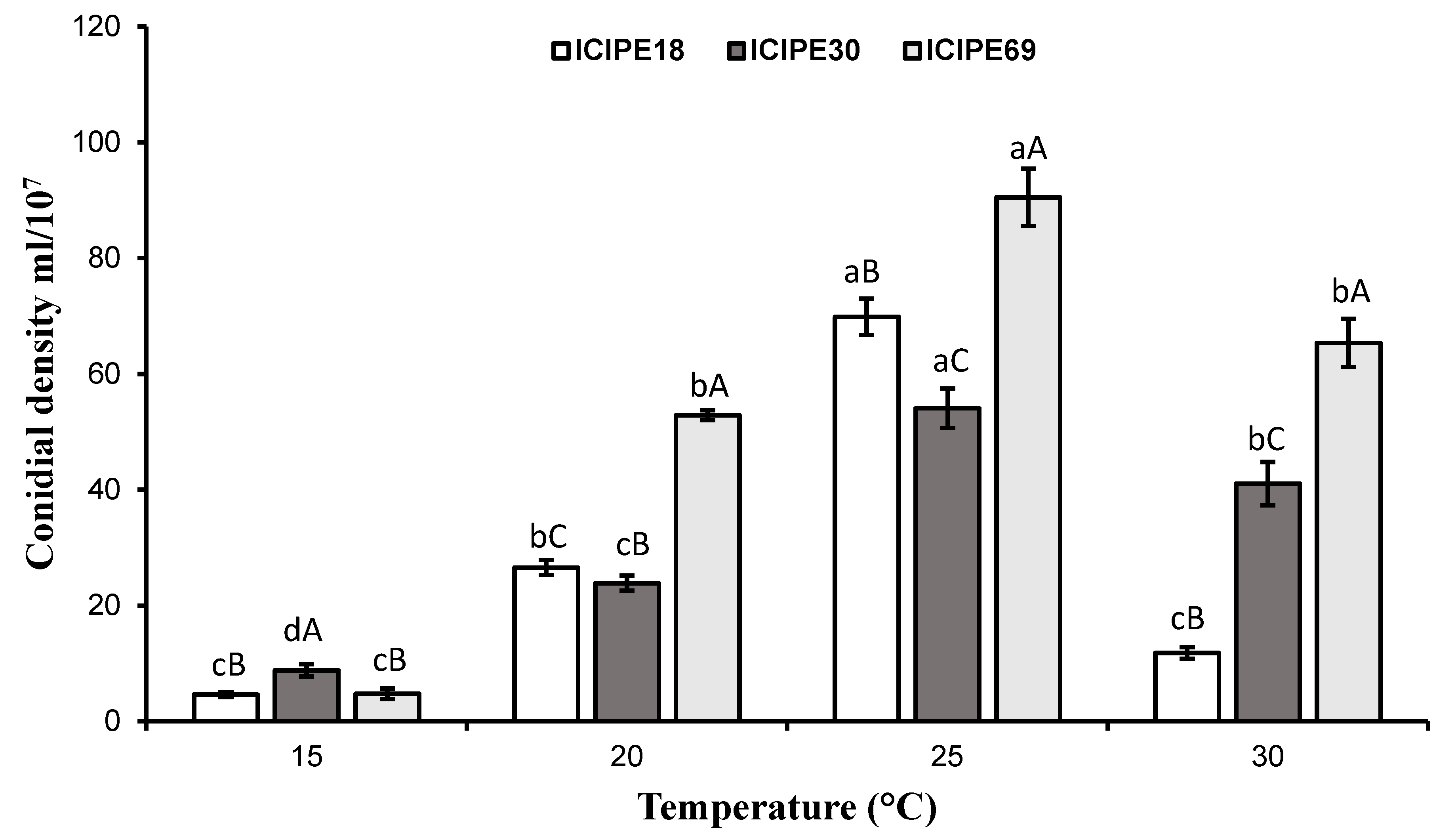
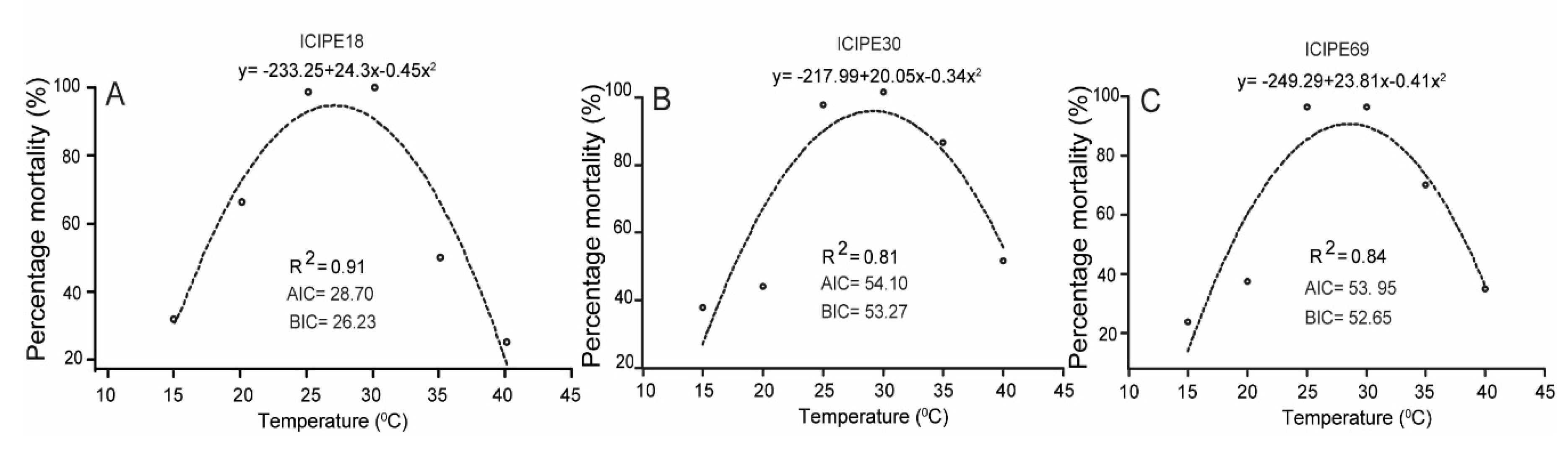
3.4. Effect of Temperature on Virulence of M. anisopliae Isolates to Z. cucurbitae
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Deguine, J.P.; Atiama–Nurbel, T.; Aubertot, J.N.; Augusseau, X.; Atiama, M.; Jacquot, M.; Reynaud, B. Agroecological management of cucurbit–infesting fruit fly: A review. Agron. Sustain. Dev. 2015, 35, 937–965. [Google Scholar] [CrossRef]
- Madzivhandila, T.; Sibanda, S.; Yamdjeu, A.W.; Moalosi, K.; Gwelo, F.A. Achieving Food Security and Nutrition. Afr. Agric. Status Rep. 2016, 19, 234–251. [Google Scholar]
- Horticultural Crops Development Authority (HCDA). Horticulture Validated Report; Kenya Bureau of Standards: Nairobi, Kenya, 2014.
- Carmo–Sousa, M.; Moreno, A.; Plaza, M.; Garzo, E.; Fereres, A. Cucurbit aphid–borne yellows virus (CABYV) modifies the alighting, settling and probing behaviour of its vector Aphis gossypii favouring its own spread. Ann. Appl. Biol. 2016, 169, 284–297. [Google Scholar] [CrossRef]
- Pareek, B.L.; Kavadia, V.S. Economic insecticidal control of two major pests of musk melon, Cucumis melo in the pumpkin beetle, Raphidopalpa spp. and the fruitfly, Dacus cucurbitae in Rajasthan, India. Int. J. Pest Manag. 1988, 34, 15–18. [Google Scholar]
- Roggero, P.; Dellavalle, G.; Lisa, V.; Stravato, V.M. First Report of Moroccan Watermelon Mosaic Potyvirus in Zucchini in Italy. Plant Dis. 2007, 82, 351. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.K.; Kim, B.S.; Chang, S.W.; Hwang, B.K. Aggressiveness to pumpkin cultivars of isolates of Phytophthora capsici from pumpkin and pepper. Plant Dis. 2001, 85, 497–500. [Google Scholar] [CrossRef]
- Zhao, Y.R.; Li, X.; Yu, K.Q.; Cheng, F.; He, Y. Hyperspectral Imaging for Determining Pigment Contents in Cucumber Leaves in Response to Angular Leaf Spot Disease. Nature 2016, 6, 27790. [Google Scholar] [CrossRef]
- Ansari, S.M.; Hasan, F.; Ahmad, N. Threats to fruit and vegetable crops: Fruit flies (Tephritidae) ecology, behaviour, and management. J. Crop Sci. Biotech. 2012, 15, 169–188. [Google Scholar] [CrossRef]
- Godefroid, M.; Cruaud, A.; Rossi, J.P.; Rasplus, J.Y. Assessing the risk of invasion by tephritid fruit flies: Intraspecific divergence matters. PLoS ONE 2015, 10, e0135209. [Google Scholar] [CrossRef]
- White, I.; Goodger, M. African Dacus (Diptera: Tephritidae); New Species and Data, with Particular Reference to the Tel Aviv University Collection. Zootaxa 2009, 49, 1–49. [Google Scholar] [CrossRef]
- Mwatawala, M.; Kudra, A.; Mkiga, A.; Godfrey, E.; Jeremiah, S.; Virgilio, M.; De Meyer, M. Preference of Zeugodacus cucurbitae (Coquillett) for three commercial fruit vegetable hosts in natural and semi natural conditions. Fruits 2015, 70, 333–339. [Google Scholar] [CrossRef]
- Mkiga, A.M.; Mwatawala, M.W. Developmental Biology of Zeugodacus cucurbitae (Diptera: Tephritidae) in Three Cucurbitaceous Hosts at Different Temperature Regimes. J. Insect Sci. 2015, 15, 160. [Google Scholar] [CrossRef] [PubMed]
- De Meyer, M.; Delatte, H.; Mwatawala, M.; Quilici, S.; Vayssières, J.F.; Virgilio, M. A review of the current knowledge on Zeugodacus cucurbitae (Coquillett) (Diptera: Tephritidae) in Africa, with a list of species included in Zeugodacus. ZooKeys 2015, 540, 539–557. [Google Scholar] [CrossRef] [PubMed]
- Kwasi, W. Assessment of Fruit Fly Damage and Implications for the Dissemination of Management Practices for Mango Production in the Upper West Region of Ghana. J. Dev. Sustain. Agric. 2008, 3, 117–134. [Google Scholar]
- Kambura, C.; Tanga, C.M.; Kilalo, D.; Muthomi, J.; Salifu, D.; Rwomushana, I.; Mohamed, S.A.; Ekesi, S. Composition, host range and host suitability of vegetable–infesting tephritids on cucurbits cultivated in Kenya. Afr. Entomol. 2018, 26, 379–397. [Google Scholar] [CrossRef]
- Bissdorf, J.; Weber, C. Field Guide to Non–Chemical Pest Management in Mango Production; Pesticide Action Network (PAN): Hamburg, Germany, 2005. [Google Scholar]
- Dhillon, M.; Singh, R.; Naresh, J.; Sharma, H. The melon fruit fly, Bactrocera cucurbitae: A review of its biology and management. J. Insect Sci. 2005, 5, 40. [Google Scholar] [CrossRef] [PubMed]
- Jager, T.; Rikken, M.G.J.; van der Poel, P. Uncertainty Analysis of EUSES: Improving Risk Management by Probabilistic Risk Assessment; National Institute of Public Health and the Environment: Bilthoven, The Netherlands, 1997. [Google Scholar]
- Jin, T.; Lin, Y.Y.; Jin, Q.A.; Wen, H.B.; Peng, Z.Q. Population susceptibility to insecticides and the development of resistance in Bactrocera cucurbitae (Diptera: Tephritidae). J. Econ. Entomol. 2016, 109, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Inglis, G.D.; Goettel, M.S.; Butt, T.M.; Strasser, H. Use of hyphomycetous fungi for managing insect pests. In Fungi as Biocontrol Agents: Problems and Potential; CAB International: Wallingford, UK, 2001; pp. 23–69. [Google Scholar]
- Onsongo, S.K.; Gichimu, B.M.; Akutse, K.S.; Dubois, T.; Mohamed, S. Pathogenicity of Metarhizium anisopliae (Metsch.) Sorokin and Beauveria bassiana (Balsamo) Vuillemin, on adults of Melon fly (Zeugodacus cucurbitae) (Diptera: Tephritidae) (Unpublished, manuscript in preparation). Mycopathologia 2003, 156, 375–382. [Google Scholar]
- Jaronski, S.T. Ecological factors in the inundative use of fungal entomopathogens. BioControl 2010, 55, 159–185. [Google Scholar] [CrossRef]
- Tefera, T.; Pringle, K. Germination, Radial Growth, and Sporulation of Beauveria bassiana and Metarhizium anisopliae Isolates and Their Virulence to Chilo partellus (Lepidoptera: Pyralidae) at Different Temperatures. Biocontrol. Sci. Technol. 2010, 13, 699–704. [Google Scholar] [CrossRef]
- Ugine, T.A. The effect of temperature and exposure to Beauveria bassiana on tarnished plant bug Lygus lineolaris (Heteroptera: Miridae) population dynamics, and the broader implications of treating insects with entomopathogenic fungi over a range of temperatures. Biol. Control 2011, 59, 373–383. [Google Scholar] [CrossRef]
- Mishra, S.; Kumar, P.; Malik, A. Effect of temperature and humidity on pathogenicity of native Beauveria bassiana isolate against Musca domestica L. J. Parasit. Dis. 2013, 39, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Tumuhaise, A.V.; Ekesi, S.; Maniania, N.K.; Tonnang, H.E.Z.; Tanga, M.; Ndegwa, P.N.; Mohamed, S.A. Temperature–Dependent Growth and Virulence, and Mass Production Potential of Two Candidate Isolates of Metarhizium anisopliae (Metschnikoff) Sorokin for Managing Maruca vitrata Fabricius (Lepidoptera: Crambidae) on Cowpea. Afr. Entomol. 2018, 26, 73–83. [Google Scholar] [CrossRef]
- Duyck, P.F.; Sterlin, J.F.; Quilici, S. Survival and development of different life stages of Bactrocera zonata (Diptera: Tephritidae) reared at five constant temperatures compared to other fruit fly species. Bull. Entomol. Res. 2004, 94, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Rwomushana, I.; Ekesi, S.; Ogol, C.K.P.O.; Gordon, I. Effect of temperature on development and survival of immature stages of Bactrocera invadens (Diptera: Tephritidae). J. Appl. Entomol. 2008, 132, 832–839. [Google Scholar] [CrossRef]
- Dimbi, S.; Maniania, N.K.; Lux, S.A.; Mueke, J.M. Effect of constant temperatures on germination, radial growth and virulence of Metarhizium anisopliae to three species of African tephritid fruit flies. BioControl 2004, 49, 83–94. [Google Scholar] [CrossRef]
- Chang, C.L.; Caceres, C.; Jang, E.B. A novel liquid diet and its rearing system for melon fly, Bactrocera cucurbitae (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2004, 97, 524–528. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with R, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Fargues, J.; Maniania, N.K.; Delmas, J.; Smits, N. Influence de la température sur la croissance in vitro d’ hyphomycètes entomopathogènes. Agronomie 1992, 12, 557–564. [Google Scholar] [CrossRef]
- Ouedraogo, A.; Fargues, J.; Goettel, M.S.; Lomer, C.J. Effect of temperature on vegetative growth among isolates of Metarhizium anisopliae and M. flavoviride. Mycopathologia 1997, 137, 37–43. [Google Scholar] [CrossRef]
- Yeo, H.; Pell, J.K.; Alderson, P.G.; Clark, S.J.; Pye, B.J. Laboratory evaluation of temperature effects on the germination and growth of entomopathogenic fungi and on their pathogenicity to two aphid species. Pest Manag. Sci. 2003, 59, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Davidson, G.; Phelps, K.; Sunderland, K.D.; Pell, J.K.; Ball, B.V.; Shaw, K.E.; Chandler, D. Study of temperature—Growth interactions of entomopathogenic fungi with potential for control of Varroa destructor (Acari: Mesostigmata) using a nonlinear model of poikilotherm development. J. Appl. Entomol. 2003, 94, 816–825. [Google Scholar] [CrossRef]
- Bayissa, W.; Ekesi, S.; Mohamed, S.A.; Kaaya, G.P.; Wagacha, J.M.; Hanna, P.; Maniania, N.K. Selection of fungal isolates for virulence against three aphid pest species of crucifers and okra. J. Pest Sci. 2017, 90, 355–368. [Google Scholar] [CrossRef]
- Arnhold, E. Easy Nonlinear Model. R Package Version 5.0. 2017. Available online: https://cran.r-project.org/web/packages/easynls/easynls.pdf (accessed on 4 July 2019).
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Altre, J.A.; Vandenberg, J.D.; Cantone, F.A. Pathogenicity of Paecilomyces fumosoroseus isolates to diamondback moth, Plutella xylostella: Correlation with spore size, germination speed, and attachment to cuticle. J. Invertebr. Pathol. 1999, 73, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Shah, F.A.; Wang, C.S.; Butt, T.M. Nutrition influences growth and virulence of the insect–pathogenic fungus Metarhizium anisopliae. FEMS Microbiol. Biol. Lett. 2005, 251, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.; Magan, N.; Mead, A.; Chandler, D. Development of a population–based threshold model of conidial germination for analysing the effects of physiological manipulation on the stress tolerance and infectivity of insect pathogenic fungi. Environ. Microb. 2006, 8, 1625–1634. [Google Scholar] [CrossRef] [PubMed]
- Talaei–hassanloui, R.; Kharazi–pakdel, A.; Goettel, M.S.; Little, S.; Mozaffari, J. Germination polarity of Beauveria bassiana conidia and its possible correlation with virulence. J. Inverteb. Pathol. 2007, 94, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Bugeme, D.M.; Maniania, N.K.; Knapp, M.; Boga, H.I. Effect of temperature on virulence of Beauveria bassiana and Metarhizium anisopliae isolates to Tetranychus evansi. Exp. Appl. Acarol. 2008, 46, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Gougouli, M.; Koutsoumanis, K.P. Modeling germination of fungal spores at constant and fluctuating temperature conditions. Int. J. Food Microbiol. 2012, 152, 153–161. [Google Scholar] [CrossRef]
- Pardo, E.; Ramos, A.J.; Sanchis, V.; Marín, S. Modelling of effects of water activity and temperature on germination and growth of ochratoxigenic isolates of Aspergillus ochraceus on a green coffee–based medium. Int. J. Food Microbiol. 2005, 98, 1–9. [Google Scholar] [CrossRef]
- Cabanillas, H.E.; Jones, W.A. Effects of Temperature and Culture Media on Vegetative Growth of an Entomopathogenic Fungus Isaria sp. (Hypocreales: Clavicipitaceae) Naturally Affecting the Whitefly. Mycopathologia 2009, 167, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Arthurs, S.; Thomas, M.B. Effects of Temperature and Relative Humidity on Sporulation of Metarhizium anisopliae var. acridum in Mycosed Cadavers of Schistocerca gregaria. J. Invertebr. Pathol. 2001, 78, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Borisade, O.A.; Magan, N. Growth and sporulation of entomopathogenic Beauveria bassiana, Metarhizium anisopliae, Isaria farinosa and Isaria fumosorosea strains in relation to water activity and temperature interactions. Biocontrol Sci. Technol. 2014, 24, 999–1011. [Google Scholar] [CrossRef]
- Chauvet, E.; Suberkropp, K. Temperature and sporulation of aquatic hyphomycetes. Appl. Environ. Microbiol. 1998, 64, 1522–1525. [Google Scholar] [PubMed]
- King, W.T.; Madden, L.V.; Ellis, M.A.; Wilson, V. Effects of Temperature on Sporulation and Latent Period of Colletotrichum spp. Infect. Strawb. Fruit Plant Dis. 2007, 81, 77–84. [Google Scholar]
- Klass, J.I.; Blanford, S.; Thomas, M.B. Development of a model for evaluating the effects of environmental temperature and thermal behaviour on biological control of locusts and grasshoppers using pathogens. Agric. Forest Entomol. 2007, 9, 189–199. [Google Scholar] [CrossRef]
- Rangel, D.E.N.; Fernandes, É.K.K.; Dettenmaier, S.J.; Roberts, D.W. Thermotolerance of germlings and mycelium of the insect–pathogenic fungus Metarhizium spp. and mycelial recovery after heat stress. J. Basic Microbiol. 2010, 50, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Tumuhaise, V. Laboratory and Field Evaluation of Entomopathogenic Fungi, Metarhizium anisopliae and Beauveria bassiana, for Management of the Legume Pod Borer, Maruca vitrata (Fabricius) on Cowpea. Ph.D. Thesis, University of Nairobi, Nairobi, Kenya, 2015. [Google Scholar]
- Street, A.; Shelly, T.E.; Kurashima, R.S. Capture of Mediterranean Fruit Flies and Melon Flies (Diptera: Tephritidae) in Food–Baited Traps in Hawaii. Proc. Hawaii Entomol. Soc. 2016, 48, 71–84. [Google Scholar]
- Ryckewaert, P.; Deguine, J.P.; Brévault, T.; Vayssières, J.F. Fruit flies (Diptera: Tephritidae) on vegetable crops in Reunion Island (Indian Ocean): State of knowledge, control methods and prospects for management. Fruits 2010, 65, 113–130. [Google Scholar] [CrossRef]
- Broza, M.; Pereira, P.M.; Stimac, J.L. The Non–susceptibility of Soil Collembola to Insect Pathogens and Their Potential as Scavengers of Microbial Pesticides. Pedobiologia 2001, 45, 523–534. [Google Scholar] [CrossRef]
- Dromph, M.K.; Vestergaard, S. Pathogenicity and Attractiveness of Entomopathogenic Hyphomycetes Fungi to Collembolans. Appl. Soil. Ecol. 2002, 21, 197–210. [Google Scholar] [CrossRef]
- Pourian, H.R.; Talaei–Hassanloui, R.; Kosari, A.A.; Ashouri, A. Effects of Metarhizium anisopliae on searching, feeding and predation by Orius albidipennis (Hem., Anthocoridae) on Thrips tabaci (Thy., Thripidae) larvae, Biocontrol. Sci. Technol. 2011, 21, 15–21. [Google Scholar]
- Lobo, L.S.; Rodrigues, J.; Luz, C. Effectiveness of Metarhizium anisopliae formulations against dengue vectors under laboratory and field conditions. Biocontrol. Sci. Technol. 2016, 26, 386–401. [Google Scholar] [CrossRef]
- Bugeme, D.M.; Knapp, M.; Ekesi, S.; Chabi–Olaye, A.; Boga, H.I.; Maniania, N.K. Efficacy of Metarhizium anisopliae in controlling the two–spotted spider mite Tetranychus urticae on common bean in screenhouse and field experiments. J. Insect Sci. 2015, 22, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Erler, F.; Pradier, T.; Aciloglu, B. Field evaluation of an entomopathogenic fungus, Metarhizium brunneum strain F52, against pear psylla, Cacopsylla pyri. Pest Manag. Sci. 2014, 70, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Magda, S.; Said, S. Efficacy of Two Entomopathogenic Fungi against Corn Pests Under Laboratory and Field Conditions in Egypt. Eur. J. Acad. Res. 2014, 1, 1–6. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
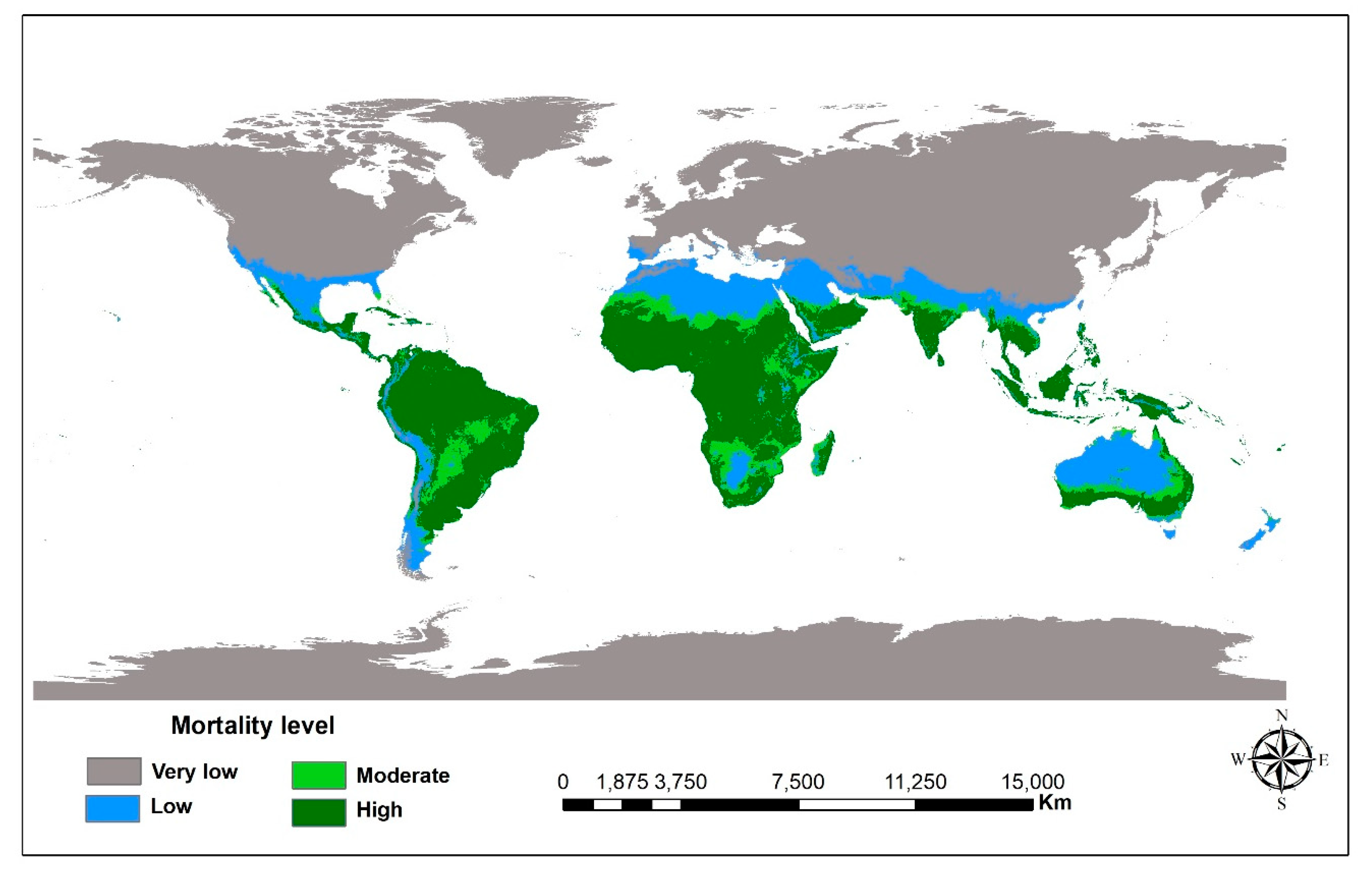{kind=link}
| Temperature | Fungal Isolates | ||
|---|---|---|---|
| ICIPE 18 | ICIPE 30 | ICIPE 69 | |
| 15 °C | 4.26 ± 0.35 cA | 3.65 ± 0.20 cAB | 2.90 ± 0.28 cB |
| 20 °C | 69.83 ± 1.82 bA | 71.74 ± 1.27 bA | 71.75 ± 2.99 bA |
| 25 °C | 98.86 ± 0.48 aA | 97.64 ± 0.45 aA | 98.96 ± 0.49 aA |
| 30 °C | 98.00 ± 0.23 aA | 97.69 ± 0.41 aA | 98.56 ± 0.27 aA |
| Temperature | F3,36 = 3084.63 | p < 0.001 | |
| Isolate | F2,36 = 1.23 | p = 0.304 | |
| Temperature x isolate | F6,36 = 1.57 | p = 0.184 | |
| Fungal Isolates | |||
|---|---|---|---|
| Temperature | ICIPE 18 | ICIPE 30 | ICIPE 69 |
| 15 °C | 1.32 ± 0.10 dA | 0.18 ± 0.07 dB | 1.0 ± 0.06 dB |
| 20 °C | 2.48 ± 0.11 cA | 1.65 ± 0.13 cB | 2.44 ± 0.15 cA |
| 25 °C | 3.15 ± 0.16 bA | 2.85 ± 0.04 bA | 3.34 ± 0.13 bA |
| 30 °C | 3.88 ± 0.18 aA | 3.79 ± 0.15 aA | 4.08 ± 0.24 aA |
| Temperature | F3,36 = 241.712 | p < 0.001 | |
| Isolate | F2,36 = 13.267 | p < 0.001 | |
| Temperature x isolate | F6,36 = 2.006 | p = 0.0904 | |
| Temperature | Fungal Isolates | ||
|---|---|---|---|
| ICIPE 18 | ICIPE 30 | ICIPE 69 | |
| 15 °C | 31.25 ± 3.15 cA | 16.25 ± 3.15 bA | 23.75 ± 5.15 bA |
| 20 °C | 66.25 ± 3.75 bA | 22.5 ± 3.23 bB | 37.5 ± 8.29 bB |
| 25 °C | 98.75 ± 1.25 aA | 76.25 ± 3.15 aB | 96.25 ± 1.25 aA |
| 30 °C | 100.00 ± 0.00 aA | 80.00 ± 2.04 aB | 96.25 ± 2.39 aA |
| Temperature | F3,36 = 214.76 | p < 0.001 | |
| isolate | F2,36 = 56.46 | p < 0.001 | |
| Temperature x isolate | F6,36 = 2.63 | p < 0.05 | |
| ICIPE 18 | ICIPE 30 | ICIPE 69 | ||||
|---|---|---|---|---|---|---|
| Temperature | LT50 (days) | LT90 (days) | LT50 (days) | LT90 (days) | LT50 (days) | LT90 (days) |
| 15 °C | 4.89 | 7.41 | 5.25 | 7.00 | 5.40 | 8.04 |
| (4.72–5.06) | (7.04–7.78) | (5.04–5.46) | (6.61–7.40) | (5.19–5.64) | (7.56–8.52) | |
| 20 °C | 3.36 | 5.07 | 5.32 | 7.69 | 4.46 | 6.53 |
| (3.3–3.41) | (4.95–5.19) | (5.10–5.54) | (7.26–8.13) | (4.34–4.58) | (6.28–6.78) | |
| 25 °C | 2.71 | 3.84 | 2.99 | 4.70 | 2.71 | 3.92 |
| (2.68–2.75) | (3.78–3.9) | (2.94–3.04) | (4.59–4.8) | (2.67–2.75) | (3.85–3.98) | |
| 30 °C | 2.63 | 3.72 | 2.99 | 4.54 | 2.61 | 3.70 |
| (2.59–2.66) | (3.66–3.77) | (2.94–3.03) | (4.45–4.63) | (2.57–2.64) | (3.65–3.76) | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Onsongo, S.K.; Gichimu, B.M.; Akutse, K.S.; Dubois, T.; Mohamed, S.A. Performance of Three Isolates of Metarhizium anisopliae and Their Virulence against Zeugodacus cucurbitae under Different Temperature Regimes, with Global Extrapolation of Their Efficiency. Insects 2019, 10, 270. https://doi.org/10.3390/insects10090270
Onsongo SK, Gichimu BM, Akutse KS, Dubois T, Mohamed SA. Performance of Three Isolates of Metarhizium anisopliae and Their Virulence against Zeugodacus cucurbitae under Different Temperature Regimes, with Global Extrapolation of Their Efficiency. Insects. 2019; 10(9):270. https://doi.org/10.3390/insects10090270
Chicago/Turabian StyleOnsongo, Susan K., Bernard M. Gichimu, Komivi S. Akutse, Thomas Dubois, and Samira A. Mohamed. 2019. "Performance of Three Isolates of Metarhizium anisopliae and Their Virulence against Zeugodacus cucurbitae under Different Temperature Regimes, with Global Extrapolation of Their Efficiency" Insects 10, no. 9: 270. https://doi.org/10.3390/insects10090270