
Mechanisms of High Temperature Resistance of Synechocystis sp. PCC 6803: An Impact of Histidine Kinase 34




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Strains
2.2. Experimental Setup
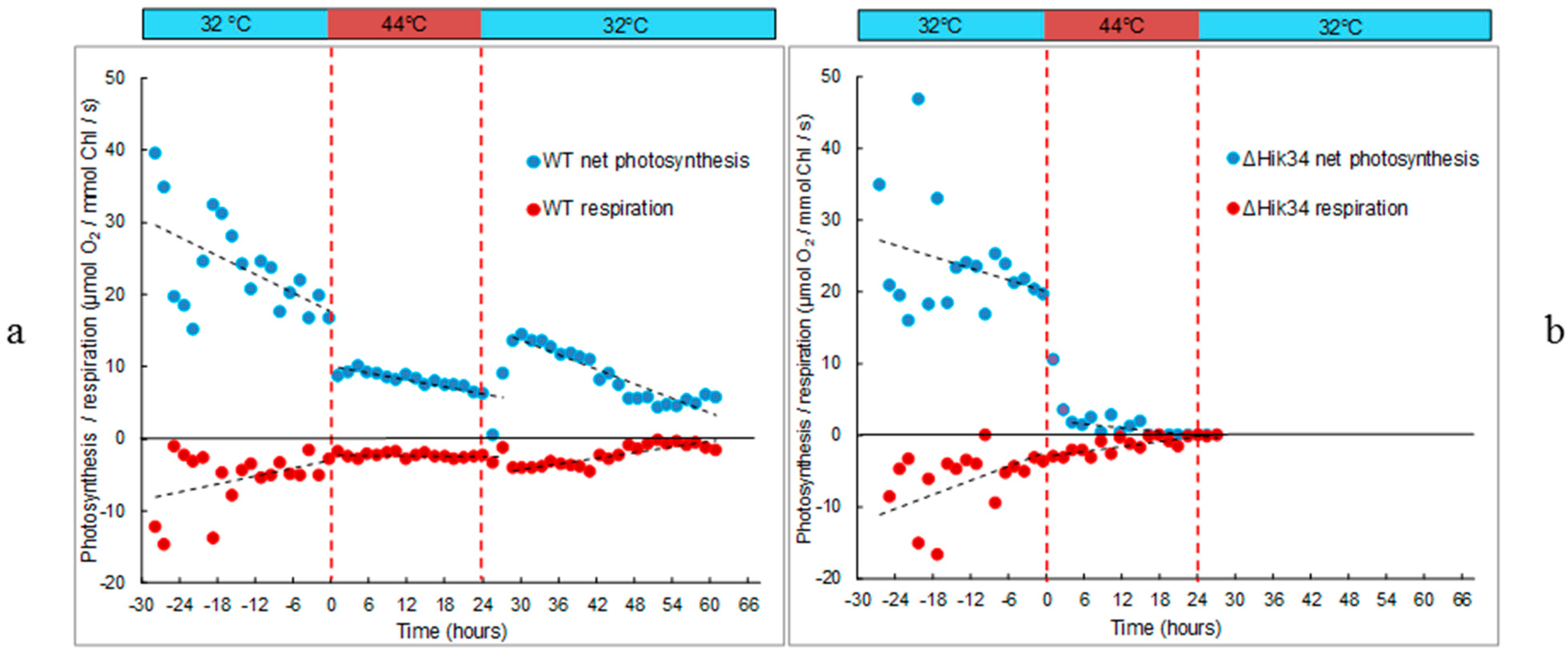
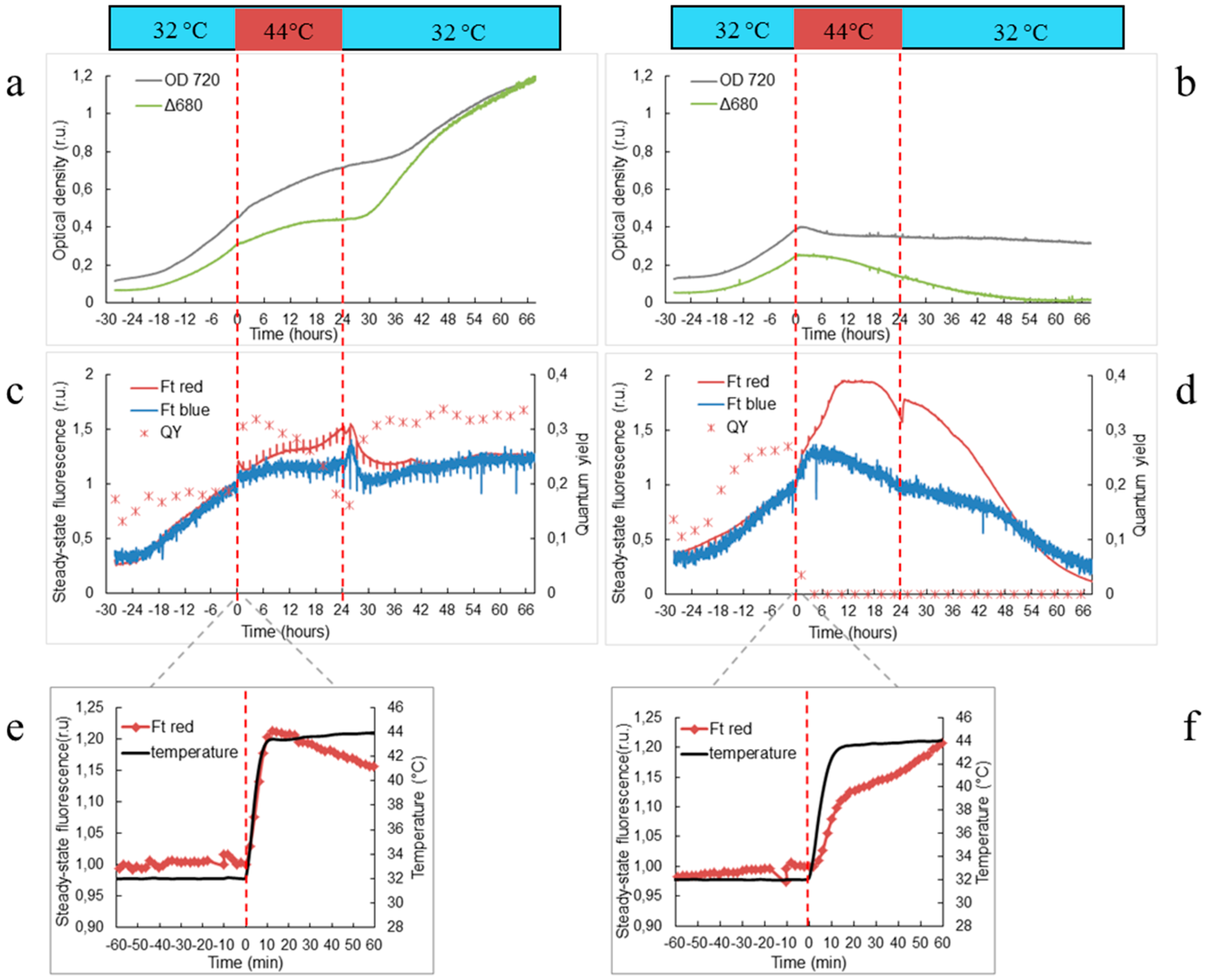
2.3. Automatic Measurements of Respiration and Photosynthesis
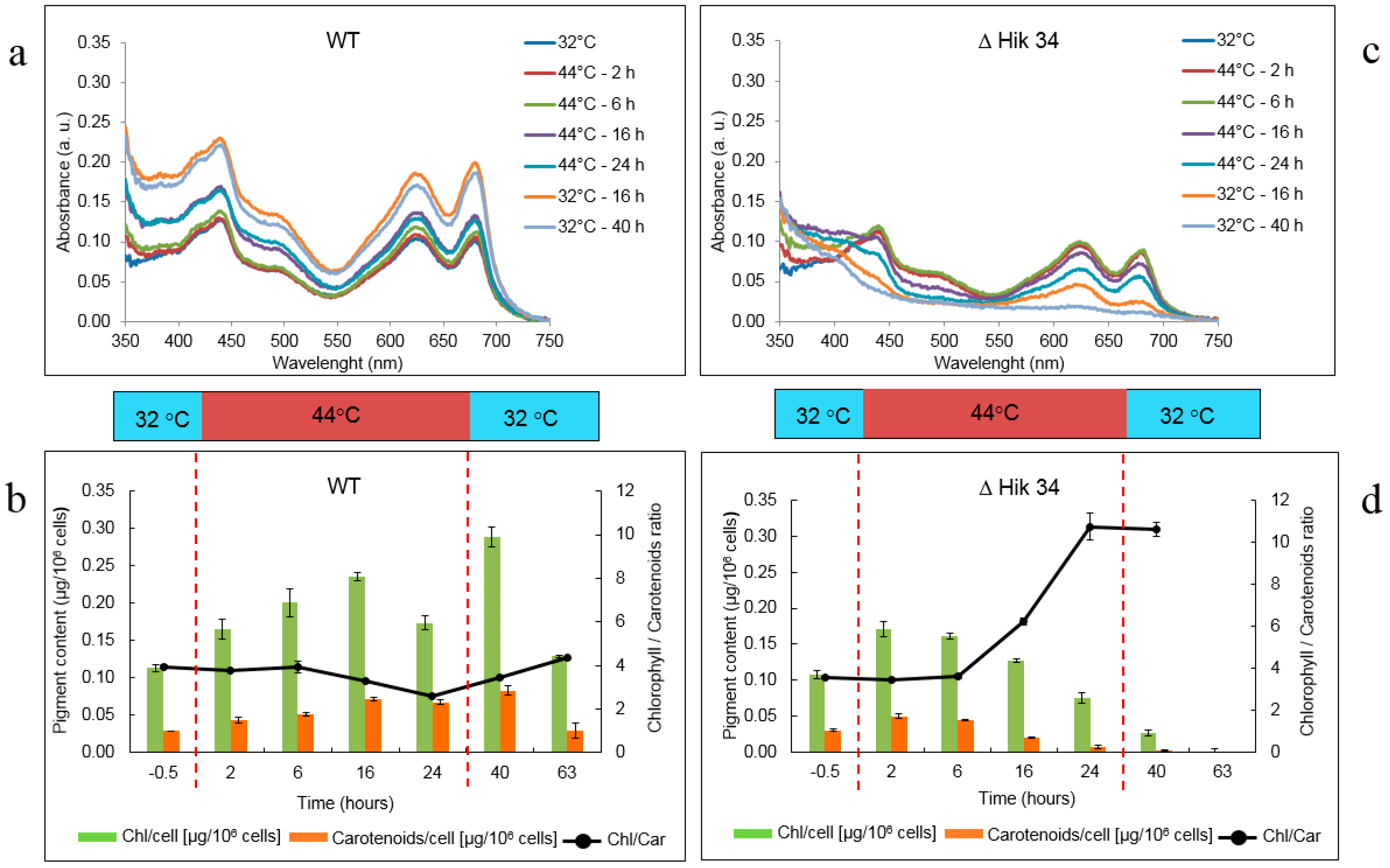
2.4. Pigment Analysis
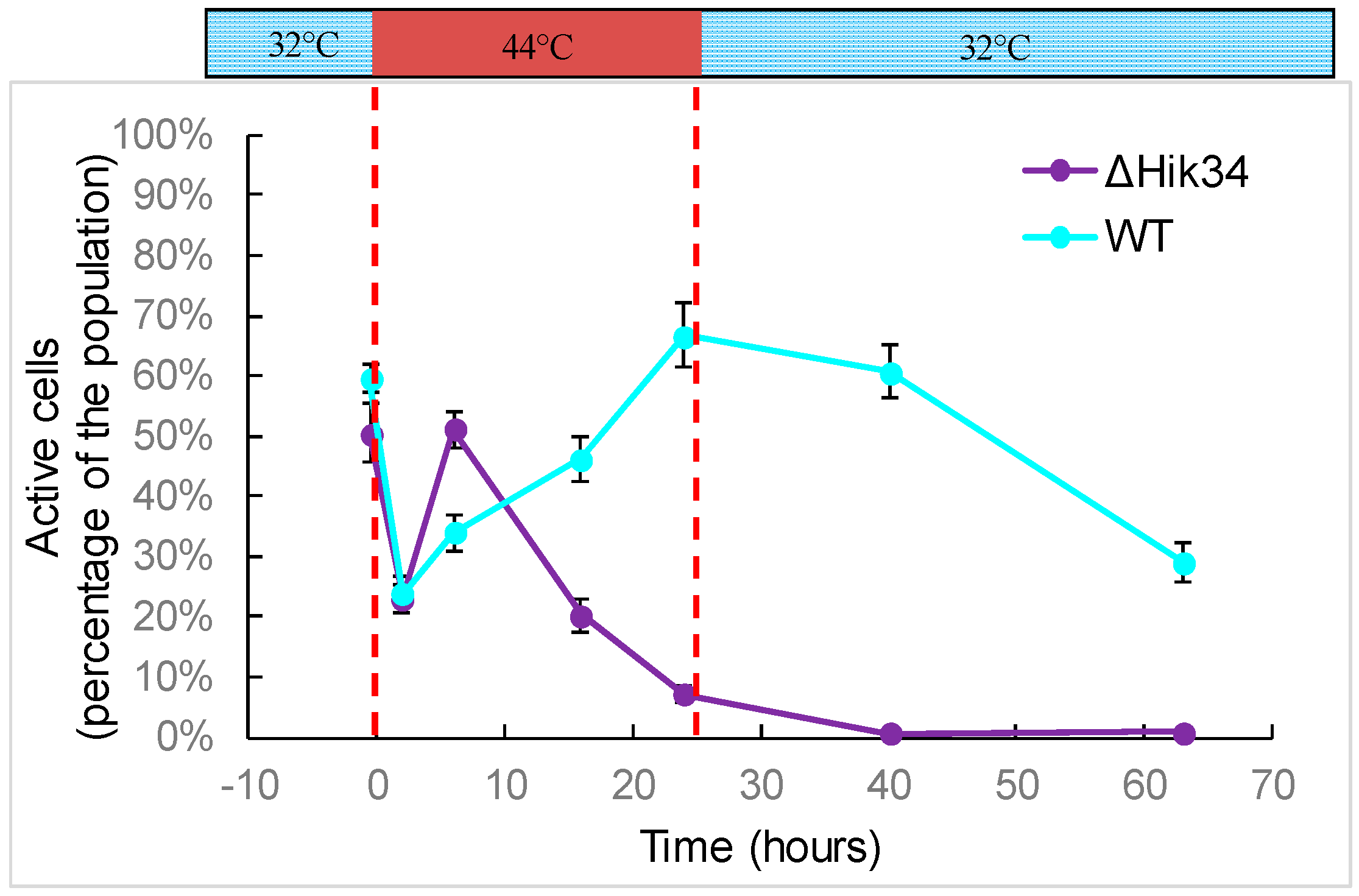
2.5. FDA Activity Test
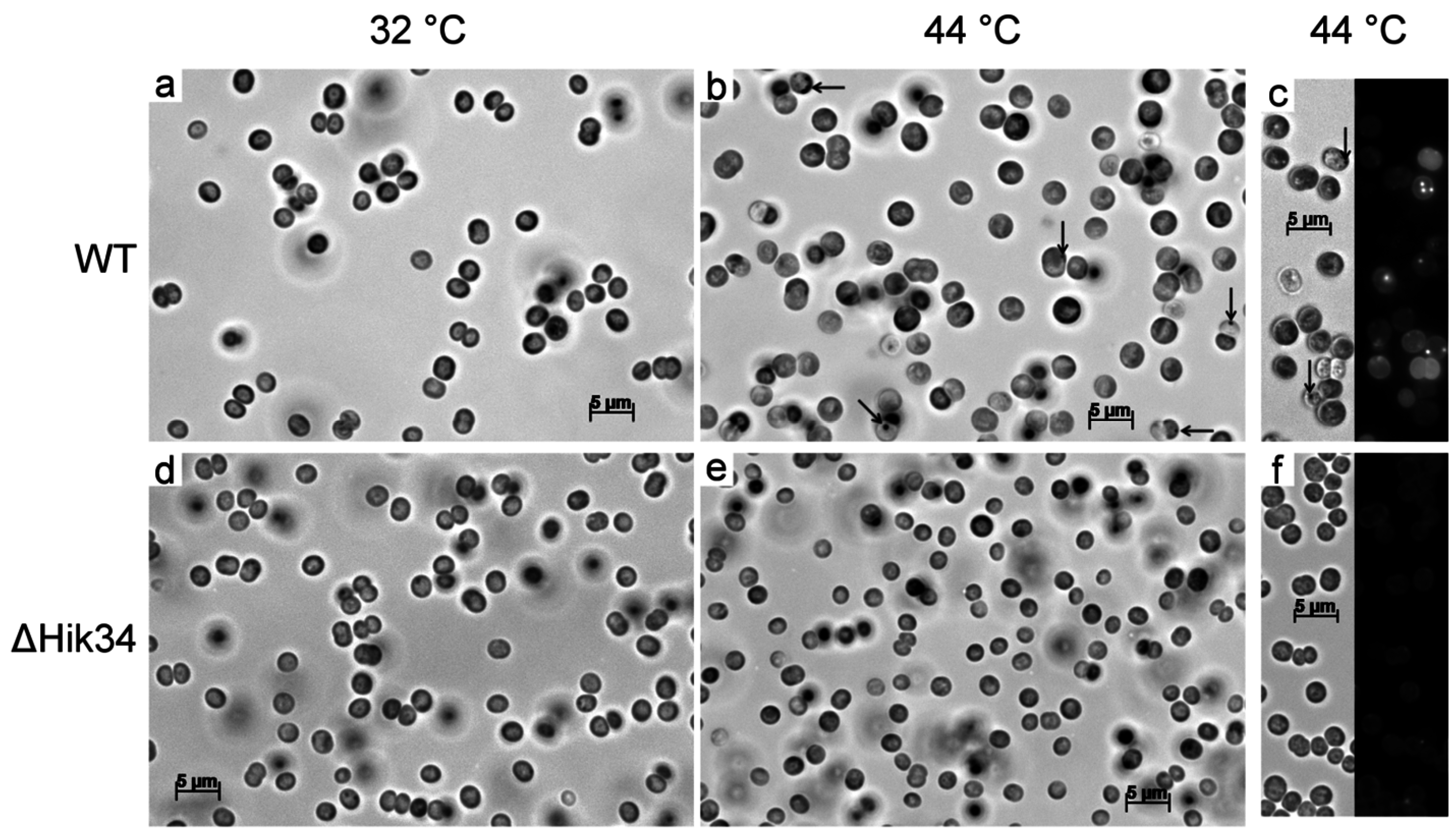
2.6. Light and Fluorescent Microscopy
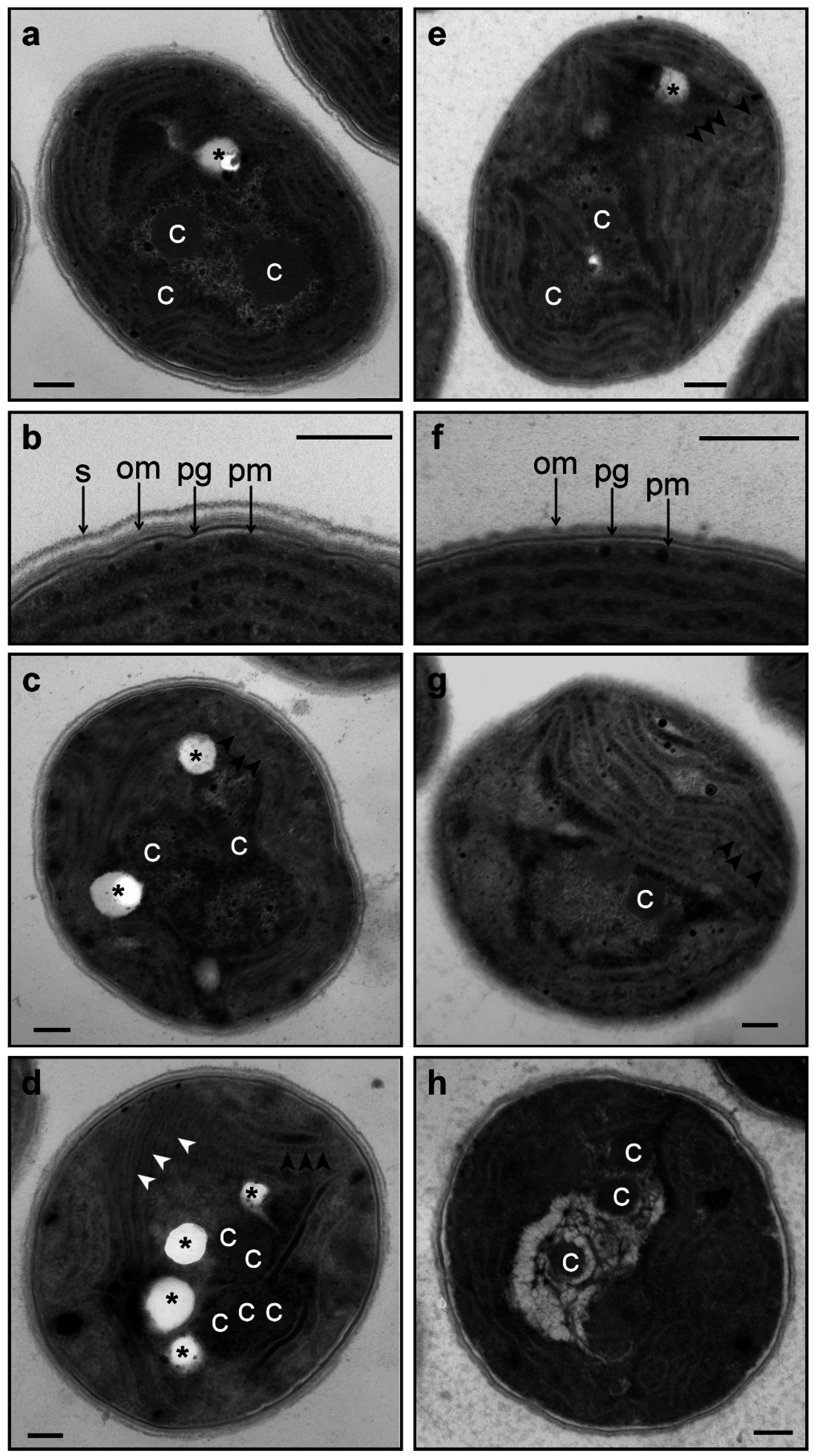
2.7. Electron Microscopy
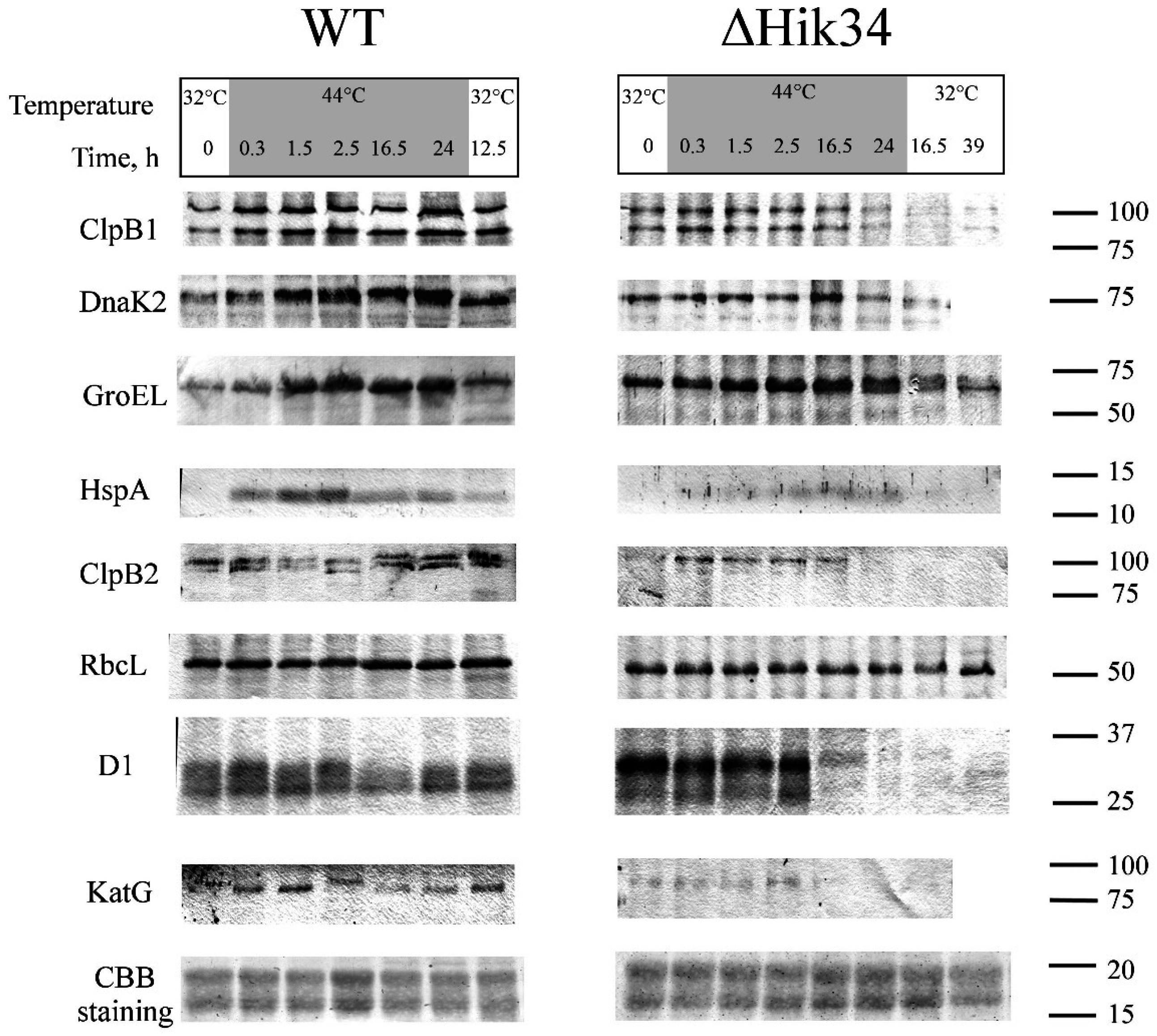
2.8. Protein Isolation and Immunoblotting
3. Results
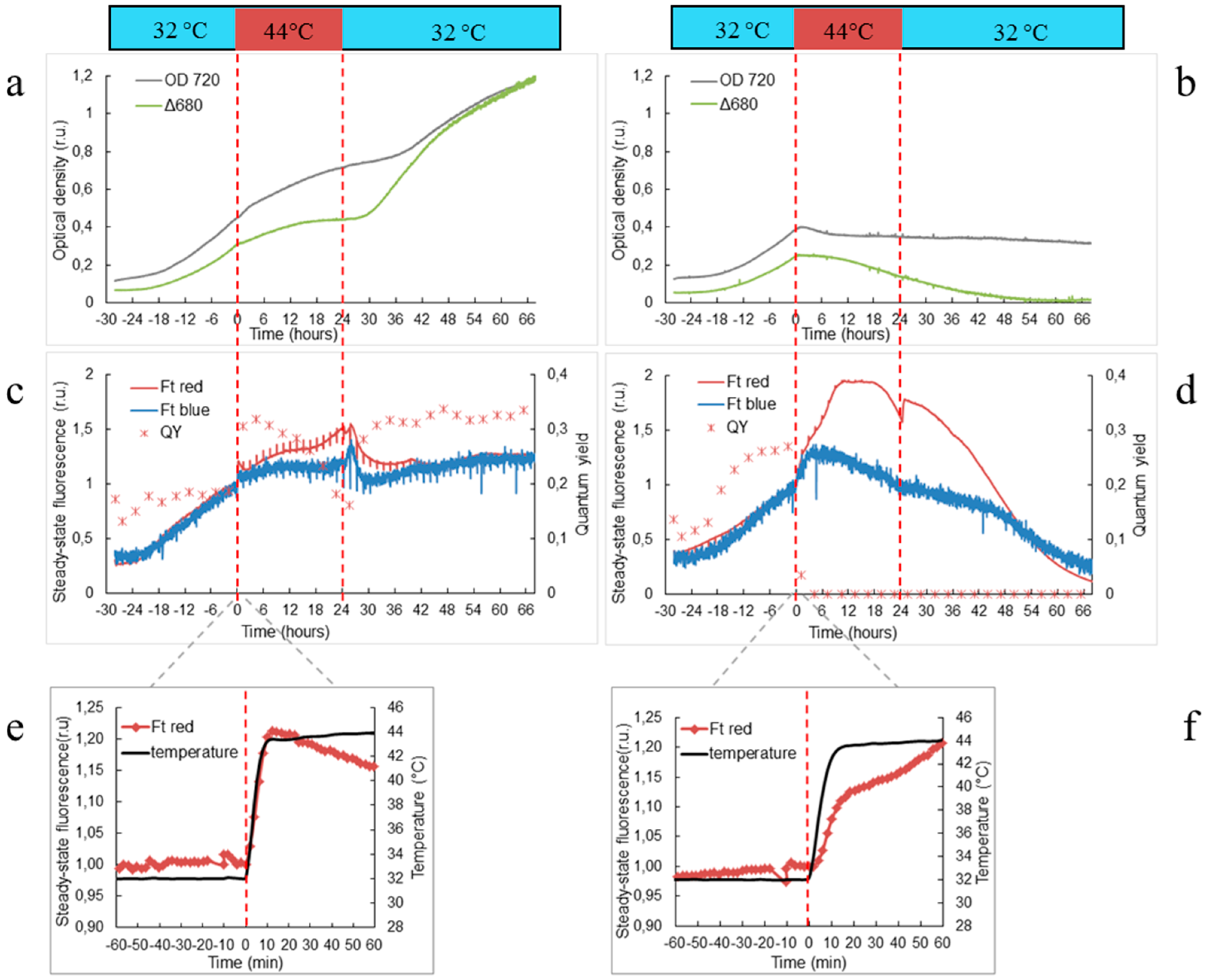
3.1. Physiological Changes in Wild-Type and ΔHik34 Mutant Strains of Synechocystis Subjected to Heat Stress for 24 h
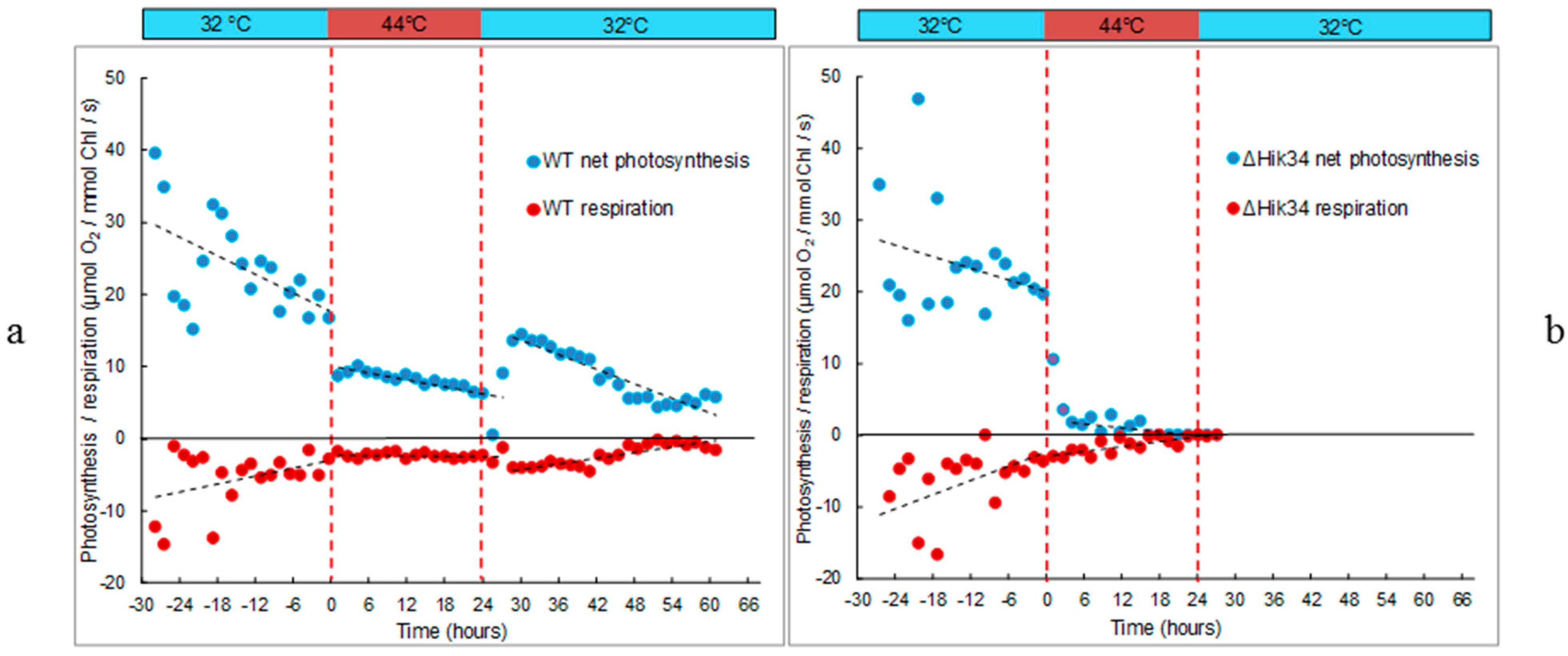
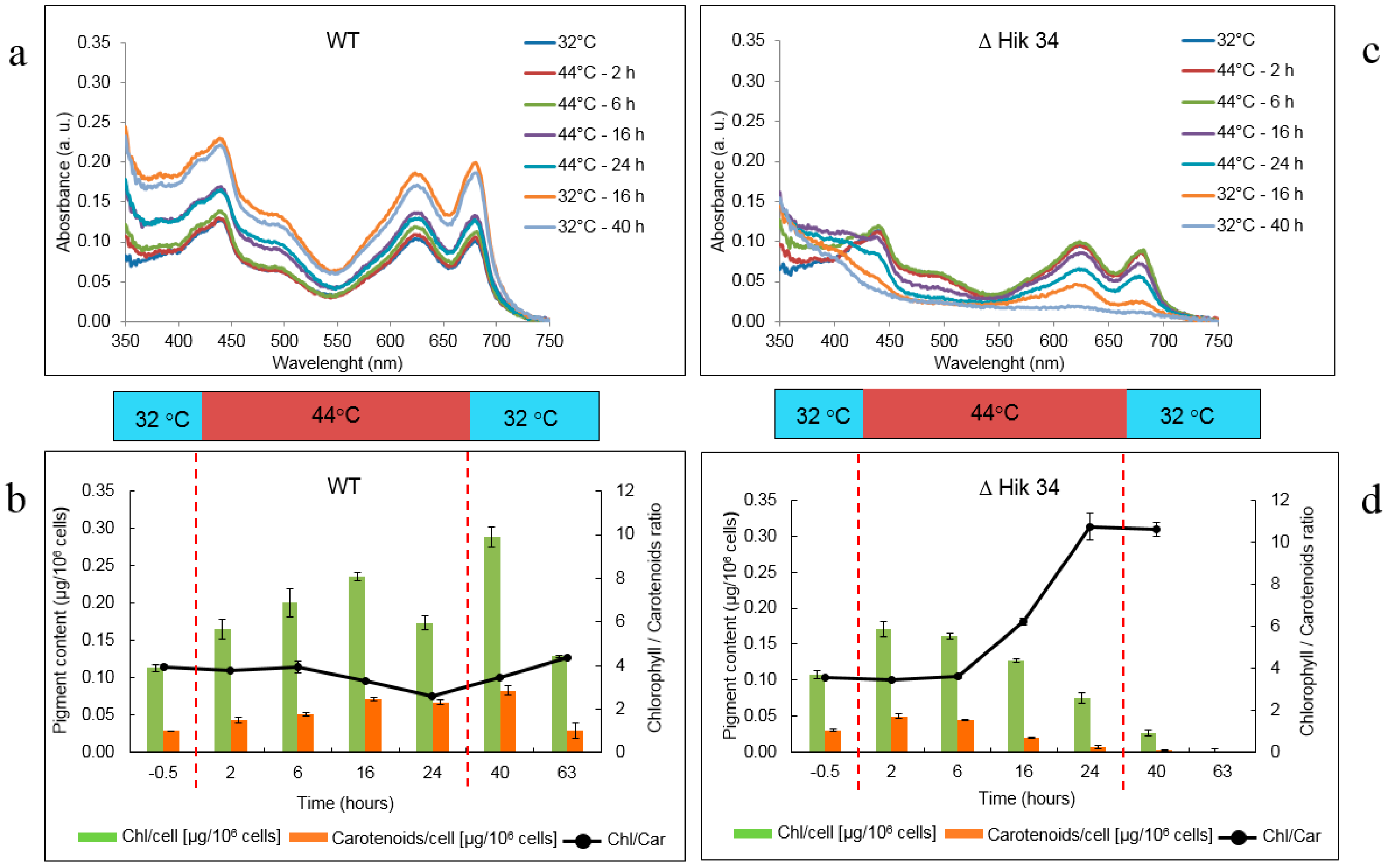
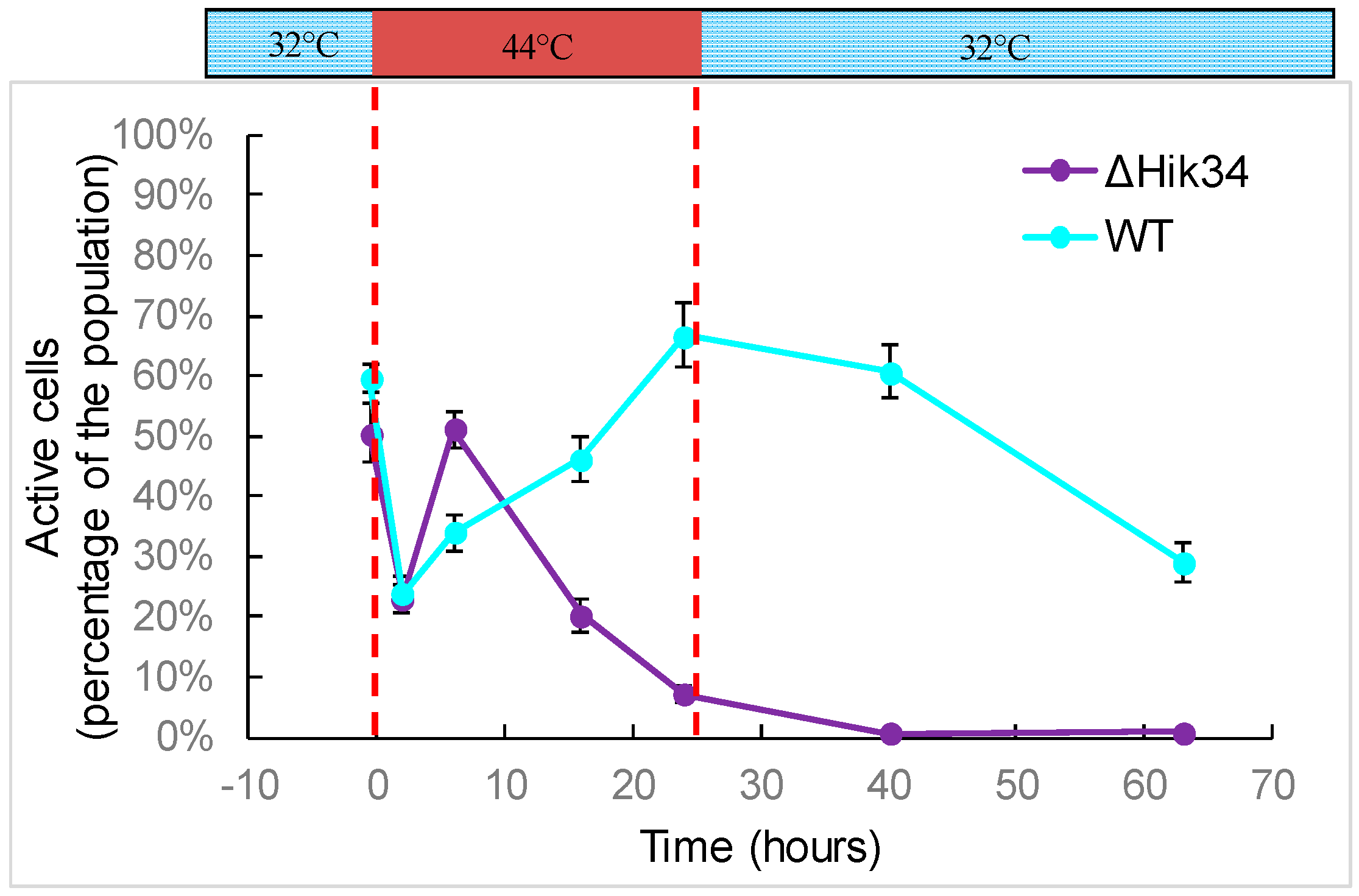
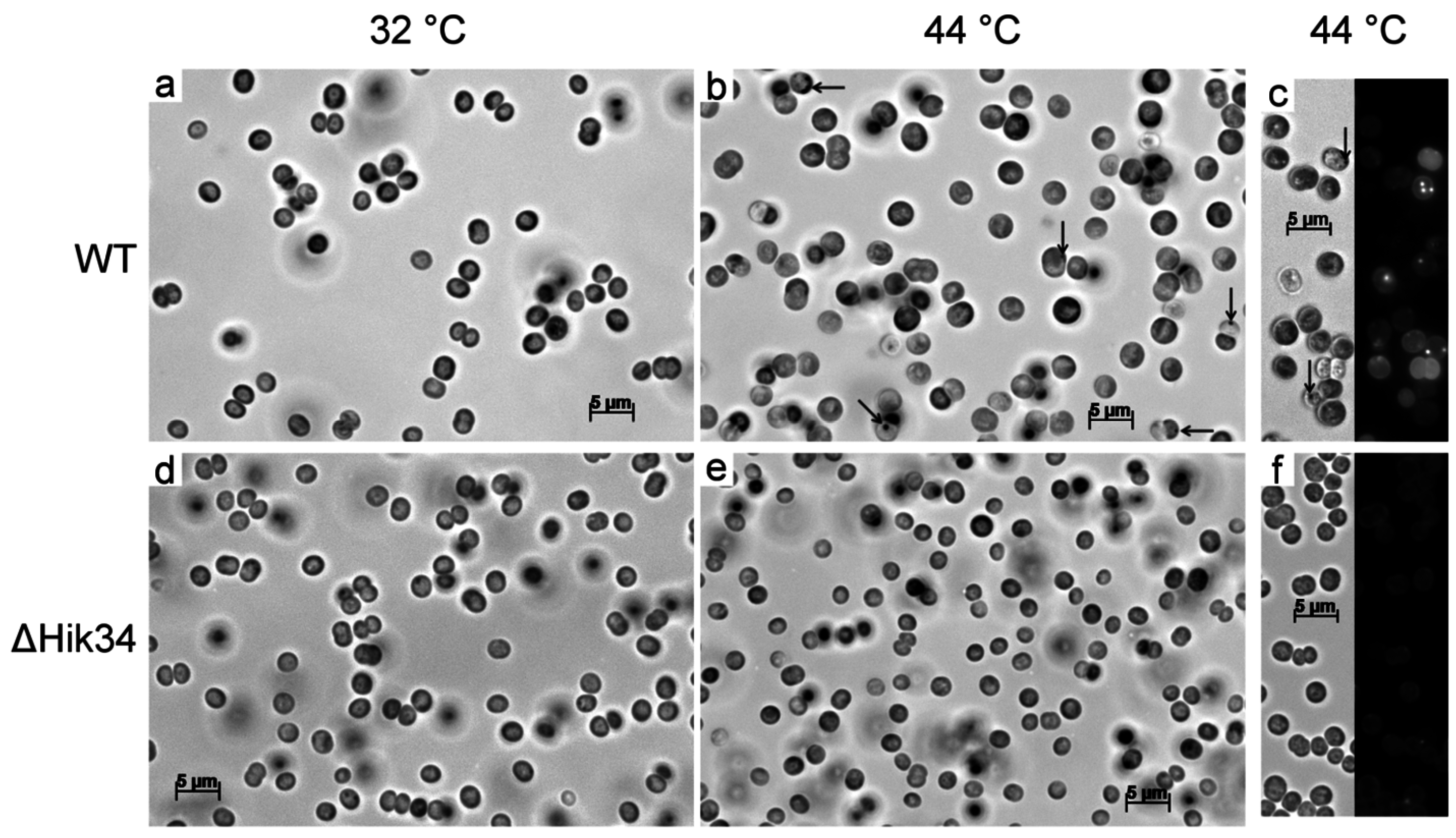
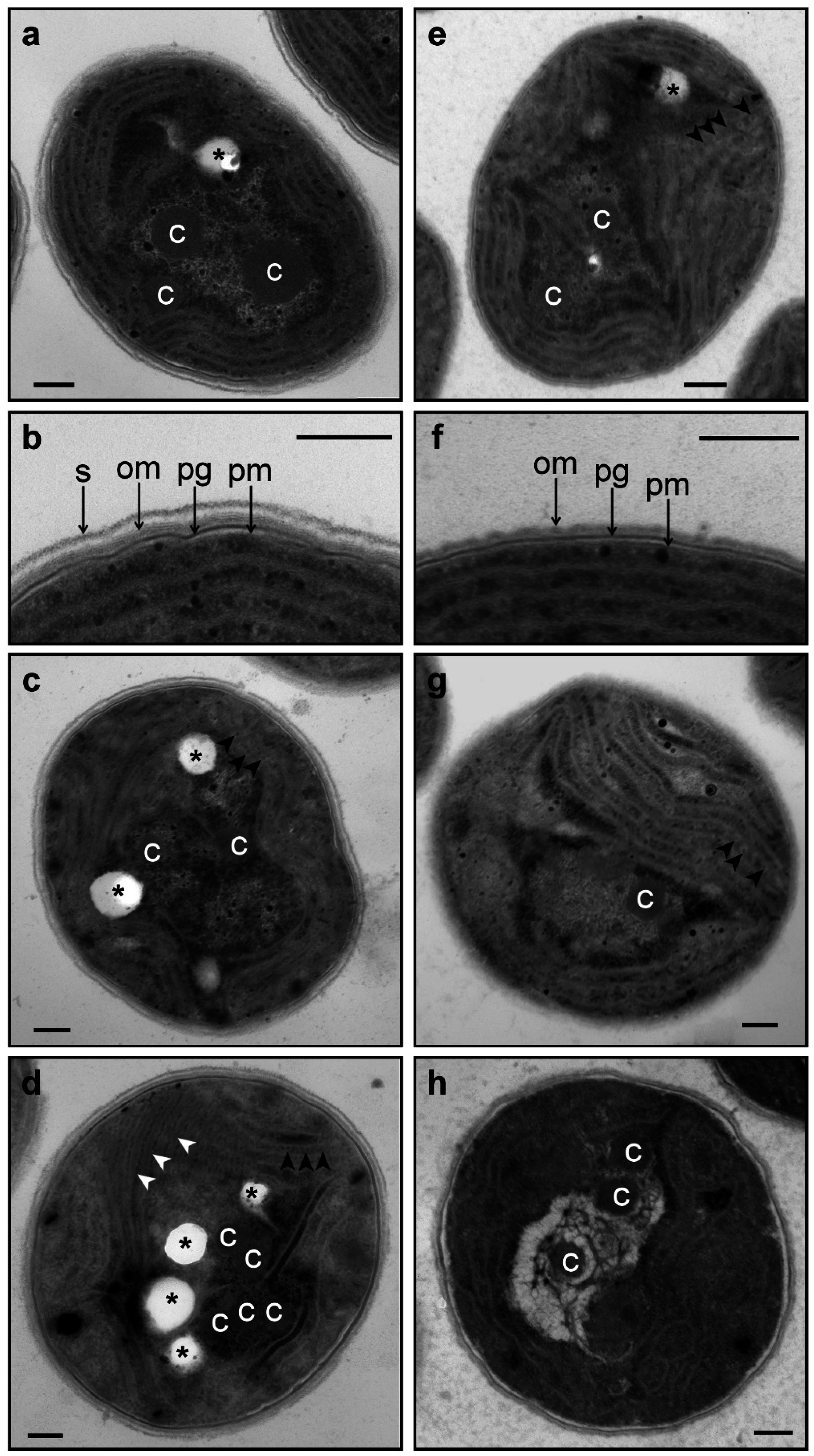
3.2. Changes in Morphology and Ultrastructure of Wild-Type and ΔHik34 Cells During Heat Stress.
- (1)
- Phycobilisomes were usually more structured and thus more apparent in ΔHik34 cells than in WT (Figure 6a,e, Figure S3a–c, S4a–c). In many ΔHik34 cells phycobilisomes formed a hatched pattern between thylakoids. Such hatched pattern was only rarely visible in WT cells under normal conditions, but it became abundant under heat stress (Figure 6c,d).
- (2)
- The outer layer (or s-layer) was apparent in WT cells, but it was absent or significantly reduced in ΔHik34 cells (Figure 6b,f).
- (3)
- After 2 h of heat stress, in WT cells a hatched pattern between thylakoids appeared (Figure 6с, Figure S3d–f), similar to that in ΔHik34 cells under normal conditions (Figure 6e, Figure S4a–c). This pattern was mostly formed by alternating phycobilisomes and electron-dense material between them. Some of WT cells lacked their s-layer (Figure S3d,f). The hatched pattern retained in wild-type cells during the whole heat stress period (Figure 6d, Figure S3g–i). After 24 h of heat stress, the s-layer and the outer membrane were damaged in many cells. In some cells thylakoids became thicker and filled with electron-dense material (Figure 6d, Figure S3h).
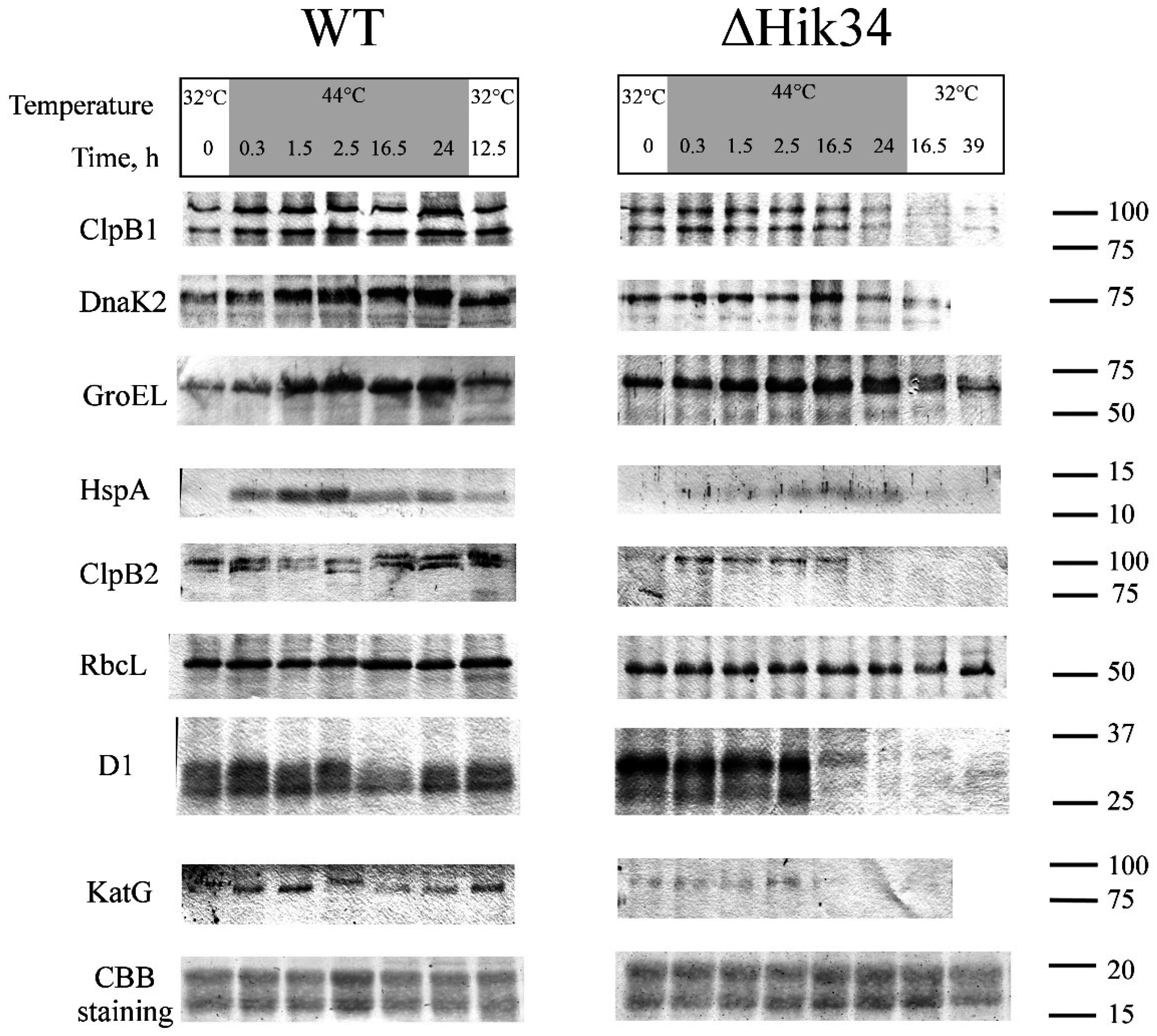
3.3. Western-Blot Analysis of Heat Shock Induced Protein Expression
4. Discussion
4.1. Physiological Changes in WT and ΔHik34 Mutant Strains of Synechocystis Cells Subjected to Heat Stress for 24 Hours
4.2. Morphological and Ultrastructural Changes in WT and ΔHik34 Mutant Strains of Synechocystis Cells Subjected to Heat Stress for 24 Hours
4.3. Protein Analysis
5. Conclusions
Acknowledgments
Author Contributions
Supplementary Materials
Conflicts of Interest
References
- Zavřel, T.; Literáková, P.; Búzová, D.; Sinetova, M.A.; Červený, J. Characterization of a model cyanobacterium Synechocystis sp. PCC 6803 autotrophic growth in a flat-panel photobioreactor. Eng. Life Sci. 2014, 15, 122–132. [Google Scholar] [CrossRef]
- Suzuki, I.; Kanesaki, Y.; Hayashi, H.; Hall, J.J.; Simon, W.J.; Slabas, A.R.; Murata, N. The histidine kinase Hik34 is involved in thermotolerance by regulating the expression of heat shock genes in Synechocystis. Plant Physiol. 2005, 138, 1409–1421. [Google Scholar] [CrossRef] [PubMed]
- Slabas, A.R.; Suzuki, I.; Murata, N.; Simon, W.J.; Hall, J.J. Proteomic analysis of the heat shock response in Synechocystis PCC 6803 and a thermally tolerant knockout strain lacking the histidine kinase 34 gene. Proteomics 2006, 6, 845–864. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Summerfield, T.C.; Li, H.; Sherman, L.A. The heat shock response in the cyanobacterium Synechocystis sp. strain PCC 6803 and regulation of gene expression by HrcA and SigB. Arch Microbiol. 2006, 186, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Tuominen, I.; Pollari, M.; von Wobeser, A.E.; Tyystjarvi, E.; Ibelings, B.W.; Matthijs, H.C.; Tyystjarvi, T. Sigma factor SigC is required for heat acclimation of the cyanobacterium Synechocystis sp. strain PCC 6803. FEBS Lett. 2008, 582, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Tuominen, I.; Pollari, M.; Tyystjarvi, E.; Tyystjarvi, T. The SigB sigma factor mediates high-temperature responses in the cyanobacterium Synechocystis sp. PCC 6803. FEBS Lett. 2006, 580, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Aminaka, R.; Taira, Y.; Kashino, Y.; Koike, H.; Satoh, K. Acclimation to the growth temperature and thermosensitivity of photosystem II in a mesophilic cyanobacterium, Synechocystis sp. PCC 6803. Plant Cell Physiol. 2006, 47, 1612–1621. [Google Scholar] [CrossRef] [PubMed]
- Murata, N.; Takahashi, S.; Nishiyama, Y.; Allakhverdiev, S.I. Photoinhibition of photosystem II under environmental stress. Biochim. Biophys. Acta 2007, 1767, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Balogi, Z.; Török, Z.; Balogh, G.; Jósvay, K.; Shigapova, N.; Vierling, E.; Vígh, L.; Horváth, I. “Heat shock lipid” in cyanobacteria during heat/light-acclimation. Arch. Biochem. Biophys. 2005, 436, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Taira, Y.; Emi, T.; Yamane, Y.; Kashino, Y.; Koike, H.; Satoh, K. Acclimation to growth temperature and the high-temperature effects on photosystem II and plasma membranes in a mezophilic cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 2001, 42, 1140–1148. [Google Scholar] [CrossRef] [PubMed]
- Török, Z.; Goloubinoff, P.; Horváth, I.; Tsvetkova, N.M.; Glatz, A.; Balogh, G.; Varvasovszki, V.; Los, D.A.; Vierling, E.; Crowe, J.H.; et al. Synechocystis HSP17 is an amphitropic protein that stabilizes heat-stressed membranes and binds denatured proteins for subsequent chaperone-mediated refolding. Proc. Natl. Acad. Sci. USA 2001, 98, 3098–3103. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, H.; Suzuki, M.; Kojima, K. Targeted inactivation of the hrcA repressor gene in cyanobacteria. FEBS Lett. 2003, 549, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Owen, H.A.; Prochaska, D.J.; Barnum, S.R. HSP16.6 is involved in the development of thermotolerance and thylakoid stability in the unicellular cyanobacterium, Synechocystis sp. PCC 6803. Curr. Microbiol. 2000, 40, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Barnum, S.R. The heat shock gene, htpG, and thermotolerance in the cyanobacterium, Synechocystis sp. PCC 6803. Curr. Microbiol. 2003, 47, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarzyk, D.; Anfelt, J.; Sarnegrim, A.; Hudson, E.P. Overexpression of sigma factor SigB improves temperature and butanol tolerance of Synechocystis sp. PCC 6803. J. Biotechnol. 2014, 182–183, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Kim, H.W.; Badalamenti, J.P.; Zhou, C.; Sridharakrishnan, S.; Krajmalnik-Brown, R.; Rittmann, B.E.; Vannela, R. Effects of temperature shifts on growth rate and lipid characteristics of Synechocystis sp. PCC 6803 in a bench-top photobioreactor. Bioresour. Technol. 2011, 102, 11218–11225. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, I.; Simon, W.J.; Slabas, A.R. The heat shock response of Synechocystis sp. PCC 6803 analysed by transcriptomics and proteomics. J. Exp. Bot. 2006, 57, 1573–1578. [Google Scholar] [CrossRef] [PubMed]
- Lehel, C.; Wada, H.; Kovács, E.; Török, Z.; Gombos, Z.; Horváth, I.; Murata, N.; Vigh, L. Heat shock protein synthesis of the cyanobacterium Synechocystis PCC 6803: Purification of the GroEL-related chaperonin. Plant Mol. Biol. 1992, 18, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Krishna, P.S.; Rani, B.R.; Mohan, M.K.; Suzuki, I.; Shivaji, S.; Prakash, J.S. A novel transcriptional regulator, Sll1130, negatively regulates heat-responsive genes in Synechocystis sp. PCC 6803. Biochem. J. 2013, 449, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Kortmann, J.; Sczodrok, S.; Rinnenthal, J.; Schwalbe, H.; Narberhaus, F. Translation on demand by a simple RNA-based thermosensor. Nucleic Acids Res. 2011, 39, 2855–2868. [Google Scholar] [CrossRef] [PubMed]
- Asadulghani; Suzuki, Y.; Nakamoto, H. Light plays a key role in the modulation of heat shock response in the cyanobacterium Synechocystis sp. PCC 6803. Biochem. Biophys. Res. Commun. 2003, 306, 872–879. [Google Scholar] [CrossRef]
- Los, D.A.; Suzuki, I.; Zinchenko, V.V.; Murata, N. Stress responses in Synechocystis: Regulated genes and regulatory systems. In The Cyanobacteria: Molecular Biology, Genomics and Evolution; Herrero, A., Flores, E., Eds.; Caister Academic Press: Norfolk, UK, 2008; pp. 117–157. [Google Scholar]
- Murata, N.; Suzuki, I. Exploitation of genomic sequences in a systematic analysis to access how cyanobacteria sense environmental stress. J. Exp. Bot. 2006, 57, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Ashby, M.K.; Houmard, J. Cyanobacterial two-component proteins: Structure, diversity, distribution, and evolution. Microbiol. Mol. Biol. Rev. 2006, 70, 472–509. [Google Scholar] [CrossRef] [PubMed]
- Marin, K.; Suzuki, I.; Yamaguchi, K.; Ribbeck, K.; Yamamoto, H.; Kanesaki, Y.; Hagemann, M.; Murata, N. Identification of histidine kinases that act as sensors in the perception of salt stress in Synechocystis sp. PCC 6803. Proc. Natl. Acad. Sci. USA 2003, 100, 9061–9066. [Google Scholar] [CrossRef] [PubMed]
- Hihara, Y.; Sonoike, K.; Kanehisa, M.; Ikeuchi, M. DNA microarray analysis of redox-responsive genes in the genome of the cyanobacterium Synechocystis sp. Strain PCC 6803. J. Bacteriol. 2003, 185, 1719–1725. [Google Scholar] [CrossRef] [PubMed]
- Paithoonrangsarid, K.; Shumskaya, M.A.; Kanesaki, Y.; Satoh, S.; Tabata, S.; Los, D.A.; Zinchenko, V.V.; Hayashi, H.; Tanticharoen, M.; Suzuki, I.; et al. Five histidine kinases perceive osmotic stress and regulate distinct sets of genes in Synechocystis. J. Biol. Chem. 2004, 279, 53078–53086. [Google Scholar] [CrossRef]
- Shumskaya, M.A.; Paithoonrangsarid, K.; Kanesaki, Y.; Los, D.A.; Zinchenko, V.V.; Tanticharoen, M.; Suzuki, I.; Murata, N. Identical Hik-Rre systems are involved in perception and transduction of salt signals and hyperosmotic signals but regulate the expression of individual genes to different extents in Synechocystis. J. Biol. Chem. 2005, 280, 21531–21538. [Google Scholar] [CrossRef] [PubMed]
- Kanesaki, Y.; Yamamoto, H.; Paithoonrangsarid, K.; Shumskaya, M.; Suzuki, I.; Hayashi, H.; Murata, N. Histidine kinases play important roles in the perception and signal transduction of hydrogen peroxide in the cyanobacterium, Synechocystis sp. PCC 6803. Plant J. 2007, 49, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Rowland, J.G.; Pang, X.; Suzuki, I.; Murata, N.; Simon, W.J.; Slabas, A.R. Identification of components associated with thermal acclimation of photosystem II in Synechocystis sp. PCC 6803. PLoS One 2010, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [PubMed]
- Nedbal, L.; Trtílek, M.; Červený, J.; Komárek, O.; Pakrasi, H.B. A photobioreactor system for precision cultivation of photoautotrophic microorganisms and for high-content analysis of suspension dynamics. Biotechnol. Bioeng. 2008, 100, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Sinetova, M.A.; Červený, J.; Zavřel, T.; Nedbal, L. On the dynamics and constraints of batch culture growth of the cyanobacterium Cyanothece sp. ATCC 51142. J. Biotechnol. 2012, 162, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Markelova, A.G.; Vladimirova, M.G.; Kuptsova, E.S. A comparison of cytochemical methods for the rapid evaluation of microalgal viability. Russ. J. Plant Physiol. 2000, 47, 815–819. [Google Scholar] [CrossRef]
- Guilbault, G.G.; Kramer, D.N. Fluorometric determination of lipase, acylase, alpha-, and gamma-chymotrypsin and inhibitors of these enzymes. Anal. Chem. 1964, 36, 409–412. [Google Scholar] [CrossRef]
- Reynolds, E.S. The use of lead citrate at high pH as an electronopaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sherman, L.A. A redox-responsive regulator of photosynthesis gene expression in the cyanobacterium Synechocystis sp. strain PCC 6803. J. Bacteriol. 2000, 182, 4268–4277. [Google Scholar] [CrossRef] [PubMed]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat stress: An overview of molecular responses in photosynthesis. Photosynth. Res. 2008, 98, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Emi, T.; Yamane, Y.; Kashino, Y.; Koike, H.; Satoh, K. Effects of high-temperature treatments on a thermophilic cyanobacterium Synechococcus vulcanus. Plant Cell Physiol. 2000, 41, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Stoitchkova, K.; Zsiros, O.; Jávorfi, T.; Páli, T.; Andreeva, A.; Gombos, Z.; Garab, G. Heat- and light-induced reorganizations in the phycobilisome antenna of Synechocystis sp. PCC 6803. Thermo-optic effect. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 750–756. [Google Scholar] [CrossRef]
- Horvath, I.; Glatz, A.; Nakamoto, H.; Mishkind, M.L.; Munnik, T.; Saidi, Y.; Goloubinoff, P.; Harwood, J.L.; Vigh, L. Heat shock response in photosynthetic organisms: Membrane and lipid connections. Prog. Lipid Res. 2012, 51, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Van de Meene, A.L.; Hohmann-Marriott, M.; Vermaas, W.J.; Roberson, R. The three-dimensional structure of the cyanobacterium Synechocystis sp. PCC 6803. Arch. Microbiol. 2006, 184, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Trautner, C.; Vermaas, W.F. The sll1951 gene encodes the surface layer protein of Synechocystis sp. strain PCC 6803. J. Bacteriol. 2013, 195, 5370–5380. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.J. Proteomic Analysis of the Heat Shock and Acclimation Responses of Cyanobacteria. Ph.D. Thesis, Durham University, Durham, UK, 2005. [Google Scholar]
- Sakthivel, K.; Watanabe, T.; Nakamoto, H. A small heat-shock protein confers stress tolerance and stabilizes thylakoid membrane proteins in cyanobacteria under oxidative stress. Arch. Microbiol. 2009, 191, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Prochaska, D.J.; Fang, F.; Barnum, S.R. A 16.6-kilodalton protein in the cyanobacterium Synechocystis sp. PCC 6803 plays a role in the heat shock response. Curr. Microbiol. 1998, 37, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Varvasovszki, V.; Glatz, A.; Shigapova, N.; Jósvay, K.; Vı́gh, L.; Horváth, I. Only one dnaK homolog, dnaK2, is active transcriptionally and is essential in Synechocystis. Biochem. Biophys. Res. Commun. 2003, 305, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Kovács, E.; van der Vies, S.M.; Glatz, A.; Török, Z.; Varvasovszki, V.; Horváth, I.; Vígh, L. The chaperonins of Synechocystis PCC 6803 differ in heat inducibility and chaperone activity. Biochem. Biophys. Res. Commun. 2001, 289, 908–915. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.; Pisareva, T.; Norling, B. Proteomic studies of the thylakoid membrane of Synechocystis sp. PCC 6803. Proteomics 2005, 5, 4905–4916. [Google Scholar] [CrossRef] [PubMed]
- Tsvetkova, N.M.; Horvath, I.; Torok, Z.; Wolkers, W.F.; Balogi, Z.; Shigapova, N.; Crowe, L.M.; Tablin, F.; Vierling, E.; Crowe, J.H.; et al. Small heat-shock proteins regulate membrane lipid polymorphism. Proc. Natl. Acad. Sci. USA 2002, 99, 13504–13509. [Google Scholar] [CrossRef] [PubMed]
- Nitta, K.; Suzuki, N.; Honma, D.; Kaneko, Y.; Nakamoto, H. Ultrastructural stability under high temperature or intensive light stress conferred by a small heat shock protein in cyanobacteria. FEBS Lett. 2005, 579, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, E.; Gathmann, S.; Fuhrmann, E.; Schneider, D. Three different DnaK proteins are functionally expressed in the cyanobacterium Synechocystis sp. PCC 6803. Microbiology 2007, 153, 1828–1841. [Google Scholar] [CrossRef] [PubMed]
- Asadulghani; Nitta, K.; Kaneko, Y.; Kojima, K.; Fukuzawa, H.; Kosaka, H.; Nakamoto, H. Comparative analysis of the hspA mutant and wild-type Synechocystis sp. strain PCC 6803 under salt stress: Evaluation of the role of hspA in salt-stress management. Arch. Microbiol. 2004, 182, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Mansilla, M.; Banchio, C.; de Mendoza, D. Signalling pathways controlling fatty acid desaturation. In Lipids in Health and Disease; Quinn, P., Wang, X., Eds.; Springer: Dordrecht, the Netherlands, 2008; Volume 49, pp. 71–99. [Google Scholar]
- Sakayori, T.; Shiraiwa, Y.; Suzuki, I. A Synechocystis homolog of SipA protein, Ssl3451, enhances the activity of the histidine kinase Hik33. Plant Cell Physiol. 2009, 50, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Červený, J.; Sinetova, M.A.; Zavřel, T.; Los, D.A. Mechanisms of High Temperature Resistance of Synechocystis sp. PCC 6803: An Impact of Histidine Kinase 34. Life 2015, 5, 676-699. https://doi.org/10.3390/life5010676
Červený J, Sinetova MA, Zavřel T, Los DA. Mechanisms of High Temperature Resistance of Synechocystis sp. PCC 6803: An Impact of Histidine Kinase 34. Life. 2015; 5(1):676-699. https://doi.org/10.3390/life5010676
Chicago/Turabian StyleČervený, Jan, Maria A. Sinetova, Tomáš Zavřel, and Dmitry A. Los. 2015. "Mechanisms of High Temperature Resistance of Synechocystis sp. PCC 6803: An Impact of Histidine Kinase 34" Life 5, no. 1: 676-699. https://doi.org/10.3390/life5010676