Exploring Linkages between Supporting, Regulating, and Provisioning Ecosystem Services in Rangelands in a Tropical Agro-Forest Frontier


 ,
,
Abstract
:1. Introduction
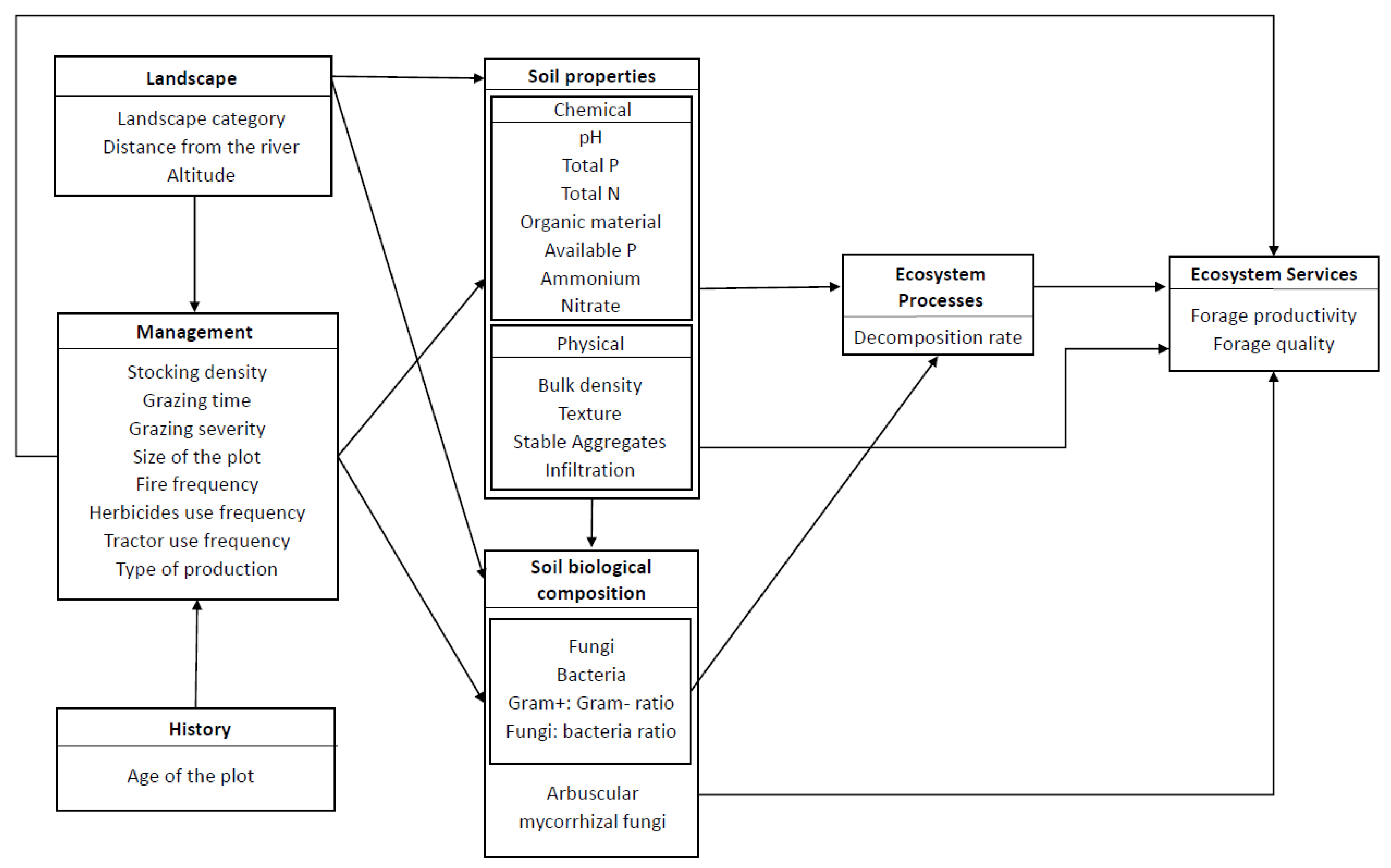
2. Materials and Methods
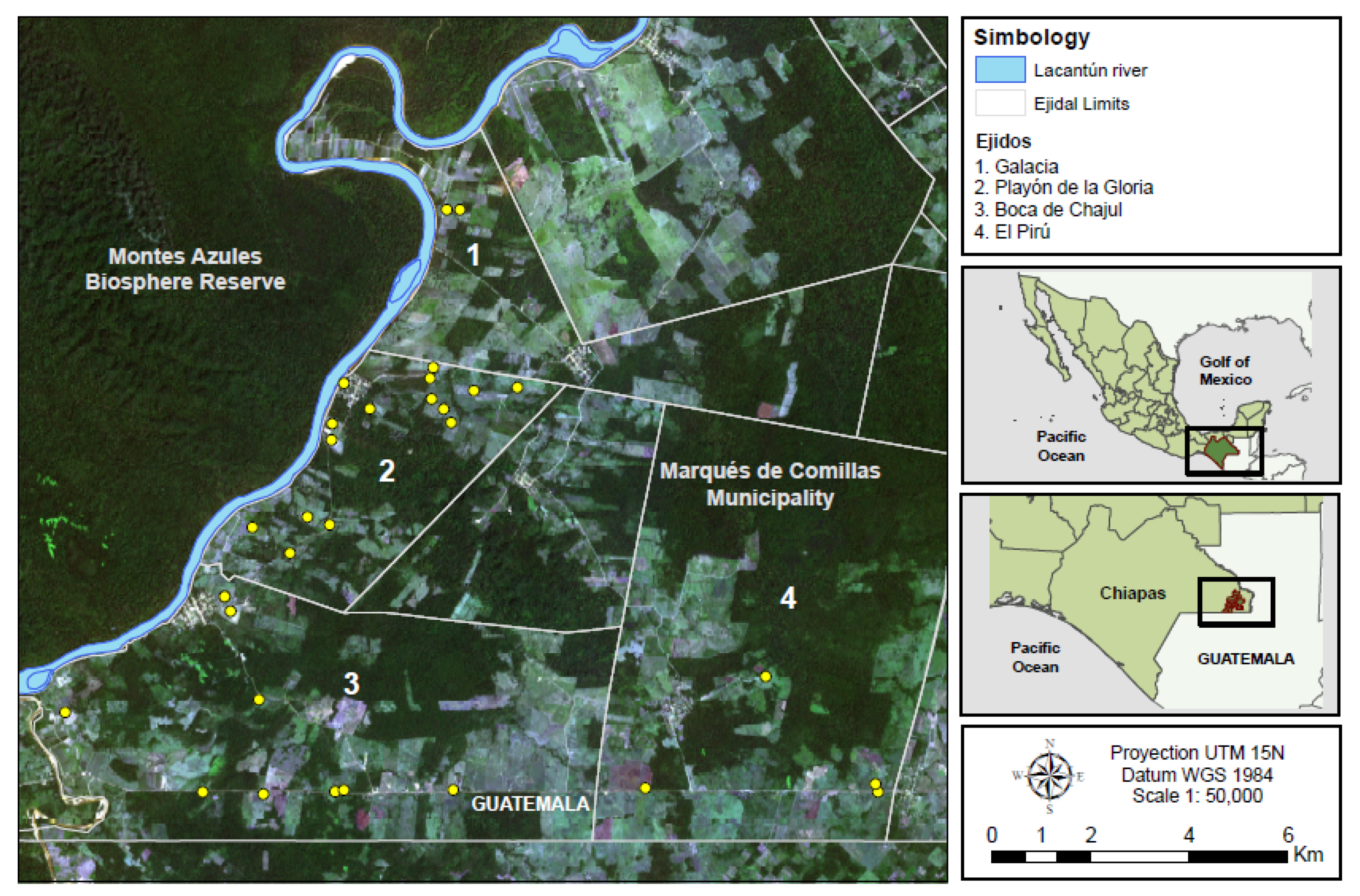
2.1. Study Site
2.2. Study Design, Plot Establishment and Plot Characteristics
2.3. Data Collection
2.3.1. Characterization of Livestock Management Practices
2.3.2. Soil Quality and Litter Decomposition
2.3.3. Biomass Production and Forage Quality
2.4. Data Analyses
3. Results
3.1. Rangeland Characteristics and Management
3.2. Soil Quality and Biomass Productivity
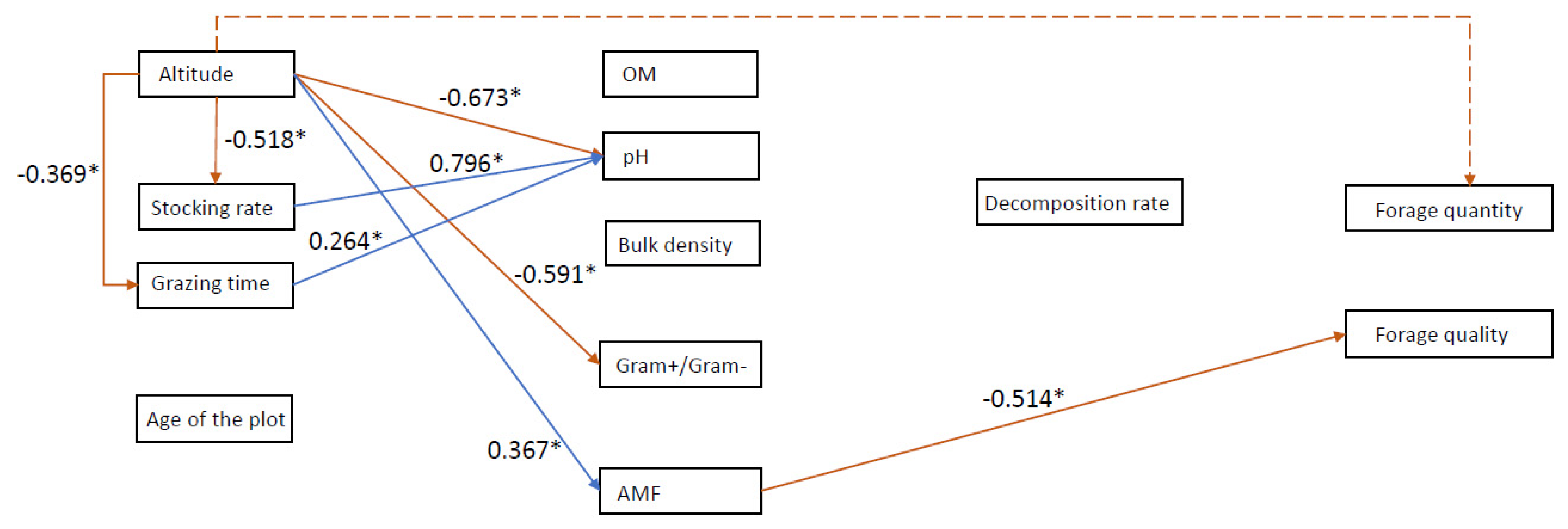
3.3. Effects of Management on Soil Quality and Ecosystem Services
4. Discussion
4.1. Landscape Position Drives Farmers Management Practices, Soil Quality and Forage Quantity but Not Quality
4.2. Soil Properties Differed Widely with Landscape Position and among Plots
4.3. Land Use History Was Not Related to Actual Management Practices and Soil Quality Indicators
4.4. Implications and Recommendations for Sustainable Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Questions to Guide Semi-Structure Interviews
- Interviewee general data
- 1.1.
- Name
- 1.2.
- Age
- 1.3.
- Town of origin
- 1.4.
- Principal productive activities
- Sketch of farm (rangeland). Indicate: (a) dimensions, (b) fences, (c) rotational system, (d) rivers, streams and other sources of water, (e) exclusion experiment, (f) outstanding trees, (g) average head of cattle present in each section of the farm, (h) years of establishment of the grassland.
- History of the farm (reconstruct land-use history)
- 3.1.
- Are you the first owner of the farm? When you became the owner, what was the existing land-use?
- 3.2.
- When was the rainforest transformed for the first time? How did you induce the grassland establishment? (Record land-use trajectory. For example: rainforest-> cropland-> grassland)
- 3.3.
- After slash-burn process, did you induce the establishment of native or commercial grasses? How? (For example, sowing, transplant of seedlings, wait for natural colonization)
- 3.4.
- When did you introduced cattle for the first time?
- 3.5.
- Does the rangeland has had resting periods? (time without cattle) Why? How many? How long?
- 3.6.
- Do you use fire? How many times have you burned the field? Why?
- 3.7.
- Do you apply agrochemicals? Which ones? When did you start using them? (Inquire about frequency and doses).
- 3.8.
- What type of cattle production do you carry out? (meat production, calf production, mixed). Why?
- 3.9.
- How many heads of cattle do you have in the rangeland we are talking about? (Inquire about the current and the last production cycle. Specify number of: cows, bulls, calves and steers).
- 3.10.
- What is the maximum/minimum number of heads of cattle you have had in this rangeland? When was that?
- 3.11.
- Calves production: In average, how many calves do you sell per productive cycle? (Specify average weigh).Meat production: How many calves do you fatten per year? What are the average initial and final weigh?
- 3.12.
- Do you sell “old cows”? How many per productive cycle? (Specify average weigh). Do you replace them?
- 3.13.
- Rotational system: Inquire how many sections exist in the rangeland and how many days the cows graze in each section. Be emphatic about the section where the exclosure was set up.
- 3.14.
- What type of grasses (species) are present in the rangeland? Describe the succession of grasses in the history of the grassland.
- 3.15.
- Do you feed the cattle with complementary food? (pollinasa, maize, salt). Specify quantities.
- 3.16.
- Have you experience any trouble related with the productivity of the grassland? (Inquire about non-palatable species or perceptions related with soil characteristics)
References
- Arroyo-Rodríguez, V.; Melo, F.P.L.; Martínez-Ramos, M.; Bongers, F.; Chazdon, R.L.; Meave, J.A.; Norden, N.; Santos, B.A.; Leal, I.R.; Tabarelli, M. Multiple successional pathways in human-modified tropical landscapes: New insights from forest succession, forest fragmentation and landscape ecology research. Biol. Rev. 2017, 92, 326–340. [Google Scholar] [CrossRef] [PubMed]
- Quezada, M.L.; Arroyo-Rodríguez, V.; Pérez-Silva, E.; Aide, T.M. Land cover changes in the Lachuá region, Guatemala: Patterns, proximate causes, and underlying driving forces over the last 50 years. Reg. Environ. Chang. 2014, 14, 1139–1149. [Google Scholar] [CrossRef]
- Murgueitio, E.; Calle, Z.; Uribe, F.; Calle, A.; Solorio, B. Native trees and shrubs for the productive rehabilitation of tropical cattle ranching lands. For. Ecol. Manag. 2011, 261, 1654–1663. [Google Scholar] [CrossRef]
- Díaz, S.; Pascual, U.; Stenseke, M.; Martín-López, B.; Watson, R.T.; Molnár, Z.; Hill, R.; Chan, K.M.A.; Baste, I.A.; Brauman, K.A.; et al. Assessing nature’s contributions to people. Science 2018, 359, 270–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millennium Ecosystem Assessment (MEA). Ecosystems and Human Well-Being: Synthesis; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Pascual, U.; Balvanera, P.; Díaz, S.; Pataki, G.; Roth, E.; Stenseke, M.; Watson, R.T.; Başak Dessane, E.; Islar, M.; Kelemen, E.; et al. Valuing nature’s contributions to people: The IPBES approach. Curr. Opin. Environ. Sustain. 2017, 26, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Carabias, J.; Gonzalez-Ortiz, M.A.; Auzias, C.; Castro-Tovar, E.; Acevedo, M.A. Ordenamiento Comunitario del Territorio de la Microregion Marqués de Comillas: Una Iniciativa Interejidal para el Mejoramiento de los Medios de Vida Rurales en la Selva Lacandona; Grupo Autónomo para la Investigación Ambiental, A.C., Natura y Ecosistemas Mexicanos, A.C.: Mexico City, México, 2012. [Google Scholar]
- Heinze, A.; Bongers, F.; Ramírez Marcial, N.; García Barrios, L.; Kuyper, T.W. The montane multifunctional landscape: How stakeholders in a biosphere reserve derive benefits and address trade-offs in ecosystem service supply. Ecosyst. Serv. 2020, 44. [Google Scholar] [CrossRef]
- Aguilar-Fernández, R. Characterizing land use management and promoting recovery of ecosystem services in agro-forested landscapes. Unpublished, manuscript in preparation.
- Mora, C.A.; Pérez, J.; Rodríguez, E.; Alonso, E.; Camacho, R.; Israel, J.; Aurelio, M.; Tagle, G. Effect of Livestock in the Composition and Diversity of Trees and Shrubs in the Tamaulipan Thornscrub. Rev. Mex. Cienc. For. 2013, 4, 124–137. [Google Scholar]
- Fernandez, A.; Torres, A.; Nava, A.D.; Rosas, A.; Muñoz, C.; Rodríguez, C.; Cortinas, C.; Rothman, D.; Quadri, G.; Bocco, G.; et al. Perspectivas del Medio Ambiente en México; Semarnat: Mexico City, México, 2004; ISBN 968-817-683-4.
- Castillo Santiago, M.A. Análisis con Imágenes Satelitales de los Recursos Forestales en el Trópico Húmedo de Chiapas: Un Estudio de Caso en Marqués de Comillas. Ph.D. Thesis, Universidad Nacional Autónoma de México, Mexico City, México, June 2009. [Google Scholar]
- Swanepoel, F.; Stroebel, A.; Moyo, S. The Role of Livestock in Developing Communities: Enhancing Multifunctionality; The Technical Centre for Agricultural and Rural Cooperation (CTA): Bloemfontein, South Africa, 2010; ISBN 978-0-86886-798-4. [Google Scholar]
- Cano-Castellanos, J.I. De Montana a ‘Reserva Forestal’: Colonización, Sentido de Comunidad, y Producción de la Conservación Ecológica en el Sureste de la Selva Lacandona, México. Ph.D. Thesis, Centro de Investigaciones y Estudios Superiores en Antropología Social, Guadalajara, Jalisco, México, September 2013. [Google Scholar]
- Báez Montoya, J.R.; Gutiérrez Perez, I.G. Descripción y costos de la ganadería para engorda a media ceba en esquemas convencionales y silvopastoril en Marqués de Comillas. Unpublished work, manuscript in preparation.
- Suazo-Ortuño, I.; Lopez-Toledo, L.; Alvarado-Díaz, J.; Martínez-Ramos, M. Land-use Change Dynamics, Soil Type and Species Forming Mono-dominant Patches: The Case of Pteridium aquilinum in a Neotropical Rain Forest Region. Biotropica 2015, 47, 18–26. [Google Scholar] [CrossRef]
- Reiners, W.A.; Bouwman, A.F.; Parsons, W.F.J.; Keller, M. Tropical rain forest conversion to pasture: Changes in vegetation and soil properties. Ecol. Appl. 1994, 4, 363–377. [Google Scholar] [CrossRef]
- Mueller, R.C.; Rodrigues, J.L.M.; Nüsslein, K.; Bohannan, B.J.M. Land use change in the Amazon rain forest favours generalist fungi. Funct. Ecol. 2016, 30, 1845–1853. [Google Scholar] [CrossRef] [Green Version]
- Flores-Rentería, D.; Sánchez-Gallén, I.; Morales-Rojas, D.; Larsen, J.; Álvarez-Sánchez, J. Changes in the Abundance and Composition of a Microbial Community Associated with Land Use Change in a Mexican Tropical Rain Forest. J. Soil Sci. Plant Nutr. 2020, 20, 1144–1155. [Google Scholar] [CrossRef]
- Martínez-Ramos, M.; Anten, N.P.R.; Ackerly, D.D. Defoliation and ENSO effects on vital rates of an understory tropical rain forest palm. J. Ecol. 2009, 97, 1050–1061. [Google Scholar] [CrossRef]
- Navarrete-Segueda, A.; Martínez-Ramos, M.; Ibarra-Manríquez, G.; Vázquez-Selem, L.; Siebe, C. Variation of main terrestrial carbon stocks at the landscape-scale are shaped by soil in a tropical rainforest. Geoderma 2018, 313, 57–68. [Google Scholar] [CrossRef]
- Navarrete-Segueda, A.; Vázquez-Selem, L.; Siebe-Grabach, C.D. Caracterización geopedológica y calidad de sitio de una selva tropical. In Conservación y Desarrollo Sustentable en la Selva Lacandona: 25 años de Actividades y Experiencias; Carabias, J., de la Maza, J., Cadena, R., Eds.; Natura y Ecosistemas Mexicanos, A.C.: Mexico City, México, 2015; pp. 87–96. [Google Scholar]
- Méndez, M. El Ejido y la Comunidad en el México del Siglo XXI: La Transición Agraria (1992–2015), 1st ed.; Porrúa: Mexico City, México, 2016; ISBN 6070922352. [Google Scholar]
- Izquierdo-Tort, S.; Ortiz-Rosas, F.; Vázquez-Cisneros, P.A. ‘Partial’ participation in Payments for Environmental Services (PES): Land enrolment and forest loss in the Mexican Lacandona Rainforest. Land Use Policy 2019, 87, 103950. [Google Scholar] [CrossRef]
- Obregón, R. Elaboración de Proyectos Para el Fortalecimiento de la Agricultura Sustentable en los Corredores de Chiapas; Conabio: Mexico City, México, 2007.
- Briske, D.D.; Derner, J.D.; Brown, J.R.; Fuhlendorf, S.D.; Teague, W.R.; Havstad, K.M.; Gillen, R.L.; Ash, A.J.; Willms, W.D. Rotational grazing on rangelands: Reconciliation of perception and experimental evidence. Rangel. Ecol. Manag. 2008, 61, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Zermeño-Hernández, I.; Méndez-Toribio, M.; Siebe, C.; Benítez-Malvido, J.; Martínez-Ramos, M. Ecological disturbance regimes caused by agricultural land uses and their effects on tropical forest regeneration. Appl. Veg. Sci. 2015, 18, 443–455. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An Examination of Degtjareff Method for Determining Soil Organic Matter and a Proposed Modification of the Chromic Acid Titration Method. Soil Sci. 1934, 37, 29–37. [Google Scholar] [CrossRef]
- Bremner, J. Nitrogen. In Methods of Soil Analysis, Part 3-Chemical Methods; Sparks, D., Page, A., Helmke, P., Loeppert, R., Soltanpour, P., Tabatabai, M., Johnston, C., Sumner, M., Eds.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Murphy, J.; Riley, J. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Harwood, J.E.; Huyser, D.J. Some aspects of the phenol-hypoclorite reaction as applied to ammonia analysis. Water Res. 1970, 4, 501–515. [Google Scholar] [CrossRef]
- Robertson, G.; Coleman, D.; Bledsoe, C.; Sollins, P. Standard Soil Methods for Long-Term Ecological Research; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Kroetsch, D.; Wang, C. Particle Size Distribution, 2nd ed.; Taylor and Francis Group, LLC: Oxfordshire, UK, 2006. [Google Scholar]
- Hallett, P.D.; Feeney, D.S.; Bengough, A.G.; Rillig, M.C.; Scrimgeour, C.M.; Young, I.M. Disentangling the impact of AM fungi versus roots on soil structure and water transport. Plant Soil 2009, 314, 183–196. [Google Scholar] [CrossRef]
- Verbist, K.; Torfs, S.; Cornelis, W.M.; Oyarzún, R.; Soto, G.; Gabriels, D. Comparison of single- and double-ring infiltrometer methods on stony soils. Vadose Zone J. 2010, 9, 462–475. [Google Scholar] [CrossRef]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Use and misuse of PLFA measurements in soils. Soil Biol. Biochem. 2011, 43, 1621–1625. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, X.; Zhao, J.; Du, X.; Ma, B. Analysis of the microbial communities of three kinds of Fen-Daqu by PLFAs. J. Inst. Brew. 2016, 122, 34–41. [Google Scholar] [CrossRef]
- Ruess, L.; Chamberlain, P.M. The fat that matters: Soil food web analysis using fatty acids and their carbon stable isotope signature. Soil Biol. Biochem. 2010, 42, 1898–1910. [Google Scholar] [CrossRef]
- Willers, C.; Van Rensburg, P.J.J.; Claassens, S. Phospholipid fatty acid profiling of microbial communities—A review of interpretations and recent applications. J. Appl. Microbiol. 2015, 119, 1207–1218. [Google Scholar] [CrossRef] [PubMed]
- Dungait, J.A.J.; Kemmitt, S.J.; Michallon, L.; Guo, S.; Wen, Q.; Brookes, P.C.; Evershed, R.P. Variable responses of the soil microbial biomass to trace concentrations of 13C-labelled glucose, using 13C-PLFA analysis. Eur. J. Soil Sci. 2011, 62, 117–126. [Google Scholar] [CrossRef]
- Liu, G.; Jin, M.; Cai, C.; Ma, C.; Chen, Z.; Gao, L. Soil microbial community structure and physicochemical properties in Amomum tsaoko-based agroforestry systems in the Gaoligong Mountains, Southwest China. Sustainability 2019, 11, 546. [Google Scholar] [CrossRef] [Green Version]
- Karberg, N.J.; Scott, N.A.; Giardina, C.P. Methods for Estimating Litter Decomposition. In Field Measurements for Forest Carbon Monitoring; Hoover, C.M.., Ed.; Springer US: New York, NY, USA, 2008; ISBN 9781402085062. [Google Scholar]
- Olson, J. Energy storage and the balance of producers and decomposers in ecological system. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Vibrans, H. Malezas de Mexico. Available online: http://www.conabio.gob.mx/malezasdemexico/2inicio/home-malezas-mexico.htm (accessed on 15 November 2020).
- Pearson, C.J.; Ison, R.L. Agronomy of Grassland Systems, 2nd ed.; Cambridge University Press: Cambridge, UK, 1997; ISBN 0521310091. [Google Scholar]
- Aguilar-Fernández, R. Análisis de los Componentes Socioambientales Para la Restauración de Claros Antropogénicos en la Selva Tropical Húmeda, Marqués de Comillas, Chiapas, México. Master’s Thesis, Universidad Nacional Autónoma de México, Mexico City, México, November 2013. [Google Scholar]
- Fanin, N.; Kardol, P.; Farrell, M.; Nilsson, M.C.; Gundale, M.J.; Wardle, D.A. The ratio of Gram-positive to Gram-negative bacterial PLFA markers as an indicator of carbon availability in organic soils. Soil Biol. Biochem. 2019, 128, 111–114. [Google Scholar] [CrossRef]
- Ho, A.; Di Lonardo, D.P.; Bodelier, P.L.E. Revisiting life strategy concepts in environmental microbial ecology. FEMS Microbiol. Ecol. 2017, 93, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, S.; Hutchings, M.I.; Mascher, T. Cell envelope stress response in Gram-positive bacteria. FEMS Microbiol. Rev. 2007, 32, 107–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unger, I.M.; Kennedy, A.C.; Muzika, R.-M. Flooding effects on soil microbial communities. Appl. Soil Ecol. 2009, 42, 1–8. [Google Scholar] [CrossRef]
- Johnson, N.C.; Wilson, G.W.T.; Bowker, M.A.; Wilson, J.A.; Miller, R.M. Resource limitation is a driver of local adaptation in mycorrhizal symbioses. Proc. Natl. Acad. Sci. USA 2010, 107, 2093–2098. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Cambridge, MA, USA, 2008; ISBN 9780080559346. [Google Scholar]
- Gavito, M.E.; Unam, Morelia, Michoacán, México. Personal Communication, 2020.
- Aryal, D.R.; Gómez-González, R.R.; Hernández-Nuriasmú, R.; Morales-Ruiz, D.E. Carbon stocks and tree diversity in scattered tree silvopastoral systems in Chiapas, Mexico. Agrofor. Syst. 2019, 93, 213–227. [Google Scholar] [CrossRef]
- Zhou, G.; Zhou, X.; He, Y.; Shao, J.; Hu, Z.; Liu, R.; Zhou, H.; Hosseinibai, S. Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: A meta-analysis. Glob. Chang. Biol. 2017, 23, 1167–1179. [Google Scholar] [CrossRef]
- Trilleras, J.M.; Jaramillo, V.J.; Vega, E.V.; Balvanera, P. Effects of livestock management on the supply of ecosystem services in pastures in a tropical dry region of western Mexico. Agric. Ecosyst. Environ. 2015, 211, 133–144. [Google Scholar] [CrossRef]
- Zermeño-Hernández, I.; Pingarroni, A.; Martínez-Ramos, M. Agricultural land-use diversity and forest regeneration potential in human- modified tropical landscapes. Agric. Ecosyst. Environ. 2016, 230, 210–220. [Google Scholar] [CrossRef]
- Puschel, D.; Janouskova, M.; Hujslova, M.; Slavikova, R.; Gryndlerova, H.; Jansa, J. Plant–fungus competition for nitrogen erases mycorrhizal growth benefits of Andropogon gerardii under limited nitrogen supply. Ecol. Evol. 2016, 6, 4332–4346. [Google Scholar] [CrossRef] [Green Version]
- Secretaria de Agricultura y Desarrollo Rural (Sagarpa). Comisión Técnico Consultiva de Coeficientes de Agostadero (COTECOCA); Sagarpa: Mexico City, México, 2014.
- Vázquez, E.; Teutscherová, N.; Sotelo, M.; Villegas, D.; Velasquez, N.; Baquero, D.; Pulleman, M.; Arango, J. Intensive short-duration rotational grazing is associated with improved soil quality within one year after establishment in Colombia. Appl. Soil Ecol. 2020, 159, 103835. [Google Scholar] [CrossRef]
- Vázquez, E.; Teutscherova, N.; Lojka, B.; Arango, J.; Pulleman, M. Pasture diversification affects soil macrofauna and soil biophysical properties in tropical (silvo)pastoral systems. Agric. Ecosyst. Environ. 2020, 302, 107083. [Google Scholar] [CrossRef]
- Webster, E.; Gaudin, A.C.M.; Pulleman, M.; Siles, P.; Fonte, S.J. Improved Pastures Support Early Indicators of Soil Restoration in Low-input Agroecosystems of Nicaragua. Environ. Manag. 2019, 64, 201–212. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variable | Average | Min | Max | Units |
|---|---|---|---|---|
| Landscape position | ||||
| distance from the river | 3.04 | 0.17 | 13.00 | km |
| altitude | 166 | 143 | 186 | masl |
| History | ||||
| age of the plot (Y) | 25 | 4 | 40 | years |
| Management | ||||
| size of the plot (SZ) | 20 | 5 | 50 | ha |
| livestock heads (LH) | 32 | 3 | 67 | head |
| stocking density (SD) | 2.16 | 0.10 | 9.00 | head ha−1 |
| grazing time (GT) | 0.48 | 0.11 | 1.00 | - |
| grazing severity (GS) | 1.01 | 0.05 | 4.05 | - |
| fire incidence (FI) | 3.42 | 1 | 11 | burns |
| fire frequency (FF) | 0.16 | 0.03 | 0.60 | burns/year |
| herbicide incidence (HI) | 5.94 | 0 | 33 | applications |
| herbicide frequency (HF) | 0.28 | 0 | 1 | - |
| tractor use (TI) | 0.13 | 0 | 1 | times |
| Ecological Disturbance Index (EDI) | 1.12 | 0.54 | 2.13 |
| Variable | Average | Min | Max | Units |
|---|---|---|---|---|
| Soil properties | ||||
| available P | 2.47 | 0.73 | 8.88 | μg g−1 |
| ammonium | 25.29 | 0.11 | 72.90 | mg g−1 |
| nitrate | 3.81 | 0 | 9.79 | mg g−1 |
| soil organic matter | 2.31 | 1.19 | 3.79 | % |
| total P | 0.22 | 0.07 | 0.72 | mg g−1 |
| total N | 0.16 | 0.06 | 0.34 | % |
| pH | 6.35 | 5.30 | 8.76 | - |
| bulk density | 1.08 | 0.77 | 1.43 | g cm−3 |
| sand content | 39.70 | 11.95 | 80.02 | % |
| clay content | 19.59 | 1.25 | 55.80 | % |
| silt content | 40.71 | 16.24 | 72.22 | % |
| infiltration | 196 | 20 | 480 | mm hr−1 |
| micro- and macro-aggregates | 67.97 | 36.53 | 84.84 | % |
| Soil biological composition | ||||
| total AMF | 8.21 | 1.56 | 17.68 | nmol g−1 soil |
| total fungi | 3.11 | 0.28 | 8.25 | nmol g−1 soil |
| total bacteria | 23.29 | 9.32 | 45.19 | nmol g−1 soil |
| Gram+/Gram- | 1.96 | 1.42 | 3.45 | - |
| bacteria/fungi | 0.15 | 0.02 | 0.80 | - |
| Ecosystem Processes | ||||
| Decomposition rate | 0.004 | 0.003 | 0.016 | day−1 |
| Ecosystem Services | ||||
| protein content | 7.3 | 4.46 | 15.37 | % |
| forage productivity (T1) | 317 | 18 | 852 | kg ha−1 |
| forage productivity (T2) | 601 | 114 | 1418 | kg ha−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguilar-Fernández, R.; Gavito, M.E.; Peña-Claros, M.; Pulleman, M.; Kuyper, T.W. Exploring Linkages between Supporting, Regulating, and Provisioning Ecosystem Services in Rangelands in a Tropical Agro-Forest Frontier. Land 2020, 9, 511. https://doi.org/10.3390/land9120511
Aguilar-Fernández R, Gavito ME, Peña-Claros M, Pulleman M, Kuyper TW. Exploring Linkages between Supporting, Regulating, and Provisioning Ecosystem Services in Rangelands in a Tropical Agro-Forest Frontier. Land. 2020; 9(12):511. https://doi.org/10.3390/land9120511
Chicago/Turabian StyleAguilar-Fernández, Rocío, Mayra E. Gavito, Marielos Peña-Claros, Mirjam Pulleman, and Thomas W. Kuyper. 2020. "Exploring Linkages between Supporting, Regulating, and Provisioning Ecosystem Services in Rangelands in a Tropical Agro-Forest Frontier" Land 9, no. 12: 511. https://doi.org/10.3390/land9120511