Planning for Dynamic Connectivity: Operationalizing Robust Decision-Making and Prioritization Across Landscapes Experiencing Climate and Land-Use Change

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
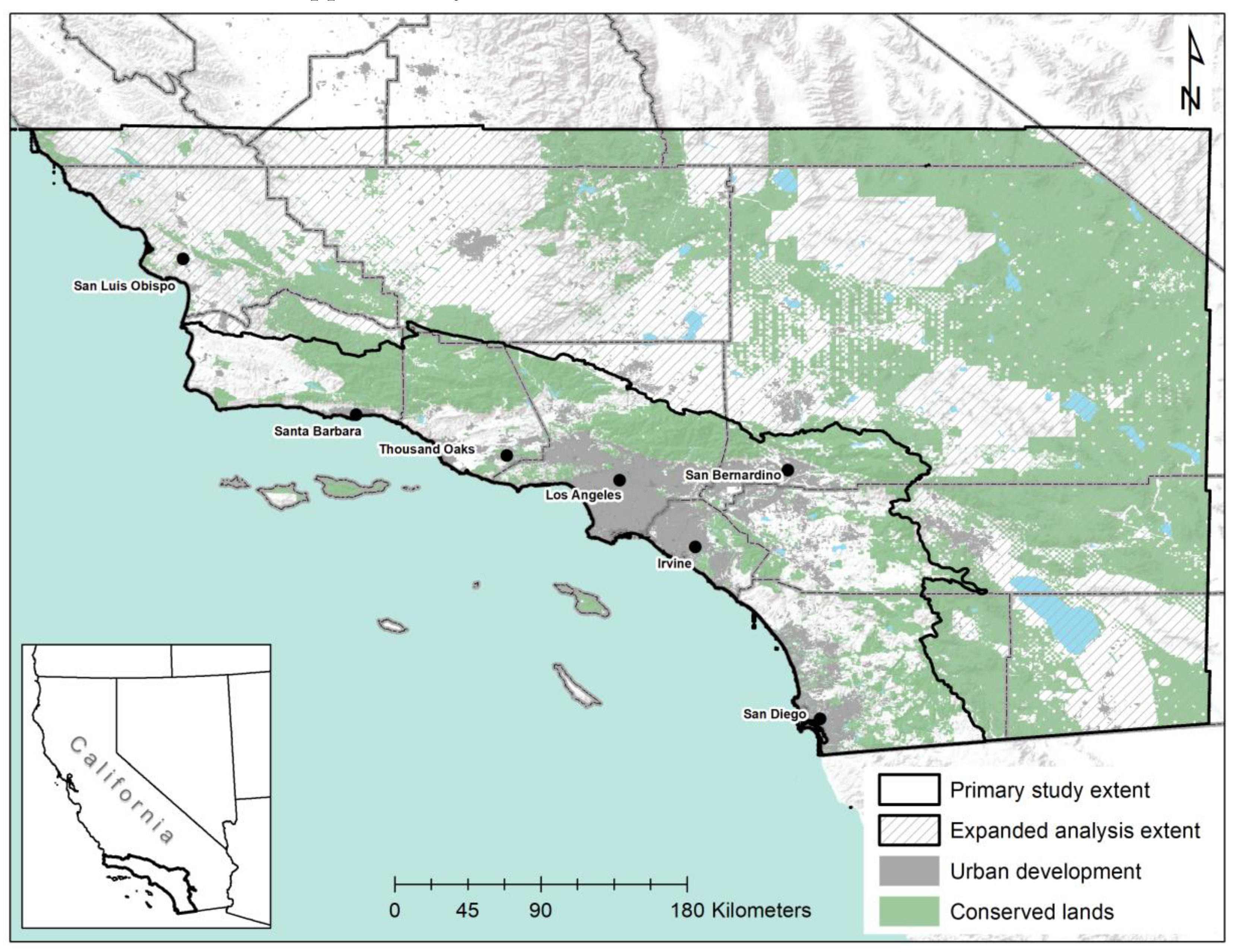
2.1. Study Area
2.2. Modeling Approach
2.2.1. Habitat Suitability, Resistance, and Core Area Modeling
2.2.2. Linkage Modeling
2.2.3. Metapopulation Modeling
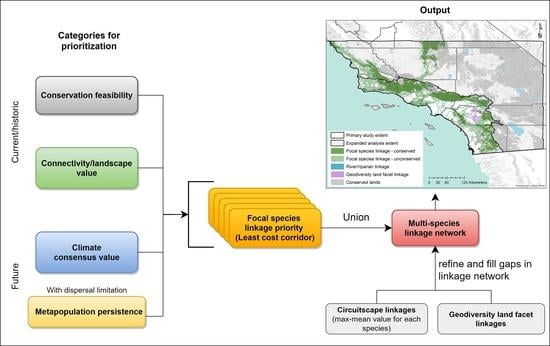
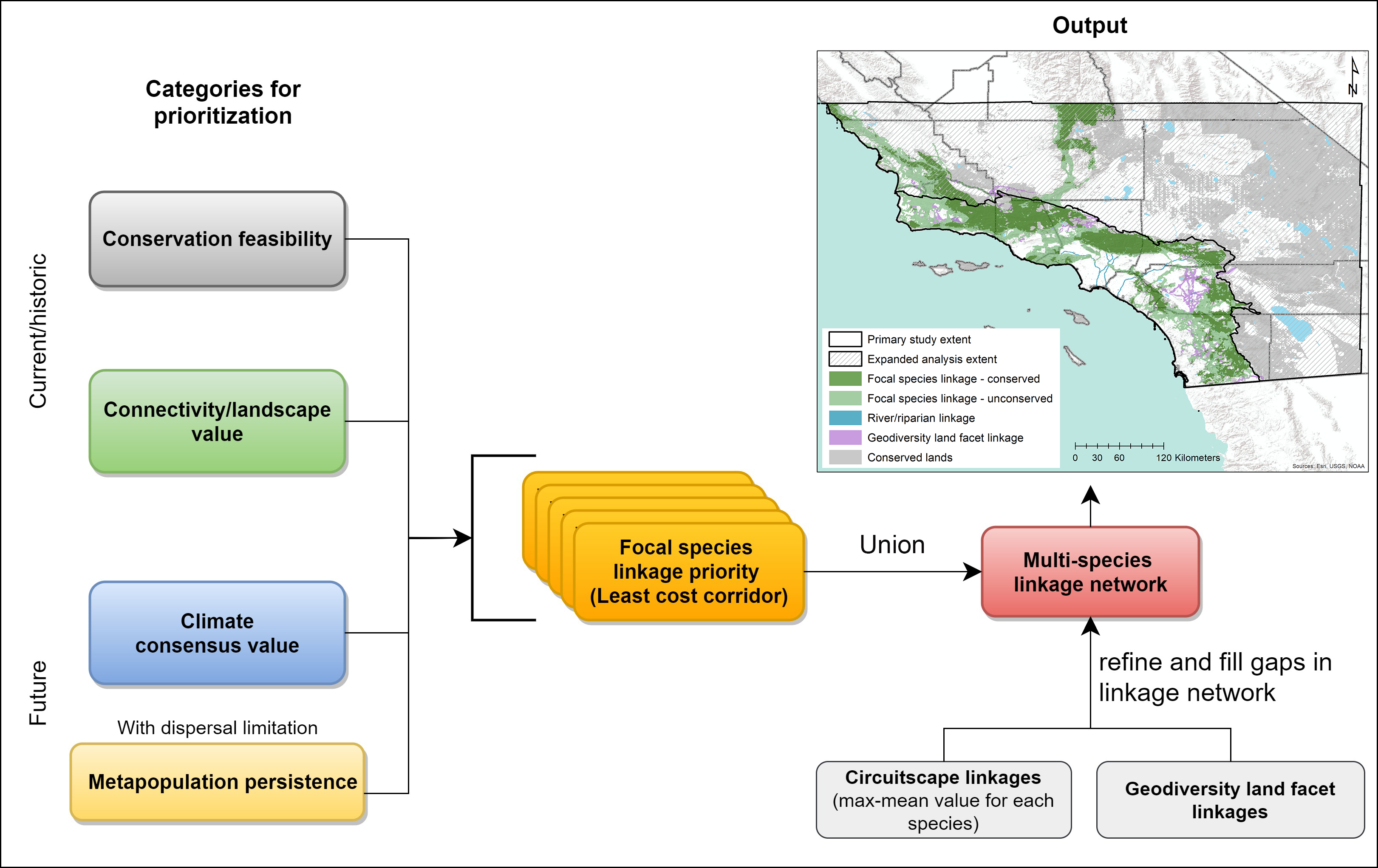
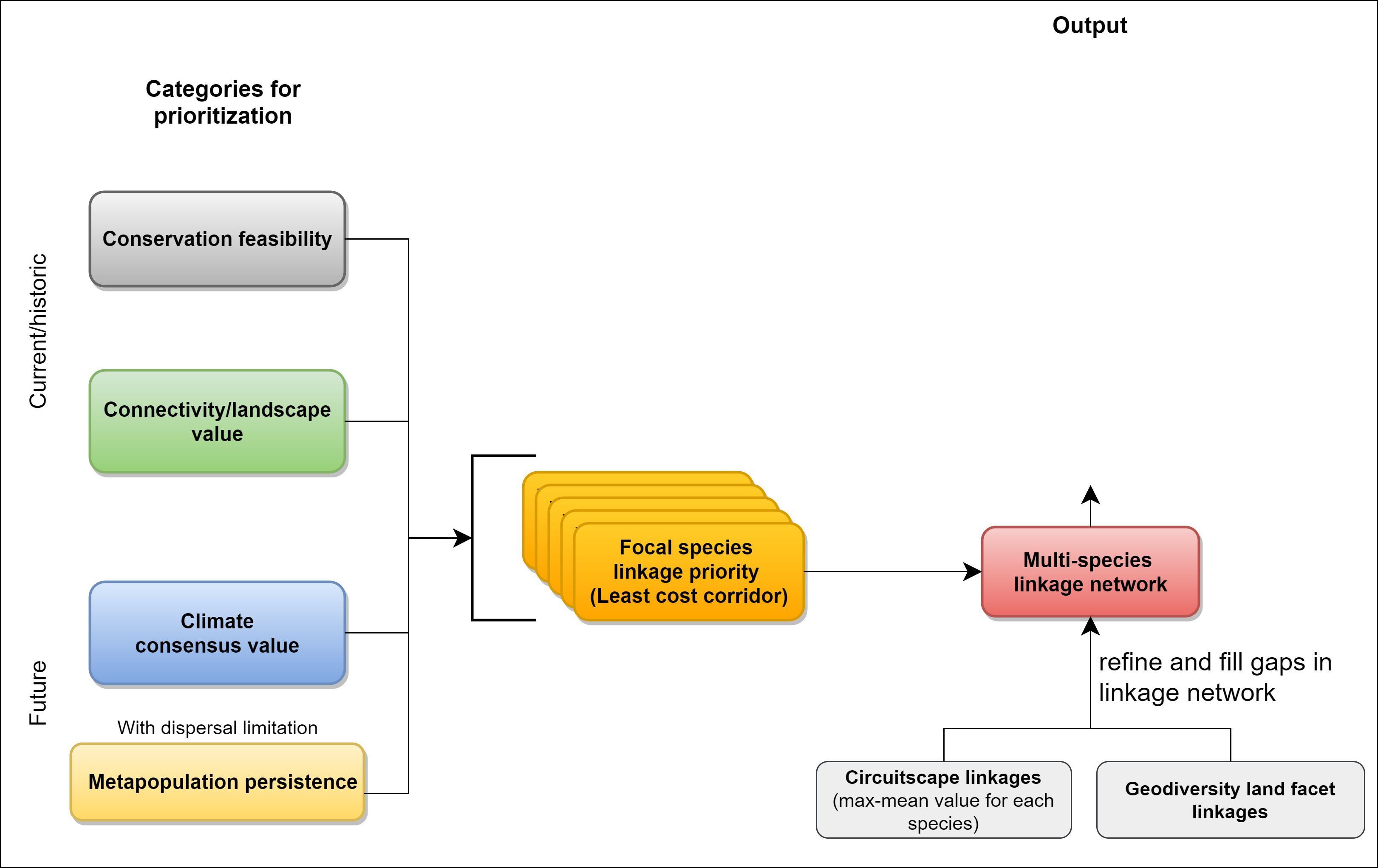
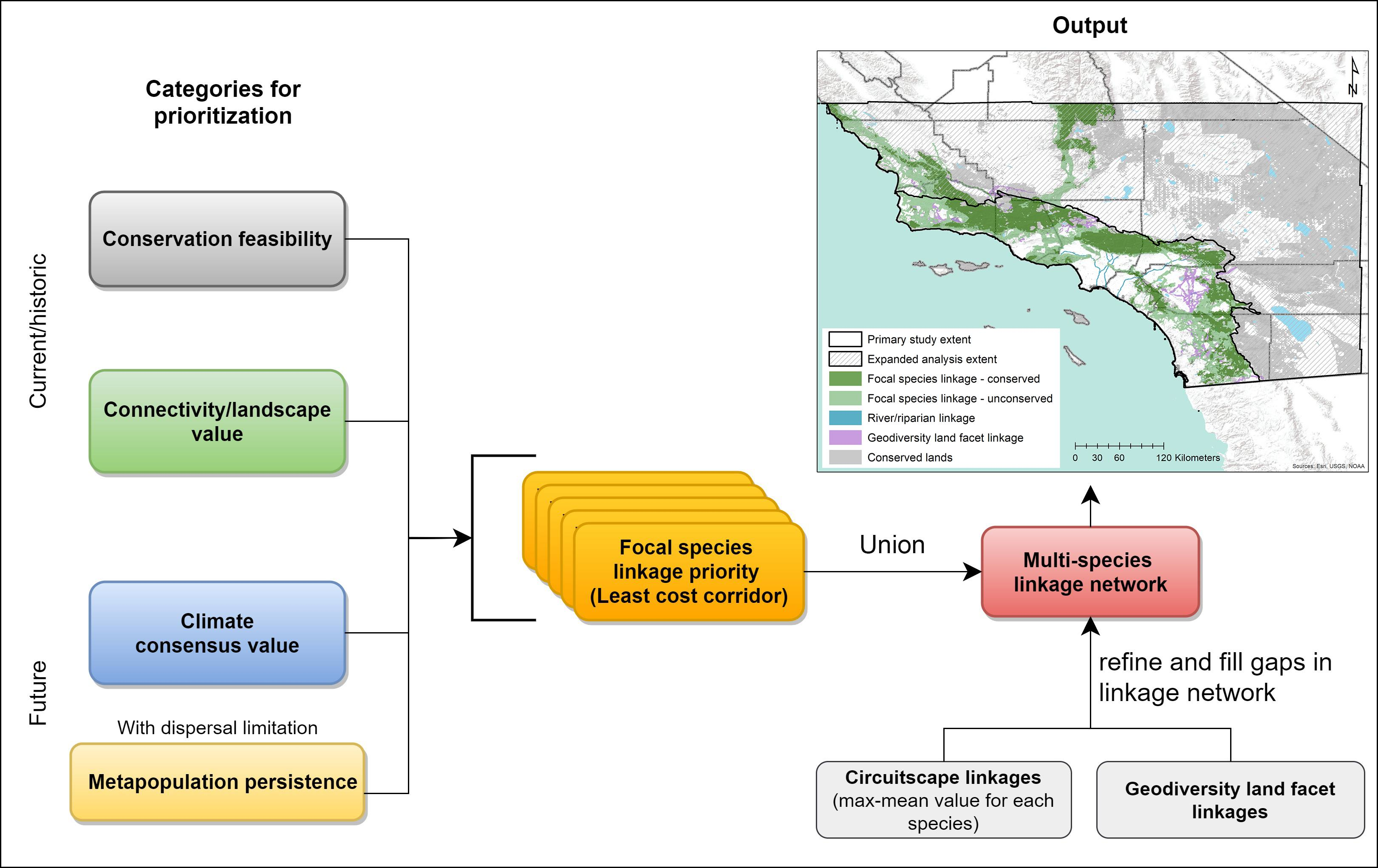
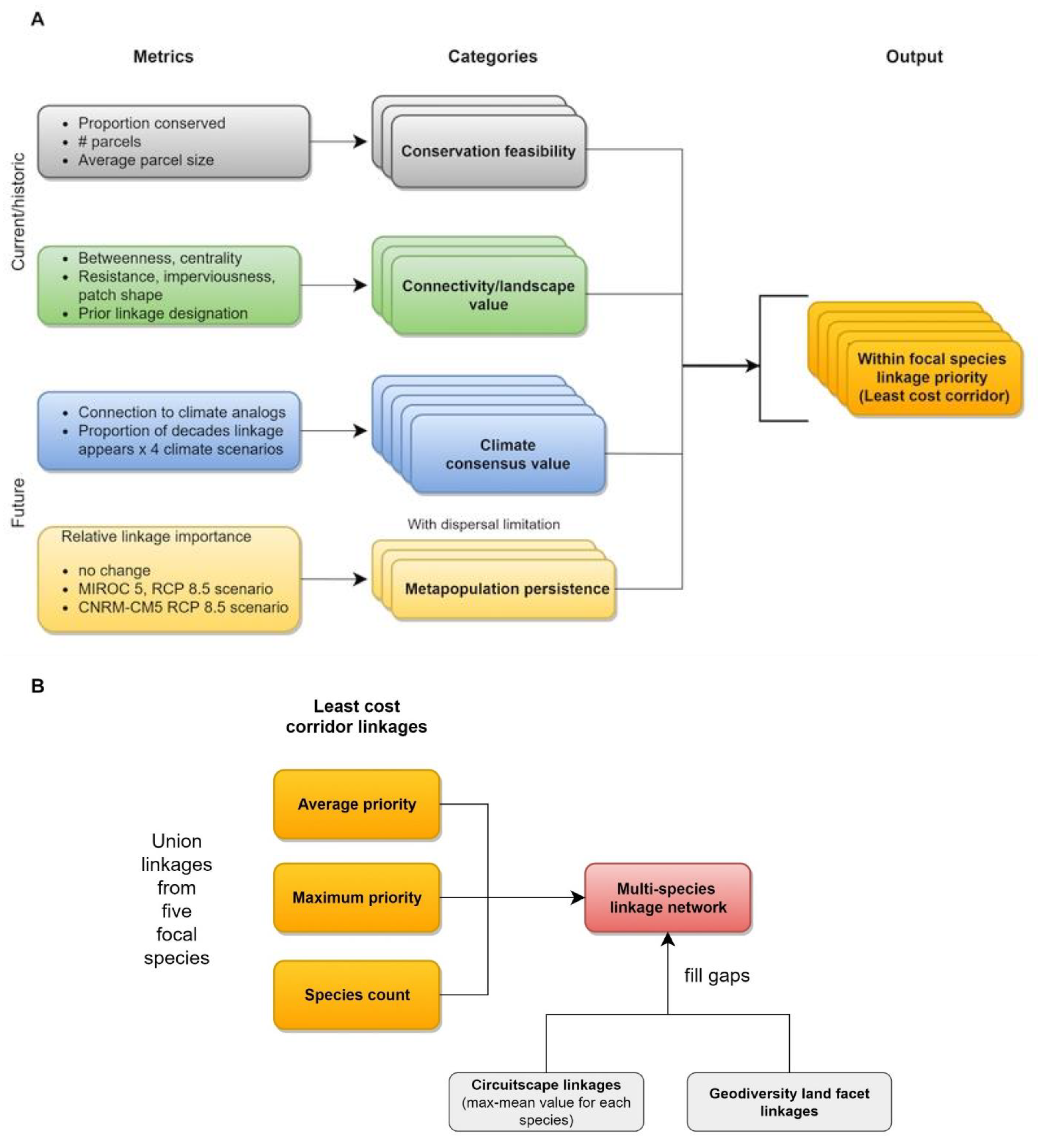
2.3. Prioritizing a Multispecies Linkage Network
2.3.1. Within-Species Prioritization
2.3.2. Multispecies Prioritization
3. Results
3.1. Habitat Suitability, Linkage, and Metapopulation Modeling
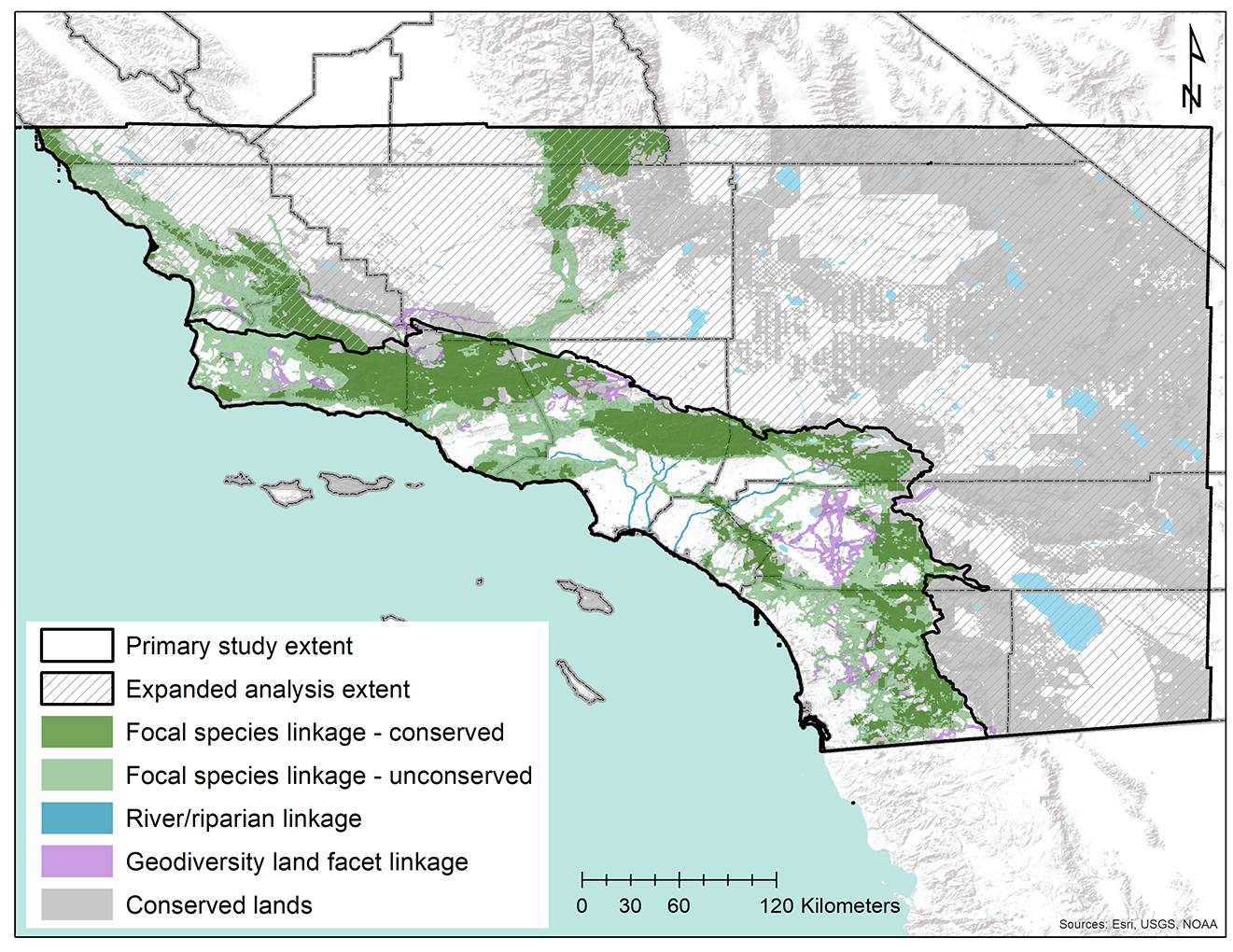
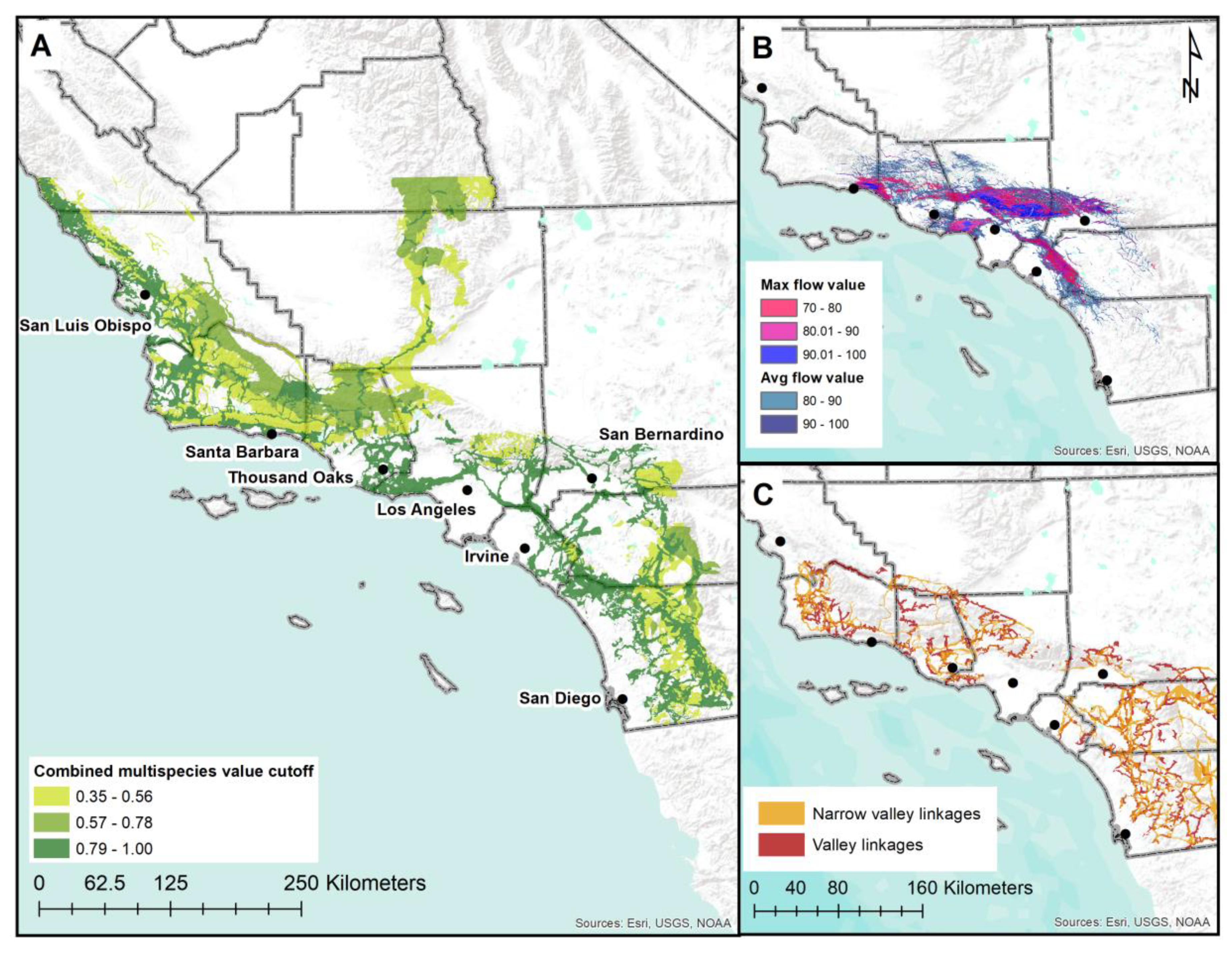
3.2. Prioritizing a Multispecies Linkage Network
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef] [Green Version]
- McRae, B.H.; Hall, S.A.; Beier, P.; Theobald, D.M. Where to Restore Ecological Connectivity? Detecting Barriers and Quantifying Restoration Benefits. PLoS ONE 2012, 7, e52604. [Google Scholar] [CrossRef] [PubMed]
- Heller, N.E.; Zavaleta, E.S. Biodiversity management in the face of climate change: A review of 22 years of recommendations. Biol. Conserv. 2009, 142, 14–32. [Google Scholar] [CrossRef]
- Keeley, A.T.H.; Ackerly, D.D.; Cameron, D.R.; Heller, N.E.; Huber, P.R.; Schloss, C.A.; Thorne, J.H.; Merenlender, A.M. New concepts, models, and assessments of climate-wise connectivity. Environ. Res. Lett. 2018, 13, 073002. [Google Scholar] [CrossRef]
- Beier, P.; Gregory, A.J. Desperately seeking sTable 50-year-old landscapes with patches and long, wide corridors. PLoS Biol. 2012, 10, e1001253. [Google Scholar] [CrossRef] [Green Version]
- Noss, R.F. Landscape connectivity: Different functions at different scales. In Landscape linkages and Biodiversity; Hudson, W.E., Ed.; Island Press: Washington, DC, USA, 1991; pp. 27–39. [Google Scholar]
- Hilty, J.A.; Lidicker, W., Jr.; Merenlender, A.M. Corridor Ecology: The Science and Practice of Linking Landscapes for Biodiversity Conservation; Island Press: Washington, DC, USA, 2006; ISBN 1559630477. [Google Scholar]
- Brown, J.H.; Kodric-Brown, A. Turnover Rates in Insular Biogeography: Effect of Immigration on Extinction. Ecology 1977, 58, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Simberloff, D.; Farr, J.A.; Cox, J.; Mehlman, D.W. Movement corridors: Conservation bargains or poor investments? Conserv. Biol. 1992, 6, 493–504. [Google Scholar] [CrossRef]
- Hannah, L.; Midgley, G.F.; Millar, D. Climate change-integrated conservation strategies. Glob. Ecol. Biogeogr. 2002, 11, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Saura, S.; Bertzky, B.; Bastin, L.; Battistella, L.; Mandrici, A.; Dubois, G. Protected area connectivity: Shortfalls in global targets and country-level priorities. Biol. Conserv. 2018, 219, 53–67. [Google Scholar] [CrossRef]
- Keeley, A.T.H.; Beier, P.; Creech, T.; Jones, K.; Jongman, R.H.G.; Stonecipher, G.; Tabor, G.M. Thirty years of connectivity conservation planning: An assessment of factors influencing plan implementation. Environ. Res. Lett. 2019, 14, 103001. [Google Scholar] [CrossRef]
- Luque, S.; Saura, S.; Fortin, M.-J. Landscape connectivity analysis for conservation: Insights from combining new methods with ecological and genetic data. Landsc. Ecol. 2012, 27, 153–157. [Google Scholar] [CrossRef]
- Dilkina, B.; Houtman, R.; Gomes, C.P.; Montgomery, C.A.; McKelvey, K.S.; Kendall, K.; Graves, T.A.; Bernstein, R.; Schwartz, M.K. Trade-offs and efficiencies in optimal budget-constrained multispecies corridor networks. Conserv. Biol. 2017, 31, 192–202. [Google Scholar] [CrossRef]
- Gippoliti, S.; Battisti, C. More cool than tool: Equivoques, conceptual traps and weaknesses of ecological networks in environmental planning and conservation. Land Use policy 2017, 68, 686–691. [Google Scholar] [CrossRef]
- Albert, C.H.; Rayfield, B.; Dumitru, M.; Gonzalez, A. Applying network theory to prioritize multispecies habitat networks that are robust to climate and land-use change. Conserv. Biol. 2017, 31, 1383–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brost, B.M.; Beier, P. Comparing linkage designs based on land facets to linkage designs based on focal species. PLoS ONE 2012, 7, e48965. [Google Scholar] [CrossRef] [Green Version]
- Krosby, M.; Breckheimer, I.; Pierce, D.J.; Singleton, P.H.; Hall, S.A.; Halupka, K.C.; Gaines, W.L.; Long, R.A.; McRae, B.H.; Cosentino, B.L.; et al. Focal species and landscape “naturalness” corridor models offer complementary approaches for connectivity conservation planning. Landsc. Ecol. 2015, 30, 2121–2132. [Google Scholar] [CrossRef]
- McGuire, J.L.; Lawler, J.J.; McRae, B.H.; Nuñez, T.A.; Theobald, D.M. Achieving climate connectivity in a fragmented landscape. Proc. Natl. Acad. Sci. USA 2016, 113, 7195–7200. [Google Scholar] [CrossRef] [Green Version]
- Hanski, I. Metapopulation Ecology; Oxford University Press: Oxford, UK, 1999; p. 313. ISBN 0198540655. [Google Scholar]
- Bengtsson, J.; Angelstam, P.; Elmqvist, T.; Emanuelsson, U.; Folke, C.; Ihse, M.; Moberg, F.; Nyström, M. Reserves, Resilience and Dynamic Landscapes. AMBIO 2003, 32, 389–396. [Google Scholar] [CrossRef]
- Vos, C.C.; van der Hoek, D.C.J.; Vonk, M. Spatial planning of a climate adaptation zone for wetland ecosystems. Landsc. Ecol. 2010, 25, 1465–1477. [Google Scholar] [CrossRef]
- Nuñez, T.A.; Lawler, J.J.; McRae, B.H.; Pierce, D.J.; Krosby, M.B.; Kavanagh, D.M.; Singleton, P.H.; Tewksbury, J.J. Connectivity Planning to Address Climate Change. Conserv. Biol. 2013, 27, 407–416. [Google Scholar] [CrossRef]
- Penrod, K.; Beier, P.; Garding, E.; Cabañero, C. A Linkage Network for the California Deserts; Technical Report for the Bureau of Land Management and The Wildlands Conservancy: Fair Oaks, CA, USA, February 2012.
- Underwood, E.C.; Viers, J.H.; Klausmeyer, K.R.; Cox, R.L.; Shaw, M.R. Threats and biodiversity in the mediterranean biome. Divers. Distrib. 2009, 15, 188–197. [Google Scholar] [CrossRef]
- Syphard, A.D.; Radeloff, V.C.; Keeley, J.E.; Hawbaker, T.J.; Clayton, M.K.; Stewart, S.I.; Hammer, R.B. Human influence on California fire regimes. Ecol. Appl. 2007, 17, 1388–1402. [Google Scholar] [CrossRef]
- Franklin, J.; Wejnert, K.E.; Hathaway, S.A.; Rochester, C.J.; Fisher, R.N. Effect of species rarity on the accuracy of species distribution models for reptiles and amphibians in southern California. Divers. Distrib. 2009, 15, 167–177. [Google Scholar] [CrossRef]
- Daly, C. Guidelines for assessing the suitability of spatial climate data sets. Int. J. Climatol. 2006, 26, 707–721. [Google Scholar] [CrossRef]
- U.S. Geological Survey National Elevation Dataset. Sioux Falls, SD: EROS. 2009. Available online: http://viewer.nationalmap.gov/viewer/ (accessed on 15 July 2016).
- Flint, L.E.; Flint, A.L. Downscaling future climate scenarios to fine scales for hydrologic and ecological modeling and analysis. Ecol. Process. 2012, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Pierce, D.W.; Cayan, D.R.; Thrasher, B.L. Statistical Downscaling Using Localized Constructed Analogs (LOCA)*. J. Hydrometeorol. 2014, 15, 2558–2585. [Google Scholar] [CrossRef]
- Jin, S.; Yang, L.; Danielson, P.; Homer, C.; Fry, J.; Xian, G. A comprehensive change detection method for updating the National Land Cover Database to circa 2011. Remote Sens. Environ. 2013, 132, 159–175. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, J.K.H.; Carlisle, D.M.; May, J.T.; Klausmeyer, K.R.; Grantham, T.E.; Brown, L.R.; Howard, J.K. Patterns and magnitude of flow alteration in California, USA. Freshw. Biol. 2018, 63, 859–873. [Google Scholar] [CrossRef] [Green Version]
- Jenness, J. DEM Surface Tools. Available online: http://www.jennessent.com/arcgis/surface_area.htm (accessed on 15 July 2016).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2017. [Google Scholar]
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. Ser. B 2011, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- biomod2: Ensemble Platform for Species Distribution Modeling. Available online: https://cran.r-project.org/web/packages/biomod2/index.html (accessed on 1 August 2020).
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. Package ‘dismo’’-Species Distribution Modeling’. Circles 2017, 9, 1–68. [Google Scholar]
- Shirk, A.; McRae, B.H. Gnarly Landscape Utilities: Core Mapper User Guide; The Nature Conservancy: Fort Collins, CO, USA, 2013. [Google Scholar]
- Adriaensen, F.; Chardon, J.P.; De Blust, G.; Swinnen, E.; Villalba, S.; Gulinck, H.; Matthysen, E. The application of ‘least-cost’ modelling as a functional landscape model. Landsc. Urban Plan. 2003, 64, 233–247. [Google Scholar] [CrossRef]
- Keeley, A.T.H.; Beier, P.; Gagnon, J.W. Estimating landscape resistance from habitat suitability: Effects of data source and nonlinearities. Landsc. Ecol. 2016, 31, 2151–2162. [Google Scholar] [CrossRef]
- Trainor, A.M.; Walters, J.R.; Morris, W.F.; Sexton, J.; Moody, A. Empirical estimation of dispersal resistance surfaces: A case study with red-cockaded woodpeckers. Landsc. Ecol. 2013, 28, 755–767. [Google Scholar] [CrossRef]
- Mateo-Sánchez, M.C.; Balkenhol, N.; Cushman, S.; Pérez, T.; Domínguez, A.; Saura, S. A comparative framework to infer landscape effects on population genetic structure: Are habitat suitability models effective in explaining gene flow? Landsc. Ecol. 2015, 30, 1405–1420. [Google Scholar] [CrossRef]
- McRae, B.H.; Kavanagh, D.M. Linkage Mapper Connectivity Analysis Software; The Nature Conservancy: Seattle, WA, USA, 2011. [Google Scholar]
- McRae, B.H.; Dickson, B.G.; Keitt, T.H.; Shah, V.B. Using Circuit Theory to Model Connectivity in Ecology, Evolution, and Conservation. Ecology 2008, 89, 2712–2724. [Google Scholar] [CrossRef] [PubMed]
- Bezanson, J.; Edelman, A.; Karpinski, S.; Shah, V.B. Julia: A fresh approach to numerical computing. SIAM Rev. 2017, 59. [Google Scholar] [CrossRef] [Green Version]
- Anantharaman, R.; Hall, K.; Shah, V.; Edelman, A. Circuitscape in Julia: High performance connectivity modelling to support conservation decisions. arXiv 2019, arXiv:1906.03542. [Google Scholar]
- Comer, P.J.; Pressey, R.L.; Hunter, M.L.; Schloss, C.A.; Buttrick, S.C.; Heller, N.E.; Tirpak, J.M.; Faith, D.P.; Cross, M.S.; Shaffer, M.L. Incorporating geodiversity into conservation decisions. Conserv. Biol. 2015, 29, 692–701. [Google Scholar] [CrossRef]
- Theobald, D.M.; Harrison-Atlas, D.; Monahan, W.B.; Albano, C.M. Ecologically-relevant maps of landforms and physiographic diversity for climate adaptation planning. PLoS ONE 2015, 10, e0143619. [Google Scholar] [CrossRef] [Green Version]
- Beier, P.; Brost, B. Use of land facets to plan for climate change: Conserving the arenas, not the actors. Conserv. Biol. 2010, 24, 701–710. [Google Scholar] [CrossRef]
- Brost, B.M.; Beier, P. Use of land facets to design linkages for climate change. Ecol. Appl. 2012, 22, 87–103. [Google Scholar] [CrossRef]
- Jenness, J.; Brost, B.; Beier, P. Land Facet Corridor Designer: Extension for ArcGIS; Jenness Enterprises: Flagstaff, AZ, USA, 2013; p. 110. [Google Scholar]
- Akçakaya, H.R.; Root, W.T. Linking Landscape Data with Population Viability Analysis (Version 5.0); Applied Mathematics: Setaukey, NY, USA, 2005. [Google Scholar]
- Sheehan, T.; Gough, M. A platform-independent fuzzy logic modeling framework for environmental decision support. Ecol. Inform. 2016, 34, 92–101. [Google Scholar] [CrossRef]
- GreenInfo Network California Protected Areas Database. 2018. Available online: https://www.calands.org/cpad/ (accessed on 31 October 2018).
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. Inter J. Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- South Coast Wildlands. South Coast Wildlands South Coast Missing Linkages: A Wildland Network for the South Coast Ecoregion; South Coast Wildlands: Fair Oaks, CA, USA, 2008; 67p. [Google Scholar]
- Spencer, W.D.; Beier, P.; Penrod, K.; Paulman, K.; Rustigian-Romsos, H.; Strittholt, J.; Parisi, M.; Pettler, A. California Essential Habitat Connectivity Project: A Strategy for Conservation a Connected California; Prepared for California Department of Transportation, California Department of Fish and Game, and Federal Highways Administration: Sacramento, CA, USA, 2010; p. 313.
- Flint, L.E.; Flint, A.L.; Thorne, J.H.; Boynton, R. Fine-scale hydrologic modeling for regional landscape applications: The California Basin Characterization Model development and performance. Ecol. Process. 2013, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Gallo, J.A.; Greene, R. Connectivity Analysis Software for Estimating Linkage Priority; Conservation Biology Institute: Corvallis, OR, USA, 2018. [Google Scholar]
- GreenInfo Network California Conservation Easements Database. 2018. Available online: https://www.calands.org/cced/ (accessed on 31 October 2018).
- Peterson, G.D.; Cumming, G.S.; Carpenter, S.R. Scenario planning: A tool for conservation in an uncertain world. Conserv. Biol. 2003, 17, 358–366. [Google Scholar] [CrossRef] [Green Version]
- Lynn; Elissa; Schwarz, A.; Anderson, J.; Correa, M. Perspectives and Guidance for Climate Change Analysis; Technical Report for California Department of Water Resources (DWR) and Climate Change Technical Advisory Group (CCTAG): Sacramento, CA, USA, August 2015.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Description and Source | Time Variant | |
|---|---|---|---|
| Climate | Source: Downscaled (to 90 m) PRISM, MIROC5 RCP4.5, MIROC5 RCP8.5, CNRM CM5 RCP4.5, CNRM CM5 RCP8.5 | ||
| Bioclim 1 | Mean temperature | Yes | |
| Bioclim 2 | Mean diurnal range (mean of monthly (max temperature-minimum temperature)) | Yes | |
| Bioclim 4 | Temperature Seasonality (Monthly standard deviation *100) | Yes | |
| Bioclim 6 | Minimum temperature of the coldest month | Yes | |
| Bioclim 12 | Mean precipitation | Yes | |
| Bioclim 14 | Precipitation of the driest month | Yes | |
| Bioclim 15 | Precipitation seasonality (coefficient of variation across months) | Yes | |
| Land- use | Source: National Land Cover Database 2011 [33] | ||
| Impervious surfaces | Used as a proxy for urban land cover | No | |
| Water Resources | Source: [34] | ||
| Distance to seasonal streams | Euclidean distance to streams with low probability of year-round flow | No | |
| Distance to perennial streams | Euclidean distance to streams with high probability of year-round flow | No | |
| Stream density | Density of all streams within a 5-km moving window | No | |
| Topography | Source: National Elevation Dataset [30] | ||
| Roughness Index | Total curvature calculated with DEM Surface Tools [35] | No | |
| Percent Slope | Derived from National Elevation Dataset | No | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jennings, M.K.; Haeuser, E.; Foote, D.; Lewison, R.L.; Conlisk, E. Planning for Dynamic Connectivity: Operationalizing Robust Decision-Making and Prioritization Across Landscapes Experiencing Climate and Land-Use Change. Land 2020, 9, 341. https://doi.org/10.3390/land9100341
Jennings MK, Haeuser E, Foote D, Lewison RL, Conlisk E. Planning for Dynamic Connectivity: Operationalizing Robust Decision-Making and Prioritization Across Landscapes Experiencing Climate and Land-Use Change. Land. 2020; 9(10):341. https://doi.org/10.3390/land9100341
Chicago/Turabian StyleJennings, Megan K., Emily Haeuser, Diane Foote, Rebecca L. Lewison, and Erin Conlisk. 2020. "Planning for Dynamic Connectivity: Operationalizing Robust Decision-Making and Prioritization Across Landscapes Experiencing Climate and Land-Use Change" Land 9, no. 10: 341. https://doi.org/10.3390/land9100341