1. Introduction
Woody alien plants are particularly important in the Northeastern United States: they make up 32% of the invasive plant species in the region and constitute about 70% of the invasive plant occurrence records [
1]. In Connecticut, invasive species constitute a high percentage of the vascular plant flora, and have created ecological damage and economic losses [
1,
2,
3,
4]. The current flora is composed of 35% percent nonindigenous species [
3]. The main invasive species impacts include direct and indirect economic costs to property, human health, and ecosystem services [
5,
6,
7,
8,
9,
10]. The cost to control invasive species and the damages they inflicted upon properties and natural resources in the U.S. is estimated at $137 billion annually [
11]. In addition, these invasive species pose an important threat to biodiversity in Connecticut [
3]. Invasive plants co-occur with nearly half of rare plant species in New England, and are most frequent and diverse in Connecticut among all New England states [
7].
One especially problematic invasive plant in Connecticut is
Berberis thunbergii (Japanese barberry). This perennial woody shrub provides excellent habitats for Lyme-disease carrying ticks thereby increasing the risk of infection in humans [
12,
13] in addition to altering ecosystem functions, such as nitrogen cycling, in invaded areas [
14]. Japanese barberry was the second most frequently observed invasive plant at rare species sites in New England, and invasive plant presence was correlated with decreases in rare species population size [
7]. Predicting future changes in invasion probability for Japanese barberry could help direct management efforts to mitigate its effects.
Land use patterns play an important role in the establishment and spread of invasive plants. Land use changes can aid plant invasions by promoting disturbances that may be a key to establishment, by creating habitats that may be more favorable to particular species, and also by creating dispersal corridors [
15,
16]. Therefore, incorporating land use change is a priority in models forecasting future invasions. Previous studies suggested that land use changes provide opportunities for particular plants to invade an area [
17,
18,
19], that the types of changes that promote one species might inhibit others [
18], and that land use changes contribute not only to invasive species establishment but also to their spread [
20,
21]. The generation of forest edges and the fragmentation of contiguous forest habitats are particularly important to woody invasive plant richness [
22]. There are no studies, however, that analyze and predict future land use changes related to invasive species that might help predict the spatial distribution of future invasive species based on the predicted land use and land cover (LULC) data.
Connecticut is the third smallest state by area, but also the fourth most densely populated state among the 50 United States [
23]. Land in Connecticut has gone through a tremendous change over the past two centuries. For example, up to 90% of the forest land was cleared for farming by the mid-1800s [
24]. As farms were abandoned, most of the lands reverted back to mixed hardwood forests. Over the past fifty years, the region has undergone significant land use changes as housing and industrial development has encroached upon formerly rural and forested lands [
18,
25]. There are a few of studies that investigate how land uses and covers have changed throughout the Connecticut region. For example, Drummond and Loveland [
26] studied land-use pressure and land transitions in the northeastern United States. Their results showed that a regional scale decline in forest cover was caused by contemporary land-use pressures in the eastern United States. Increasing crop production, pasturing, residential and industrial development, and uses of fuel wood and other resources caused nearly half or more of the natural forests to be cleared in the past three centuries.
Hurd et al. [
27] analyzed forest fragmentation in the Salmon River watershed in Connecticut. They found that the interior forest area declined, while edge, transitional and perforated forest and urban areas increased between 1985 and 1999 [
27,
28]. Parent et al. [
29] also studied the Salmon River watershed and found that the forest fragmentation in the region was mostly the result of land cover changes associated with building construction. They indicated that suburban development was a major contributor to forest fragmentation in the northeastern United States. Road development and socio-economic data were not taken into consideration in the study, however. It is also worth noting that the Center for Land Use Education And Research (CLEAR) at the University of Connecticut developed a temporal series of basic land cover information for the Thames watershed and surrounding towns to help gain a better understanding of the extent of land cover changes occurring in the Connecticut landscape [
28].
Although the general picture of land use change in the past two centuries in Connecticut is known, prediction of future land use in the region remains difficult. And there is little research to predict the future land use change in this region. Complicating the prediction further, land use changes are not simple processes. It is difficult to conduct an accurate prediction of land use changes for a region without sufficient data and knowledge about the study area and well-developed and theoretically-informed models [
30,
31,
32,
33]. The models should have the ability to represent dynamically the processes of land-use change and biophysical processes. This has to be based on dynamic system models, which represent functional complexity [
34]. In addition, future predictions of invasive plant distributions in the region have not incorporated land use change despite its importance [
16,
19,
23,
35] due to lack of future land use change data.
In this study, we aim to fill the data gap of future land use change predictions by: (1) explaining land use changes in Connecticut over 1996–2006; (2) constructing a Multi-layer Perceptron_Markov Chain (MLP_MC) model to predict short-term (12 years) changes in land use; and (3) investigate how short-term land use changes may affect landscape scale Japanese barberry invasion probability. The land use models provide a unique perspective into the drivers of land use change in our study region, while the short-term future land use predictions provide an insight into possible changes related to an important regional invasive plant.
3. Results and Analyses
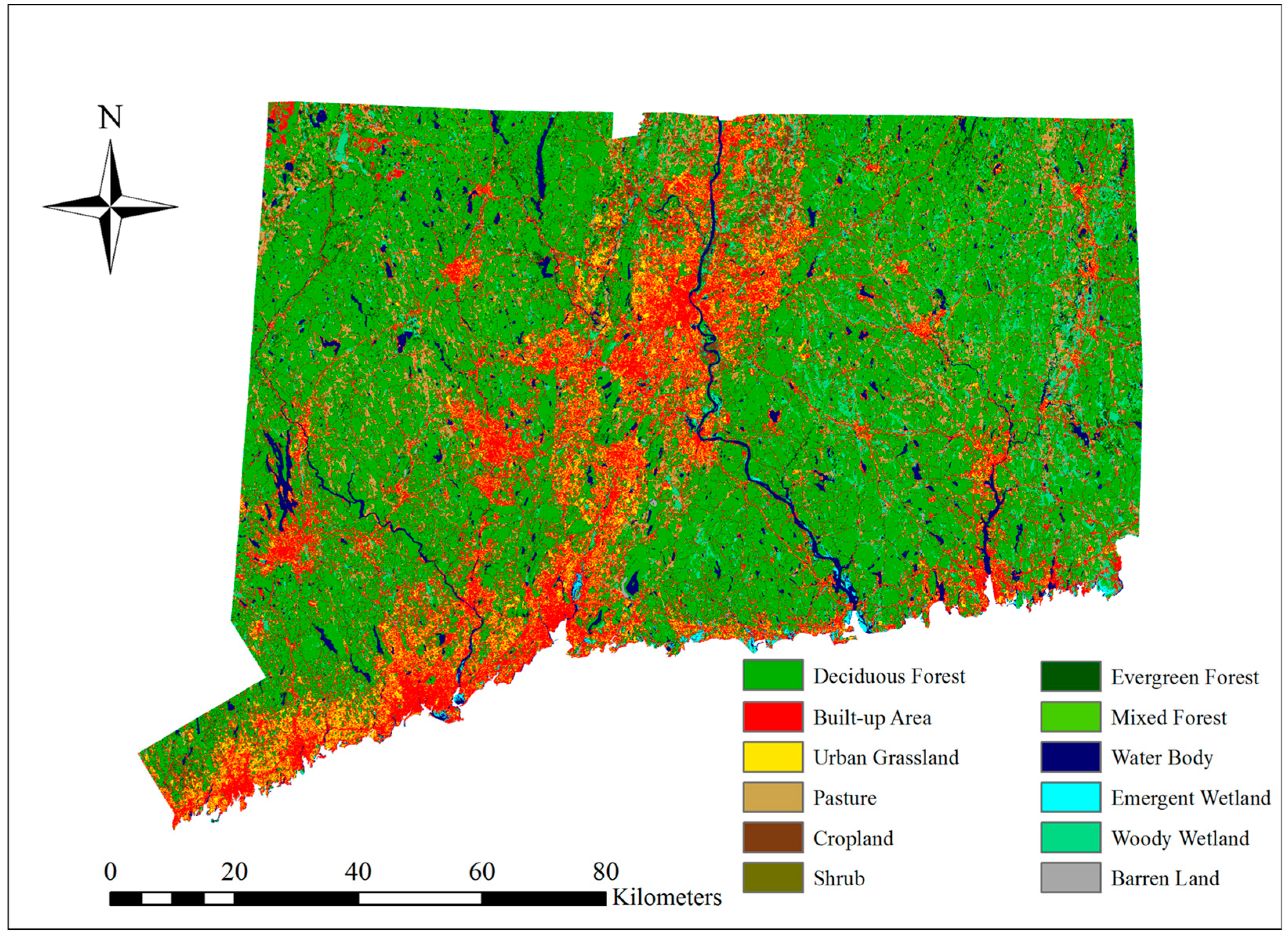
3.1. Observed LULC Changes
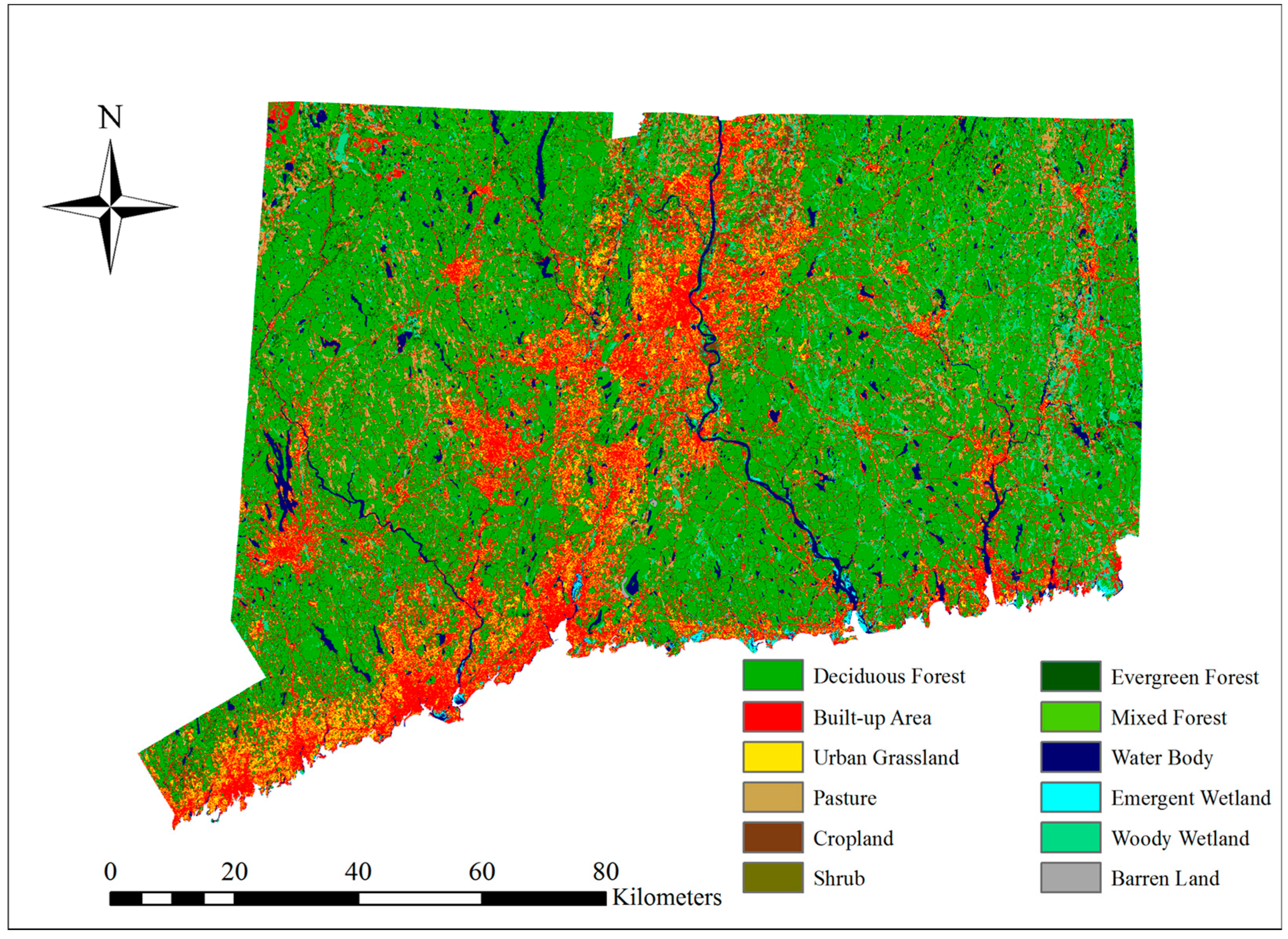
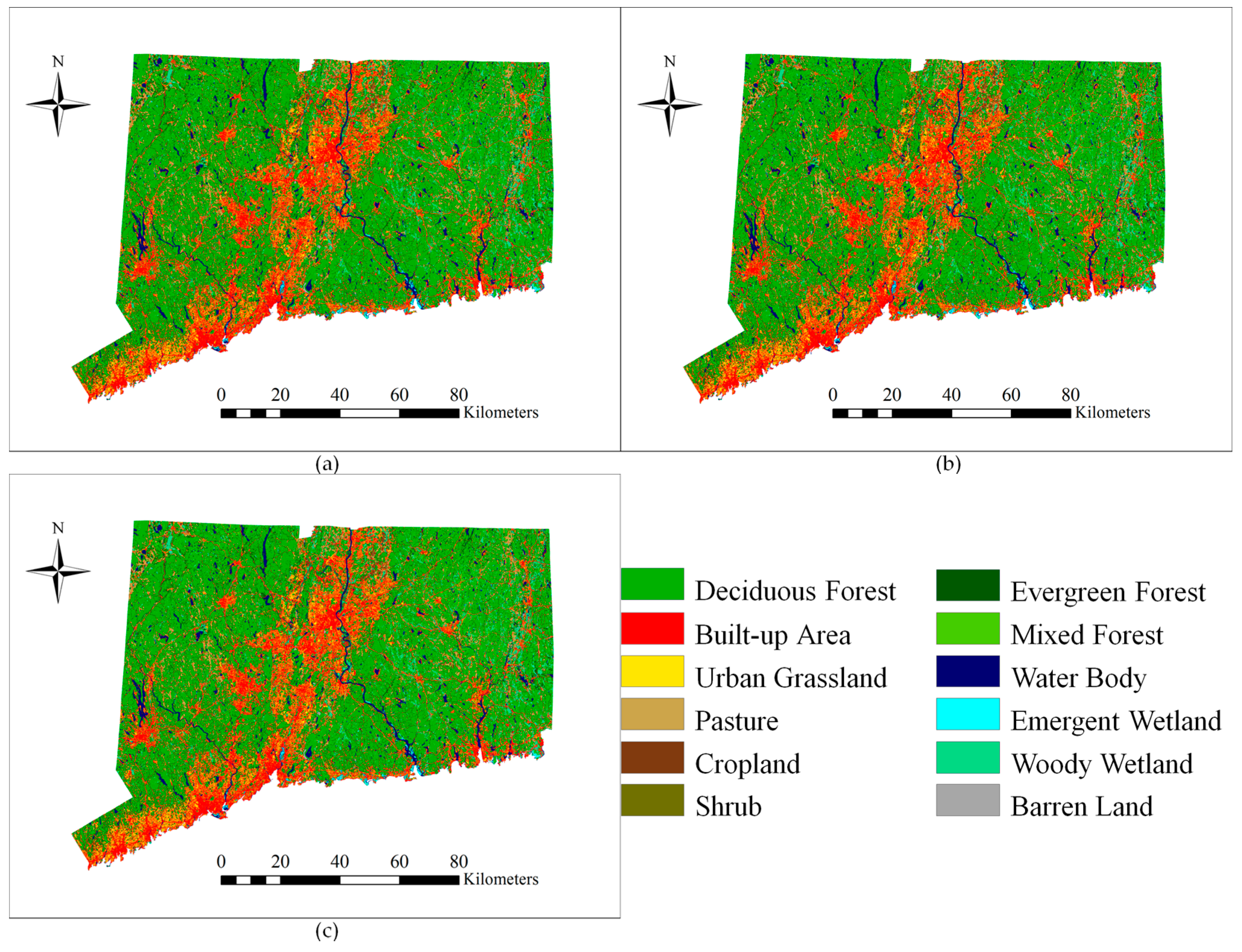
The results of LULC distribution for the years 1996, 2001 and 2006 show that deciduous forest was the dominant LULC class (from 48.04% to 47.67%) and the secondary class was built-up area (from 19.07% to 19.21%). There is a notable increasing trend for built-up area and agriculture classes, whereas a decreasing trend can be observed for forest classes (
Table 5).
According to
Table 6, the net deciduous forest loss for the period of 1996–2001 is 2289 ha. Drummond and Loveland [
26] indicated that the largest contribution to forest expansion was the forest gain from grassland and shrub during the 1973–2000 period in the eastern United States. The situation, however, is different in Connecticut. A large portion of the deciduous forest loss was due to the transition of deciduous forest to shrub, which was 1242 ha, equivalent to 54.26% of the total deciduous forest loss. Shrub’s gain was also the result of the transition from evergreen forest to shrub (5.89%). The reader will recall that the “succession” process occurs when cleared areas transit naturally from grassland to shrub and then to forests. The gain of forest from scrub/shrub is small relative to its loss to other LULC classes, and it is possible that some deforested areas were classified into shrub. After a longer period, the areas will go through enough succession to become forest again.
The transition of shrub to forest should be observed in the next period (
Table 7). The transition from deciduous forest to agriculture land (including cropland and pasture) was the second largest change, which was 527 ha, equivalent to 23.02% of the total deciduous forest loss. The third was the conversion of deciduous forest into built-up area, that is 229 ha and equivalent to 10% of the total deciduous loss. The shrub class increased by 1262 ha in total for this period, and the pasture class increased by 331 ha in total, which was mostly from deciduous forest. In some places, however, pasture was converted into built-up area (38 ha), cropland (36 ha) and urban grassland (31 ha). Alig et al. [
69] and Tyrrell et al. [
28] indicated that some pasture lands were relinquished to reforestation and frequently to development in the Northeastern United States, following the decline of the dairy industry. The situation was similar in Connecticut, but pasture lands were more likely to be converted into built-up area and cropland instead of forest. Cropland increased by 169 ha in total, which was mostly from deciduous forest. In some places, however, cropland was converted into barren land (22 ha) and built-up area (7 ha).
For the period 2001–2006, the net deciduous forest loss was 2505 ha. Most of the deciduous forest was lost to built-up area, which was 1206 ha, equivalent to 48.14% of the total deciduous loss. The transition of deciduous forest to agriculture land (including cropland and Pasture) was the second largest in area at 783 ha or equivalent to 31.26% of the total deciduous loss. Third was the conversion into urban grassland, accounting for 13.25% of the total deciduous loss. The built-up area increased by 1547 ha in total for this period, and most of the gain (77.76%) resulted from the conversion of deciduous forest. The urban grassland class increased by 408 ha, also gaining from deciduous forest. It was observed that urban and agricultural expansions were the main driving forces for the changes in forest because of increased population, residential development and proximity to rapidly developing area during this period. The pasture class increased by 601 ha, mostly from deciduous forest, evergreen forest and mixed forest. In some places, however, pasture was converted into shrub (71 ha), cropland (41 ha) and built-up area (33 ha). Similar to the LULC changes in the previous period, the loss of pasture land was related to the change in dairy production.
Figure 6 shows how widespread the loss of deciduous forest from 1996 to 2006 was. The colored areas indicate where forest cover was lost.
3.2. Driving Factor Analysis
The overall CVC values are shown in
Table 8. This index describes the quantitative levels of associations of a driving factor with individual or all LULC classes. The overall CVC value considers all variables together by Equation (1). It can help us understand how much a factor can influence LULC change. Normally, CVCs of about 0.15 or higher are considered important [
70]. Although some factors’ overall CVCs are slightly less than 0.15, the factors may still have relatively high values for specific LULC classes. For example, the overall CVC of elevation is 0.132, but CVCs of elevation with deciduous forest, urban grassland, built-up area, and water body are 0.306, 0.179, 0.170, and 0.156, respectively. This means that elevation has a relatively stronger relationship with the spread of deciduous forest. CVCs of slope with water, deciduous forest, and woody wetlands are 0.336, 0.307, and 0.158, respectively. Thus, slope appears to be a good predictor of water bodies and wetlands, because they are generally located in flat and lower areas. The results also indicate that slope has a similar influence on deciduous forest, as does elevation. Because of the small variation in topography and socioeconomic complexity, however, elevation and slope do not have large overall CVCs [
71,
72].
The DTR variable shows a good association with built-up area and deciduous forest, with CVCs of 0.483 and 0.356, respectively. Hence, it is one of the major drivers for forest change, because the development of new settlements tends to occur near existing settlements. CVCs of the DTC variable with cropland, pastures, and deciduous forest are 0.290, 0.176, and 0.142, respectively; and the CVCs of the DTP variable with pastures, deciduous forest, and built-up area are 0.501, 0.176, and 0.164, respectively. Both factors are useful in explaining the conversion of deciduous forest into agriculture. The CVC value of DTW with deciduous forest is 0.174, which indicates that the proximity to water has a relationship with forest change. CVCs of DTD with built-up area and urban grassland are 0.294 and 0.285, respectively, meaning that urban sprawl has a strong relationship with deforestation. The CVC of DTM with evergreen forest is 0.155. Similarly, the CVC of DTE with mixed forest is 0.154. Those two factors can partly explain the conversion between mixed forest and evergreen forest. CVCs of DTB with built-up area, deciduous forest, and urban grassland are 0.430, 0.380 and 0.266, respectively.
Apparently, the DTB plays an important role in LULC change in Connecticut, such as reduction in forest areas and increased new settlement areas. Not surprisingly, deforestation is stronger at places closer to an existing built-up area. And the areas closer to existing built-up areas have higher probabilities of conversion to built-up areas. CVC values of population with deciduous forest, urban grassland, and built-up area are 0.209, 0.205 and 0.197, respectively. These results indicate that population pressure should be an important driving force to the increase of the built-up area and conversion of forest to other land use classes.
3.3. Model Validation
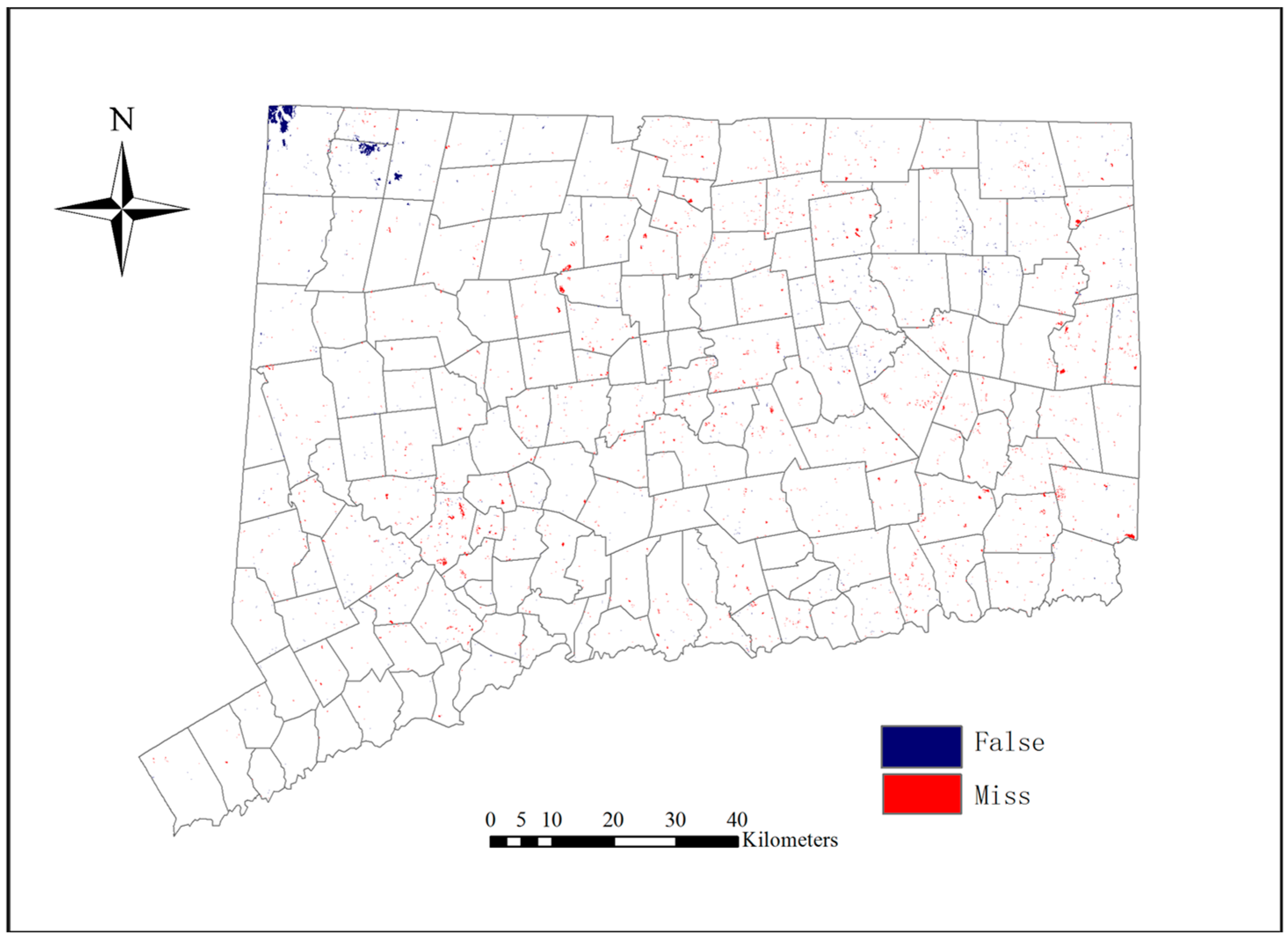
A predicted map for model validation was created for the year 2006 using the transition probability matrix estimated from the 1996 and 2001 LULC maps. This validation aimed to evaluate the quality of the predicted land use map in comparison with the real land use map. A three-way cross-tabulation between the real 2001 map, the real 2006 map, and the predicted 2006 map was conducted for the validation. By comparing the real 2001 LULC map with the real 2006 LULC map, the actual change locations can be discovered, and by comparing the real 2001 LULC map with the predicted 2006 LULC map, the predicted change locations can also be identified. Then, comparing the actual change locations with the predicted change locations can find out where the prediction errors are located.
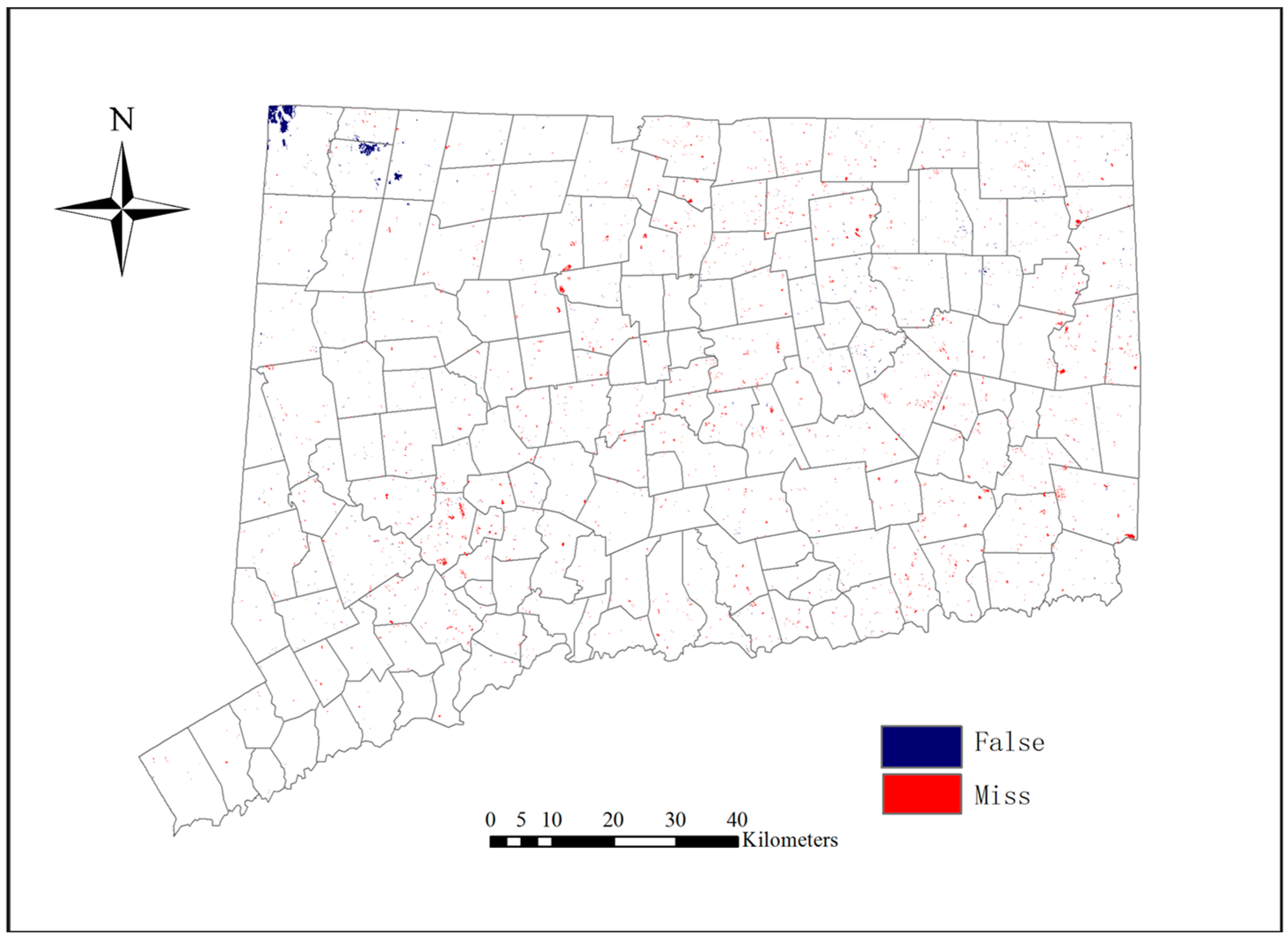
Figure 6 indicates that the LULC change model worked effectively because the predicted map and the real map for 2006 coincided in most places and only minor differences occurred. The difference map is shown in
Figure 7.
IDRISI Selva [
73] provides a Validation Module, which can measure the agreement between two categorical maps. Cohen’s kappa coefficient is a commonly used index, which measures inter-rater agreement for qualitative (categorical) items. Kappa is always less than or equal to 1. A value of 1 means perfect agreement. Pontius [
74,
75] suggested, however, that the standard Kappa coefficient cannot provide enough information because it does not have ability to distinguish the quantification error with the location error [
74,
75]. In order to solve this problem, two more kappa statistics are provided: Kappa for no information (Kno), and Kappa for grid-cell level location (Klocation). The Kno statistic is an improved general statistic over the traditional Kappa Index of Agreement (Kstandard), as it penalizes large quantity errors and rewards correct location classifications. Klocation indicates how well the grid cells are located on the landscape. In order to calculate those Kappa statistics, the real 2006 LULC map and the predicted 2006 LULC map were used as the reference map and the comparison map, respectively. Furthermore, five statistics were calculated to indicate how well the comparison map agrees with the reference map [
67,
68]: agreement due to chance (AgreementChance), agreement due to quantity (AgreementQuantity), agreement due to location at the grid cell level (AgreementGridcell), disagreement due to location at the grid cell level (DisagreeGridcell), and disagreement due to quantity (DisagreeQuantity) (
Table 9).
The results in
Table 9 indicate that the MLP_MC model has a very high capability to predict future LULC changes. It is also important to note that the DisagreeGridcell and DisagreeQuantity indices can help us understand the predicted results. In
Table 9, the DisagreeGridcell is larger than the DisagreeQuantity. This means that the model has a higher ability to predict the LULC changes in quantity than in location in the study area. To reduce the DisagreeGridcell value, more explanatory variables about location should be considered in further studies.
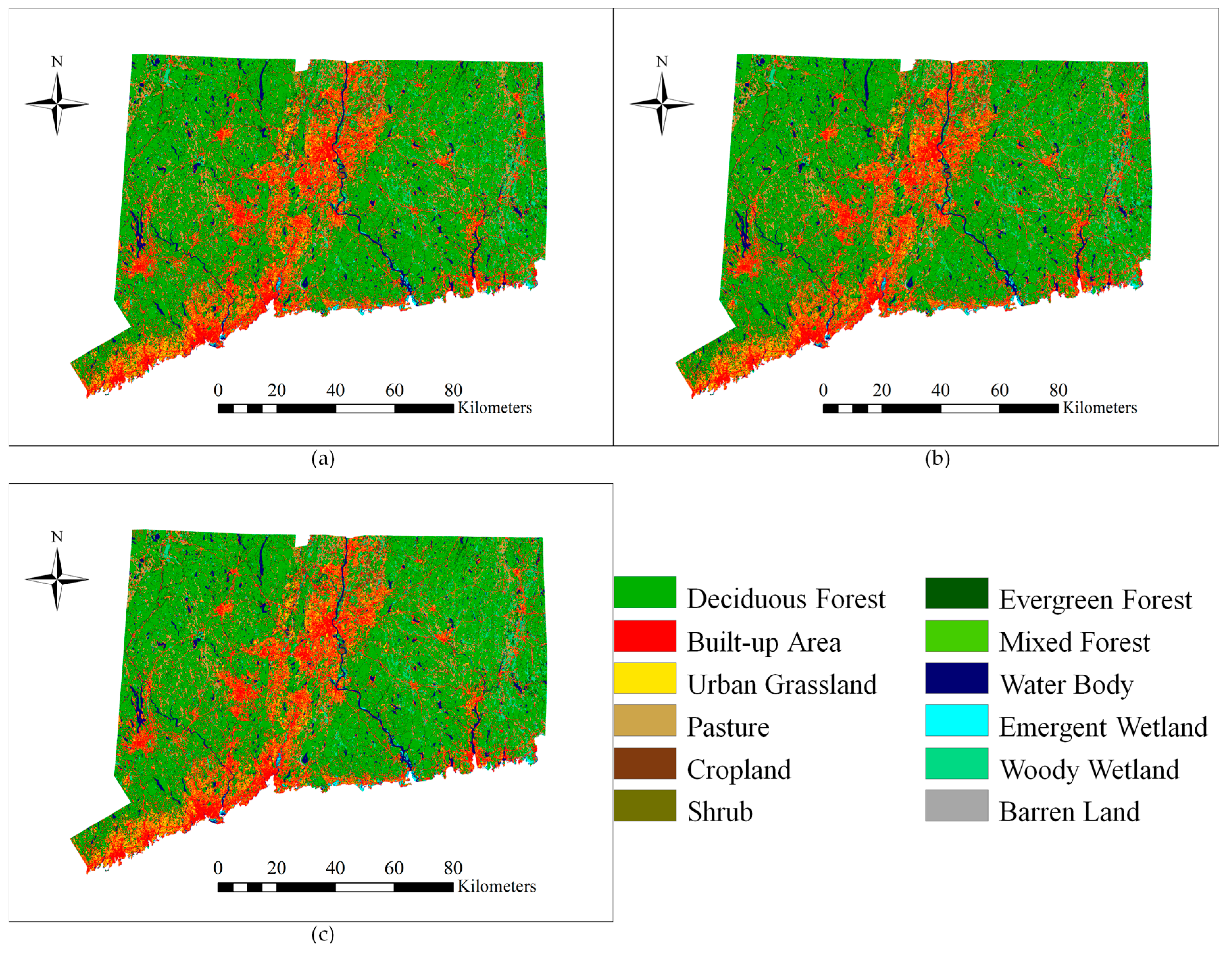
3.4. Change Predictions
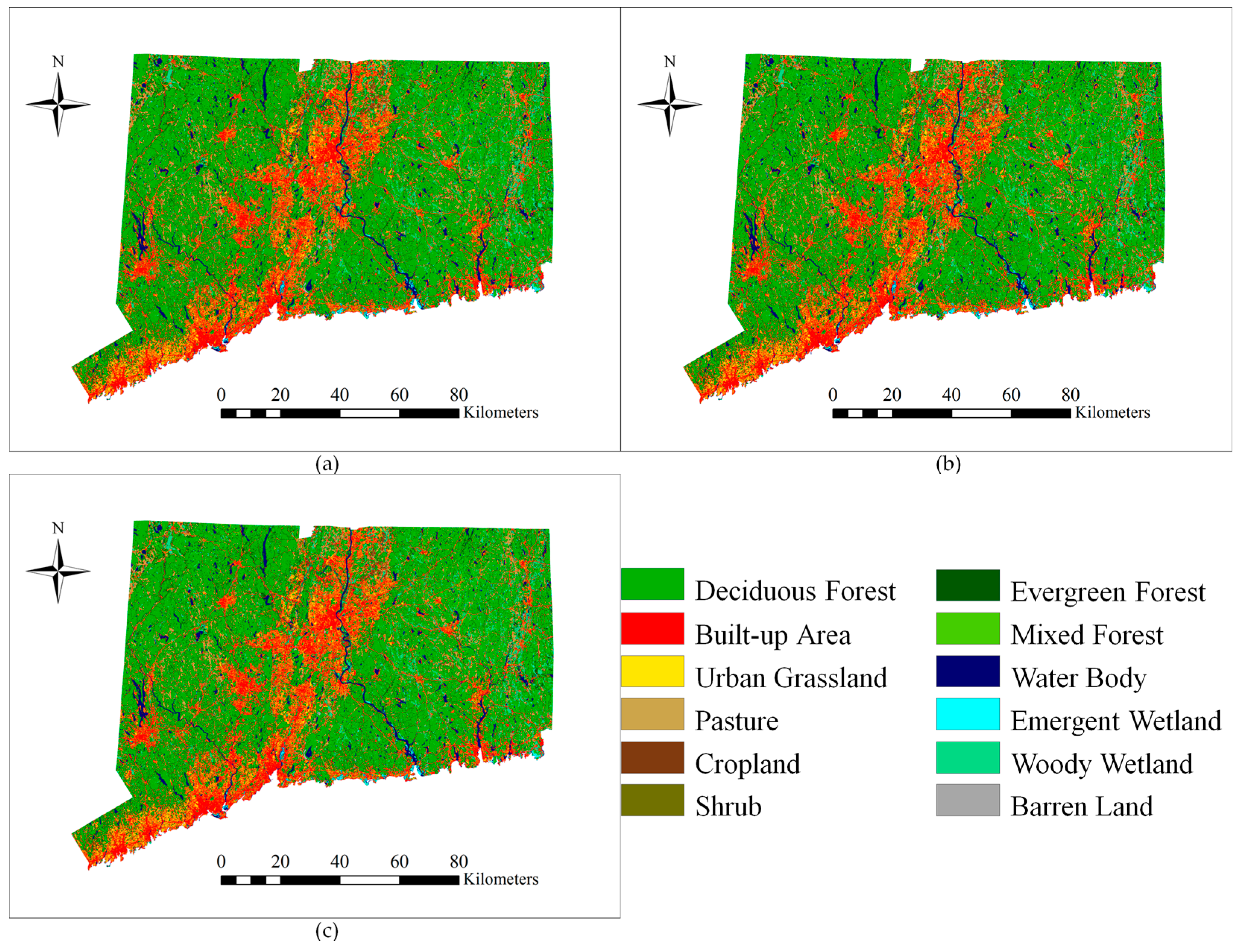
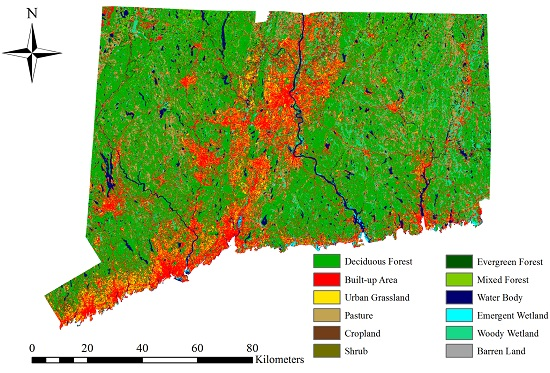
The LULC map for the year 2018 (
Figure 8) was predicted using the LULC maps of 2001 and 2006. The results of a predicted LULC scenario (
Figure 9) show an obvious increase in developed area (built-up area and urban grassland) and agriculture land (cropland and pasture). In addition, it shows a drastic decrease in forest area (deciduous, mixed, and evergreen forest). The model predicts that the study area will lose 5535 ha deciduous forest and gain 3502 ha in built-up area from 2006 to 2018. The conversion of deciduous forest into built-up area is predicted to be 2884 ha in the 12 years by 2018, accounting for 52.10% of the total deciduous forest loss. Urban growth will continue in the future, but the rate of deforestation will slow down. Because of an increasing demand for agriculture area driven by population pressure, pasture and cropland will increase by 2325 ha and 253 ha, respectively. The conversion of deciduous forest into agriculture land is predicted to be 1903 ha by 2018, equivalent to 34.38% of the total deciduous forest loss. In addition, areas near the built-up area and agriculture land appear to be vulnerable to conversion. These areas may need more protection measures if the total forest area is to be maintained in the future. Mature forest stands are necessary to maintain a diversity of wildlife and native plants.
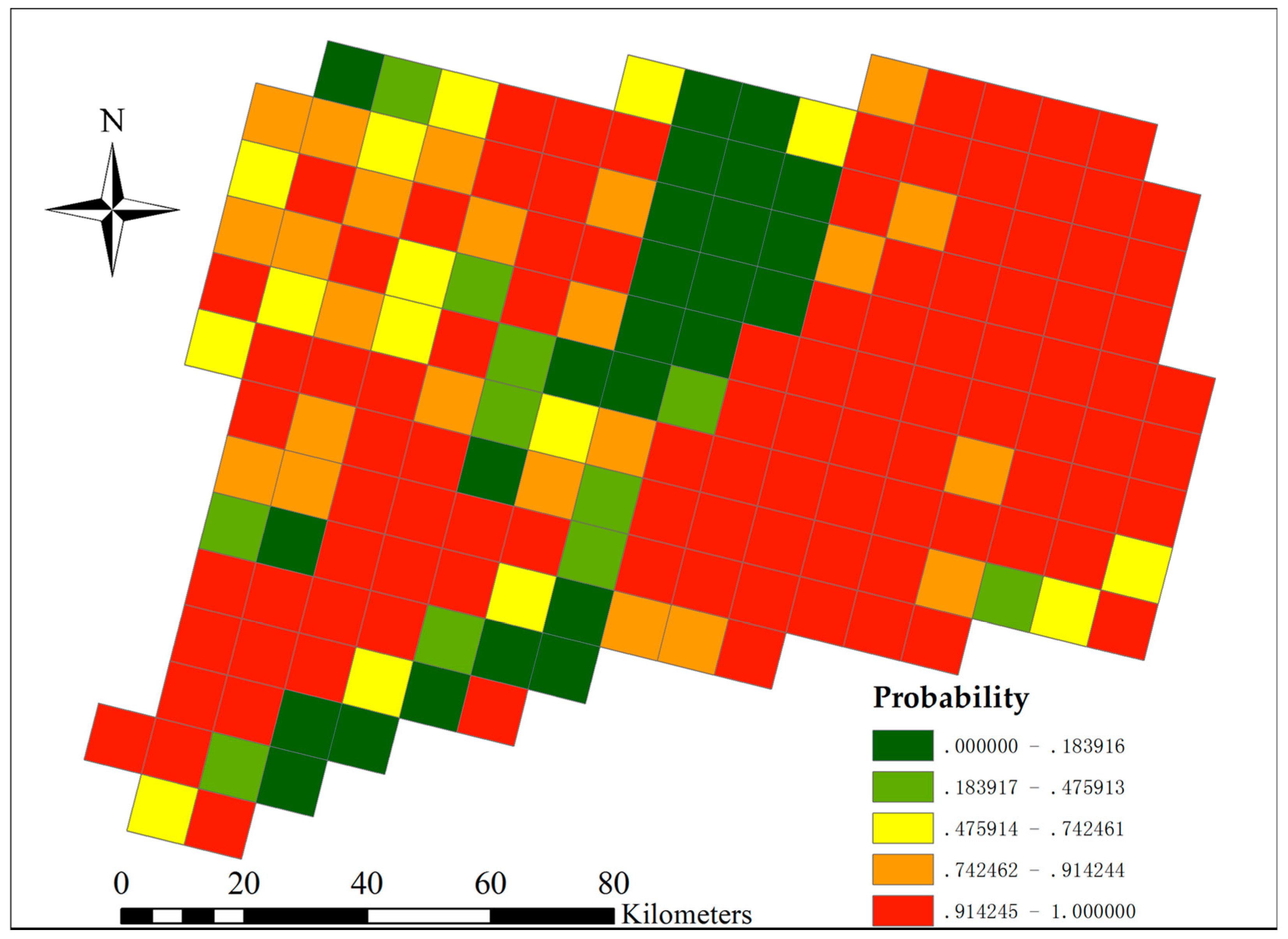
3.5. Landscape Scale Japanese Barberry Invasion Probability Affected by Short-Term Land Use Changes
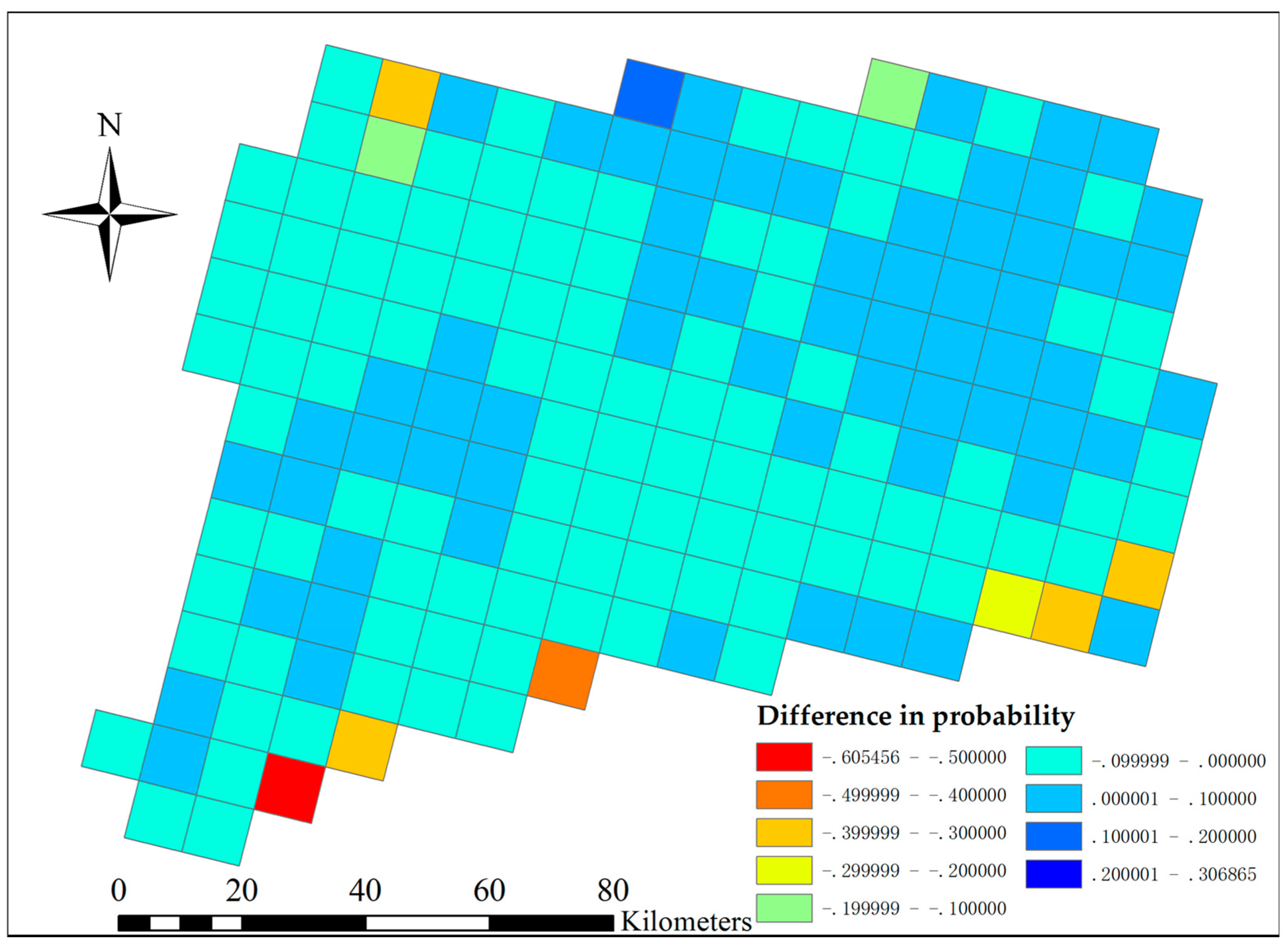
The projected LULC changes by 2018 yielded subtle, but measurable changes in the predicted probability of Japanese barberry. The northwest and south-central areas of the state had slightly reduced probabilities of barberry presence, whereas the north-central and northeast areas of the state had slightly higher risk with projected LULC change (
Figure 10). The probability change for each grid cell was generally 0.1 more or less than the 2006 baseline estimate, with only about 6% of the cells falling outside of that range. The changes in invasion risk do not negate the overall risk, however; areas that had high probability of presence (e.g., most areas except the Connecticut River Valley) in 2006 continue to be at high risk in the future (
Figure 11). Landscape-scale loss in the percentage of deciduous and mixed forest cover was significantly correlated with decreased invasion risk for this species (r = 0.16,
p = 0.03) and gain in developed cover was correlated with increased risk (r = 0.64,
p < 0.0001).
4. Discussions
Changes in LULC will influence forest distribution in the study area. Since urbanization can increase local temperature (e.g., the urban heat island effect), human activities, and damage to the natural environment, it is possible to see a shift in species composition between urban and rural areas. Human activities in urban areas also change the characteristics of soils. For example, soils in urban areas have higher concentrations of heavy metals and organic matter than in rural areas. For some species, this kind of change could reduce habitat suitability. Moreover, forests adjacent to residential areas tend to have a higher number of invasive species compared to those surrounded by industrial lands.
In the Northeast, dairy production was dominated by small dairies, which disappeared rapidly because of increasing mechanization and industry competition [
76]. Therefore, decreasing dairy production led to pasture’s natural conversion to shrub. Cropland increased by 217 ha in total, mostly converted from deciduous forest and pasture. Rapidly increasing population brought the pressure to existing agriculture land by replacing forest.
Forest utilization, such as timber harvesting, extraction of non-timber products, construction of logging and transport roads and facilities for logging camps, and conversion of natural forest to plantations, may have direct and indirect negative impacts on forest biodiversity by promoting the invasion of alien species. Some studies have shown that forest fragmentation may accelerate the spread of invasive species [
32,
36]. Simply, a high proportion of evergreen forest in a landscape tends to deter invasive plants [
20,
64]. But the portion of evergreen forest area is relatively low in Connecticut; and based on our prediction, some evergreen forest is expected to be lost (272 ha) by 2018. Therefore, to stem the spread of invasive plants, it would make sense for policy makers to take actions to preserve evergreen forest in the region.
Moreover, road systems provide essential access to forests for timber extraction. A good road system is an important requirement for sustainable forest management. But without quality design and maintenance, roads are often the cause of a variety of environmental problems associated with forest harvesting operations [
77]. In some situations, roads may also initiate or accelerate the invasion of non-native species that ultimately displace native species. Additionally, the increased level of human activities in previously inaccessible areas (activities that are facilitated by roads) can cause environmental problems, including the possible introduction of alien species [
78]. Mosher et al. [
18] indicated that the greatest incidence of invasion is in post-agricultural settings, e.g., fields that are currently abandoned or have reverted to forest. Our study shows that portions of the pasture land and cropland were converted to urban grassland during 1996 to 2006 in Connecticut, which might increase the current and future richness of invasive species.
5. Conclusions
In this study, LULC change in Connecticut from 1996 to 2006 was analyzed and LULC change from 2006 to 2018 was predicted using the MLP_MC model. In addition, the effect of LULC change on the invasion risk of an abundant invasive shrub, Japanese barberry, was studied. The results show that the total area of forests, especially deciduous forest, has been decreasing. That decrease is primarily caused by urban development and other human activity in the study area. We also found that forest areas near built-up areas and agriculture lands appear to be more vulnerable to conversion. These predicted LULC changes translated into subtle changes in the invasion risk by the invasive shrub Japanese barberry. While the magnitude of change in Japanese barberry risk was low over this relatively short time-scale, it could be compounded by longer-term changes in LULC and changes in climate. Future work should focus on these interacting global change factors and the generalization of their effects on invasive plants in the region and beyond.
In summary, this study provides reliable LULC data, which is useful for the study of invasive species in the Connecticut region. The information is also useful for early detection and in the future refinement of conservation policy aimed at reducing the spread of invasive species. Our findings suggest that forest conversion should be controlled and managed in service of invasive species control. In addition, our research indicates that the MLP_MC model can predict future LULC with a reasonable accuracy. Nevertheless, it is important to note that predicting the actual location of LULC change is challenging. Explanatory variables about locations are usually not sufficient and important social economical drivers vary with time. Future work will consider dealing with these limitations.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}