Linking Land Cover Data and Crop Yields for Mapping and Assessment of Pollination Services in Europe

Abstract
:1. Introduction
2. Methods
2.1. General Outline of the Pollination Supply Model
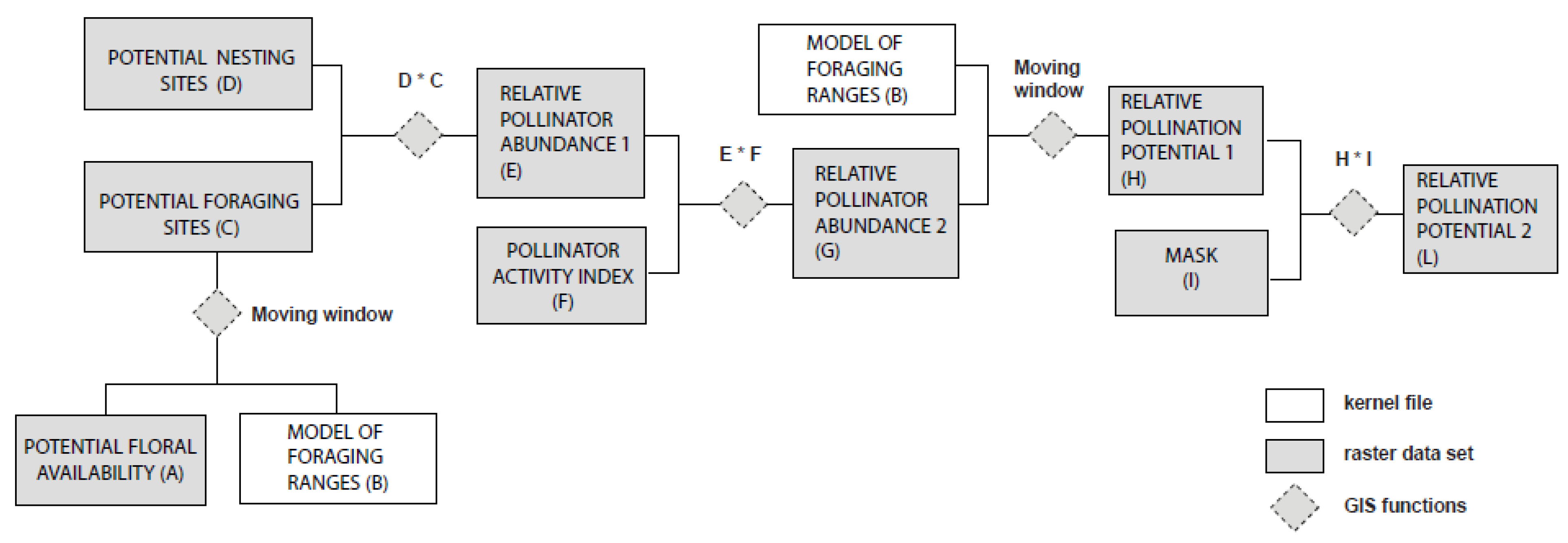
2.2. Nesting Suitability and Floral Availability
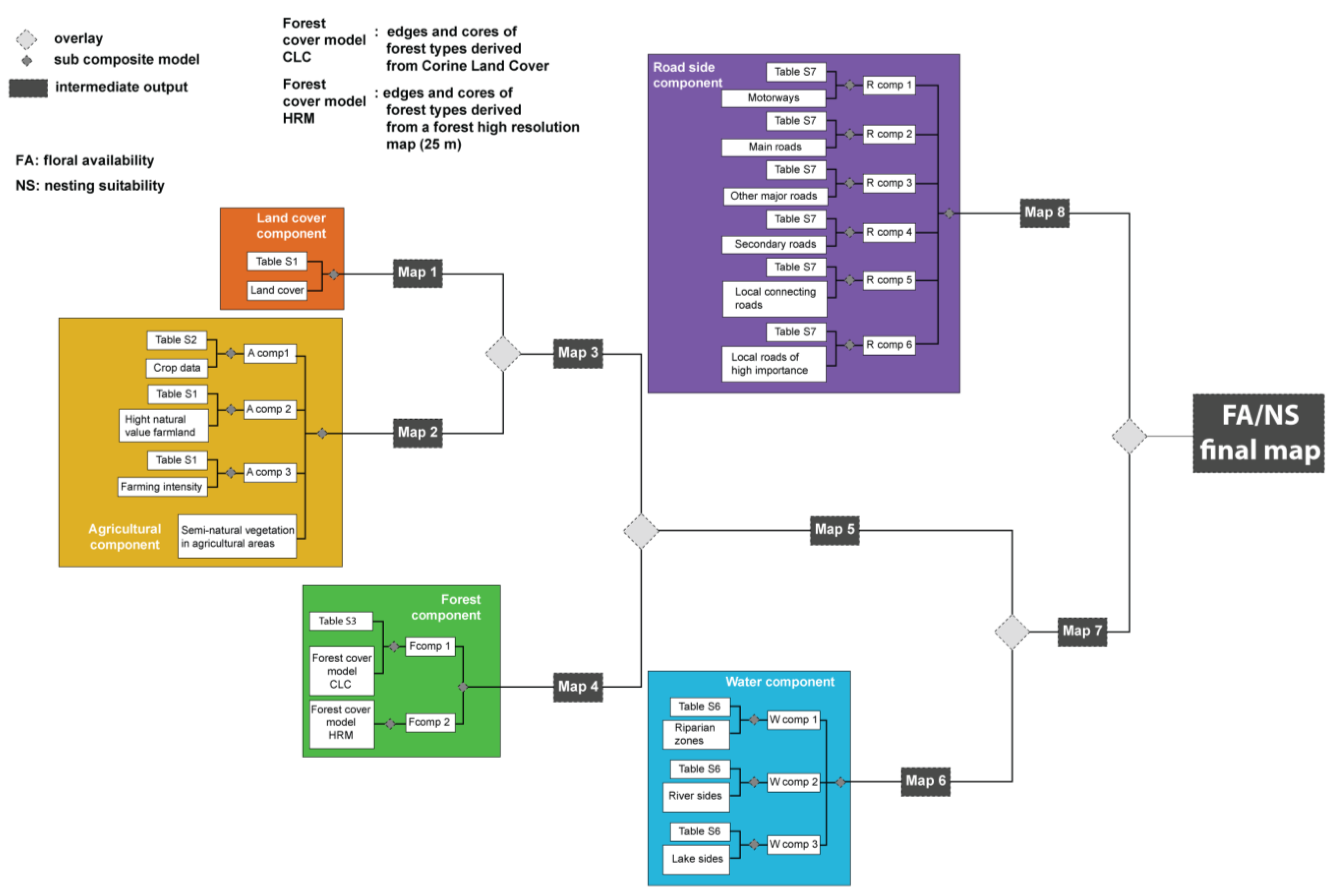

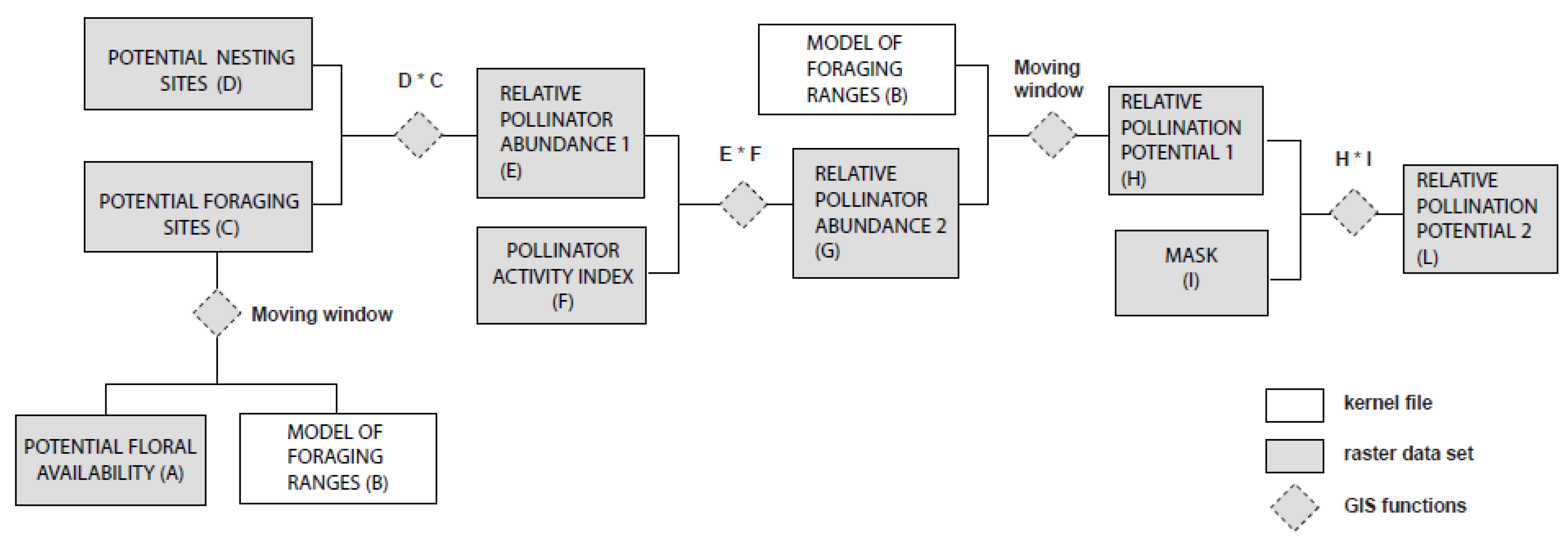{kind=link}
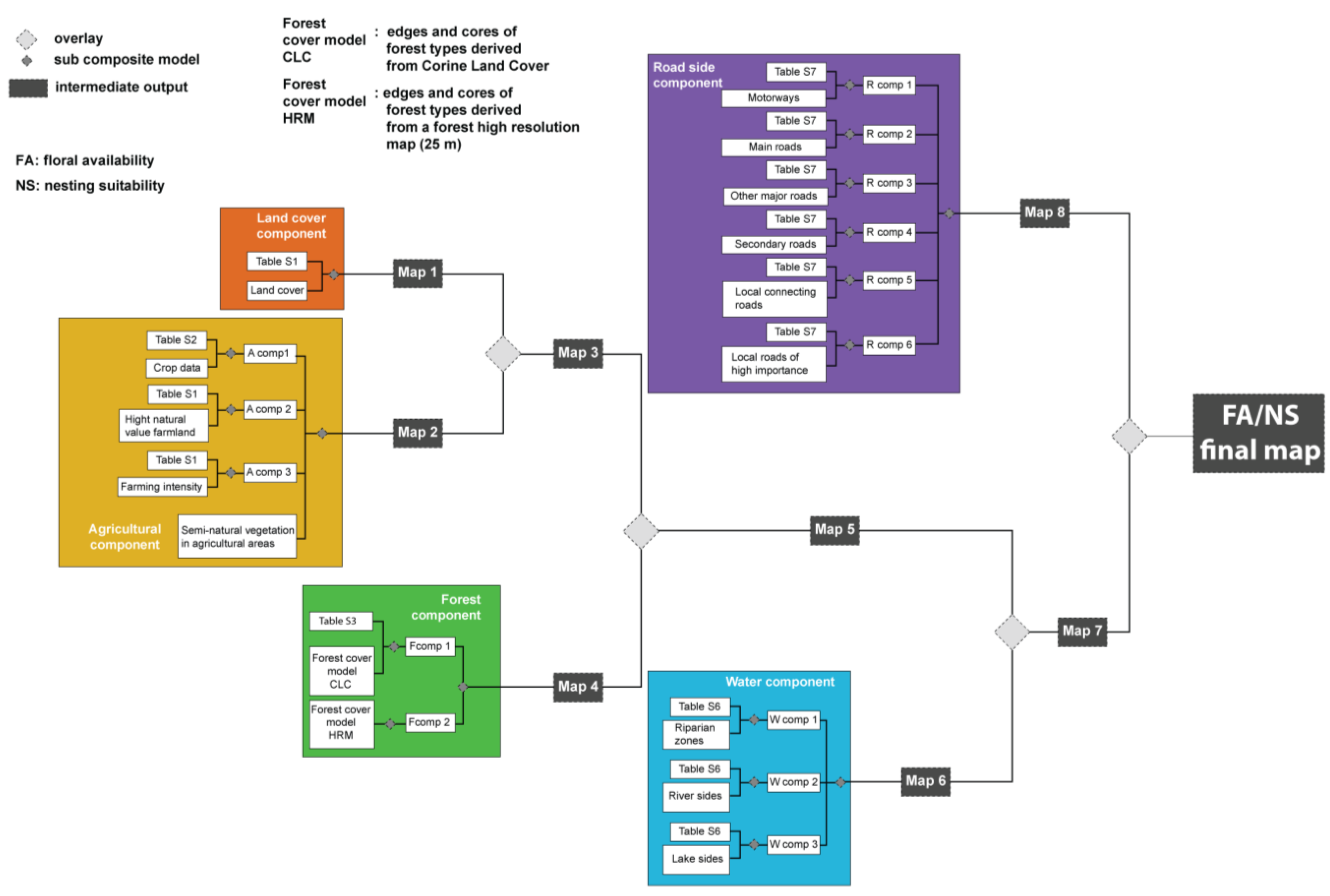{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Data Description | Resolution |
|---|---|---|
| Land cover | CORINE Land Cover 2000 (CLC2000) raster data—version 13 (02/2010) Source: EEA, 2010 Map of the European environmental landscape based on interpretation of satellite images with land cover types in 44 standard classes. | 100 m |
| Agricultural land use | Crop yield data The CAPRI model results in crop yield statistics for homogeneous clusters of 1 km2 pixels (HSMU), identified on the basis of the Farm Structure Survey regions (NUTS 2 or 3, depending on the Member State, EUROSTAT 2003), land cover (CLC2000), soil mapping units (European Soil Database V2.0, European Commission, 2004) and slope [26]. | 1,000 m |
| Olive farming data [28] | 100 m | |
| High Nature Value Farmland (HNV) data. HNV is defined as areas in Europe where agriculture is a major (usually the dominant) land use and where that agriculture supports, or is associated with, either a high species and habitat diversity or the presence of species of European conservation concern, or both. Source: JRC [29] | 100 m | |
| Presence of semi-natural vegetation at European scale. Source JRC, unpublished | 100 m | |
| Road network | TeleAtlas® MultiNet™ dataset (version 2007.10) | |
| Water | Riparian zones [30] | 25 m |
| CCM2 data (river network and small lakes) Source: JRC/EEA | ||
| CLC2000 data (main lakes) Source: EEA | ||
| Forest cover | CLC2000 data Source: EEA | 100 m |
| Pan-European Forest/Non-Forest Map 2006, Source: JRC [31] | 25 m | |
| Activity index | AGRI4CAST interpolated grid [32] | 25 km |
2.2.1. Cropland
2.2.2. Forest
2.2.3. Riparian Zones
2.2.4. Road Sides
2.3. Foraging Range Model
2.4. Activity
2.5. Regional Pollination Deficit
2.6. Benefit for Crop Production
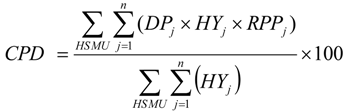
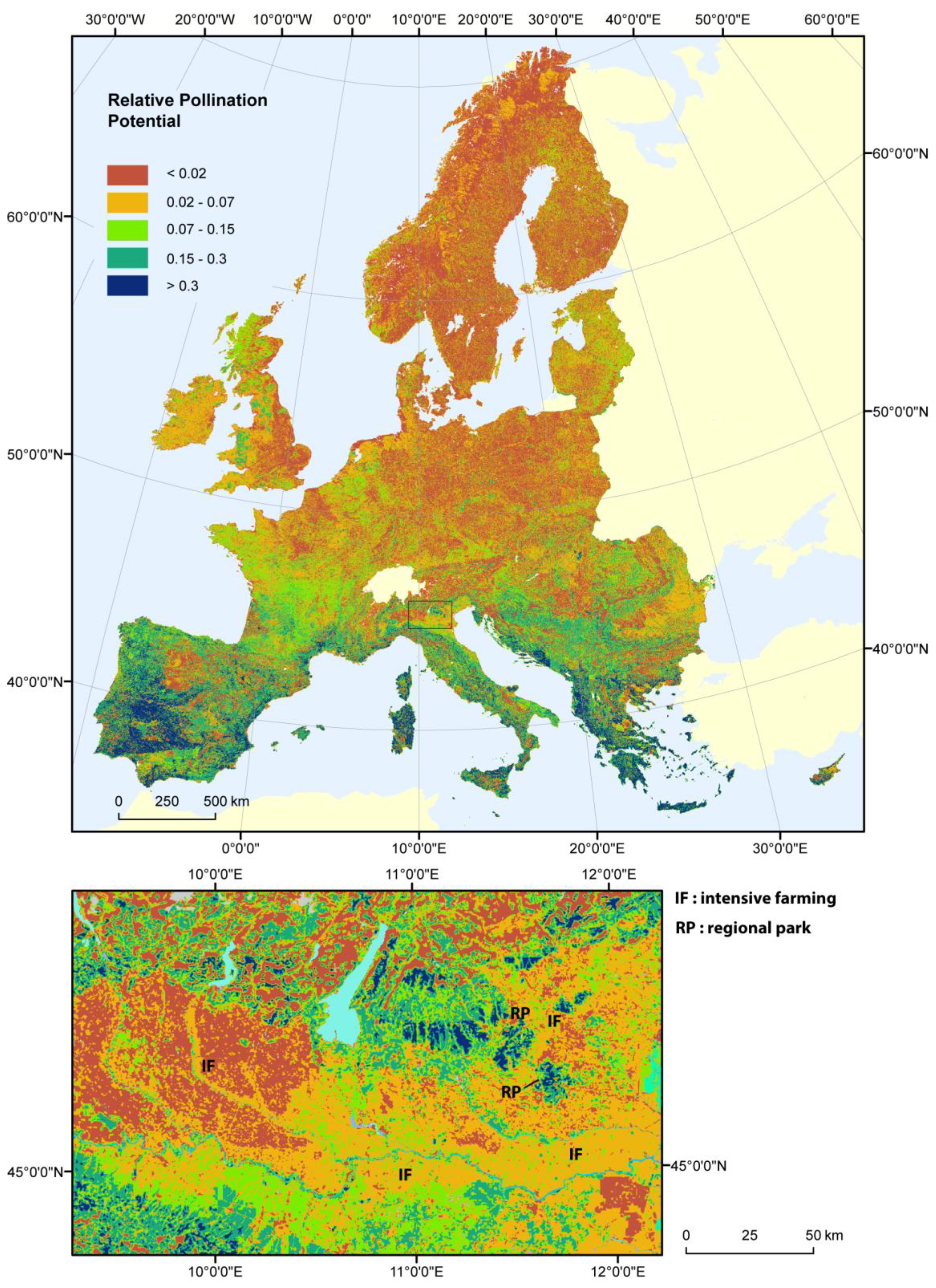
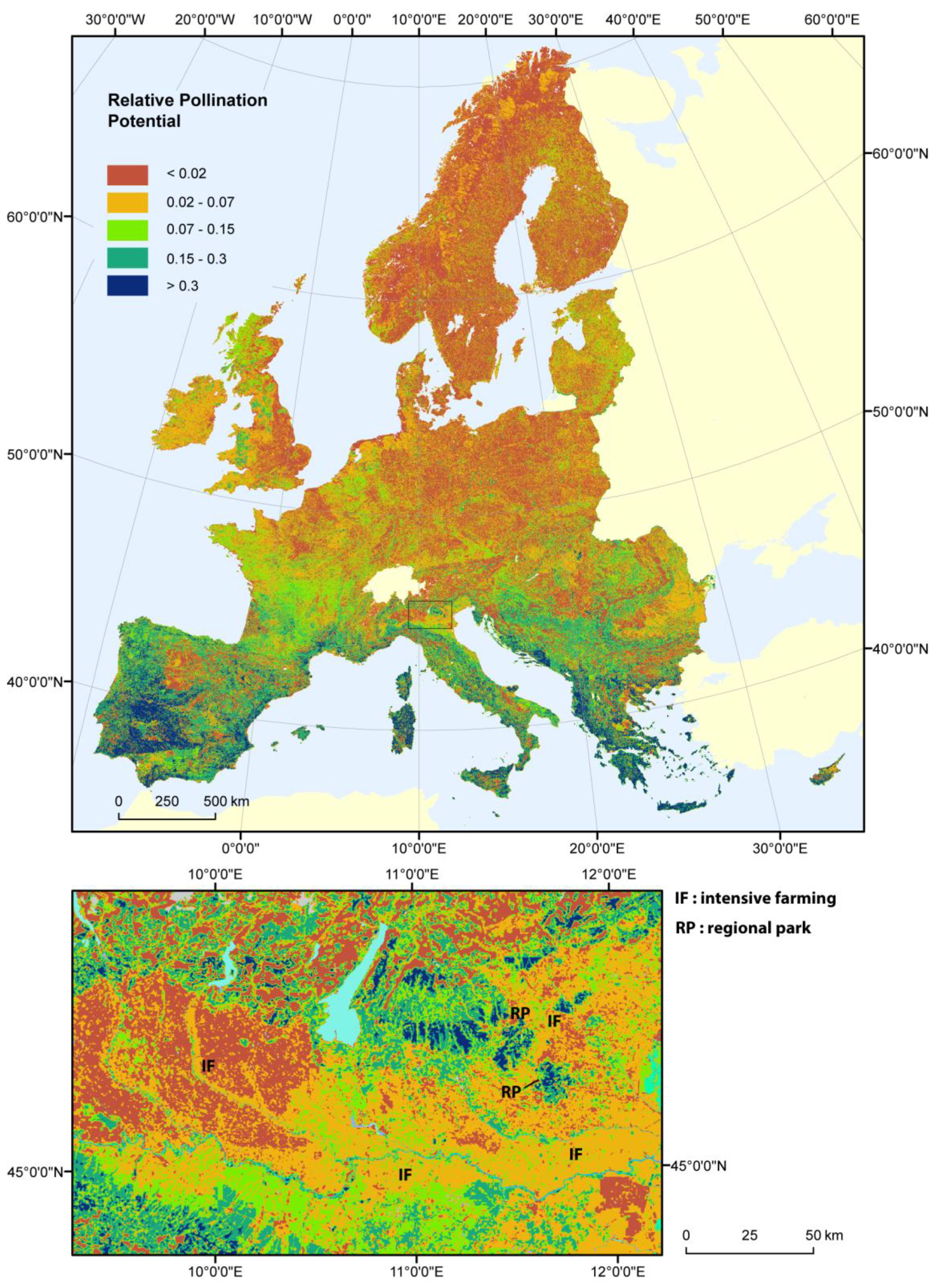
3. Results
3.1. Relative Pollination Potential
3.2. Benefits of Pollination for Crop Yield
| Country | CPD (%) (RPP = 1) | CPD (%) (RPP = 1) | CPD (%) (RPP) |
|---|---|---|---|
| Source: EUROSTAT | Source: CAPRI | Source: CAPRI | |
| Austria | 47.7 | 45.0 | 3.83 |
| Belgium | 26.6 | 39.7 | 3.26 |
| Bulgaria | 30.8 | 23.4 | 1.78 |
| Croatia | 45.7 | ||
| Cyprus | 23.9 | ||
| Czech Republic | 28.8 | 27.4 | 1.29 |
| Denmark | 26.0 | 25.1 | 0.79 |
| Estonia | 26.0 | 24.2 | 1.15 |
| Finland | 34.1 | 20.9 | 0.71 |
| France | 32.3 | 29.1 | 2.82 |
| Germany | 30.7 | 29.3 | 1.28 |
| Greece | 34.8 | 20.1 | 4.11 |
| Hungary | 40.9 | 35.4 | 2.44 |
| Ireland | 8.7 | 29.3 | 1.11 |
| Italy | 30.7 | 23.9 | 2.95 |
| Latvia | 46.2 | 36.0 | 1.91 |
| Lithuania | 25.8 | 27.9 | 1.04 |
| Luxembourg | 36.8 | 51.4 | 5.14 |
| Malta | 43.6 | ||
| Netherlands | 32.5 | 32.3 | 2.71 |
| Poland | 45.3 | 45.0 | 2.09 |
| Portugal | 21.1 | 7.5 | 2.16 |
| Romania | 46.8 | 31.3 | 3.6 |
| Slovakia | 26.4 | 26.8 | 1.79 |
| Slovenia | 57.9 | 57.0 | 6.86 |
| Spain | 27.9 | 8.4 | 1.67 |
| Sweden | 28.9 | 26.8 | 0.78 |
| United Kingdom | 22.4 | 21.5 | 0.69 |
| European Union | 32.6 | 25.1 | 2.46 |
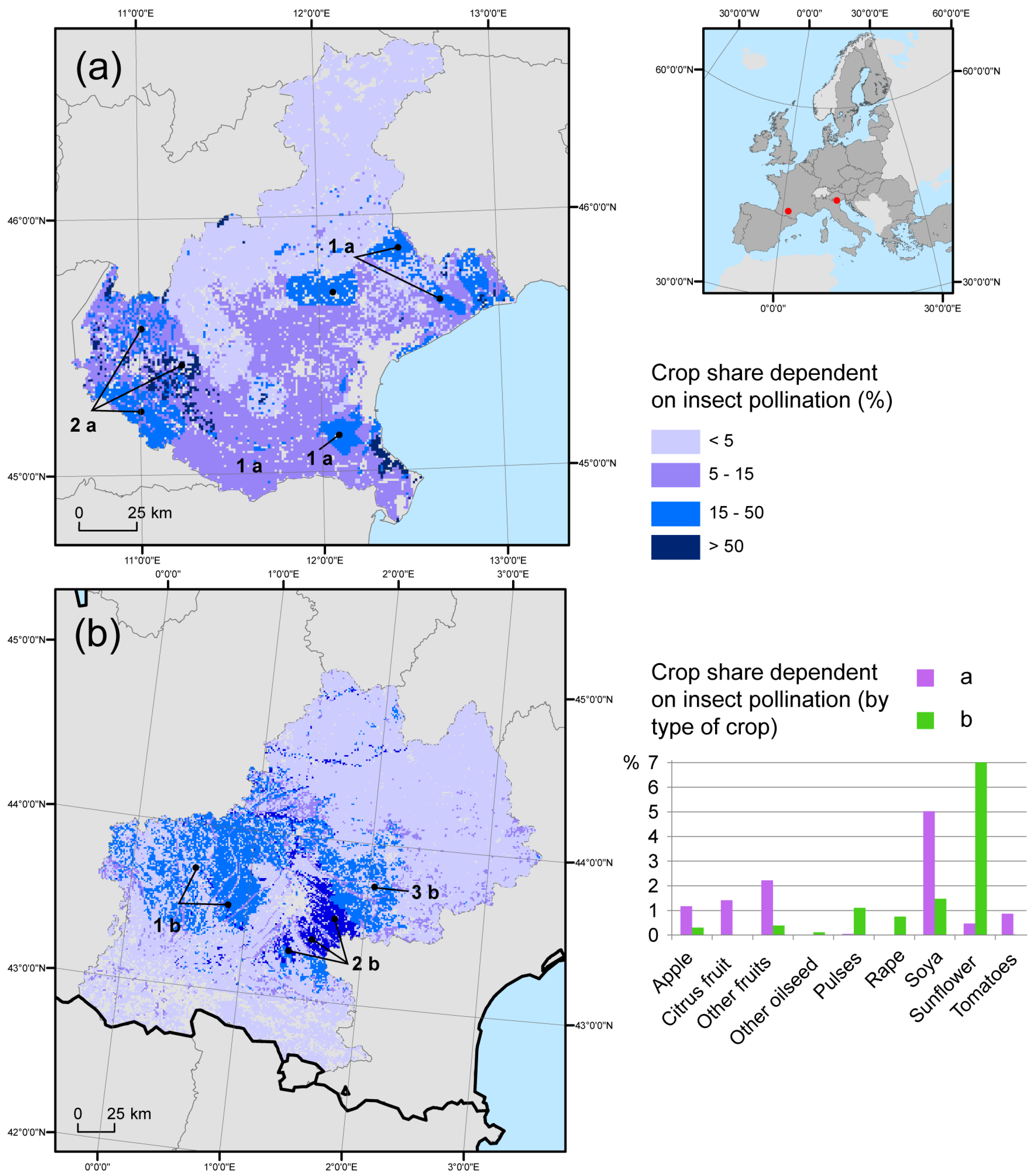
3.3. Regional Gap Analysis
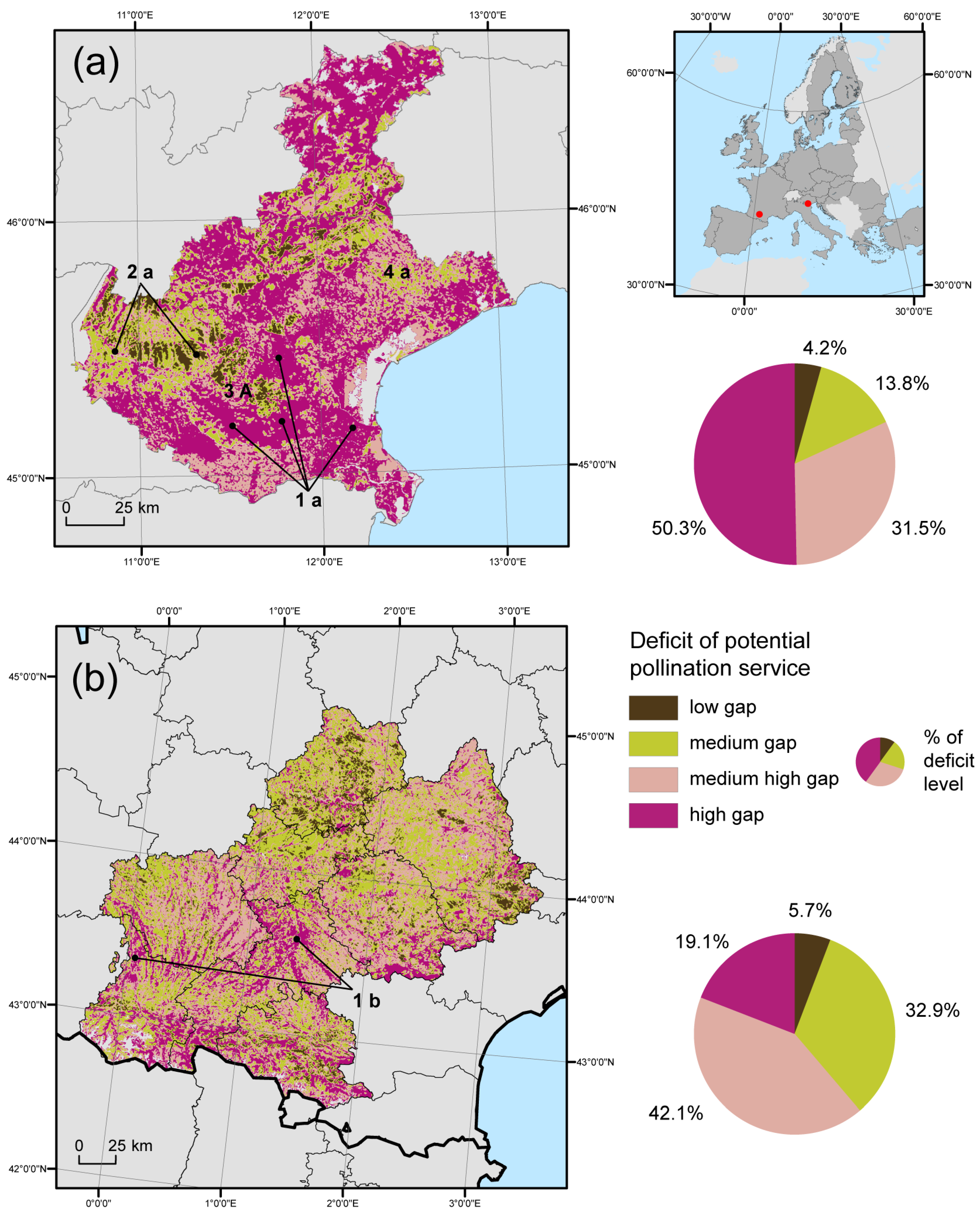
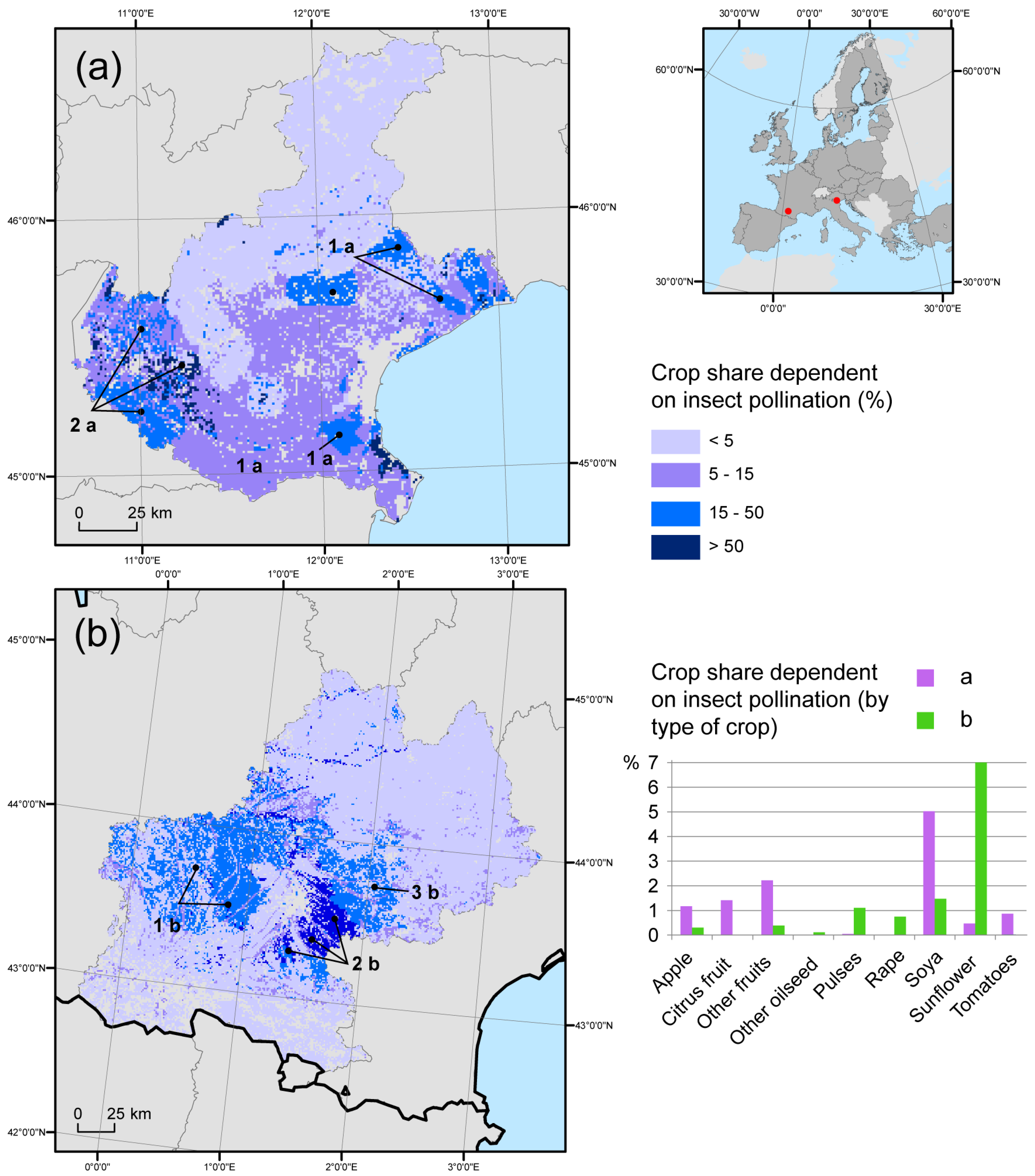
4. Discussion
4.1. Strengths and Applications
4.2. Limitations
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 340, 1608–1611. [Google Scholar]
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Aizen, M.A.; Garibaldi, L.A.; Cunningham, S.A.; Klein, A.M. Long-term global trends in crop yield and production reveal no current pollination shortage but increasing pollinator dependency. Curr. Biol. 2008, 18, 1572–1575. [Google Scholar] [CrossRef]
- Pimentel, D.; Wilson, C.; McCullum, C.; Huang, R.; Dwen, P.; Flack, J.; Tran, Q.; Saltman, T.; Cliff, B. Economic and environmental benefits of biodiversity: The annual economic and environmental benefits of biodiversity in the United States total approximately $300 billion. BioScience 1997, 47, 747–757. [Google Scholar] [CrossRef]
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef] [Green Version]
- Lautenbach, S.; Seppelt, R.; Liebscher, J.; Dormann, C.F. Spatial and temporal trends of global pollination benefit. PLoS One 2012, 7, e35954. [Google Scholar]
- Kremen, C.; Ostfeld, R.S. A call to ecologists: Measuring, analyzing, and managing ecosystem services. Front. Ecol. Environ. 2005, 3, 540–548. [Google Scholar] [CrossRef]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trend. Ecol. Evol. 2012, 28, 230–238. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trend. Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and The Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Winfree, R.; Aguilar, R.; Vázquez, D.P.; LeBuhn, G.; Aizen, M.A. A meta-analysis of bees’ responses to anthropogenic disturbance. Ecology 2009, 90, 2068–2076. [Google Scholar] [CrossRef]
- Williams, I.H. The dependence of crop production within the European Union on pollination by honey bees. Agric. Zool. Rev. 1994, 6, 229–257. [Google Scholar]
- Winfree, R.; Williams, N.M.; Dushoff, J.; Kremen, C. Native bees provide insurance against ongoing honey bee losses. Ecol. Lett. 2007, 10, 1105–1113. [Google Scholar] [CrossRef]
- Our Life Insurance, Our Natural Capital: An EU Biodiversity Strategy to 2020; European Commission: Brussels, Belgium, 2011.
- Maes, J.; Egoh, B.; Willemen, L.; Liquete, C.; Vihervaara, P.; Schägner, J.P.; Grizzetti, B.; Drakou, E.G.; Notte, A.L.; Zulian, G.; et al. Mapping ecosystem services for policy support and decision making in the European Union. Ecosyst. Serv. 2012, 1, 31–39. [Google Scholar] [CrossRef]
- Haines-Young, R.H.; Potschin, M.P. The Links between Biodiversity, Ecosystem Services and Human Well-Being. In Ecosystem Ecology: A New Synthesis; Raffaelli, D.G., Frid, C.L.J., Eds.; Cambridge University Press: Cambridge, UK, 2010; p. 162. [Google Scholar]
- Kells, A.R.; Goulson, D. Preferred nesting sites of bumblebee queens (hymenoptera: Apidae) in agroecosystems in the UK. Biol. Conserv. 2003, 109, 165–174. [Google Scholar] [CrossRef]
- Svensson, B.; Lagerlöf, J.; Svensson, B.G. Habitat preferences of nest-seeking bumble bees (hymenoptera: Apidae) in an agricultural landscape. Agric. Ecosyst. Environ. 2000, 77, 247–255. [Google Scholar] [CrossRef]
- Westphal, C.; Steffan-Dewenter, I.; Tscharntke, T. Mass flowering crops enhance pollinator densities at a landscape scale. Ecol. Lett. 2003, 6, 961–965. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Veldtman, R.; Shenkute, A.G.; Tesfay, G.B.; Pirk, C.W.W.; Donaldson, J.S.; Nicolson, S.W. Natural and within-farmland biodiversity enhances crop productivity. Ecol. Lett. 2011, 14, 251–259. [Google Scholar] [CrossRef]
- Aizen, M.A.; Feinsinger, P. Forest fragmentation, pollination, and plant reproduction in a chaco dry forest, Argentina. Ecology 1994, 75, 330–351. [Google Scholar] [CrossRef]
- Aizen, M.A.; Garibaldi, L.A.; Cunningham, S.A.; Klein, A.M. How much does agriculture depend on pollinators? Lessons from long-term trends in crop production. Ann. Bot. 2009, 103, 1579–1588. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Kremen, C.; Morales, J.M.; Bommarco, R.; Cunningham, S.A.; Carvalheiro, L.G.; Chacoff, N.P.; Dudenhöffer, J.H.; Greenleaf, S.S.; et al. Stability of pollination services decreases with isolation from natural areas despite honey bee visits. Ecol. Lett. 2011, 14, 1062–1072. [Google Scholar] [CrossRef]
- Lonsdorf, E.; Kremen, C.; Ricketts, T.; Winfree, R.; Williams, N.; Greenleaf, S. Modelling pollination services across agricultural landscapes. Ann. Bot. 2009, 103, 1589–1600. [Google Scholar] [CrossRef]
- Kareiva, P.; Tallis, H.; Ricketts, T.H.; Daily, G.C.; Polasky, S. Natural Capital—Theory and Practice of Mapping Ecosystem Services; Oxford University Press: New York, NY, USA, 2011. [Google Scholar]
- Britz, W.; Witzke, H.P. Capri Model Documentation 2008: Version 2; Institute for Food and Resource Economics, University of Bonn: Bonn, Germany, 2008. Available online: http://www.capri-model.org/dokuwiki/doku.php/ (accessed on 22 August 2013).
- Corbet, S.A. Temperature and the pollinating activity of social bees. Ecol. Entomol. 1993, 18, 17–30. [Google Scholar] [CrossRef]
- Weissteiner, C.J.; Strobl, P.; Sommer, S. Assessment of status and trends of olive farming intensity in EU-mediterranean countries using remote sensing time series and land cover data. Ecol. Indic. 2011, 11, 601–610. [Google Scholar] [CrossRef]
- Paracchini, M.L.; Petersen, J.E.; Hoogeveen, Y.; Bamps, C.; Burfield, I.; van Swaay, C. High Nature Value Farmland in Europe. An Estimate of the Distribution Patterns on the Basis of Land Cover and Biodiversity Data; Publications Office of the European Union: Luxembourg, 2008. [Google Scholar]
- Clerici, N.; Weissteiner, C.J.; Paracchini, M.L.; Boschetti, L.; Baraldi, A.; Strobl, P. Pan-European distribution modelling of stream riparian zones based on multi-source earth observation data. Ecol. Indic. 2013, 24, 211–223. [Google Scholar] [CrossRef]
- Kempeneers, P.; Sedano, F.; Seebach, L.; Strobl, P.; San-Miguel-Ayanz, J. Data fusion of different spatial resolution remote sensing images applied to forest-type mapping. IEEE Trans. Geosci. Remote Sens. 2011, 49, 4977–4986. [Google Scholar] [CrossRef]
- Baruth, B.; Genovese, G.; Leo, O. Cgms Version 9.2 User Manual and Technical Documentation; Publications Office of the European Union: Luxembourg, 2007. [Google Scholar]
- Ricketts, T.H.; Daily, G.C.; Ehrlich, P.R.; Michener, C.D. Economic value of tropical forest to coffee production. Proc. Natl. Acad. Sci. USA 2004, 101, 12579–12582. [Google Scholar]
- Andersson, G.K.S.; Rundlöf, M.; Smith, H.G. Organic farming improves pollination success in strawberries. PLoS One 2012, 7, e31599. [Google Scholar] [CrossRef]
- Holzschuh, A.; Steffan-Dewenter, I.; Tscharntke, T. Agricultural landscapes with organic crops support higher pollinator diversity. Oikos 2008, 117, 354–361. [Google Scholar] [CrossRef]
- Holzschuh, A.; Dudenhöffer, J.H.; Tscharntke, T. Landscapes with wild bee habitats enhance pollination, fruit set and yield of sweet cherry. Biol. Conserv. 2012, 153, 101–107. [Google Scholar] [CrossRef]
- Winfree, R.; Williams, N.M.; Gaines, H.; Ascher, J.S.; Kremen, C. Wild bee pollinators provide the majority of crop visitation across land-use gradients in New Jersey and Pennsylvania, USA. J. Appl. Ecol. 2008, 45, 793–802. [Google Scholar]
- Hopwood, J.L. The contribution of roadside grassland restorations to native bee conservation. Biol. Conserv. 2008, 141, 2632–2640. [Google Scholar] [CrossRef]
- Henriksen, C.I.; Langer, V. Road verges and winter wheat fields as resources for wild bees in agricultural landscapes. Agric. Ecosyst. Environ. 2013, 173, 66–71. [Google Scholar] [CrossRef]
- Gathmann, A.; Tscharntke, T. Foraging ranges of solitary bees. J. Anim. Ecol. 2002, 71, 757–764. [Google Scholar] [CrossRef]
- Ricketts, T.H.; Regetz, J.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Bogdanski, A.; Gemmill-Herren, B.; Greenleaf, S.S.; Klein, A.M.; Mayfield, M.M.; et al. Landscape effects on crop pollination services: Are there general patterns? Ecol. Lett. 2008, 11, 499–515. [Google Scholar] [CrossRef]
- Zurbuchen, A.; Landert, L.; Klaiber, J.; Müller, A.; Hein, S.; Dorn, S. Maximum foraging ranges in solitary bees: Only few individuals have the capability to cover long foraging distances. Biol. Conserv. 2010, 143, 669–676. [Google Scholar] [CrossRef]
- Westphal, C.; Bommarco, R.; Carré, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyörgyi, H.; Tscheulin, T.; et al. Measuring bee diversity in different european habitats and biogeographical regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef]
- Henry, M.; Béguin, M.; Requier, F.; Rollin, O.; Odoux, J.-F.; Aupinel, P.; Aptel, J.; Tchamitchian, S.; Decourtye, A. A common pesticide decreases foraging success and survival in honey bees. Science 2012, 336, 348–350. [Google Scholar] [CrossRef]
- Whitehorn, P.R.; O’Connor, S.; Wackers, F.L.; Goulson, D. Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 2012, 336, 351–352. [Google Scholar] [CrossRef]
- Maes, J.; Hauck, J.; Paracchini, M.L.; Ratamäki, O.; Hutchins, M.; Termansen, M.; Furman, E.; Pérez-Soba, M.; Braat, L.; Bidoglio, G. Mainstreaming ecosystem services into EU policy. Curr. Opin. Environ. Sustain. 2013, 5, 128–134. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zulian, G.; Maes, J.; Paracchini, M.L. Linking Land Cover Data and Crop Yields for Mapping and Assessment of Pollination Services in Europe. Land 2013, 2, 472-492. https://doi.org/10.3390/land2030472
Zulian G, Maes J, Paracchini ML. Linking Land Cover Data and Crop Yields for Mapping and Assessment of Pollination Services in Europe. Land. 2013; 2(3):472-492. https://doi.org/10.3390/land2030472
Chicago/Turabian StyleZulian, Grazia, Joachim Maes, and Maria Luisa Paracchini. 2013. "Linking Land Cover Data and Crop Yields for Mapping and Assessment of Pollination Services in Europe" Land 2, no. 3: 472-492. https://doi.org/10.3390/land2030472