Phosphorus Solubilizing and Releasing Bacteria Screening from the Rhizosphere in a Natural Wetland

1
Department of Municipal Engineering, School of Civil Engineering, Southeast University, Nanjing 210096, China
2
Department of Environmental Engineering, Nanjing Institute of Technology, Nanjing 211167, China
*
Author to whom correspondence should be addressed.
Water 2018, 10(2), 195; https://doi.org/10.3390/w10020195
Submission received: 24 December 2017
/
Revised: 29 January 2018
/
Accepted: 30 January 2018
/
Published: 12 February 2018
(This article belongs to the Special Issue Sponge Cities: Emerging Approaches, Challenges and Opportunities)
Abstract
:Inorganic phosphorus (P)-solubilizing bacteria (IPSB) and organic P-mineralizing bacteria (OPMB) were isolated from bacteria that were first extracted from the rhizosphere soil of a natural wetland and then grown on either tricalcium phosphate or lecithin medium. The solubilizing of inorganic P was the major contribution to P availability, since the isolated bacteria released much more available P from inorganic tricalcium phosphate than lecithin. IPSB No. 5 had the highest P release rate, that is, 0.53 mg·L−1·h−1 in 96 h, and R10′s release rate was 0.52 mg·L−1·h−1 in 10 days. The bacteria were identified as Pseudomonas sp. and Pseudomonas knackmussii, respectively. R10 released as much as 125.88 mg·L−1 dissolved P from tricalcium phosphate medium, while R4 released the most dissolved P from organic P medium among the isolates, with a concentration of 1.88 mg·L−1 and a releasing rate of 0.0078 mg·L−1·h−1 in ten days. P releasing increased with a pH decrease only when it was from inorganic P, not organic lecithin, and there was no significant correlation between the culture pH and P solubilizing. High-throughput sequencing analysis revealed that the dominant phylum in the studied wetland rhizosphere consisted of Acidobacteria, Proteobacteria, Bacteroidetes and Chloroflexi, accounting for 34.9%, 34.2%, 8.8% and 4.8%, respectively.
1. Introduction
Wetlands are useful for controlling pollution related to phosphorus (P) [1,2,3,4], as it may be taken up by vegetation before being washed into lakes or rivers. Much research is concerned with the effects of P removal by plants in wetlands [1,5,6], especially with the control of non-point source pollution [7]. However, plants can only absorb P in dissolved forms such as the free orthophosphate ions, H2PO4− and HPO42− [8], while the available P in natural soil can be easily converted into insoluble complexes such as iron and aluminum hydrous oxides, crystalline and amorphous aluminum silicate, and calcium carbonate [9]. P can be dissolved by several mechanisms including Fe reduction [10], Ca removal and ligand exchange. Under anaerobic conditions, Fe3+ is microbially reduced to soluble Fe2+, which releases into solution the PO43− bound to Fe2+ oxides [11]. Ca2+ or PO43− removal, such as that absorbed into the cell, may promote the dissociation of tricalcium phosphate. Organic acids can affect phosphate dissolution by competition for adsorption sites, the dissolution of adsorbents, a change in the surface charge of adsorbents, the creation of new adsorption sites through the adsorption of metal ions, and the retardation of the crystal growth of poorly ordered Al and Fe oxides [12].
There are many studies focusing on P-solubilizing and -releasing bacteria in agricultural fields. Inorganic P-solubilizing bacteria (IPSB), such as Bacillus thuringiensis, Enterobacter intermedium, B. megaterium, Burkholderia caryophylli, Pseudomonas cichorii, Pseudomonas syringae. Bacillus sp. and Burkholderia sp. [13,14,15,16], have been screened and identified for solubilizing insoluble forms of P, in order to make them available for the growth of crops [17,18]. As organic P accounts for 30–80% of the total P in most agricultural soils [19], organic P-mineralizing bacteria (OPMB) have been considered as predominant for P availability. OPMBs like Bacillus cereus and Bacillus megaterium that were isolated from agricultural soils have also been reported [14].
In natural wetlands, the rhizosphere bacteria’s ability to transform P, such as through the release of available P, has not been reported. Bacteria with better P-solubilizing and -releasing abilities for extraction from natural wetlands have not been studied so far. It is not known which bacteria may helpful and how they promote the uptake of P by wetland plants, although it would be helpful to remove extra P from the environment in order to reduce the eutrophication problems. Unlike agricultural soil, wetlands have a much higher level of inorganic P and moisture content, with the inorganic P accounting for 60–97% of the total P in wetland areas; total P was also lower than in fertilized fields [20,21,22,23,24]. Due to the different physicochemical properties and vegetation, the microbial community of wetlands is different from that of farmland soil [10,25,26]. Rhizosphere microorganisms are able to convert insoluble forms of P to an accessible soluble form, where different strains show different efficiencies in P solubilization [16]. Reasons for the differences in the duration of P solubilization and release of bacteria strains may include: (1) different growth rate of the bacteria; (2) the production and secretion of different P solubilizing materials such as H+ in the form of different acids, reducing substances to convert Fe3+ to Fe2+- and Ca2+-bound proteins out of the cell to solubilize tricalcium phosphate; (3) adsorption and absorption of Ca2+ or PO43− into the cell to promote the dissociation of tricalcium phosphate; and (4) production and secretion of related hydrolytic enzymes for releasing P from organic P, such as lecithin.
Certain bacteria species like IPSB and OPMB could be isolated, screened and applied in the rhizosphere of wetlands in order to promote the P absorption of plants, to enhance the P removal efficiency of wetlands. However, phosphorus solubilizing and releasing bacteria screening studies of a natural wetland rhizosphere have not been reported to a great extent so far. In this study, the composition of the bacteria community in the rhizosphere from the sampling wetland was examined using a high-throughput sequencing analysis. IPSB and OPMB were isolated using tricalcium phosphate and lecithin medium from the rhizosphere of Acorus calamus L., reed and Iris tectorum Maxim. in the studied wetland. The pH values were also recorded during the incubation, because the rate of phosphate uptake decreases as the pH value of the external solution increases [27]. In other words, P availability decreased with increase in the pH value of aquaponic nutrient solutions [28], and high pH values resulted in the formation of insoluble calcium phosphate species.
The objectives of this work are: (i) extract bacteria strains with better P solubilizing and releasing abilities from the natural wetland rhizosphere; (ii) test their ability to solubilize and release P from different compounds (organic P and inorganic P); (iii) identify the bacteria that solubilize and release the most P; and (iv) test the composition of the bacteria community in the studied wetland rhizosphere to show the proportion of the identified P solubilizing and releasing bacteria and to predict the potential P solubilizing and releasing microbes to be cultured and screened. The novelty of this work is the extraction of P solubilizing and releasing bacteria from the rhizosphere of natural wetland plants. These screened bacteria may more adaptable in the wetland environment when applied back to the field because they were wetland rhizosphere species. The work will also provide strains and a research basis for the future use of bacteria to promote the removal of phosphorus pollutants by wetland plants. This work used a high-throughput sequencing analysis to illustrate the composition of the bacteria community in the rhizosphere of the wetland, to understand the species and their proportion of wetland rhizosphere bacteria to the greatest extent and not be restricted to the bacteria that can be cultured in a lab.
2. Material and Methods
2.1. Material
Acorus calamus L., reed and Iris tectorum Maxim. were chosen because they are common wetland plants in East China. The experimental vegetation and their rhizosphere material were sampled from the wetland area of Tinyin Lake near the egret natural reserve in Nanjing, China, of which the water quality was monitored every year, showing that the average concentration of total phosphorus was lower than the level III baseline in the Chinese National Surface Water Environmental Quality Standard (GB3838-2002, 2002). Rhizosphere samples were taken from a mixture of soil on the roots (in 5 mm) of the plants that were gathered around the wetland area of the lake.
2.2. Methods
2.2.1. Extraction, Isolation and Purification of Rhizosphere Bacteria
The root surface soil and epidermis were first scraped off the plants, and then treated with a shaking technique to extract liquids with rhizosphere bacteria. The shaking technique here was to add 1 g of root surface soil and epidermis into a sterile conical flask and fill sterile water to 100 mL. The liquid was then placed on a constant temperature oscillator (HZ-81B, Shanghai Yuejin medical instruments factory, Shanghai, China) at 30 °C for 30 min. There was a dilution rate of 10−2 extract. The extracts were then prepared under sterile conditions with dilution rates of 10−3, 10−4, 10−5 and 10−6, respectively. The separation and purification of microorganisms in the extracts were carried out using plate culture and plate streak methods in the aseptic environment [29] (Superclean bench, CJ-1FD, Shanghai Boxun medical bioinstrument Co., Ltd., Shanghai, China; Biochemistry cultivation cabinet, GZP250S, Shanghai Xinnuo instrument and equipment Co., Ltd., Shanghai, China). The bacterial collective medium contained: beef paste (3 g), NaCl (5 g), peptone (10 g), agar (15 g), distilled water (1000 mL), with a pH value of 7.0–7.2. Different bacteria colonies grown on the plates were selected based on their morphological characteristics such as size, shape, surface condition, color, edge and cross-section shape.
2.2.2. Preparation of P Medium
(1) Inorganic P (Ca3(PO4)2) liquid medium (IPLM)
The isolation and purification of IPSB strains were carried out using IPLM based on tricalcium phosphate medium (TPM) [30], which contained: glucose (10 g), (NH4)2SO4 (0.5 g), NaCl (0.3 g), KCl (0.3 g), MgSO4·7H2O (0.3 g), FeSO4·7H2O (0.03 g), MnSO4·H2O (0.03 g), Ca3(PO4)2 (0.7 g) (140 mg·L−1 total P) and distilled water (1000 mL) with a pH value of 7.0–7.2.
(2) Organic P (lecithin) liquid medium (OPLM)
A liquid lecithin medium (OPLM) for the OPMB strains was prepared based on the Menkina medium [31], which was composed of: Glucose (10 g), (NH4)2SO4 (0.5 g), NaCl (0.3 g), KCl (0.3 g), MgSO4·7H2O (0.3 g), FeSO4·7H2O (0.03 g), MnSO4·H2O (0.03 g), CaCO3 (5 g), lecithin (2 g) (85.10 mg·L−1 total P), and distilled water 1000 mL, with a pH value of 7.0–7.2.
Inorganic P (Ca3(PO4)2) solid medium (IPSM) and organic P (lecithin) solid medium (OPSM) were made by adding 18 g agar to the liquid medium described above. All media were adjusted to proper pH values (7.0–7.2) with 1 mol·L−1 NaOH or HCl, and sterilized using an autoclave (LDZX-40KBS, Shanghai shenan medical equipment factory, Shanghai, China) for 20 min at 121 °C.
2.2.3. Measurement of P-Solubilizing and -Releasing Capability
(1) Measurement of P-solubilizing capability of bacteria in solid medium
The preliminarily-screened P-solubilizing bacteria strains were first cultured on IPSM and OPSM respectively, and then incubated at 30 °C for 96 h, then the P solubilizing ring was observed and its size was measured by 2b/a, where a is the total diameter of the ring and colony, and 2b is the difference between the outer and inner rings. The P-solubilizing ring is a clarified area around the bacterial colony on IPSM/OPSM, which indicates that the bacteria may have stronger P-solubilizing ability than those without the ring. By observing whether there is a P solubilizing ring and its size, it is more convenient to screen out the bacteria with a stronger P solubilizing ability from a large number of bacteria.
(2) Measurement of P-solubilizing and -releasing capability of bacteria in liquid medium and pH
To determine the short-term P-solubilizing and -releasing rate of the bacteria, each strain was first cultured in 100 mL liquid medium, and then incubated in a constant-temperature oscillator at 30 °C for 96 h (130 r·min−1), when the pH and soluble P was examined from the supernatant of centrifugal medium ((7000 r·min−1) for 20 min) every 48 h. To determine the long-term P solubilizing and releasing rate, the bacteria strain was first cultured in the liquid medium and then incubated at 30 °C for 10 days, when the concentration of soluble P from the supernatant of centrifugal medium (Centrifuge, TGL-16G-XTA10, Shanghai Anting scientific instrument factory , Shanghai, China) was tested every 24 h.
(3) Analysis of the samples
Total P and soluble phosphates were determined using the ammonium molybdate spectrophotometric method (Spectrophotometer, 752N, Shanghai Youke instrument Co., Ltd., Shanghai, China) (Determination of total phosphorus of water quality ammonium molybdate spectrophotometric method Chinese GB 11893-1989) and represented as P concentration (mg·L−1). Samples of total P were pretreated by potassium persulfate digestion. Three replicates of the control treatment were included in the experiment for each isolate and the results were expressed as mean values.
2.2.4. P-Solubilizing and -Releasing Rate Calculation
The bacterial P-solubilizing and -releasing rate η (mg·L−1·h−1) was calculated using the following formula:
where C0 is the initial soluble P concentration in the liquid medium, Cn is the soluble P concentration in the liquid medium after n hours, and t is the incubation time.
2.2.5. Microbial Community Analysis
A total of 15 g of rhizosphere soil sample was sent to Shanghai Majorbio Bio-pharm Technology Co., Ltd. (Shanghai, China) and analyzed using a high-throughput sequencing analysis. The sequencing type, primer name and sequencing platform were bacterial 16S rRNA, 338F_806R and PE300, respectively.
3. Results and Discussion
3.1. P-Solubilizing Capacity of Inorganic Insoluble P (Ca3(PO4)2)
(1) P-solubilizing and -releasing capacity of insoluble Ca3(PO4)2 in the solid medium
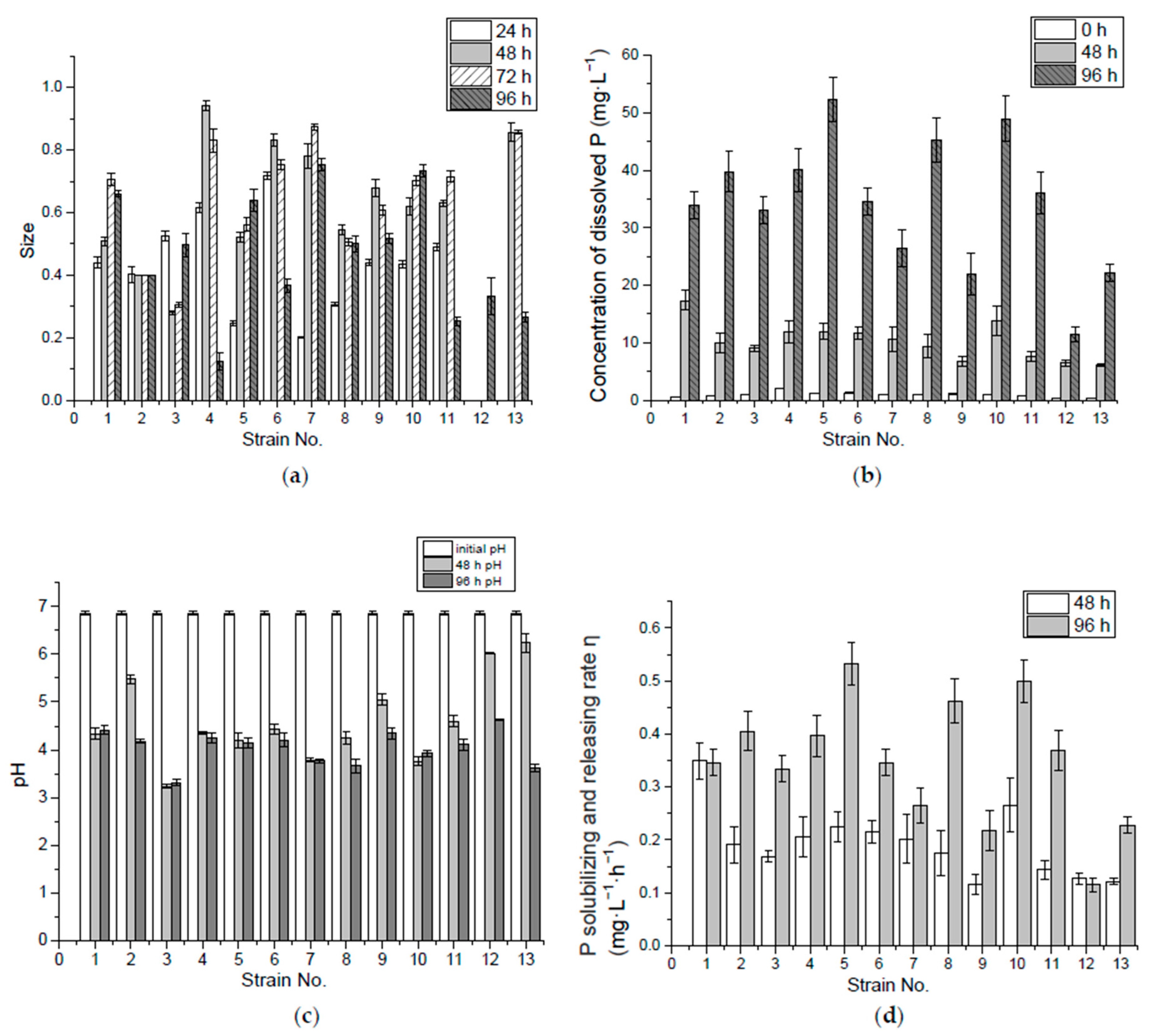
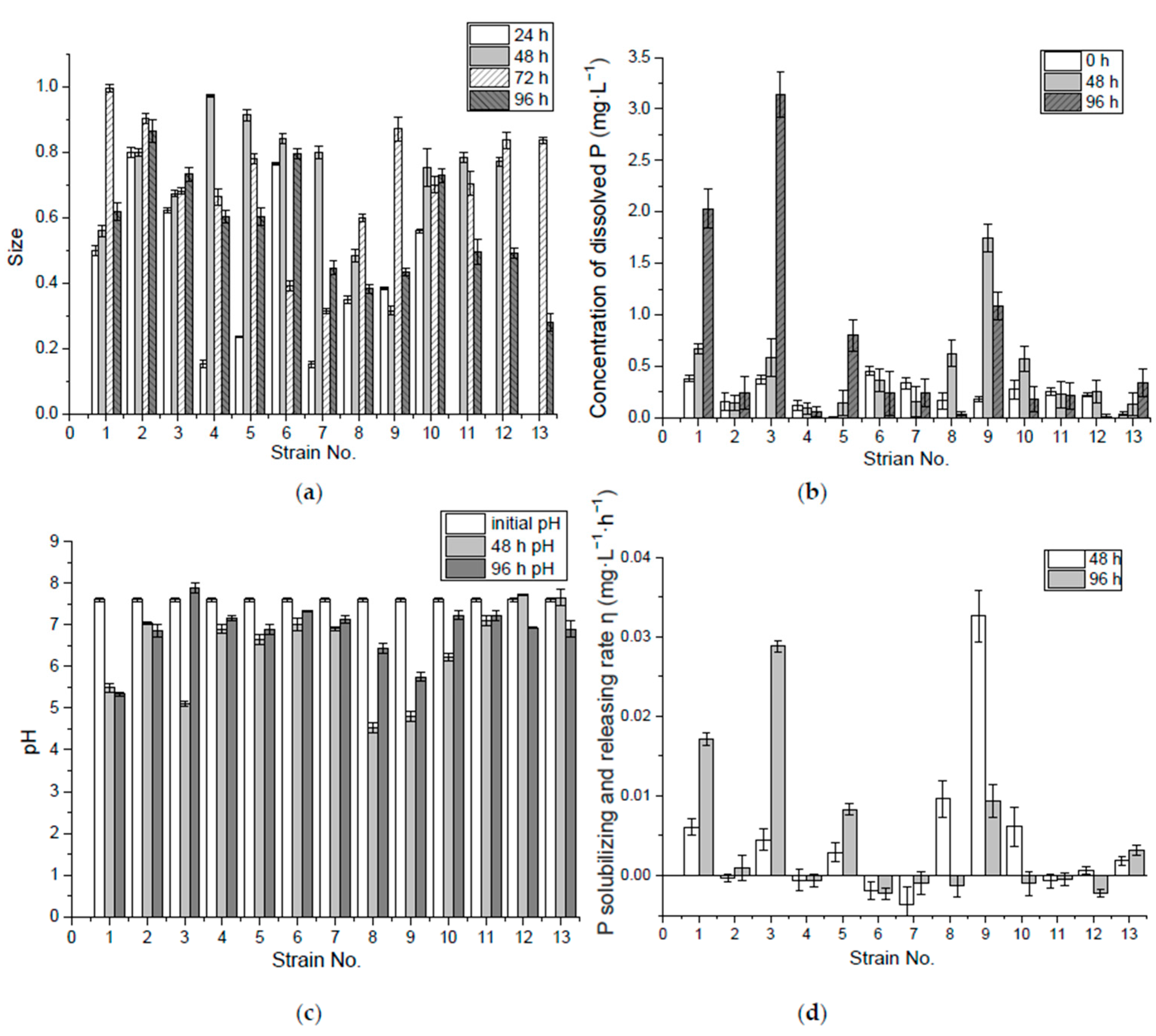
It could be seen in the experiment with isolated bacteria strains growing on IPSM that only some strains showed the visible P solubilizing ring. Among the 13 strains selected and cultured on IPSM for 96 h, colonies of strain No. 1 to No. 11 formed the P solubilizing ring in the first 24 h, while No. 12 and 13 formed the P solubilizing ring later, after around 96 h and 72 h, respectively, illustrating the differences in the duration of the P-solubilizing and -releasing of bacteria stains. Among these isolates, 15.4%, 30.1%, 30.1% and 23.1% reached the maximum size in 24 h, 48 h, 72 h and 96 h, respectively. In terms of the average ring size grown at 96 h, strains No. 4, 6, 7 and 10 were better in insoluble P-solubilizing and -releasing in the solid medium (Figure 1a).
The insoluble tricalcium phosphate (Ca3(PO4)2) in the solid medium appeared turbid when the ring area appeared clarified, which means the tricalcium phosphate was dissolved/consumed. The biochemical interactions may include: (1) the tricalcium phosphate dissolved in the low pH environment caused by acid production from the bacteria; (2) the adsorption or absorption of Ca2+ by bacteria thus releasing PO43−; and (3) the absorption of PO43− by the bacteria. If the ring was caused by the third reason, and the PO43− was absorbed by bacteria, and then the bacteria cannot be judged as P-releasing bacteria. Thus, these bacteria have to be incubated in liquid medium and the soluble P level in the solution must be tested. In IPLM, if the insoluble tricalcium phosphate dissolved during the incubation, the higher soluble P level would indicate that the bacteria have a better P-solubilizing and -releasing ability. Otherwise the lower soluble P level when the insoluble tricalcium phosphate dissolved would mean that the bacteria could solubilize P not release much of the available P, which may be taken up by the bacteria. In this case, this type of bacteria cannot be used to promote P uptake by wetland plants since P is not released.
(2) P-solubilizing and -releasing capacity of insoluble Ca3(PO4)2 in liquid medium
In the first 48 h of incubation, No. 1 and 10 released more dissolved P. By 96 h, however, No. 5, 8 and 10 released more in the liquid medium, as shown in Figure 1b. The decrease in pH value helped P release, since orthophosphate is mostly available at pH 3.0–5.5 in the aquaponic nutrient solution [28]. In this experiment, most of the isolates which brought the medium pH value down below 5 in the first 48 h might have a better capability of P solubilization and release. Thus, strains, like No. 12, 13 and 9, that were unable to lower the pH below 5 at the beginning showed poor performance in P release. However, the decrease in pH value was not the only cause of P release, although No. 3 and 7 had lowered the pH value below 4 by the end of the first 48 h, the concentration of the dissolved P in the medium was relatively low, compared with the dissolved P in the media of the other isolates. Take glucose in the medium as substrate, for example, under aerobic conditions the sugar is first metabolized to pyruvic acid through glycolysis, and the intermediate products are fructose 1,6-bisphosphate, glyceraldehyde 3-phosphate, and 2-phosphoglycerate, sequentially, during which two phosphates have been consumed to generate a net yield of two adenosine triphosphate (ATP) molecules. Then the pyruvate from the glycolysis is converted to acetyl-Coenzyme A (acetyl-CoA) by the action of pyruvate dehydrogenase complex, and acetyl-CoA is completely oxidized to CO2 and water through citric acid cycle. O2 consumption and pyruvate oxidation were found to be stimulated by some four-carbon dicarboxylic acids (fumarate, succinate, malate and oxaloacetate), five-carbon dicarboxylic acid (a-ketoglutarate), or six-carbon tricarboxylic acids (citrate, isocitrate, cis-aconitate) [29,32]. These citric acid intermediates are important sources of biosynthetic precursors. In aerobic respiration one molecular of glucose may yield 36–38 ATP and P-containing bio-molecules like nucleic acid, phospholipid, polyphosphate and P-containing enzymes are synthesized [29]. Due to this large consumption/biosynthesis of P, the dissolved P in the liquid medium would be low when a large number of bacteria cells or P-containing bio-molecules are produced under aerobic conditions. Under anaerobic conditions, some facultative or anaerobic bacteria species undergo fermentation where glucose is metabolized to pyruvate. As a result of the need to produce high-energy, phosphate-containing, organic compounds (generally in the form of CoA-esters) fermentative bacteria use Nicotinamide adenine dinucleotide (NADH) and other cofactors to produce many different reduced metabolic by-products, often including H2 and small organic acids and alcohols derived from pyruvate, such as ethanol, acetate, lactate, and butyrate [32].
Many studies have shown that P solubilizing microorganisms can secrete a variety of low-molecular organic acids during metabolism, such as malic acid, propionic acid, lactic acid, acetic acid and citric acid [33]. These organic acid anions can react with calcium ions in the liquid medium to release P from modestly soluble phosphates [34]. Nos. 5, 8 and 10 had higher solubilizing and releasing rates in incubation during the 96 h, which were 0.53 mg·L−1·h−1, 0.46 mg·L−1·h−1 and 0.49 mg·L−1·h−1, respectively. The pH values of super P-releasing isolates (Nos. 5, 8 and 10) in the liquid medium with Ca3(PO4)2 eventually reached 4.5–3.8, as shown in the results in Figure 1c, similar to the final pH value of 3.99 found in a culture of Pseudomonas K3, which was the best in P solubilizing and screened from the rhizosphere of the calcareous soil [35].
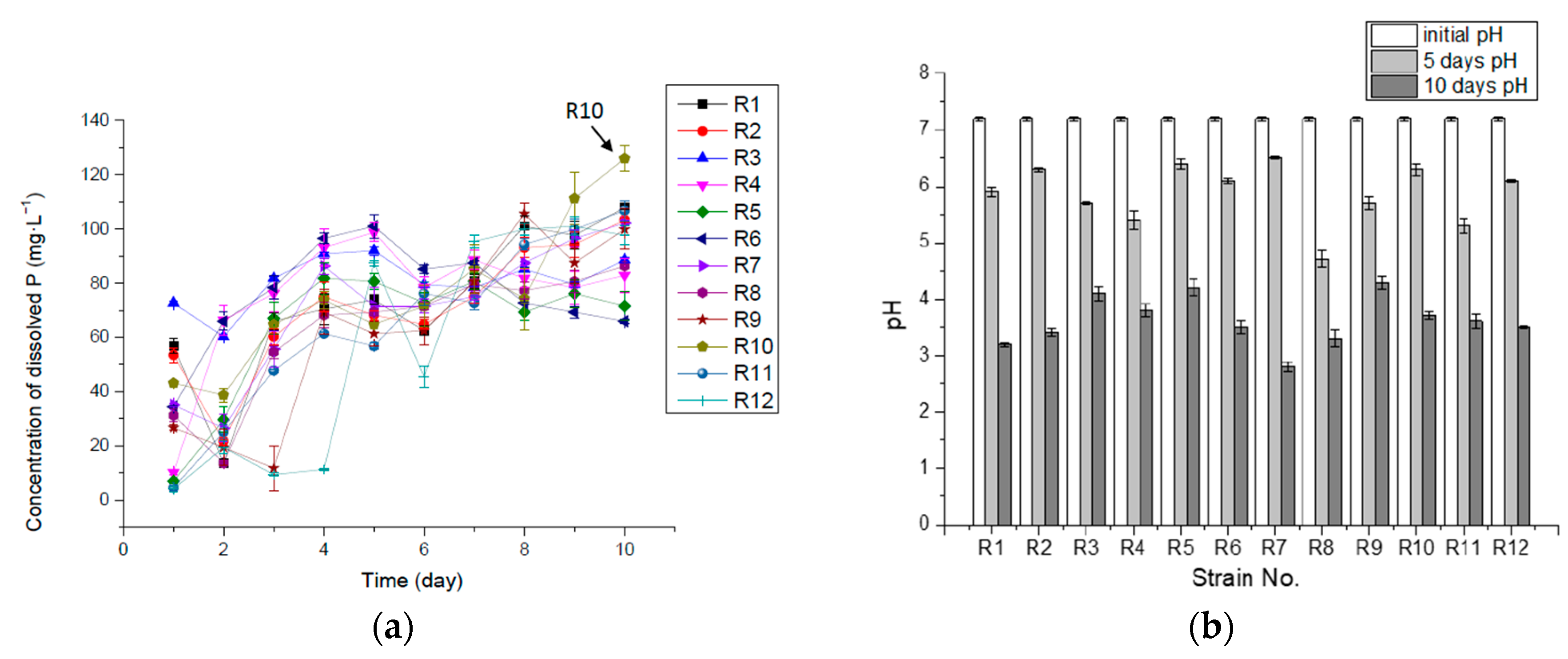
(3) P-solubilizing and -releasing capacity of isolates from reed in liquid medium with insoluble Ca3(PO4)2
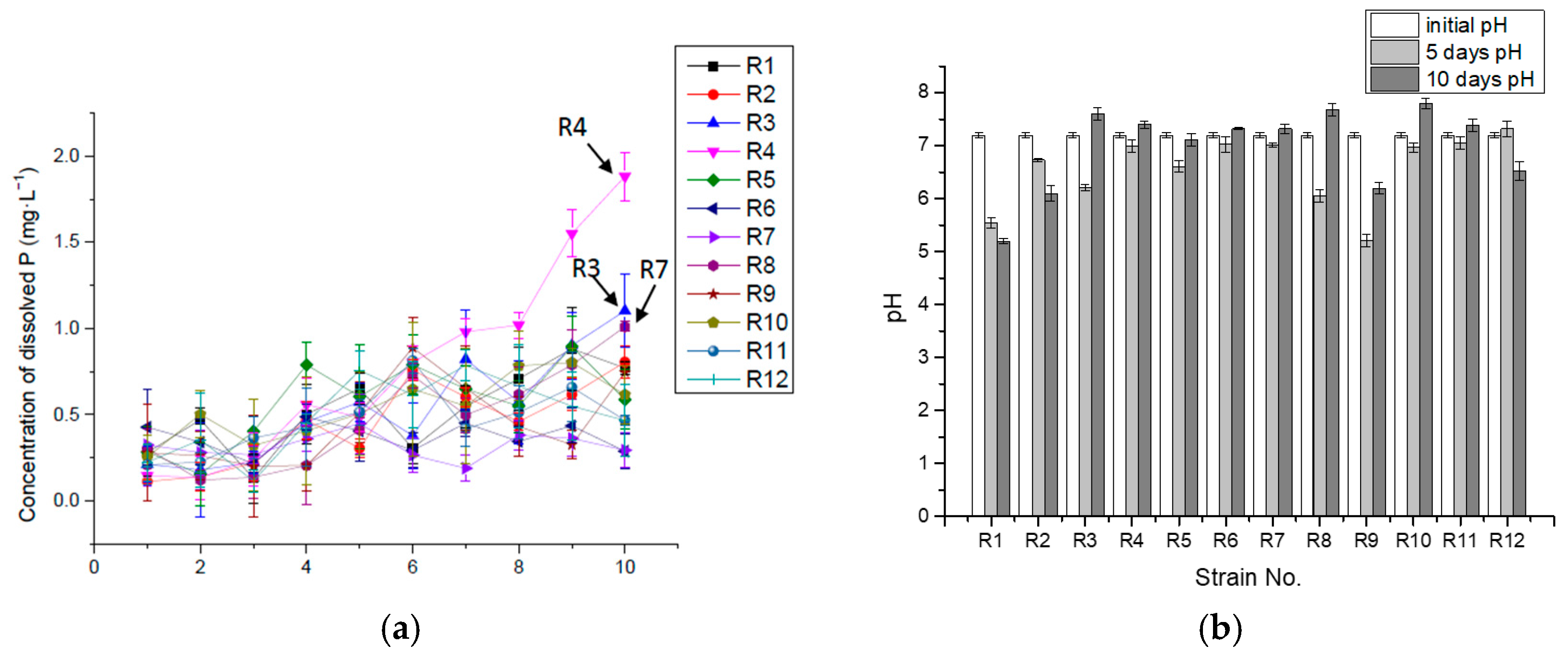
Isolate R10 was the best at releasing available P from insoluble Ca3(PO4)2 in the liquid medium, releasing 125.88 mg·L−1 of solubilized P, accounting for about 90.0% of the total P ten days into the incubation. Other isolates released 47.1–77.0% of the total P in the culture medium. The rest of the total P might be either absorbed by the bacteria and synthesized to organic matter, such as phospholipid, polyphosphate and nucleic acid, or deposited as insoluble tricalcium phosphate, iron (III) phosphate or magnesium phosphate. Some isolates (R1, R2, R3, R7, R8, R9 and R10) released much of the dissolved P at the beginning, possibly due to the fast production of organic acid and carbonic acid from glucose when seeded into the medium. The organic acid might either release hydrogen ions to dissolve Ca3(PO4)2 or combine with calcium ions to release P [36]. Then the concentration of dissolved P kept decreasing for one or two days, which could have been caused by the absorption and assimilation of P in the growth-lag phase of bacterial cells, followed by a period of fast growth, when more acid was produced, leading to the continuously increasing levels of dissolved P (Figure 2).
Figure 2b shows that, although the pH kept decreasing during the incubation, the dissolved P in some cultures started to decline from the middle of the process, implying that it could not have been caused by pH or acid production, but might have resulted from the absorption of P into bacterial cells and the formation of organic P through biosynthesis.
Table 1 shows the maximum concentration of dissolved P and their corresponding time during the incubation process, where R1, R2, R7, R8, R10 and R11 reached their maximum concentration on the tenth day, suggesting that these isolates could have solubilized and released available P for a longer term and at a higher concentration if more total P had existed in the medium. A total of 66.7% of the isolates (R1, R2, R3, R6, R7, R8, R9 and R10) reached the highest rate of dissolution and release P in the first 24 h, with R8 in 48 h, R5 and R11 in 72 h and R12 in 120 h, respectively. R10 had the highest P-releasing rate in the 10 days, with 0.52 mg·L−1·h−1. The results are summarized in Table 1.
(4) Identification of bacteria species
No. 5 and R10 were selected and sent to Shanghai Majorbio Bio-pharm Technology Co., Ltd. to be sequenced and contrasted, and were then identified as Pseudomonas sp. and Pseudomonas knackmussii, both belonging to the Proteobacteria phylum.
3.2. P-Solubilizing Capacity of Organic P (Lecithin)
(1) P-solubilizing capacity of organic P (lecithin) in solid medium
It was observed that the P-solubilizing rings formed in organic solid medium with lecithin were larger in size and clearer than those in inorganic solid medium. The largest size was formed between 48 and 72 h. No. 2, 6 and 10 were the best in forming a P-solubilizing ring in organic solid medium with lecithin, based on the average size from Figure 3a. However, the solubilized ring around the bacteria colony could only indicate that the “cloudy” lecithin was hydrolyzed. Whether the phosphate in the lecithin molecules was released as soluble P requires further confirmation. The hydrolyzed products can be fatty acids and other lipophilic substances such as arachidonic acid, diacylglycerol, phosphatidic acid, alcohol, inositol 1,4,5-trisphosphate, diacyl glycerol, etc. [37,38,39] Even when soluble P was released, it could be further absorbed by bacteria. Thus, the soluble P levels in the liquid medium during the bacterial incubation should be tested to analyze the bacteria’s P-solubilizing and -releasing ability.
(2) P-solubilizing and -releasing capacity of organic P (lecithin) in liquid medium
The concentration of dissolved P and release rate from lecithin were much lower than those from Ca3(PO4)2, where No. 1, 3 and 9 were better at releasing dissolved phosphates. Decreasing the pH or producing acid did not promote the release of dissolved P. The pH value did not decline as much as under incubation in the liquid medium with Ca3(PO4)2, and it even increased, which may have resulted from the neutralization of acid by alkaline substances such as choline and ammonia produced by the bacteria. Since lecithin contains not only PO43−, but also N in the molecule, during the bacterial metabolism N may be released in the form of choline or ammonia [40]. Many bacteria could produce phospholipase to hydrolyze phospholipids into fatty acids and other lipophilic substances. There are four major classes, termed A, B, C and D, distinguished by the type of reaction that they catalyze. Among these, phospholipase C cleaves before the phosphate, releasing diacylglycerol and a phosphate-containing head group. Phospholipase D cleaves after the phosphate, releasing phosphatidic acid and an alcohol. Bacteria such as Bacillus cereus and Pseudomonas aeruginosa [41] could produce phospholipase C, and bacteria such as Stenotrophomonas maltophilia [42] and Streptomyces prunicolor [43] could produce phospholipase D. Figure 3b shows that there was a limitation for most of the isolates to release P since the production volume of dissolved P was small, possibly due to the limitation in production of phospholipase C and D, which can hydrolyze lecithin and release phosphatidic acid, a free head group, such as choline, and further release phosphates [37,44,45] (Figure 3).
(3) P-solubilizing and -releasing capacity of isolates from reed in liquid medium with organic P (lecithin)
The 10-day incubation pH varied from 5.2 to 7.8, illustrating that even when the P was released, it would combine with Ca2+ to form insoluble phosphate, which might thus restrict the concentration of dissolved P in the medium. For R3, R4 and R8 described above, the pH value increased to over 7.5 later in the incubation. These increased alkalinity and dissolved organic compounds (DOC) prevented Ca2+ from binding with PO43− ions. DOC, in the form of organic matter, has charges related to the pH-dependent characteristics of organic acid functional groups [46]. Under alkaline conditions, the organic matter and bacteria surface, which is made of peptidoglycan, eichoic acid, and capsule polysaccharide, have negatively charged cation exchange sites. Therefore, ions with positive charges, such as Ca2+, were attracted to the organic matter surface due to electrostatic interactions [28], thus helping phosphate dissolution in the medium with a higher pH. The partially alkaline environment was more conducive to releasing P in the organic phosphorus culture (Figure 4).
R2, R3, R4 and R8 kept increasing the dissolved P in the medium during the incubation, as shown in Figure 4. R3, R4 and R8 obtained relatively higher maximum concentrations of P release combined with the corresponding time (10th day) (Table 2), implying that these isolates were better at releasing available P for a longer term and higher concentration in the liquid medium with lecithin. The mineralization of organic P does not increase with the decrease of pH.
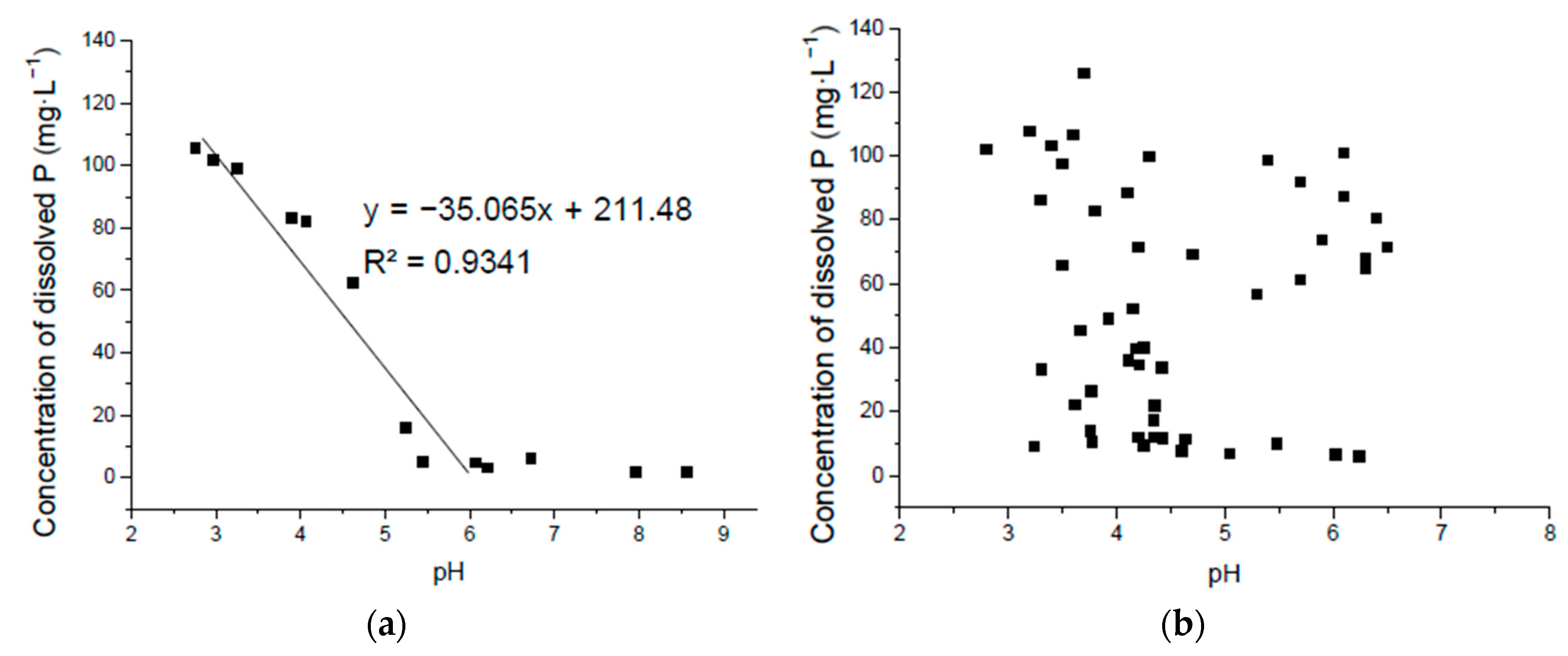
3.3. Correlation of Solubilized P and pH in Ca3(PO4)2 Liquid Medium
The pure Ca3(PO4)2 liquid medium without seeding had a significantly negative linear correlation of P solubilization and pH from about 6.0 and lower. However when pH was higher than this, the liner relationship did not exist. When cultured with the isolated strains, there was no significant correlation between the culture pH and the P-solubilizing amount of the different isolates (Figure 5). Although compared to the P released from the lecithin medium, the pH decreased much more and solubilized more P in Ca3(PO4)2 liquid medium, as discussed above. Thus, other factors such as P absorption, secreted organic acid and related enzymes [14], the reaction with organic molecules in the environment and growing patterns of isolated bacteria species are also important in P solubilization in Ca3(PO4)2 liquid medium.
Since there was only small decrease of pH and no significant correlation between the culture pH and the P mineralized by the OPMB strains grown in lecithin medium [15], the correlation of phosphorus solubilization and pH in lecithin medium was not discussed.
3.4. Microbial Community Analysis
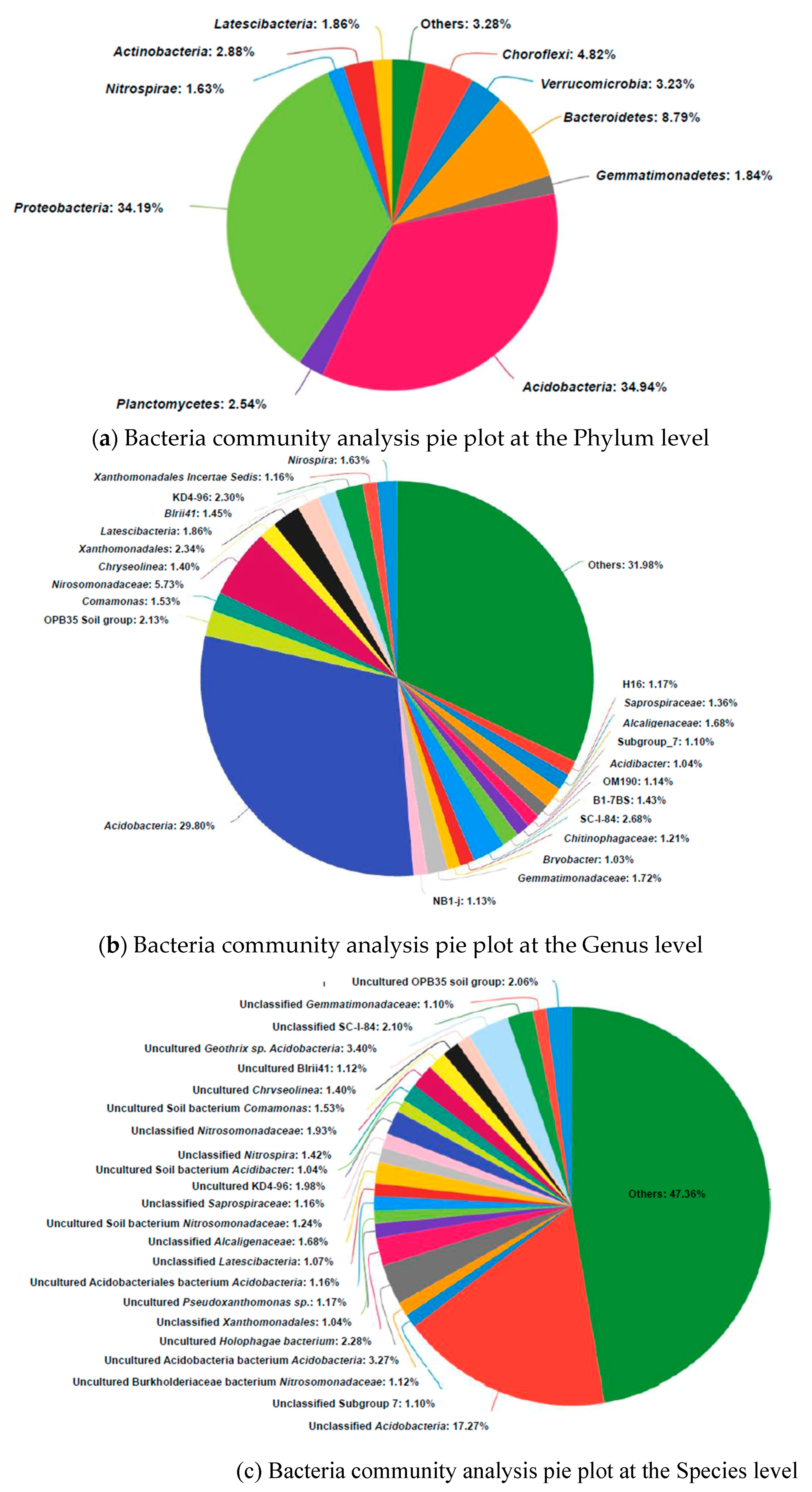
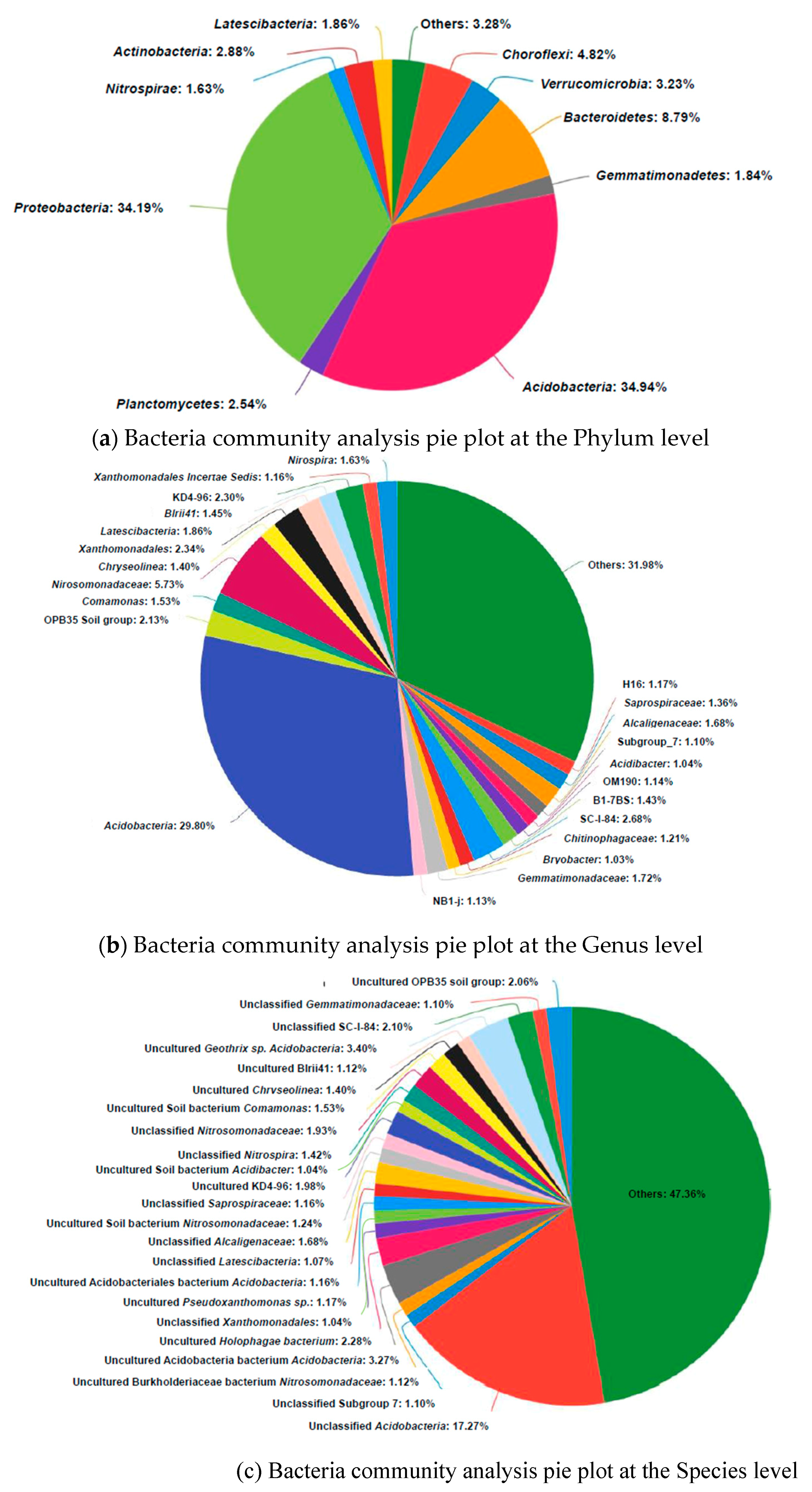
MiSeq analysis of the samples on bacteria 16S rRNA from rhizospheres of the experimental plant yielded 30212 sequences and 445 OTUs. Relatively abundant among the most dominant bacteria genera living in the sampled wetland rhizospheres was Acidobacteria (Figure 6). Some were acidophilic, and since they were only recently discovered and the large majority have not been cultured, the ecology and metabolism of these bacteria is not well understood [49]. Their acidophilic ability may account for tolerance at low pH levels and help dissolve mineral P such as Ca3(PO4)2. No. 5 and R10, which were identified as Pseudomonas sp. and Pseudomonas knackmussii, both belonging to the pseudomonas genus, accounted for less than 0.1%.
The natural wetland rhizosphere bacteria community was compared with other wetland environments in Table 3. The dominant phylum and its proportion were unique. Acidobacteria, Proteobacteria, Bacteroidetes and Chloroflexi were the most abundant in the samples, accounting for 34.9%, 34.2%, 8.8% and 4.8%, respectively. This is different to the Yellow River Delta Wetland and floating treatment wetland where Proteobacteria and Acidobacteria account for only 6.36% and less than 0.3%, respectively. Thus, in the studied natural wetland, rhizosphere Acidobacteria may play an important role in nutrient cycling. Because many of these species are uncultured, new methods other than incubation are required to illustrate their function in P solubilization and release.
4. Practical Applications and Future Perspectives
Through this study, the effects of various bacteria on the formation of dissolved P in the rhizosphere of wetland plants and the basis of the P cycle in the root–microbial–soil–water system of wetland ecosystems could be further elucidated. The bacteria with better P solubilization and release could be preserved for future applications, for instance, seeding on the rhizosphere of wetland plants or directly in the wetland field to promote plant removal of P in the environment, especially for the removal of plant-unavailable P. This thus provides a new idea and approach to enhance the P pollution control efficiency of wetlands.
The bacteria’s P-solubilizing and -releasing effects on other soil forms of P (Fe-P, Al-P, Phytate phosphorus, etc.) could be tested in the future research. The effects of P solubilization and release and plant absorption should be further analyzed when the screened bacteria are applied back in the rhizosphere or in the wetland field. Since the community analysis of the rhizosphere indicated that much of the bacteria species were not cultured, more types of culture technique and screening methods are required to maximize the number of species in the study of P-solubilizing and -releasing bacteria in the rhizosphere of wetlands. Varieties of screening condition should be used, for example, some of the bacteria might have better P-solubilizing and -releasing abilities under anaerobic or facultative conditions. A combination of bacteria species or microbes other than single bacteria strains, fungi or archaea also could be screened and analyzed from the rhizosphere of wetlands and applied to P removal and pollution control in the following studies.
5. Conclusions
The natural wetland rhizospheres in this experiment were dominated by Acidobacteria. The isolates’ solubilizing and releasing rate of inorganic phosphorus was much higher than that of organic P, demonstrating their different impacts on organic and inorganic P. No. 5, 8 and 10 isolated from Acorus calamus L. and Iris tectorum Maxim. and R10 from reed released more available P in the Ca3(PO4)2 medium, while No. 1, 3 and 9, as well as R3, R4 and R8 were better at releasing available P from lecithin. A decrease in pH would increase P solubilization from tricalcium phosphate, but there was no significant correlation between the culture pH and the P solubilization. The mineralization of organic P did not increase with the decrease in pH.
Acknowledgments
Funding: This work was supported by the National Natural Science Foundation of China [31600091], Nanjing Institute of Technology Innovation Fund Project [CKJB201409], Key State Laboratory of Lakes and Environment Open Fund [2016SKL009] and Teaching Reformation and Construction Fund of Nanjing Institute of Technology [JS201719]. Thanks to Professor Dafang Fu for leading and motivating us to finish the research. We thank Rajendra Prasad Singh for helping the submission and revision of the paper. Thanks also to Feng Yang, Changfei Zhang, Fenghua Gao, Mengyun Zhou, Huan Sun, Qi Mei, Anqi Xu, Caiju Qiu, Derong Yang and Tiansheng Wen who dedicate their time and passion in the difficult field sampling and lab investigation. Thanks to Keqiang Ding and Hongyi Li who encourage us to finish the work.
Author Contribution
Ying Cao and Dafang Fu conceived and designed the experiments; Ying Cao, Tingfeng Liu and Guang Guo performed the experiments; Ying Cao, Dafang Fu and Tingfeng Liu analyzed the data; Ying Cao, Dafang Fu, Tingfeng Liu, Guang Guo, Zhixin Hu contributed reagents/materials/analysis tools; Ying Cao and Dafang Fu wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zou, Y.; Zhang, L.; Wang, L.; Zhang, S.; Yu, X. Effects of Aeration, Vegetation, and Iron Input on Total P Removal in a Lacustrine Wetland Receiving Agricultural Drainage. Water 2018, 10, 61. [Google Scholar] [CrossRef]
- Jin, M.; Carlos, J.; McConnell, R.; Hall, G.; Champagne, P. Peat as Substrate for Small-Scale Constructed Wetlands Polishing Secondary Effluents from Municipal Wastewater Treatment Plant. Water 2017, 9, 928. [Google Scholar] [CrossRef]
- Li, W.; Cui, L.; Zhang, Y.; Cai, Z.; Zhang, M.; Xu, W.; Zhao, X.; Lei, Y.; Pan, X.; Li, J.; et al. Using a Backpropagation Artificial Neural Network to Predict Nutrient Removal in Tidal Flow Constructed Wetlands. Water 2018, 10, 83. [Google Scholar] [CrossRef]
- Zhai, J.; Xiao, J.; Rahaman, M.H.; John, Y.; Xiao, J. Seasonal Variation of Nutrient Removal in a Full-Scale Artificial Aerated Hybrid Constructed Wetland. Water 2016, 8, 551. [Google Scholar] [CrossRef]
- Jia, X.; Otte, M.L.; Liu, Y.; Qin, L.; Tian, X.; Lu, X.; Jiang, M.; Zou, Y. Performance of Iron Plaque of Wetland Plants for Regulating Iron, Manganese, and Phosphorus from Agricultural Drainage Water. Water 2018, 10, 42. [Google Scholar] [CrossRef]
- Ping, Y.; Pan, X.; Cui, L.; Li, W.; Lei, Y.; Zhou, J.; Wei, J. Effects of Plant Growth Form and Water Substrates on the Decomposition of Submerged Litter: Evidence of Constructed Wetland Plants in a Greenhouse Experiment. Water 2017, 9, 827. [Google Scholar] [CrossRef]
- Cai, S.; Shi, H.; Pan, X.; Liu, F.; Cui, Y.; Xie, H. Integrating Ecological Restoration of Agricultural Non-Point Source Pollution in Poyang Lake Basin in China. Water 2017, 9, 745. [Google Scholar] [CrossRef]
- Penn, C.; Chagas, I.; Klimeski, A.; Lyngsie, G. A Review of Phosphorus Removal Structures: How to Assess and Compare Their Performance. Water 2017, 9, 583. [Google Scholar] [CrossRef]
- Altomare, C.; Norvell, W.A.; Bjorkman, T.; Harman, G.E. Solubilization of phosphates and micronutrients by the plant growth promoting and biocontrol fungus Trichoderma harzianum Fifai1295-22. Appl. Environ. Microbiol. 1999, 65, 2926–2933. [Google Scholar] [PubMed]
- Reddy, K.R.; Delaune, R.D. Biogeochemistry of wetlands: Science and applications. Soil Sci. Soc. Am. J. 2008, 73, 1779. [Google Scholar]
- Shenker, M.; Seitelbach, S.; Brand, S.; Haim, A.; Litaor, M.I. Redox reactions and phosphorus release in re-flooded soils of an altered wetland. Eur. J. Soil Sci. 2005, 56, 515–525. [Google Scholar] [CrossRef]
- Borggaard, O.K.; Raben-Lange, B.; Gimsing, A.L.; Strobel, B.W. Influence of humic substances on phosphate adsorption by aluminum and iron oxides. Geoderma 2005, 127, 270–279. [Google Scholar] [CrossRef]
- Delfim, J.; Schoebitz, M.; Paulino, L.; Hirzel, J.; Zagal, E. Phosphorus Availability in Wheat, in Volcanic Soils Inoculated with Phosphate-Solubilizing Bacillus thuringiensis. Sustainability 2018, 10, 144. [Google Scholar] [CrossRef]
- Kim, C.H.; Han, S.H.; Kim, K.Y.; Cho, B.H.; Kim, Y.H.; Koo, B.S.; Kim, Y.C. Cloning and expression of pyrroloquinoline quinone (PQQ) genes from a phosphate-solubilizing bacterium Enterobacter intermedium. Curr. Microbiol. 2004, 47, 6. [Google Scholar] [CrossRef]
- Tao, G.C.; Tian, S.J.; Cai, M.Y.; Guang, H.X. Phosphate-Solubilizing and -Mineralizing Abilities of Bacteria Isolated from Soils. Pedosphere 2008, 4, 515–523. [Google Scholar] [CrossRef]
- Oliveira, C.A.; Alves, V.M.C.; Marriel, I.E.; Gomes, E.A.; Scotti, M.R.; Carneiro, N.P.; Guimarães, C.T.; Schaffert, R.E.; Sá, N.M.H. Phosphate solubilizing microorganisms isolated from rhizosphere of maize cultivated in an oxisol of the Brazilian Cerrado Biome. Soil. Biol. Biochem. 2009, 41, 1782–1787. [Google Scholar] [CrossRef]
- Goldstein, A. Recent progress in understanding the moleculargenetics and biochemistry of calcium phosphate solubilization by gram negative bacteria. Biol. Agric. Hortic. 1995, 12, 185–193. [Google Scholar] [CrossRef]
- Illmer, P.; Schinner, F. Solubilization of inorganic phosphateby microorganisms isolated from forest soils. Soil. Biol. Biochem. 1992, 24, 389–395. [Google Scholar] [CrossRef]
- Li, S.M.; Li, L.; Zhang, F.S.; Tang, C. Acid Phosphatase Role in Chickpea/Maize Intercropping. Ann. Bot. 2004, 94, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Zhai, J.H.; Zeng, C.S.; Tong, C.; Wang, W.Q.; Liao, J. Organic and Inorganic Phosphorus in Sediments of the Min River Estuarine Wetlands: Contents and Profile Distribution. J. Subtropic. Res. Environ. 2010, 5, 9–14. [Google Scholar] [CrossRef]
- Cao, X.Y.; Chong, Y.X.; Yu, G.W.; Zhong, H.T. Difference of P Content in Different Area Substrate of Constructed Wetland. Environ. Sci. 2012, 33, 4033–4039. [Google Scholar] [CrossRef]
- Lu, J.; Gao, B.; Hao, H. Study on the occurrence of phosphorus in the sediments of artificial wetland. Spectrosc. Spectr. Anal. 2014, 11, 3162–3165. [Google Scholar] [CrossRef]
- Liang, W.; Shao, X.X.; Wu, M.; Li, W.H.; Ye, X.Q.; Jiang, K.Y. Phosphorus fraction in the sediments from different vegetation type in hangzhou bay coastal wetlands. Acta Ecol. Sin. 2012, 32, 5025–5033. [Google Scholar] [CrossRef]
- Kong, L.Z. Study on the Morphological Characteristics and Adsorption Desorption Characteristics of Phosphorus in Wetland Soil under Different Tillage Year. Ph.D. Thesis, Anhwei Normal University, Wuhu, China, 2014. [Google Scholar]
- Lynch, J.M.; Whipps, J.M. Substrate flow in the rhizosphere. Plant Soil 1990, 129, 1–10. [Google Scholar] [CrossRef]
- Richardson, A.E.; Simpson, R.J. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiol. 2011, 156, 989. [Google Scholar] [CrossRef] [PubMed]
- White, P.J. Ion Uptake Mechanisms of Individual Cells and Roots: Shortdistance Transport. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 7–47. [Google Scholar]
- Cerozi, B.S.; Fitzsimmons, K. The effect of pH on phosphorus availability and speciation in an aquaponics nutrient solution. Bioresour. Technol. 2016, 219, 778–781. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.Y.; Gao, T.Y. Environmental Engineering Microbiology, 3rd ed.; Higher Education Press: Beijing, China, 2010; pp. 380–448. ISBN 978-7-04-022265-4. (In Chinese) [Google Scholar]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Niewolak, S. Occurrence of microorganisms in fertilized lakes. II. Lecithin-mineralizing microorganisms. Pol. Arch. Hydrobiol. 1980, 27, 53–71. [Google Scholar]
- Pepper, L.L.; Gerba, C.P.; Terry, J. Environmental Microbiology, 3rd ed.; Academic Press: Cambridge, MA, USA, 2014; ISBN 978-0-12-394626-3. [Google Scholar]
- Lin, Q.M.; Wang, H.; Zhao, X.R.; Zhao, Z.J. The solubilizing ability of some bacteria and fungi and its mechanisms. Microbiol. China 2001, 28, 26–30. [Google Scholar] [CrossRef]
- Yu, Q.Y.; Chen, S.Y.; Ma, Z.Y.; Wang, J.F.; Wei, L. Screening of phosphorus bacteria and its effects on maize growth at seedling stage. Ecol. Environ. Sci. 2012, 7, 1257–1261. [Google Scholar] [CrossRef]
- Yu, W.B.; Yang, X.M.; Shen, Q.R.; Xu, Y.C. Mechanism on phosphate solubilization of pseudomonas sp. K_3 and its phosphate solubilization ability under buffering condition. Plant Nutr. Fertile Sci. 2010, 16, 354–361. [Google Scholar]
- Moorberg, C.J.; Vepraskas, M.J.; Niewoehner, C.P. Phosphorus dissolution in the rhizosphere of bald cypress trees in restored wetland soils wetland soils. Soil Sci. Soc. Am. J. 2015, 79, 343–355. [Google Scholar] [CrossRef]
- Kadamur, G.; Ross, E.M. Mammalian phospholipase C. Annu. Rev. Physiol. 2013, 75, 127–154. [Google Scholar] [CrossRef] [PubMed]
- Titball, R.W. Bacterial phospholipase C. Microbiol. Rev. 1993, 57, 347–366. [Google Scholar] [PubMed]
- Essen, L.O.; Perisic, O.; Katan, M.; Wu, Y.; Roberts, M.F.; Williams, R.L. Structural mapping of the catalytic mechanism for a mammalian phosphoinositide-specific phospholipase C. Biochemistry 1997, 36, 1704. [Google Scholar] [CrossRef] [PubMed]
- Kolesnikov, Y.S.; Nokhrina, K.P.; Kretynin, S.V.; Volotovski, I.D.; Martinec, J.; Romanov, G.A.; Kravets, V.S. Molecular structure of phospholipase D and regulatory mechanisms of its activity in plant and animal cells. Biochem. Biokhimiia 2012, 77, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.S. Screening of Phospholipase Producing Strain C and Study on Its Enzymatic Properties. Ph.D. Thesis, Hunan Agricultural University, Changsha, China, 2010. [Google Scholar]
- Dai, S.L.; Zhang, J.; Shang, J. Screening and identification of high efficiency producing bacteria from phospholipase D. Jiangsu Agric. Sci. 2013, 41, 309–311. [Google Scholar]
- Yang, X.L.; Zhang, L.; Wang, Y.D. Screening and identification of high yield phospholipase D strain and optimization of fermentation conditions. Jiangsu Agric. Sci. 2016, 44, 521–525. [Google Scholar]
- Foster, D.A. Targeting Phospholipase D-mediated Survival Signals in Cancer. Curr. Signal Transduct. Ther. 2006, 1, 295–303. [Google Scholar] [CrossRef]
- Balboa, M.A.; Firestein, B.L.; Godson, C.; Bell, K.S.; Insel, P.A. Protein kinase C alpha mediates phospholipase D activation by nucleotides and phorbol ester in Madin-Darby canine kidney cells. Stimulation of phospholipase D is independent of activation of polyphosphoinositide-specific phospholipase C and phospholipase A. J. Biol. Chem. 1994, 269, 10511–10516. [Google Scholar] [PubMed]
- Pierzynski, G.M.; Sims, J.T.; Vance, G.F. Soils and Environmental Quality, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 145–146. ISBN 0-8493-1616-2. [Google Scholar]
- Li, Q. Study on Soil Microbial Community Structure Based on High Throughput Sequencing Technology. Ph.D. Thesis, Shandong Normal University, Jinan, China, 2015. [Google Scholar]
- Gao, L.; Zhou, W.; Huang, J.; He, S.; Yan, Y.; Zhu, W. Nitrogen removal by the enhanced floating treatment wetlands from the secondary effluent. Bioresour. Technol. 2017, 234, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Quaiser, A.; Ochsenreiter, T.; Lanz, C.; Schuster, S.C.; Treusch, A.H.; Eck, J. Acidobacteria form a coherent but highly diverse group within the bacterial domain: Evidence from environmental genomics. Mol. Microbiol. 2003, 50, 563–575. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The size (2b/a) of the P solubilizing ring of 13 preliminary screened bacteria from Acorus calamus L. and Iris tectorum Maxim. (a); Concentration of dissolved P (b), pH (c) and P-solubilizing and -releasing rate (d) by 0 hour, 48 h, and 96 h in Ca3(PO4)2 liquid medium. Error bars represent the standard errors.
Figure 1.
The size (2b/a) of the P solubilizing ring of 13 preliminary screened bacteria from Acorus calamus L. and Iris tectorum Maxim. (a); Concentration of dissolved P (b), pH (c) and P-solubilizing and -releasing rate (d) by 0 hour, 48 h, and 96 h in Ca3(PO4)2 liquid medium. Error bars represent the standard errors.
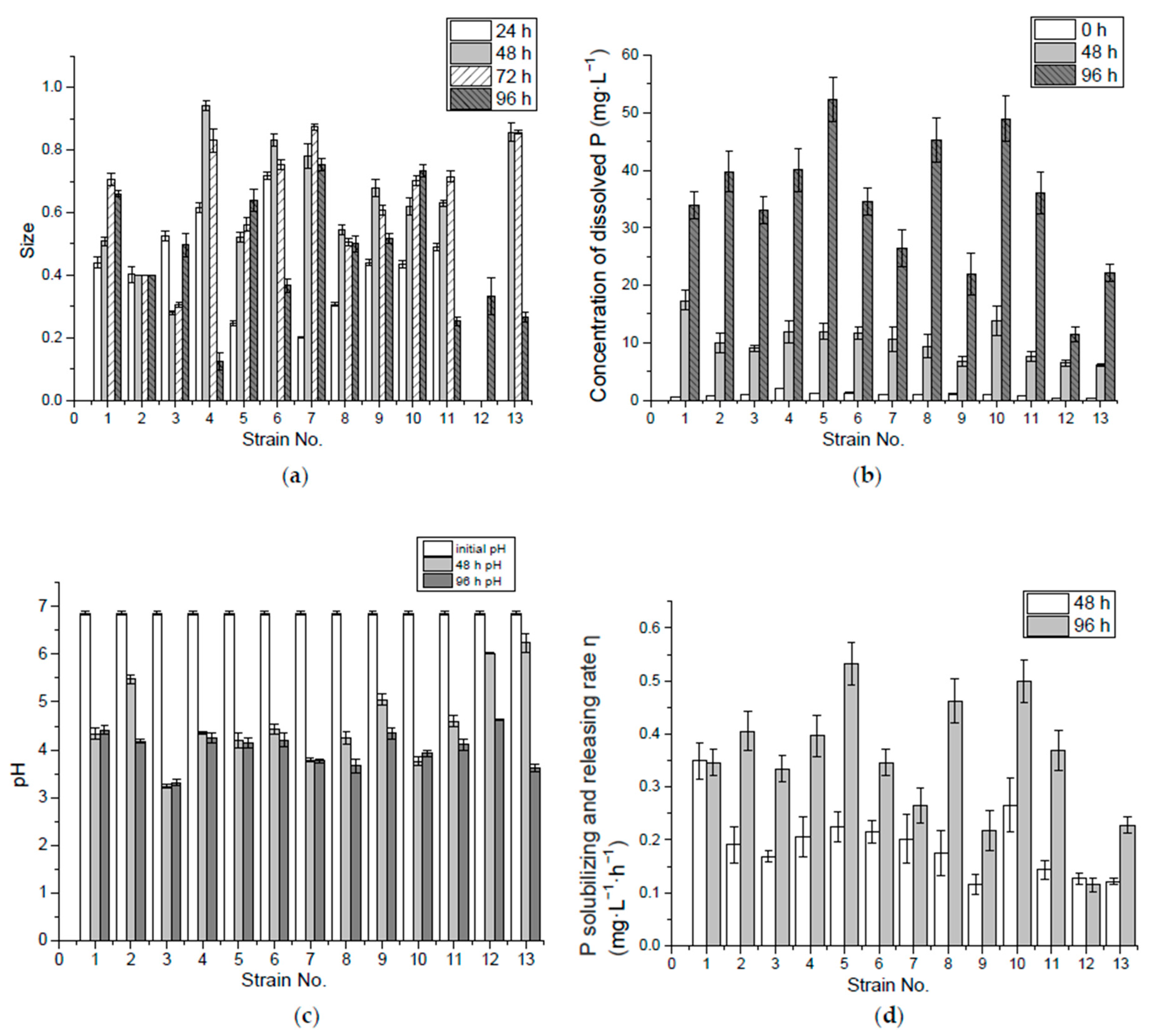
Figure 2.
Concentration of dissolved P (mg·L−1) (a) and pH (b) in 10-day incubation in liquid medium with insoluble Ca3(PO4)2.
Figure 2.
Concentration of dissolved P (mg·L−1) (a) and pH (b) in 10-day incubation in liquid medium with insoluble Ca3(PO4)2.
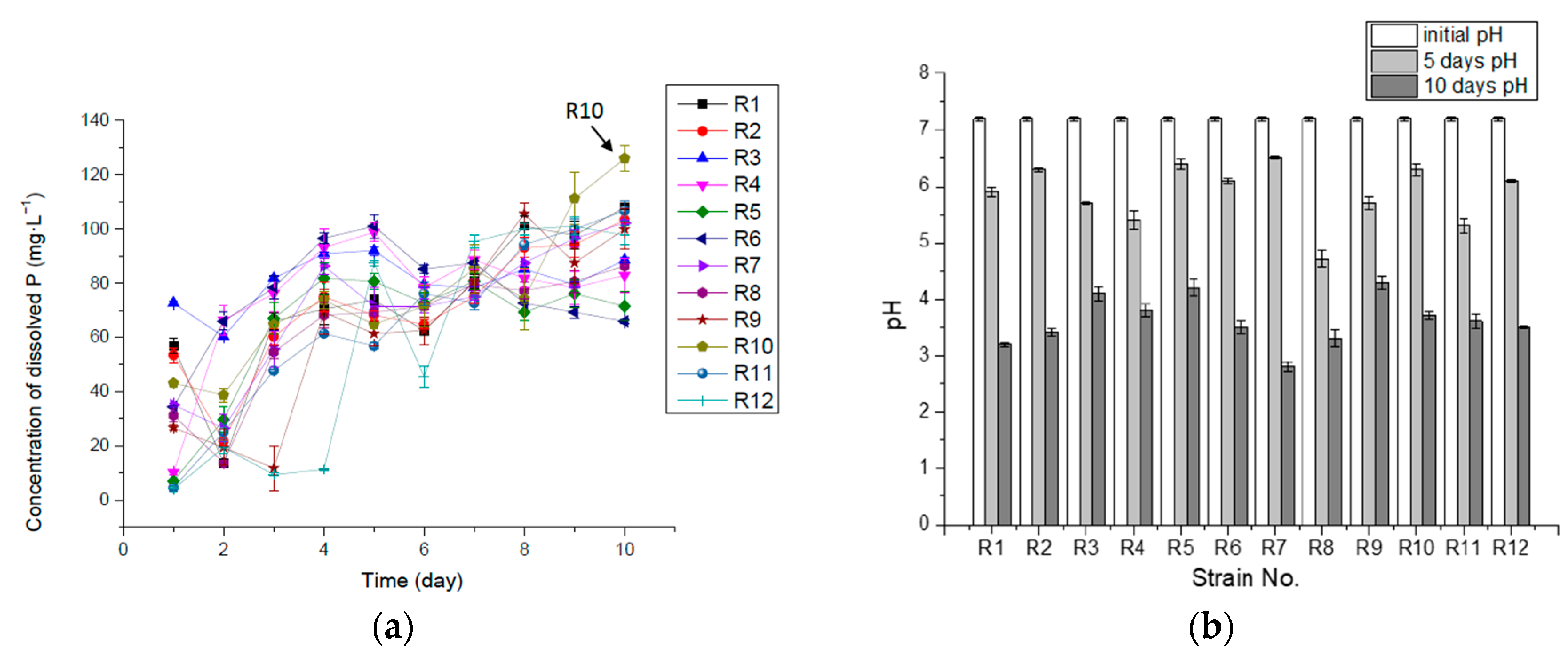
Figure 3.
Size of P-solubilizing ring formed by 13 preliminary screened bacteria in the solid medium with lecithin. The microbes were sampled from rhizospheres of Acorus calamus L., and Iris tectorum Maxim. around Tinyin Lake. (a) shows that the ring size varying by 24, 48, 72 and 96 h. Concentration of dissolved P (b), pH (c) and P solubilizing and releasing rate (d) by 0 hour, 48 h and 96 h in liquid medium with organic P (lecithin).
Figure 3.
Size of P-solubilizing ring formed by 13 preliminary screened bacteria in the solid medium with lecithin. The microbes were sampled from rhizospheres of Acorus calamus L., and Iris tectorum Maxim. around Tinyin Lake. (a) shows that the ring size varying by 24, 48, 72 and 96 h. Concentration of dissolved P (b), pH (c) and P solubilizing and releasing rate (d) by 0 hour, 48 h and 96 h in liquid medium with organic P (lecithin).
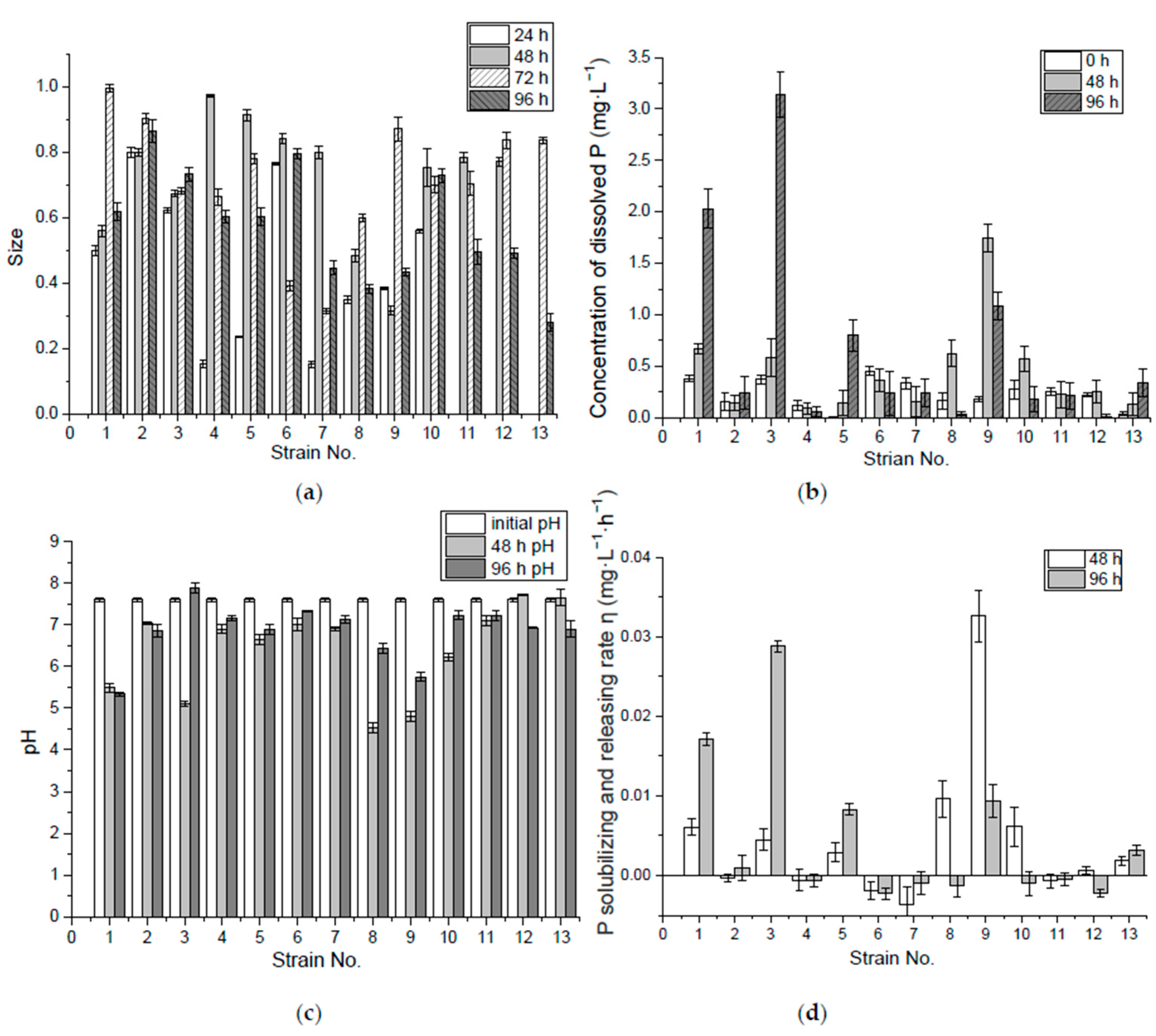
Figure 4.
Concentration of dissolved P (mg·L−1) (a) and pH (b) in 10-day incubation in liquid medium with organic P (lecithin).
Figure 4.
Concentration of dissolved P (mg·L−1) (a) and pH (b) in 10-day incubation in liquid medium with organic P (lecithin).
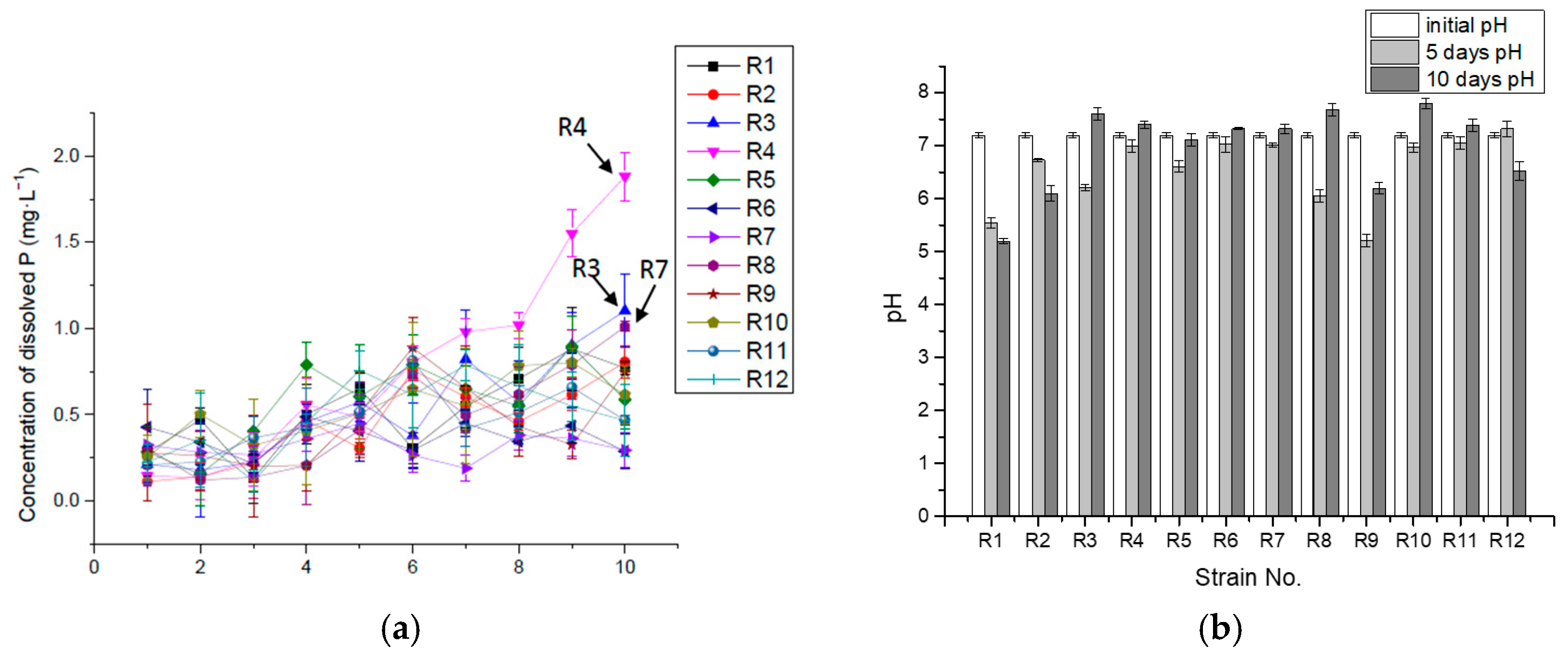
Figure 5.
Correlation of solubilized P and pH in Ca3(PO4)2 liquid medium without seeding the isolates where the pH was adjusted by hydrochloric acid (a) and seeding with the isolated bacteria (b).
Figure 5.
Correlation of solubilized P and pH in Ca3(PO4)2 liquid medium without seeding the isolates where the pH was adjusted by hydrochloric acid (a) and seeding with the isolated bacteria (b).
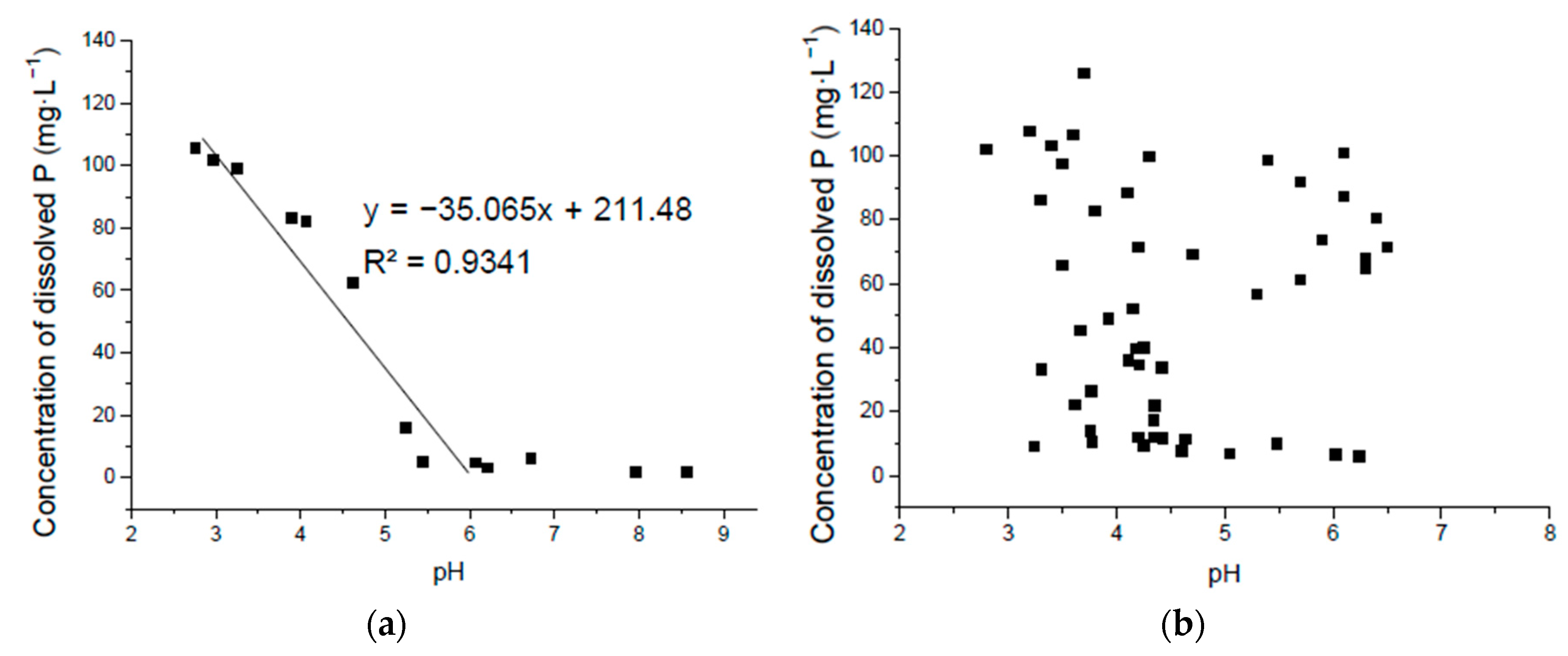
Figure 6.
Bacteria community analysis pie plot at the Phylum (a), Genus (b) and Species (c) level sampled from rhizosphere of the experimental plant. Genera making up less than 1% of the total composition are defined as “others”.
Figure 6.
Bacteria community analysis pie plot at the Phylum (a), Genus (b) and Species (c) level sampled from rhizosphere of the experimental plant. Genera making up less than 1% of the total composition are defined as “others”.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Maximum concentration and rate of P release in 10-day incubation in liquid medium with insoluble Ca3(PO4)2.
Table 1.
Maximum concentration and rate of P release in 10-day incubation in liquid medium with insoluble Ca3(PO4)2.
| Strain No. | R1 | R2 | R3 | R4 | R5 | R6 | R7 | R8 | R9 | R10 | R11 | R12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Max Concentration of dissolved P (mg·L−1) | 107.77 | 103.24 | 91.92 | 98.71 | 81.73 | 100.97 | 102.11 | 86.26 | 105.5 | 125.88 | 106.63 | 100.97 |
| Max Con. Day (day) | 10 | 10 | 5 | 5 | 4 | 5 | 10 | 10 | 8 | 10 | 10 | 9 |
| Max rate of solubilizing P (mg·L−1·h−1) | 2.37 | 2.23 | 3.03 | 1.06 | 0.93 | 1.44 | 1.46 | 1.30 | 1.11 | 1.80 | 0.66 | 0.73 |
| Max rate time (hours) | 24 | 24 | 24 | 48 | 72 | 24 | 24 | 24 | 24 | 24 | 72 | 120 |
| P releasing rate η in 10 days (mg·L−1·h−1) | 0.45 | 0.43 | 0.37 | 0.35 | 0.3 | 0.27 | 0.43 | 0.36 | 0.42 | 0.52 | 0.44 | 0.41 |
Table 2.
Maximum concentration and rate of P release for 10-day incubation in liquid medium with organic P (lecithin).
Table 2.
Maximum concentration and rate of P release for 10-day incubation in liquid medium with organic P (lecithin).
| Strain No. | R1 | R2 | R3 | R4 | R5 | R6 | R7 | R8 | R9 | R10 | R11 | R12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Max Concentration of P (mg·L−1) | 0.88 | 0.81 | 1.10 | 1.88 | 0.90 | 0.49 | 0.45 | 1.01 | 0.89 | 0.80 | 0.81 | 0.79 |
| Max Con. Day (day) | 9 | 10 | 10 | 10 | 9 | 4 | 5 | 10 | 6 | 9 | 6 | 7 |
| Max rate of releasing P (mg·L−1·h−1) | 0.0117 | 0.0053 | 0.0088 | 0.0078 | 0.0118 | 0.0178 | 0.0135 | 0.0126 | 0.0116 | 0.0106 | 0.0087 | 0.0093 |
| Max rate time (hours) | 24 | 144 | 24 | 240 | 24 | 24 | 24 | 24 | 24 | 24 | 24 | 24 |
| P releasing rate η in 10 days (mg·L−1·h−1) | 0.0032 | 0.0033 | 0.0046 | 0.0078 | 0.0024 | 0.0012 | 0.0012 | 0.0042 | 0.0031 | 0.0026 | 0.0019 | 0.0019 |
Table 3.
Dominant phylum, high quality sequences, richness and diversity estimators of the rhizosphere sample (CaoR1-B), and comparison with other wetland environments.
Table 3.
Dominant phylum, high quality sequences, richness and diversity estimators of the rhizosphere sample (CaoR1-B), and comparison with other wetland environments.
| Sample | OTU | Sequences | Shannon | Simpson | Ace | Chao | Species | Genus | Dominant Phylum | Proportion of Dominant |
|---|---|---|---|---|---|---|---|---|---|---|
| CaoR1-B | 445 | 30212 | 5.521541 | 0.00652 | 445 | 445 | 274 | 179 | Acidobacteria | 34.9% |
| Yellow river Delta Wetland[47] | 30874 | Proteobacteria | 44.67% | |||||||
| Floating treatment wetland [48] | 11477 | 78242 | 8.49 | 0.9931 | 2105 | 1603 | Proteobacteria | 46.4% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cao, Y.; Fu, D.; Liu, T.; Guo, G.; Hu, Z. Phosphorus Solubilizing and Releasing Bacteria Screening from the Rhizosphere in a Natural Wetland. Water 2018, 10, 195. https://doi.org/10.3390/w10020195
AMA Style
Cao Y, Fu D, Liu T, Guo G, Hu Z. Phosphorus Solubilizing and Releasing Bacteria Screening from the Rhizosphere in a Natural Wetland. Water. 2018; 10(2):195. https://doi.org/10.3390/w10020195
Chicago/Turabian StyleCao, Ying, Dafang Fu, Tingfeng Liu, Guang Guo, and Zhixin Hu. 2018. "Phosphorus Solubilizing and Releasing Bacteria Screening from the Rhizosphere in a Natural Wetland" Water 10, no. 2: 195. https://doi.org/10.3390/w10020195
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.