Gut Microbiota of Great Spotted Cuckoo Nestlings is a Mixture of Those of Their Foster Magpie Siblings and of Cuckoo Adults

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Field Work
2.2. Ethics Statement
2.3. Laboratory Work
2.4. Sequence Data Analysis and Taxonomic Identification
2.5. Statistics
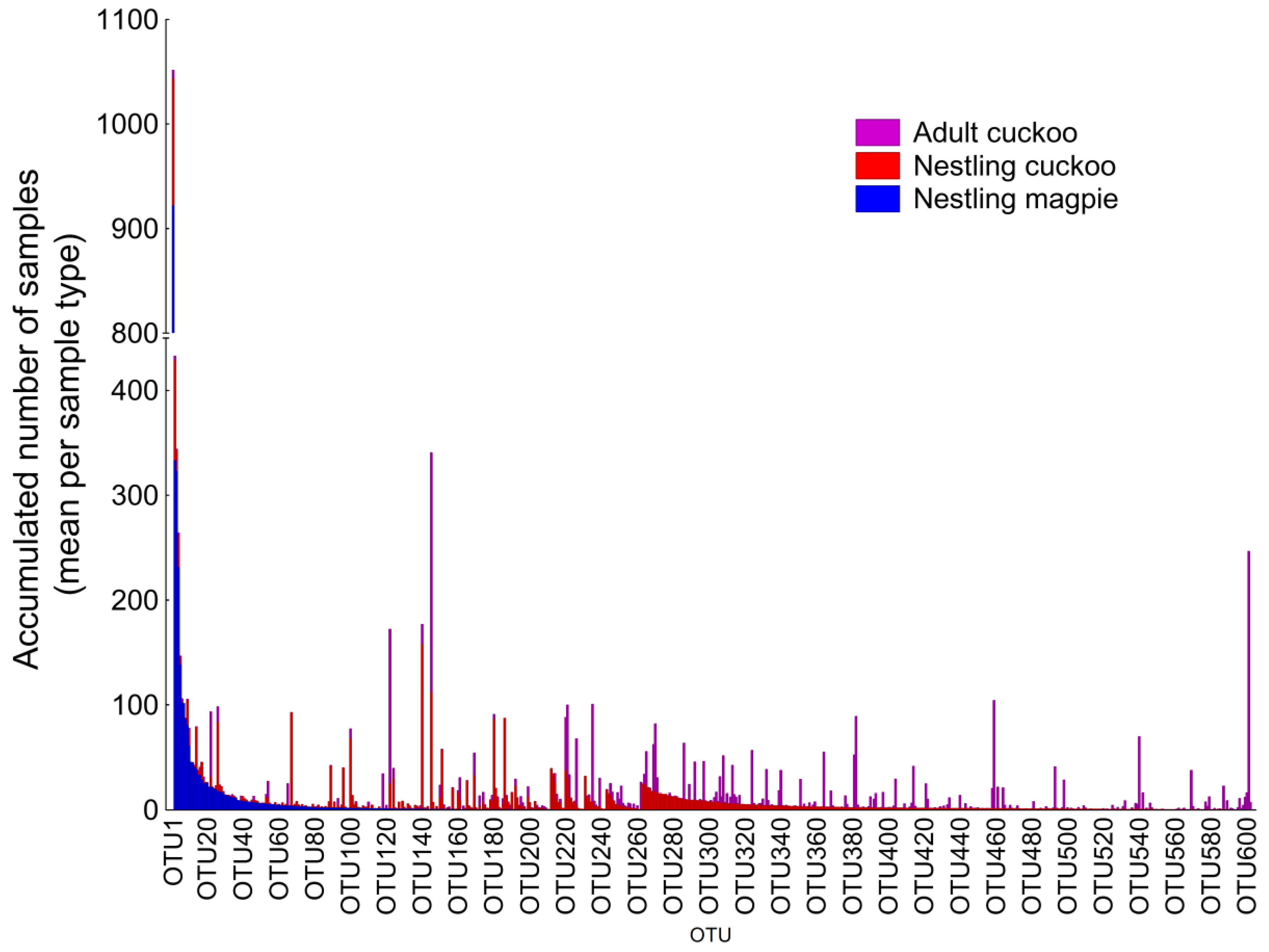
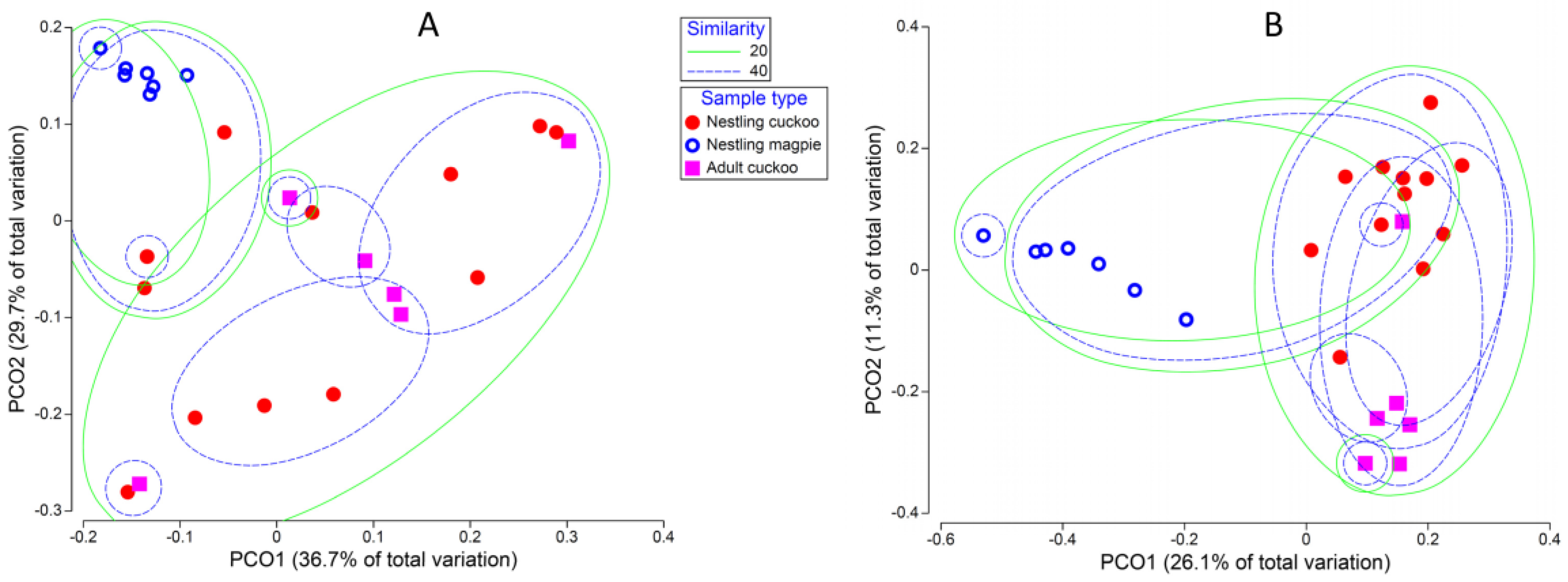
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Parfrey, L.W.; Moreau, C.S.; Russell, J.A. Introduction: The host-associated microbiome: Pattern, process and function. Mol. Ecol. 2018, 27, 1749–1765. [Google Scholar] [CrossRef] [PubMed]
- McFall-Ngai, M.; Hadfield, M.G.; Bosch, T.C.G.; Carey, H.V.; Domazet-Loso, T.; Douglas, A.E.; Dubilier, N.; Eberl, G.; Fukami, T.; Gilbert, S.F.; et al. Animals in a bacterial world, a new imperative for the life sciences. Proc. Natl. Acad. Sci. USA 2013, 110, 3229–3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colston, T.J.; Jackson, C.R. Microbiome evolution along divergent branches of the vertebrate tree of life: What is known and unknown. Mol. Ecol. 2016, 25, 3776–3800. [Google Scholar] [CrossRef] [PubMed]
- Bordenstein, S.R.; Theis, K.R. Host biology in light of the microbiome: Ten principles of holobionts and hologenomes. PLoS Biol. 2015, 13, e1002226. [Google Scholar] [CrossRef] [PubMed]
- Doolittle, W.F.; Booth, A. It’s the song, not the singer: An exploration of holobiosis and evolutionary theory. Biol. Philos. 2017, 32, 5–24. [Google Scholar] [CrossRef]
- Moran, N.A.; Sloan, D.B. The hologenome concept: Helpful or hollow? PLoS Biol. 2015, 13, e1002311. [Google Scholar] [CrossRef] [PubMed]
- Fraune, S.; Bosch, T.C.G. Why bacteria matter in animal development and evolution. BioEssays 2010, 32, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Hird, S.M. Evolutionary biology needs wild microbiomes. Front. Microbiol. 2017, 8, 725. [Google Scholar] [CrossRef] [PubMed]
- Lucas, F.S.; Heeb, P. Environmental factors shape cloacal bacterial assemblages in great tit Parus major and blue tit P. caeruleus nestlings. J. Avian Biol. 2005, 36, 510–516. [Google Scholar] [CrossRef]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [PubMed]
- Muegge, B.D.; Kuczynski, J.; Knights, D.; Clemente, J.C.; González, A.; Fontana, L.; Henrissat, B.; Knight, R.; Gordon, J.I. Diet drives convergence in gut microbiome functions across mammalian phylogeny and within humans. Science 2011, 332, 970–974. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2013, 505, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donaldson, G.P.; Ladinsky, M.S.; Yu, K.B.; Sanders, J.G.; Yoo, B.B.; Chou, W.C.; Conner, M.E.; Earl, A.M.; Knight, R.; Bjorkman, P.J.; et al. Gut microbiota utilize immunoglobulin a for mucosal colonization. Science 2018, 360, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Playfair, J.; Bancroft, G. Infection and Immunity, 2nd ed.; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Randal, B.R.; Everett, M.L.; Palestrant, D.; Love, S.D.; Lin, S.S.; Parker, W. Human secretory immunoglobulin a may contribute to biofilm formation in the gut. Immunology 2003, 109, 580–587. [Google Scholar] [CrossRef]
- Davenport, E.R. Elucidating the role of the host genome in shaping microbiome composition. Gut Microbes 2016, 7, 178–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohl, K.D.; Dearing, M.D.; Bordenstein, S.R. Microbial communities exhibit host species distinguishability and phylosymbiosis along the length of the gastrointestinal tract. Mol. Ecol. 2018, 27, 1874–1883. [Google Scholar] [CrossRef] [PubMed]
- Amato, K.R. Co-evolution in context: The importance of studying gut microbiomes in wild animals. Microbiome Sci. Med. 2013, 1, 10–29. [Google Scholar] [CrossRef]
- Hird, S.M.; Carstens, B.C.; Cardiff, S.W.; Dittmann, D.L.; Brumfield, R.T. Sampling locality is more detectable than taxonomy or ecology in the gut microbiota of the brood-parasitic brown-headed cowbird (Molothrus ater). PeerJ 2014, 2, e321. [Google Scholar] [CrossRef] [PubMed]
- Roldán, M.; Soler, M. Parental-care parasitism: How do unrelated offspring attain acceptance by foster parents? Behav. Ecol. 2011, 22, 679–691. [Google Scholar] [CrossRef]
- Rodríguez, J.M.; Murphy, K.; Stanton, C.; Ross, R.P.; Kober, O.I.; Juge, N.; Avershina, E.; Rudi, K.; Narbad, A.; Jenmalm, M.C.; et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb. Ecol. Health Dis. 2015, 26, 26050. [Google Scholar] [CrossRef] [PubMed]
- Benskin, C.M.H.; Rhodes, G.; Pickup, R.W.; Wilson, K.; Hartley, I.R. Diversity and temporal stability of bacterial communities in a model passerine bird, the zebra finch. Mol. Ecol. 2010, 19, 5531–5544. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Rodríguez, M.; Lucas, F.S.; Heeb, P.; Soler, J.J. Differences in intestinal microbiota between avian brood parasites and their hosts. Biol. J. Linn. Soc. 2009, 96, 406–414. [Google Scholar] [CrossRef] [Green Version]
- Soler, M.; Martínez, J.G.; Soler, J.J.; Møller, A.P. Preferential allocation of food by magpie Pica pica to great spotted cuckoo Clamator glandarius chicks. Behav. Ecol. Sociobiol. 1995, 37, 7–13. [Google Scholar] [CrossRef]
- Soler, J.J.; Møller, A.P.; Soler, M.; Martínez, J.G. Interactions between a brood parasite and its host in relation to parasitism and immune defence. Evol. Ecol. Res. 1999, 1, 189–210. [Google Scholar]
- Koch, H.; Schmid-Hempel, P. Socially transmitted gut microbiota protect bumble bees against an intestinal parasite. Proc. Natl. Acad. Sci. USA 2011, 108, 19288–19292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Rodríguez, M.; Soler, J.J.; Lucas, F.S.; Heeb, P.; Palacios, M.; Martín-Gálvez, D.; De Neve, L.; Pérez-Contreras, T.; Martínez, J.G.; Soler, M. Bacterial diversity at the cloaca relates to an immune response in magpie Pica pica and to body condition of great spotted cuckoo Clamator glandarius nestlings. J. Avian Biol. 2009, 40, 42–48. [Google Scholar] [CrossRef]
- Gloor, G.B.; Hummelen, R.; Macklaim, J.M.; Dickson, R.J.; Fernandes, A.D.; MacPhee, R.; Reid, G. Microbiome profiling by illumina sequencing of combinatorial sequence-tagged PCR products. PLoS ONE 2010, 5, e15406. [Google Scholar] [CrossRef] [PubMed]
- Bartram, A.K.; Lynch, M.D.J.; Stearns, J.C.; Moreno-Hagelsieb, G.; Neufeld, J.D. Generation of multimillion-sequence 16S rRNA gene libraries from complex microbial communities by assembling paired-end illumina reads. Appl. Environ. Microbiol. 2011, 77, 3846–3852. [Google Scholar] [CrossRef] [PubMed]
- Del Hoyo, J.; Elliott, A.; Christie, D.A. Handbook of the Birds of the World; Lynx Edicions: Barcelona, Spain, 1997; Volume IV. [Google Scholar]
- Martínez, J.G.; Soler, M.; Soler, J.J.; Paracuellos, M.; Sánchez, J. Alimentación de los pollos de urraca (Pica pica) en relacion con la edad y disponibilidad de presas. Ardeola 1992, 39, 35–48. [Google Scholar]
- Kohl, K.D. Diversity and function of the avian gut microbiota. J. Comp. Physiol. B-Biochem. Syst. Environ. Physiol. 2012, 182, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Waite, D.; Taylor, M. Exploring the avian gut microbiota: Current trends and future directions. Front. Microbiol. 2015, 6, 673. [Google Scholar] [CrossRef] [PubMed]
- Teyssier, A.; Lens, L.; Matthysen, E.; White, J. Dynamics of gut microbiota diversity during the early development of an avian host: Evidence from a cross-foster experiment. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Soler, J.J.; Soler, M. Brood-parasite interactions between great spotted cuckoos and magpies: A model system for studying coevolutionary relationships. Oecologia 2000, 125, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Soler, J.J.; Martin-Galvez, D.; De Neve, L.; Soler, M. Brood parasitism correlates with the strength of spatial autocorrelation of life history and defensive traits in magpies. Ecology 2013, 94, 1338–1346. [Google Scholar] [CrossRef] [PubMed]
- Soler, M.; Soler, J.J. Growth and development of great spotted cuckoos and their magpie host. Condor 1991, 93, 49–54. [Google Scholar] [CrossRef]
- Martinez-Garcia, A.; Martin-Vivaldi, M.; Rodriguez-Ruano, S.M.; Peralta-Sanchez, J.M.; Valdivia, E.; Soler, J.J. Nest bacterial environment affects microbiome of hoopoe eggshells, but not that of the uropygial secretion. PLoS ONE 2016, 11, e0158158. [Google Scholar] [CrossRef] [PubMed]
- Earth-Microbiome-Project. 16S Illumina Amplicon Protocol. Available online: http://press.igsb.anl.gov/earthmicrobiome/protocols-and-standards/16s/ (accessed on 20 Jun 2017).
- Nikkari, S.; Lopez, F.A.; Lepp, P.W.; Cieslak, P.R.; Ladd-Wilson, S.; Passaro, D.; Danila, R.; Relman, D.A. Broad-range bacterial detection and the analysis of unexplained death and critical illness. Emerg. Infect. Dis. 2002, 8, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qian, P.Y. Conservative fragments in bacterial 16S rRNA genes and primer design for 16S ribosomal DNA amplicons in metagenomic studies. PLoS ONE 2009, 4, e7401. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navas-Molina, J.A.; Peralta-Sánchez, J.M.; González, A.; McMurdie, P.J.; Vázquez-Baeza, Y.; Xu, Z.; Ursell, L.K.; Lauber, C.; Zhou, H.; Song, S.J.; et al. Advancing our understanding of the human microbiome using qiime. In Methods in Enzymology; DeLong, E.F., Ed.; Academic Press: Cambridge, MA, USA, 2013; Volume 531, pp. 371–444. [Google Scholar]
- Dumbrell, A.J.; Ferguson, R.M.W.; Clark, D.R. Microbial community analysis by single-amplicon high-throughput next generation sequencing: Data analysis—From raw output to ecology. In Hydrocarbon and Lipid Microbiology Protocols: Microbial Quantitation, Community Profiling and Array Approaches; McGenity, T.J., Timmis, K.N., Nogales, B., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 155–206. [Google Scholar]
- Knight, R.; Vrbanac, A.; Taylor, B.C.; Aksenov, A.; Callewaert, C.; Debelius, J.; Gonzalez, A.; Kosciolek, T.; McCall, L.-I.; McDonald, D.; et al. Best practices for analysing microbiomes. Nat. Rev. Microbiol. 2018, 16, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Aronesty, E. ea-utils: Command-Line Tools for Processing Biological Sequencing Data. Available online: https://github.com/ExpressionAnalysis/ea-utils (accessed on 20 June 2017).
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rideout, J.R.; He, Y.; Navas-Molina, J.A.; Walters, W.A.; Ursell, L.K.; Gibbons, S.M.; Chase, J.; McDonald, D.; Gonzalez, A.; Robbins-Pianka, A.; et al. Subsampled open-reference clustering creates consistent, comprehensive OTU definitions and scales to billions of sequences. PeerJ 2014, 2, e545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16s rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of Bacteria and Archaea. ISME J. 2011, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from illumina amplicon sequencing. Nat. Methods 2012, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Knight, R. Unifrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed]
- Dell-Inc. Dell Statistica (data analysis software system), version 13.
- Anderson, M.; Gorley, R.; Clarke, K. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Davies, N.B. Cuckoos, Cowbirds, and Others Cheats; T & A D Poyser: London, UK, 2000. [Google Scholar]
- Brooks, A.W.; Kohl, K.D.; Brucker, R.M.; van Opstal, E.J.; Bordenstein, S.R. Phylosymbiosis: Relationships and functional effects of microbial communities across host evolutionary history. PLoS Biol. 2016, 14, e2000225. [Google Scholar] [CrossRef] [PubMed]
- McNab, J.M. The avian caeca: A review. World’s Poult. Sci. J. 2007, 29, 251–263. [Google Scholar] [CrossRef]
- Mead, G.C. Microbes of the avian cecum: Types present and substrates utilized. J. Exp. Zool. 1989, 252, 48–54. [Google Scholar] [CrossRef]
- Józefiak, D.; Rutkowski, A.; Martin, S.A. Carbohydrate fermentation in the avian ceca: A review. Anim. Feed Sci. Technol. 2004, 113, 1–15. [Google Scholar] [CrossRef]
- Soler, M.; Soler, J.J. Innate versus learned recognition of conspecifics in great spotted cuckoos clamator glandarius. Anim. Cogn. 1999, 2, 97–102. [Google Scholar] [CrossRef]
- Van Veelen, H.P.J.; Salles, J.F.; Tieleman, B.I. Microbiome assembly of avian eggshells and their potential as transgenerational carriers of maternal microbiota. ISME J. 2018, 12, 1375–1388. [Google Scholar] [CrossRef] [PubMed]
- Soler, J.J.; Peralta-Sánchez, J.M.; Martínez-Bueno, M.; Martín-Vivaldi, M.; Martín-Gálvez, D.; Vela, A.I.; Briones, V.; Pérez-Contreras, T. Brood parasitism is associated with increased bacterial contamination of host eggs: Bacterial loads of host and parasitic eggs. Biol. J. Linn. Soc. 2011, 103, 836–848. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| VARIABLE CONSIDERED | TYPE OF SAMPLE | COMPARISONS | ||||||
|---|---|---|---|---|---|---|---|---|
| (A) | (B) | (C) | ANOVA | POST-HOC (Tukey tests) | ||||
| Adult cuckoos | Nestling cuckoos | Nestling magpies | ||||||
| (N = 6) | (N = 12) | (N = 7) | A vs. B | A vs. C | B vs. C | |||
| Mean | Mean | Mean | F2,22 | p | p | p | p | |
| (SE) | (SE) | (SE) | (SE) | (SE) | ||||
| Bacterial Richness | 73.8 | 123 | 52.3 | 14.42 | 0.0001 | 0.008 | 0.405 | 0.0003 |
| (12.1) | (8.5) | (11.2) | (0.19) | (0.000008) | ||||
| α-diversity (Shannon index) | 3.92 | 4.45 | 2.66 | 6.50 | 0.006 | 0.585 | 0.106 | 0.005 |
| (0.43) | (0.31) | (0.4) | (0.03) | (0.00001) | ||||
| PERMANOVA | POST-HOC (Pair-wise t-tests) | |||||||
| Microbiome composition | Pseudo-F (SE) | p (SE) | p (t-value) | p (t-value) | p (t-value) | |||
| Weighed unifrac β-diversity | 4.35 | 0.00022 | 0.559 | 0.0002 | 0.0001 (2.63) | |||
| (0.01) | (0.00004) | (0.85) | (2.87) | |||||
| Unweighed unifrac β-diversity | 5.37 | 0.0001 | 0.0002 | 0.0007 | 0.0001 | |||
| (0.07) | (0.0001) | (1.72) | (2.47) | (2.61) | ||||
| Abundance of Families (Bray−Curtis distance matrices) | 5.33 | 0.00013 | 0.58 | 0.0008 | 0.002 (2.94) | |||
| (0.03) | (0.00002) | (0.9) | (3.00) | |||||
| Prevalence of Families (Jaccard distance matrices) | 6.43 | 0.0001 | 0.0034 | 0.0006 | 0.0001 (2.86) | |||
| (0.12) | (0.00001) | (1.62) | (2.55) | |||||
| Using Information of Microbial Abundance | Using Information of Microbial Prevalence | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bray-Curtis | Mean(SE) | ANOVAs | Jaccard | Prevalence (%) | Pearson χ2 | ||||||||
| Phylum | Family | Entered Best Models | Adult Cuckoos (N = 6) | Nestling Cuckoos (N = 12) | Nestling Magpies (N = 7) | F(2,22) | p | Entered Best Models | Adult Cuckoos (N = 6) | Nestling Cuckoos (N = 12) | Nestling Magpies (N = 7) | χ2 | p |
| Actinobacteria | Bifidobacteriaceae | 1 | 0.022 (0.020) | - | - | 1.931 | 0.169 | ||||||
| Corynebacteriaceae | 1–4 | 50.00 | 75.00 | 71.43 | 1.201 | 0.548 | |||||||
| Propionibacteriaceae | 1–3 | 100.00 | 66.67 | 100.00 | 5.159 | 0.076 | |||||||
| Nocardioidaceae | 1, 4 | 16.67 | 0.00 | 42.86 | 6.045 | 0.049 | |||||||
| Micrococcaceae | 4 | 0.00 | 0.00 | 42.86 | 8.767 | 0.012 | |||||||
| Bacteroidetes | Bacteroidaceae | 1–4 | 0.113 (0.072) | 0.148 (0.058) | 0.003 (0.002) | 1.732 | 0.2 | ||||||
| Porphyromonadaceae | 1–4 | 83.33 | 83.33 | 42.86 | 4.096 | 0.129 | |||||||
| Rikenellaceae | 1–4 | 83.33 | 83.33 | 0.00 | 14.583 | <0.001 | |||||||
| Odoribacteriaceae | 2, 3 | 33.33 | 66.67 | 0.00 | 8.333 | 0.016 | |||||||
| Barnesiellaceae | 1–4 | 83.33 | 58.33 | 14.29 | 6.542 | 0.038 | |||||||
| Paraprevotellaceae | 4 | 66.67 | 16.67 | 0.00 | 8.553 | 0.014 | |||||||
| Cyanobacteria | Cyanobacteria | 1–4 | 66.67 | 75.00 | 0.00 | 10.644 | 0.005 | ||||||
| Firmicutes | Lachnospiraceae | 1–4 | 0.220 (0.095) | 0.228 (0.065) | 0.012 (0.008) | 3.036 | 0.069 | ||||||
| Other clostridiales | 2, 3 | 0.169 (0.103) | 0.093 (0.046) | 0.006 (0.005) | 2.083 | 0.148 | 1, 3 | 100.00 | 100.00 | 42.86 | 12.245 | 0.002 | |
| Ruminococcaceae | 1, 3, 4 | 0.074 (0.034) | 0.074 (0.019) | 0.00004 (0.00004) | 3.547 | 0.046 | |||||||
| Clostridiaceae | 1–4 | 0.002 (0.001) | 0.014 (0.007) | 0.070 (0.047) | 2.146 | 0.141 | 1–4 | 33.33 | 91.67 | 85.71 | 7.965 | 0.019 | |
| Veillonellaceae | 2, 3 | 0.103 (0.084) | 0.008 (0.004) | 0 | 2.251 | 0.129 | 1–4 | 66.67 | 66.67 | 0.00 | 8.974 | 0.011 | |
| Erypsipelotrichaceae | 2 | 100.00 | 100.00 | 28.57 | 16.071 | <0.001 | |||||||
| Staphyllococcaceae | 4 | 50.00 | 50.00 | 42.86 | 0.103 | 0.95 | |||||||
| Other Lactobacillales | 2, 3 | 16.67 | 33.33 | 85.71 | 7.317 | 0.026 | |||||||
| Eubacteriaceae | 1, 2 | 16.67 | 25.00 | 0.00 | 2.059 | 0.357 | |||||||
| Enterococcaceae | 1–4 | 0.00 | 16.67 | 71.43 | 9.647 | 0.008 | |||||||
| Proteobacteria | Enterobacteriaceae | 1–4 | 0.005 (0.003) | 0.104 (0.061) | 0.630 (0.112) | 17.447 | <0.001 | ||||||
| Campylobacteraceae | 1, 2 | 0 | 0.021 (0.021) | 0.159 (0.129) | 1.555 | 0.233 | |||||||
| Helicobacteriaceae | 4 | 0 | 0.015 (0.015) | 0 | 0.526 | 0.598 | |||||||
| Other Alphaproteobacteria | 1 | 50.00 | 91.67 | 0.00 | 15.192 | <0.001 | |||||||
| Pseudomonadaceae | 2 | 0.00 | 25.00 | 57.14 | 5.336 | 0.069 | |||||||
| Brucellaceae | 4 | 16.67 | 16.67 | 0.00 | 1.326 | 0.515 | |||||||
| Succinivibrionaceae | 1 | 50.00 | 0.00 | 0.00 | 10.795 | 0.0045 | |||||||
| Alcanigenaceae | 3, 4 | 33.33 | 75.00 | 0.00 | 10.457 | 0.0054 | |||||||
| Verrucomicrobia | Verrucomicrobiaceae | 3 | 16.67 | 41.67 | 0.00 | 4.441 | 0.109 | ||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Rodríguez, M.; Martín-Vivaldi, M.; Martínez-Bueno, M.; Soler, J.J. Gut Microbiota of Great Spotted Cuckoo Nestlings is a Mixture of Those of Their Foster Magpie Siblings and of Cuckoo Adults. Genes 2018, 9, 381. https://doi.org/10.3390/genes9080381
Ruiz-Rodríguez M, Martín-Vivaldi M, Martínez-Bueno M, Soler JJ. Gut Microbiota of Great Spotted Cuckoo Nestlings is a Mixture of Those of Their Foster Magpie Siblings and of Cuckoo Adults. Genes. 2018; 9(8):381. https://doi.org/10.3390/genes9080381
Chicago/Turabian StyleRuiz-Rodríguez, Magdalena, Manuel Martín-Vivaldi, Manuel Martínez-Bueno, and Juan José Soler. 2018. "Gut Microbiota of Great Spotted Cuckoo Nestlings is a Mixture of Those of Their Foster Magpie Siblings and of Cuckoo Adults" Genes 9, no. 8: 381. https://doi.org/10.3390/genes9080381