A Herpes Simplex Virus Thymidine Kinase-Induced Mouse Model of Hepatocellular Carcinoma Associated with Up-Regulated Immune-Inflammatory-Related Signals


Abstract
:1. Introduction
2. Methods
2.1. Generation of HSV-tk Transgenic Mice
2.2. Immunofluorescence Analysis
2.3. The Incidence of HCC and Histopathological or Immunohistochemistry Analysis in HSV-tk Mice
2.4. Microarray Data Analysis
2.5. Gene Functional Enrichment Analyses
2.6. Comparison of HCC Gene Expression Profiles between HSV-tk and Notch Transgenic Mice
3. Results
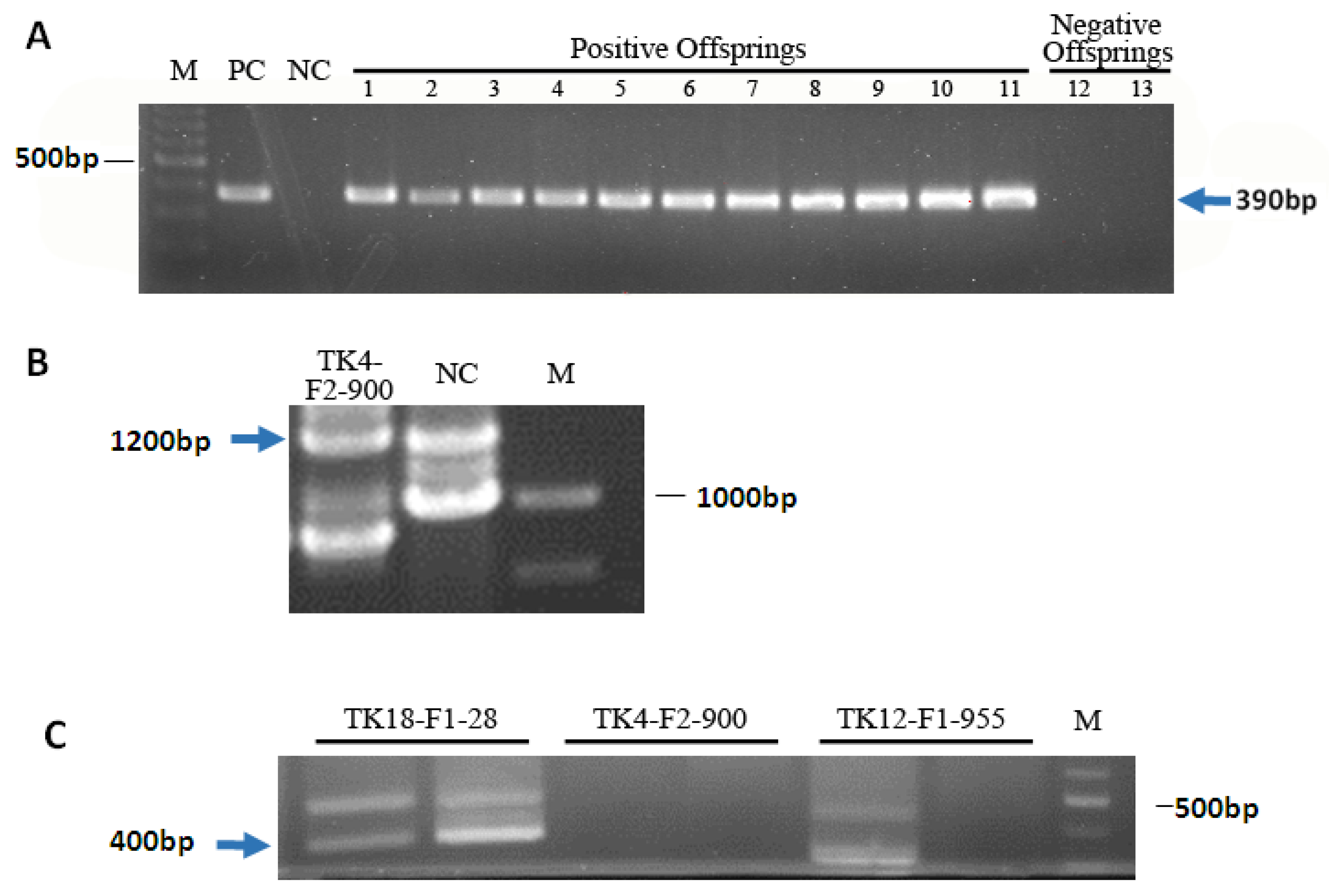
3.1. Generation and Characterization of HSV-tk Mice
3.2. The Incidence of HCC and Histopathological Analyses in HSV-tk Mice
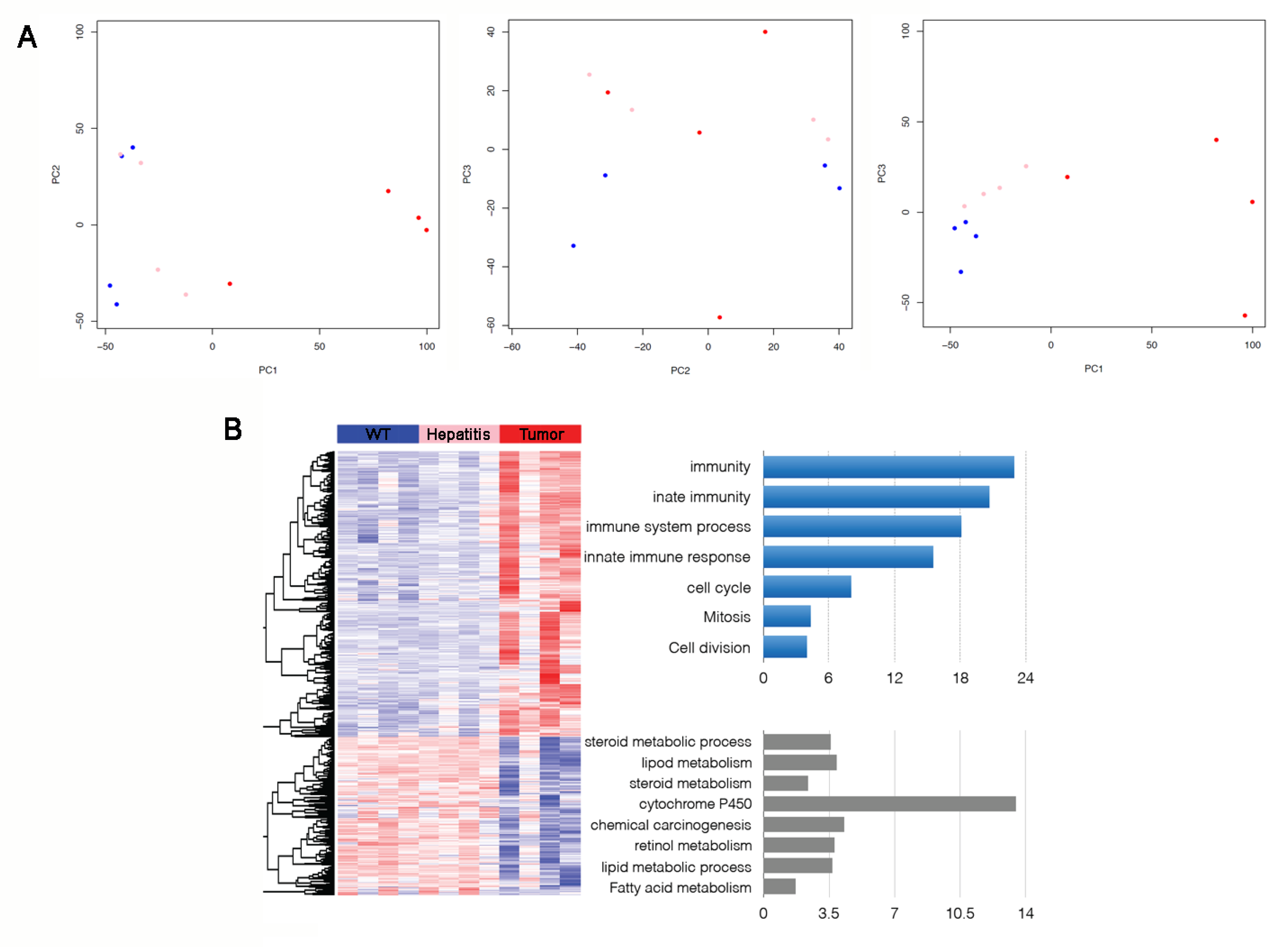
3.3. Immune/Inflammatory and Cell Cycle Abnormalities in HSV-tk Mice Tumors
3.4. Comparison of HCC Gene Expression Profiles between HSV-tk and Notch Transgenic Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Michelotti, G.A.; Machado, M.V.; Diehl, A.M. NAFLD, NASH and liver cancer. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Forner, A.; Llovet, J.M.; Bruix, J. Hepatocellular carcinoma. Lancet 2012, 379, 1245–1255. [Google Scholar] [CrossRef]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Ladd, B.; O’Konek, J.J.; Ostruszka, L.J.; Shewach, D.S. Unrepairable DNA double-strand breaks initiate cytotoxicity with HSV-TK/ganciclovir. Cancer Gene Ther. 2011, 18, 751–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rainov, N.G.; Fels, C.; Droege, J.W.; Schäfer, C.; Kramm, C.M.; Chou, T.C. Temozolomide enhances herpes simplex virus thymidine kinase/ganciclovir therapy of malignant glioma. Cancer Gene Ther. 2001, 8, 662–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nestler, U.; Wakimoto, H.; Siller-Lopez, F.; Aguilar, L.K.; Chakravarti, A.; Muzikansky, A.; Stemmer-Rachamimov, A.; Chiocca, E.A.; Aguilar-Cordova, E.; Hochberg, F.H. The combination of adenoviral HSV TK gene therapy and radiation is effective in athymic mouse glioblastoma xenografts without increasing toxic side effects. J. Neurooncol. 2004, 67, 177–188. [Google Scholar] [CrossRef] [PubMed]
- De Melo, S.M.; Bittencourt, S.; Ferrazoli, E.G.; da Silva, C.S.; da Cunha, F.F.; da Silva, F.H.; Stilhano, R.S.; Denapoli, P.M.; Zanetti, B.F.; Martin, P.K.; et al. The anti-tumor effects of adipose tissue mesenchymal stem cell transduced with HSV-TK gene on U-87-driven brain tumor. PLoS ONE 2015, 10, e0128922. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Huang, J.; Jiang, X.; Zhang, J.; Yu, H.; Ye, J.; Zhang, D. A combination hepatoma-targeted therapy based on nanotechnology: pHRE-Egr1-HSV-TK/131i-antiAFPMcAb-GCV/MHF. Sci. Rep. 2016, 6, 33524. [Google Scholar] [CrossRef] [PubMed]
- Fillat, C.; Carrio, M.; Cascante, A.; Sangro, B. Suicide gene therapy mediated by the Herpes Simplex virus thymidine kinase gene/Ganciclovir system: Fifteen years of application. Curr. Gene Ther. 2003, 3, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Chendeb, M.; Schneider, R.; Davidson, I.; Fadloun, A. Selective elimination of long INterspersed element-1 expressing tumour cells by targeted expression of the HSV-TK suicide gene. Oncotarget 2017, 8, 38239–38250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, S.Z.; Wang, S.; Zeng, Y.T. Development of an HSV-tk transgenic mouse model for study of liver damage. FEBS J. 2005, 272, 2207–2215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, W.; Lu, H. A novel joint analysis framework improves identification of differentially expressed genes in cross disease transcriptomic analysis. BioData Min. 2018, 11, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iguchi, Y.; Ito, Y.M.; Kataoka, F.; Nomura, H.; Tanaka, H.; Chiyoda, T.; Hashimoto, S.; Nishimura, S.; Takano, M.; Yamagami, W.; et al. Simultaneous analysis of the gene expression profiles of cancer and stromal cells in endometrial cancer. Genes Chromosomes Cancer 2014, 53, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.L.; Chukhman, M.; Lu, Y.; Liu, C.; Liu, S.Y.; Lu, H. RNA-seq based transcription characterization of fusion breakpoints as a potential estimator for its oncogenic potential. BioMed Res. Int. 2017, 2017, 9829175. [Google Scholar] [CrossRef] [PubMed]
- Drilon, A.; Wang, L.; Arcila, M.E.; Balasubramanian, S.; Greenbowe, J.R.; Ross, J.S.; Stephens, P.; Lipson, D.; Miller, V.A.; Kris, M.G.; et al. Broad, hybrid capture-based next-generation sequencing identifies actionable genomic alterations in lung adenocarcinomas otherwise negative for such alterations by other genomic testing approaches. Clin. Cancer Res. 2015, 21, 3631–3639. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Wan, S.; Yang, Z.; Teschendorff, A.E.; Zou, Q. Tumor origin detection with tissue-specific miRNA and DNA methylation markers. Bioinformatics 2018, 34, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, N.; Knief, J.; Kacprowski, T.; Lazar-Karsten, P.; Keck, T.; Billmann, F.; Schmid, S.; Luley, K.; Lehnert, H.; Brabant, G.; et al. MicroRNA analysis of gastroenteropancreatic neuroendocrine tumors and metastases. Oncotarget 2018, 9, 28379–28390. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Wang, X.; Zeng, Y.; Zou, Q. Identification of DEP domain-containing proteins by a machine learning method and experimental analysis of their expression in human HCC tissues. Sci. Rep. 2016, 6, 39655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.J.; Wang, M.C.; Zhang, F.H.; Kong, X. An integrated analysis of genome-wide DNA methylation and gene expression data in hepatocellular carcinoma. FEBS Open Biol. 2018, 8, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Ding, N.; Rodriguez-Paton, A.; Zou, Q. Probability-based collaborative filtering model for predicting gene-disease associations. BMC Med. Genomics 2017, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Leng, J.Y.; Zhang, R.K.; Zhang, L.; Zhang, B.; Jiang, W.Y.; Tong, L. Functional analysis of gene expression profiling-based prediction in bladder cancer. Oncol. Lett. 2018, 15, 8417–8423. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, X.; Genchev, G.Z.; Lu, H. Multi-omics facilitated variable selection in Cox-regression model for cancer prognosis prediction. Methods 2017, 124, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez Bautista, R.; Ortega Gomez, A.; Hidalgo Miranda, A.; Zentella Dehesa, A.; Villarreal-Garza, C.; Avila-Moreno, F.; Arrieta, O. Long non-coding RNAs: Implications in targeted diagnoses, prognosis, and improved therapeutic strategies in human non- and triple-negative breast cancer. Clin. Epigenet. 2018, 10, 88. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Ma, Q.; Lu, Y.; Wang, J.; Hu, W.; Gong, Z.; Cai, L.; Huang, Y.; Huang, S.Z.; Zeng, F. PhiC31 integrase induces efficient site-specific recombination in the Capra hircus genome. DNA Cell Biol. 2014, 33, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villanueva, A.; Alsinet, C.; Yanger, K.; Hoshida, Y.; Zong, Y.; Toffanin, S.; Rodriguez-Carunchio, L.; Solé, M.; Thung, S.; Stanger, B.Z.; et al. Notch signaling is activated in human hepatocellular carcinoma and induces tumor formation in mice. Gastroenterology 2012, 143, 1660–1669.e1667. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Wada, T.; Suzuki, T.; Kubota, Y.; Inagaki, N. Singar1, a novel RUN domain-containing protein, suppresses formation of surplus axons for neuronal polarity. J. Biol. Chem. 2007, 282, 19884–19893. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Wang, J.; Tang, W.; Li, Y.; Peng, Y.; Zhang, H.; Liu, G.; Huang, X.; Zhao, J.; Li, A.; et al. Rufy3 promotes metastasis through epithelial-mesenchymal transition in colorectal cancer. Cancer Lett. 2017, 390, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, Q.; Song, Y.; Wang, X.; Guo, Q.; Zhang, J.; Li, J.; Han, Y.; Miao, Z.; Li, F. PAK1 regulates RUFY3-mediated gastric cancer cell migration and invasion. Cell Death Dis. 2015, 6, e1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros, S.A.; Tennant, R.W.; Cannon, R.E. Molecular structure and characterization of a novel murine ABC transporter, Abca13. Gene 2003, 307, 191–200. [Google Scholar] [CrossRef]
- Katz, S.F.; Lechel, A.; Obenauf, A.C.; Begus-Nahrmann, Y.; Kraus, J.M.; Hoffmann, E.M.; Duda, J.; Eshraghi, P.; Hartmann, D.; Liss, B.; et al. Disruption of Trp53 in livers of mice induces formation of carcinomas with bilineal differentiation. Gastroenterology 2012, 142, 1229–1239.e1223. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, K.; Van Bockstaele, D.R.; Berneman, Z.N. The cell cycle: A review of regulation, deregulation and therapeutic targets in cancer. Cell Prolif. 2003, 36, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Coulouarn, C.; Factor, V.M.; Conner, E.A.; Thorgeirsson, S.S. Genomic modeling of tumor onset and progression in a mouse model of aggressive human liver cancer. Carcinogenesis 2011, 32, 1434–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, R.E.; Lo, D.; Pinkert, C.A.; Widera, G.; Flavell, R.A.; Palmiter, R.D.; Brinster, R.L. Infertility in male transgenic mice: Disruption of sperm development by HSV-tk expression in postmeiotic germ cells. Biol. Reprod. 1990, 43, 684–693. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Verma, I.M. NF-kappaB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Mayo, M.W.; Baldwin, A.S. The transcription factor NF-kappaB: control of oncogenesis and cancer therapy resistance. Biochim. Biophys. Acta 2000, 1470, M55–M62. [Google Scholar] [PubMed]
- Lin, A.; Karin, M. NF-kappaB in cancer: A marked target. Semin. Cancer Biol. 2003, 13, 107–114. [Google Scholar] [CrossRef]
- Lavon, I.; Goldberg, I.; Amit, S.; Landsman, L.; Jung, S.; Tsuberi, B.Z.; Barshack, I.; Kopolovic, J.; Galun, E.; Bujard, H.; et al. High susceptibility to bacterial infection, but no liver dysfunction, in mice compromised for hepatocyte NF-kappaB activation. Nat. Med. 2000, 6, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Lavon, I.; Pikarsky, E.; Gutkovich, E.; Goldberg, I.; Bar, J.; Oren, M.; Ben-Neriah, Y. Nuclear factor-kappaB protects the liver against genotoxic stress and functions independently of p53. Cancer Res. 2003, 63, 25–30. [Google Scholar] [PubMed]
- Jeyasuria, P.; Subedi, K.; Suresh, A.; Condon, J.C. Elevated levels of uterine anti-apoptotic signaling may activate NFKB and potentially confer resistance to caspase 3-mediated apoptotic cell death during pregnancy in mice. Biol. Reprod. 2011, 85, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Pikarsky, E.; Porat, R.M.; Stein, I.; Abramovitch, R.; Amit, S.; Kasem, S.; Gutkovich-Pyest, E.; Urieli-Shoval, S.; Galun, E.; Ben-Neriah, Y. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature 2004, 431, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Petrescu, A.D.; Grant, S.; Frampton, G.; Kain, J.; Hadidi, K.; Williams, E.; McMillin, M.; DeMorrow, S. Glucocorticoids cause gender-dependent reversal of hepatic fibrosis in the MDR2-knockout mouse model. Int. J. Mol. Sci. 2017, 18, 2389. [Google Scholar] [CrossRef] [PubMed]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harb Perspect. Biol. 2015, 7, a008656. [Google Scholar] [CrossRef] [PubMed]
- Conner, E.A.; Lemmer, E.R.; Sanchez, A.; Factor, V.M.; Thorgeirsson, S.S. E2F1 blocks and c-Myc accelerates hepatic ploidy in transgenic mouse models. Biochem. Biophys. Res. Commun. 2003, 302, 114–120. [Google Scholar] [CrossRef]
- Su, C.; Zhang, C.; Tecle, A.; Fu, X.; He, J.; Song, J.; Zhang, W.; Sun, X.; Ren, Y.; Silvennoinen, O.; et al. Tudor staphylococcal nuclease (Tudor-SN), a novel regulator facilitating G1/S phase transition, acting as a co-activator of E2F-1 in cell cycle regulation. J. Biol. Chem. 2015, 290, 7208–7220. [Google Scholar] [CrossRef] [PubMed]
- Phesse, T.J.; Myant, K.B.; Cole, A.M.; Ridgway, R.A.; Pearson, H.; Muncan, V.; van den Brink, G.R.; Vousden, K.H.; Sears, R.; Vassilev, L.T.; et al. Endogenous c-Myc is essential for p53-induced apoptosis in response to DNA damage in vivo. Cell Death Differ. 2014, 21, 956–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, K.; Fry, E.A.; Frazier, D.P. Transcription factors that interact with p53 and Mdm2. Int. J. Cancer 2016, 138, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- Harbour, J.W.; Dean, D.C. The Rb/E2F pathway: Expanding roles and emerging paradigms. Genes Dev. 2000, 14, 2393–2409. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.F.; Chang, H.Y.; Tsai, H.W.; Hsieh, W.C.; Kuo, Y.H.; Su, I.J.; Lin, Y.J. Liver regeneration accelerates hepatitis B virus-related tumorigenesis of hepatocellular carcinoma. Mol. Oncol. 2018, 2, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Condiotti, R.; Goldenberg, D.; Giladi, H.; Schnitzer-Perlman, T.; Waddington, S.N.; Buckley, S.M.; Heim, D.; Cheung, W.; Themis, M.; Coutelle, C.; et al. Transduction of fetal mice with a feline lentiviral vector induces liver tumors which exhibit an E2F activation signature. Mol. Ther. 2014, 22, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Chen, S.; Yin, H.; Tammela, T.; Papagiannakopoulos, T.; Joshi, N.S.; Cai, W.; Yang, G.; Bronson, R.; Crowley, D.G.; et al. CRISPR-mediated direct mutation of cancer genes in the mouse liver. Nature 2014, 514, 380–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.S.; Salmena, L.; Pandolfi, P.P. The functions and regulation of the PTEN tumour suppressor. Nat. Rev. Mol. Cell Biol. 2012, 13, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Matter, M.S.; Marquardt, J.U.; Andersen, J.B.; Quintavalle, C.; Korokhov, N.; Stauffer, J.K.; Kaji, K.; Decaens, T.; Quagliata, L.; Elloumi, F.; et al. Oncogenic driver genes and the inflammatory microenvironment dictate liver tumor phenotype. Hepatology 2016, 63, 1888–1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Sequence (5′ to 3′) | ||
|---|---|---|
| Forward | Reverse | |
| PCR for HSV-tk | GTATACCGGTATGCCCACGCTACTGCGG | GATGGCGGTGAAGATGAGG |
| first-round iPCR | GAGCATGAGGTGACACTACT | CTAGGCTGTGAGGATACA AG |
| second-round iPCR | ATACCATCATTCCGGACGTG | CTAGGCTGTGAGGATACAAG |
| Lines of HSV-tk Mice | Chromosomal Location | Intergenic Region | Intron of Gene |
|---|---|---|---|
| TK5 | 5E1-E2 | No | Rufy3 |
| TK11 | 11A1-A2 | No | Abca13 |
| TK4 | 8C3-C4 | Yes | No |
| HSV-tk Lines | Incidence of Tumor Development (%) | ||||
|---|---|---|---|---|---|
| 3 Months | 6 Months | 9 Months | 12 Months | >13 Months | |
| TK5 | 0 (0/7) | 100 (4/4) | 100 (10/10) | 100 (11/11) | 100 (2/2) |
| TK11 | 0 (0/4) | 0 (0/3) | 50 (3/6) | 100 (6/6) | 100 (6/6) |
| TK4 | 0 (0/5) | 0 (0/4) | 25 (1/4) | 40 (6/15) | 46 (16/35) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, Z.; Ma, Q.; Wang, X.; Cai, Q.; Gong, X.; Genchev, G.Z.; Lu, H.; Zeng, F. A Herpes Simplex Virus Thymidine Kinase-Induced Mouse Model of Hepatocellular Carcinoma Associated with Up-Regulated Immune-Inflammatory-Related Signals. Genes 2018, 9, 380. https://doi.org/10.3390/genes9080380
Gong Z, Ma Q, Wang X, Cai Q, Gong X, Genchev GZ, Lu H, Zeng F. A Herpes Simplex Virus Thymidine Kinase-Induced Mouse Model of Hepatocellular Carcinoma Associated with Up-Regulated Immune-Inflammatory-Related Signals. Genes. 2018; 9(8):380. https://doi.org/10.3390/genes9080380
Chicago/Turabian StyleGong, Zhijuan, Qingwen Ma, Xujun Wang, Qin Cai, Xiuli Gong, Georgi Z. Genchev, Hui Lu, and Fanyi Zeng. 2018. "A Herpes Simplex Virus Thymidine Kinase-Induced Mouse Model of Hepatocellular Carcinoma Associated with Up-Regulated Immune-Inflammatory-Related Signals" Genes 9, no. 8: 380. https://doi.org/10.3390/genes9080380