Genome Wide Identification, Evolutionary, and Expression Analysis of VQ Genes from Two Pyrus Species


School of Life Sciences, Anhui Agricultural University, Hefei 230036, China
*
Author to whom correspondence should be addressed.
Genes 2018, 9(4), 224; https://doi.org/10.3390/genes9040224
Submission received: 14 March 2018
/
Revised: 18 April 2018
/
Accepted: 18 April 2018
/
Published: 23 April 2018
(This article belongs to the Section Plant Genetics and Genomics)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The VQ motif-containing gene, a member of the plant-specific genes, is involved in the plant developmental process and various stress responses. The VQ motif-containing gene family has been studied in several plants, such as rice (Oryza sativa), maize (Zea mays), and Arabidopsis (Arabidopsis thaliana). However, no systematic study has been performed in Pyrus species, which have important economic value. In our study, we identified 41 and 28 VQ motif-containing genes in Pyrus bretschneideri and Pyrus communis, respectively. Phylogenetic trees were calculated using A. thaliana and O. sativa VQ motif-containing genes as a template, allowing us to categorize these genes into nine subfamilies. Thirty-two and eight paralogous of VQ motif-containing genes were found in P. bretschneideri and P. communis, respectively, showing that the VQ motif-containing genes had a more remarkable expansion in P. bretschneideri than in P. communis. A total of 31 orthologous pairs were identified from the P. bretschneideri and P. communis VQ motif-containing genes. Additionally, among the paralogs, we found that these duplication gene pairs probably derived from segmental duplication/whole-genome duplication (WGD) events in the genomes of P. bretschneideri and P. communis, respectively. The gene expression profiles in both P. bretschneideri and P. communis fruits suggested functional redundancy for some orthologous gene pairs derived from a common ancestry, and sub-functionalization or neo-functionalization for some of them. Our study provided the first systematic evolutionary analysis of the VQ motif-containing genes in Pyrus, and highlighted the diversification and duplication of VQ motif-containing genes in both P. bretschneideri and P. communis.
1. Introduction
The VQ motif-containing gene, a member of the plant-specific genes, is involved in the plant developmental process and various stress responses. The VQ motif-containing gene encoding domain contains a conserved VQ motif, which possesses 50–60 amino acids with a high conserved FXXhVQXhTG (h refers to hydrophobic amino acids and x refers to any amino acids) [1]. The VQ domain plays a critical role in the biological function of VQ motif-containing proteins. For example, the mutation of AtVQ14 (i.e., changes from IVQQ to EDLE) in the VQ domain leads to the smaller seed of the mutant strain, while mutations in other locations do not have this phenotype [2]. It is reported that the first VQ motif-containing protein was identified from Arabidopsis thaliana using the yeast two-hybrid method [1]. Subsequently, the VQ motif-containing genes have been identified in many plants. A total of 39, 18, 74, 18, 61, 34, and 23 members have been identified in rice, moss, soybean, grape, maize, A. thaliana, and Fragaria vesca, respectively [3,4,5,6,7,8]. Evolutionary analysis shows that members of the VQ motif-containing gene family can be clustered into 9 groups, named from I-X, and further reveals the molecular mechanisms of these genes [3,9]. Among them, some closely related VQ motif-containing genes, which belong to the same subgroup, may be functionally redundant, such as AtVQ1 and AtVQ10 (group VIII), AtVQ24 and AtVQ15 (group X), and AtVQ12 and AtVQ29 (group IV) [3].
The published articles have reported that VQ motif-containing proteins participate not only in plant stress response to drought; salt; and pathogenic bacteria, fungi, and oomycetes, but also in the regulation of various life processes of plants, such as the development of seeds, hypocotyls, flowers, leaves, and fruits [3,4,5,6,7,9,10,11,12]. Remarkably, some VQ motif-containing proteins may form complexes with WRKYs through physical interactions [13], such as AtVQ23 and AtVQ16 that appear to act as activators of AtWRKY33 in plant defenses [13]. AtVQ15 (AtCaMBP25) is a calmodulin-binding protein, and its overexpression transgenic plants are highly sensitive to osmotic stress and NaCl in seed germination and seedling growth [14]. Transgenic plants overexpressing AtVQ21 (MKS1) showed significantly increased resistance to Pseudomonas syringae [15], while showing decreased resistance to Botrytis cinerea [16,17]. The AtVQ29 plays an important role in the light morphogenesis of seedlings in A. thaliana [10]. For example, the expression level of AtVQ29 in stems is significantly higher than that in roots, rosette leaves, flowers, and pods [10]. Transgenic plants overexpressing AtVQ29 led to significantly longer hypocotyl lengths than the wild type under far-red or low light conditions, while the A. thaliana mutant plants’ hypocotyl lengths were significantly shorter than that of wild type [10]. The mutant plants of A. thaliana AtVQ8 gene showed yellowish-green leaves and growth retardation throughout their whole growth and development period, and the growth of overexpression plants of VQ17, VQ18, and VQ22 was also severely inhibited [3].
Recently, some published papers have confirmed that there are differences in the evolutionary pattern of gene families between adjacent species [8,18]. Zhong et al. [8] explored the evolution and functional of VQ motif-containing genes in six species of Fragaria, and suggested that the expansion pattern of this gene family was different among species [8]. Both Fragaria and Pyrus belong to Rosaceae family. Pyrus, which produces fruits that have great commercial value, is also one of the most important economical crops worldwide. Compared to the largely well characterized VQ motif-containing genes in A. thaliana [9,19], VQ motif-containing genes have not been functionally characterized in Pyrus (i.e., Pyrus bretschneideri and Pyrus communis). It is well-known that P. bretschneideri and P. communis have undergone at least two rounds of whole-genome duplication (WGD) events [20,21], which play an essential role in the expansion of the genes. Previous studies have reported that orthologous genes may contribute to some of the evolutionary innovations and may develop non-function, sub-function, or neo-function after evolving from a common ancestry in plants [22,23]. Thus, it is of interest to characterize orthologous VQ motif-containing genes between P. bretschneideri and P. communis. Our study identified 41 and 28 VQ motif-containing genes in P. bretschneideri and P. communis, respectively, and comprehensively characterized their evolutionary outcomes and gene duplication types based on the microcollinearity, gene structure, and expression patterns analyses.
2. Materials and Methods
2.1. Identification and Classification of P. bretschneideri and P. communis VQ Motif-Containing Genes
Predicted proteomes from P. bretschneideri (http://gigadb.org/site) and P. communis (https://www.rosaceae.org) were selected and downloaded. The VQ motif-containing genes of A. thaliana were obtained from The Arabidopsis Information Resource (TAIR) database (http://arabidopsis.org). According to the Hidden Markov Model (HMM) profile of VQ domains (PF05678) and A. thaliana VQ motif-containing proteins, we performed basic local alignment search tool (BLAST) (E-value = 1 × 10−3) searches using HMMER3 software [24]. These putative VQ motif-containing sequences, which were confirmed to contain the VQ domains by the National Center for Biotechnology Information (NCBI) Conserved Domain database, Pfam database, and SMART database [25,26,27], were used for further analysis. The VQ motif-containing sequences from Pyrus and A. thaliana were aligned using MAFFT (Multiple Alignment using Fast Fourier Transform) with default parameters [28]. According to the alignment results, we constructed a phylogenetic tree using the neighbor-joining (NJ) method, as implemented in MEGA 5 software [29]. The classification information of VQ motif-containing genes was extracted from the previously published articles [9].
2.2. Chromosome Locations and Microcollinearity Analysis of Pyrus VQ Motif-Containing Genes
In order to understand the chromosome location information, we obtained the GFF (General Feature Format) files of P. bretschneideri and P. communis from GigaDB (http://gigadb.org/site) and GDR database (https://www.rosaceae.org), respectively. Protein BLAST (BLASTP) search was carried out against P. bretschneideri and P. communis genomes, with an E-value below 1 × 105. According to the results of BLASTP and GFF, we determined the genome-wide duplication (GWD)/segmental duplication using MCScanX software [30]. The Ka (non-synonymous substitution)/Ks (synonymous substitution), Ka, and Ks values were estimated using DnaSP 5.0 software [31]. The microcollinearity visualization of the duplicated VQ motif-containing genes was performed by our Perl script (Supplementary Data S1). The Circos software was used to map the chromosome location among the P. bretschneideri and P. communis genomes [32].
2.3. Intron–Exon Structure and Domain Search of P. bretschneideri and P. communis VQ Motif-Containing Genes
According to the extracted information from the GFF files, we diagrammed the domain and intron–exon structures of P. bretschneideri and P. communis using R software and our Perl script (Supplementary Data S1). The domain information of P. bretschneideri and P. communis VQ motif-containing genes was obtained from the Pfam database [27]. The multiple EM for motif elicitation (MEME) online tool was used to identify the conserved VQ motifs with the following parameters: optimum motif length = 6–200 residues; maximum number of motifs = 20; the number of repetition = any [33].
2.4. Expression Data Analysis
To determine the expression of PbVQ genes in P. bretschneideri, RNA-Seq data were obtained from the NCBI database. The accession numbers and sample details or treatments for these data are presented in the availability of data and materials section. The clean reads, which were removed low-quality base-calls (Q < 20) using FASTX-toolkit (http://hannonlab.cshl.edu/fastx_toolkit), were mapped to the reference genome using TopHat2 software with default parameters, and were assembled using Cufflinks software [34,35]. The expression profiles were visualized using R software with our R script (Supplementary Data S1).
2.5. Expression Correlation of Homologous VQ Motif-Containing Genes between P. bretschneideri and P. communis
Expression profiles of homologous VQ motif-containing genes were manually gathered from RNA-Seq data. Using Pearson’s correlation coefficient, we calculated the similarity between the expression patterns of the homologous gene pair. According to previous studies [36,37], significant values were suggested to confirm the degree of expression diversity. Generally, r < 0.3, 0.3 < r < 0.5, and r > 0.5 indicates divergence, ongoing divergent, and non-divergence, respectively [36,37].
2.6. Plant Material, RNA Extraction, and Real-Time Polymerase Chain Reaction Analysis
The ‘Dangshansuli’ (P. bretschneideri) and ‘Starkrimson’ (P. communis) were picked from 30-year-old pear trees grown on a pear orchard (Dangshan, Anhui, China). In the present study, we selected ten healthy and robust trees, which were managed in a consistent manner. When the trees were at the bud stage (i.e., April 2016), we selected short branches bearing buds of similar developmental stages from the middle-crown area on the south side of each tree, and then marked them. In each short branch, only two fruits were kept. We collected the fruits from seven developmental stages, including fruit_stage1 (15 days after full blooming (DAB)), fruit_stage2 (30 DAB), fruit_stage3 (55 DAB), fruit_stage4 (85 DAB), fruit_stage5 (115 DAB), fruit_stage6 (mature stage), and fruit_stage7 (fruit senescence stage), respectively. After removing the peels of the fruits, we extracted total RNA of flesh for the synthesis of the first-strand complementary DNA (cDNA) using Trizol reagent (Invitrogen, Carlsbad, CA, USA). The quantitative real-time PCR (qRT-PCR) reactions were executed with 40 cycles in the Bio-rad CFX96 Real-Time PCR Detection system (BioRad, Hercules, CA, USA) using the TransStart Tip Green qPCR SuperMix (TransGen Biotech, Beijing, China). According to a previous workflow, the normalization of each reaction threshold cycle (Ct) value was determined by pear tubulin gene [38,39]. Beacon Designer v7.9 was used to design specific primers of the qRT-PCR analysis [18,39], which are presented in Table S1. All expression data were obtained from three biological repeats. The SPSS v19.0 software was used to perform statistical significance with the t test [18,39].
2.7 Availability of Data and Materials
Stop-growth pollen tube, Accession: SRX1356346; Pollen tube, Accession: SRX1356343; Hydrated pollen grains, Accession: SRX1356152; Mature pollen grains of pear, Accession: SRX1356151; Pear fruit developmental stage 1 (15DAB), Accession: SRX1595645; Pear fruit developmental stage 2 (30DAB), Accession: SRX1595646; Pear fruit developmental stage 3 (55DAB), Accession: SRX1595647; Pear fruit developmental stage 4 (85 DAB), Accession: SRX1595648; Pear fruit developmental stage 5 (115 DAB), Accession: SRX1595650; Pear fruit developmental stage 6 (mature stage), Accession: SRX1595651; Pear fruit developmental stage 7 (fruit senescence stage), Accession: SRX1595652; Pear pericarp-russet, Accession: SRX707274; Pear pericarp-green, Accession: SRX316593; Pear leaf with salt treatment, Accession: SRX525736; Pear leaf with inoculated black spot (Alternarlia alternate) 2, Accession: SRX864865; Pear leaf with inoculated distilled water 1, Accession: SRX864860; Pear leaf with inoculated distilled water 2, Accession: SRX864862; Pear leaf with inoculated black spot (Alternarlia alternate) 3, Accession: SRX864861; Pear fruit with Gibberellin treatment, Accession: SRX532394.
3. Results
3.1. Identification and Classification of P. bretschneideri and P. communis VQ Motif-Containing Genes
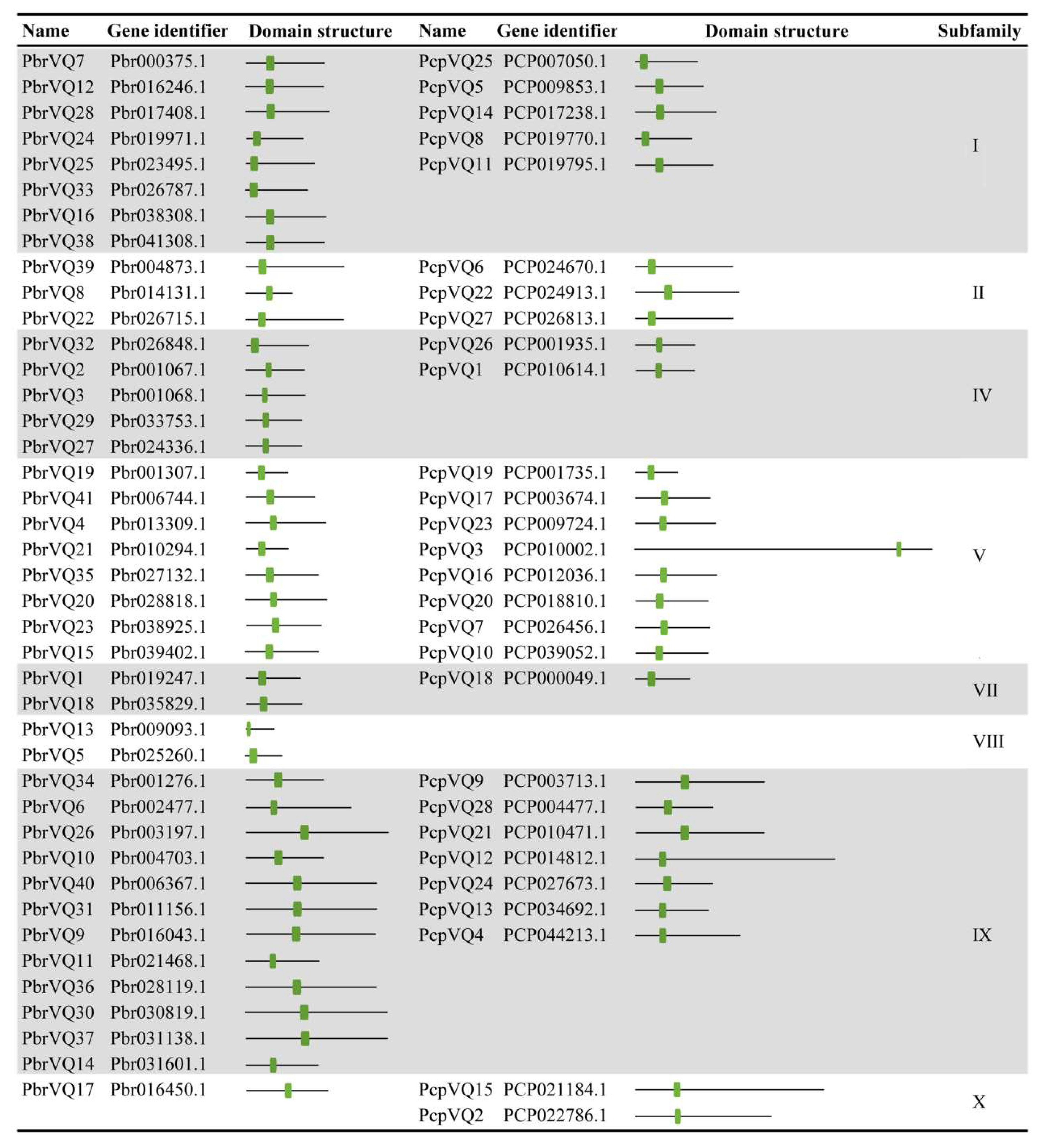
To identify the VQ motif-containing gene family members encoded by P. bretschneideri and P. communis genomes, we searched for these genomes using HMMER 3.0 software and the Pfam database with the VQ domain (PF05678). We identified 41 and 28 non-redundant VQ motif-containing genes for the following analysis in P. bretschneideri and P. communis, respectively. Detailed information (including gene identifier, domain structure, and subfamily) concerning VQ motif-containing genes is listed in Figure 1.
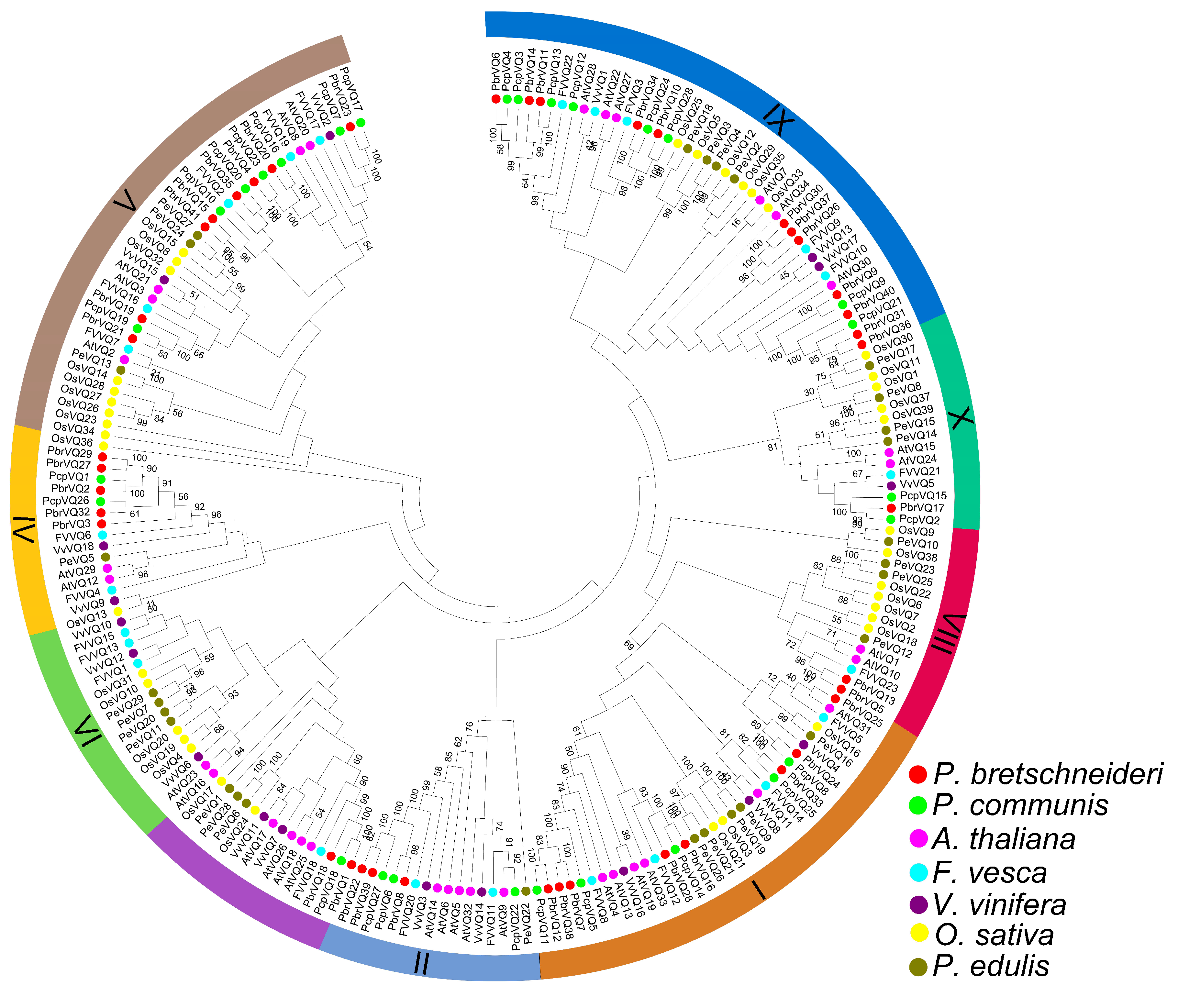
The classification of VQ motif-containing genes subfamilies has been carried out by Pascal Pecher et al. [13]. To further understand the phylogenetic relationships of VQ motif-containing gene family members in P. bretschneideri and P. communis, a NJ phylogenetic tree was constructed. As shown in Figure 2, the VQ proteins from monocotyledonous (Oryza sativa and Phyllostachys edulis) and dicotyledonous (P. bretschneideri, P. communis, Vitis vinifera, A. thaliana, and F. vesca) could be divided into nine subfamilies (I-X), based on the previous studies [9]. Remarkably, when compared with the other subfamilies, we noticed that the V and IX subfamily member sizes were significantly larger. These results were consistent with previous articles on A. thaliana, O. sativa, and Z. mays VQ motif-containing genes [4,7,19]. However, we also found that some subfamilies only contained VQ motif-containing genes from one or several species, indicating that they might have undergone dramatic changes after their origination during the long evolutionary period. For example, subfamily II contained A. thaliana, P. edulis, F. vesca, P. communis, P. bretschneideri, and V. vinifera VQ motif-containing genes, while subfamily VIII only contained P. edulis, F. vesca, A. thaliana, O. sativa, and P. bretschneideri VQ motif-containing genes. Interestingly, VQ motif-containing genes from P. bretschneideri and P. communis showed a higher similarity with each other, based on the evolutionary relationships. This was not surprising because P. bretschneideri and P. communis belong to Pyrus species.
3.2. Chromosome Locations and Duplication Events of P. bretschneideri and P. communis VQ Motif-Containing Genes
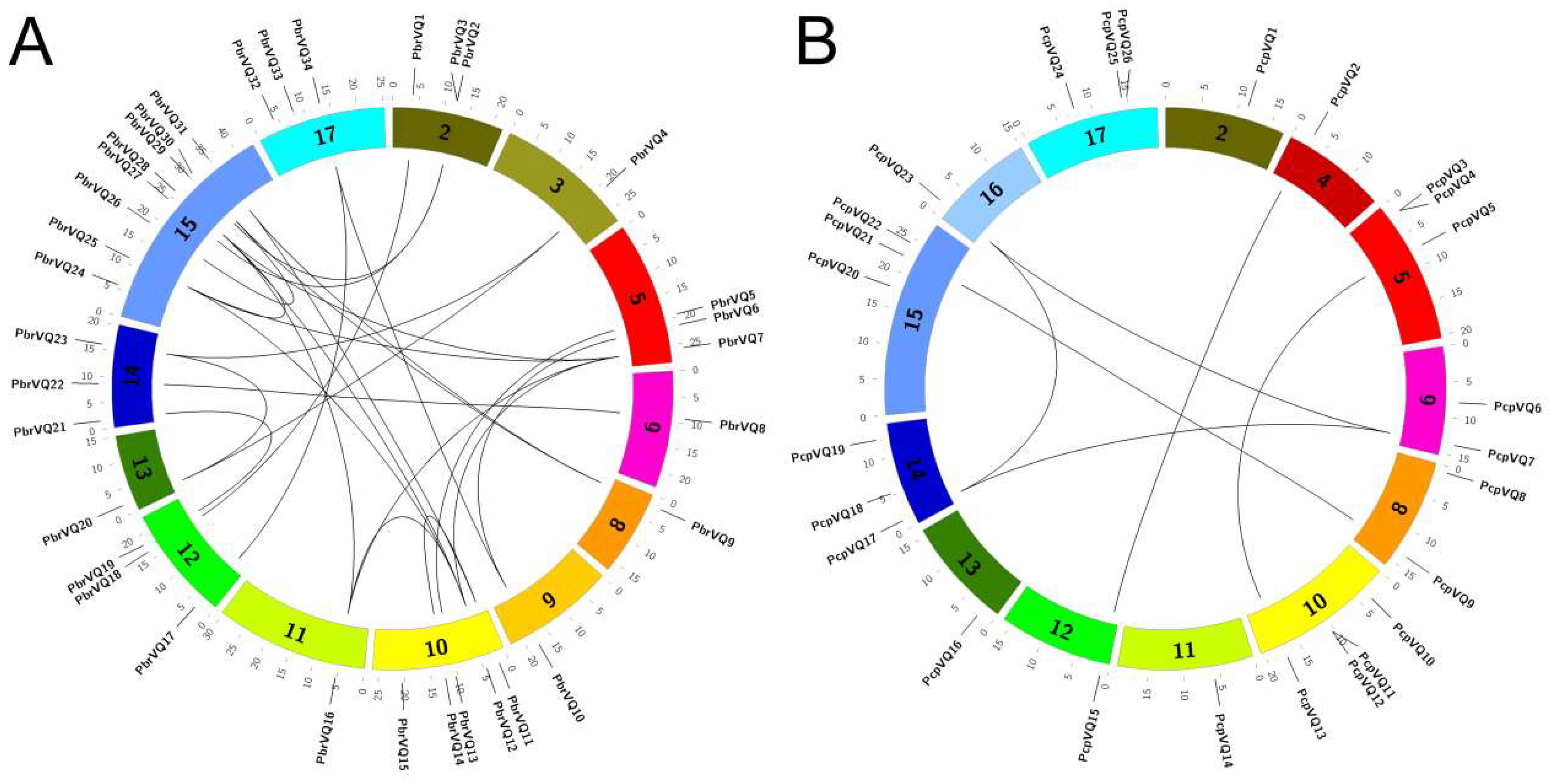
The Pyrus genome contained 17 chromosomes. To further understand the distribution of VQ motif-containing genes in the Pyrus genome, we mapped P. bretschneideri and P. communis VQ motif-containing genes to chromosome positions. In P. bretschneideri, the distribution of VQ motif-containing genes was among the chromosomes 2, 3, 5, 6, 8, 9, 10, 11, 12, 13, 14, 15, and 17, especially in chromosome 15 (Figure 3A). In P. communis, the distribution of VQ motif-containing genes was among the chromosomes 2, 4, 5, 6, 8, 10, 11, 12, 13, 14, 15, 16, and 17, especially in chromosome 10 (Figure 3B). As we know, the pear genome has undergone two WGD events. It has been reported that a recent WGD was estimated at 30–45 MYA (Millions of years ago) (Ks ∼ 0.15–0.3), while an ancient WGD was estimated at ∼140 MYA (Ks ∼ 1.5–1.8). To gain insight into the relationship between VQ motif-containing gene expansions and WGD events, we identified 32 and 8 collinearity events among P. bretschneideri and P. communis, respectively (Figure 3). In P. bretschneideri, we found that most Ks ratios of these collinearity events varied from 0.15–0.3 (Table S2). However, only three Ks ratios ranging from 0.15–0.3 were detected in P. communis. Remarkably, the ancient WGD events were not found in the VQ motif-containing genes of these two species, because their Ks values were not distributed in the 1.5–1.8 range (Table S2). The results indicated that a recent GWD might have contributed to the expansions of P. bretschneideri VQ motif-containing gene members. At the same time, these results further explain why the number of VQ motif-containing genes in the P. bretschneideri genome was far more than in the P. communis genome.
We identified 32 and 8 segmentally arranged P. bretschneideri and P. communis VQ motif-containing genes, respectively, indicating that they might be the results of segmental duplication events (Figure 3 and Table S2). In P. bretschneideri, the high frequency of segmental duplication occurred between chromosomes 10 and 15, which possessed three segmental duplications. Interestingly, no collinearity events were arranged in tandem duplication events for both P. bretschneideri and P. communis, indicating that segmental duplication events might have played major roles in the amplification of VQ motif-containing genes in these two Pyrus species (Figure 3 and Table S2).
3.3. Syntenic Blocks Analysis of P. bretschneideri and P. communis VQ Motif-Containing Genes
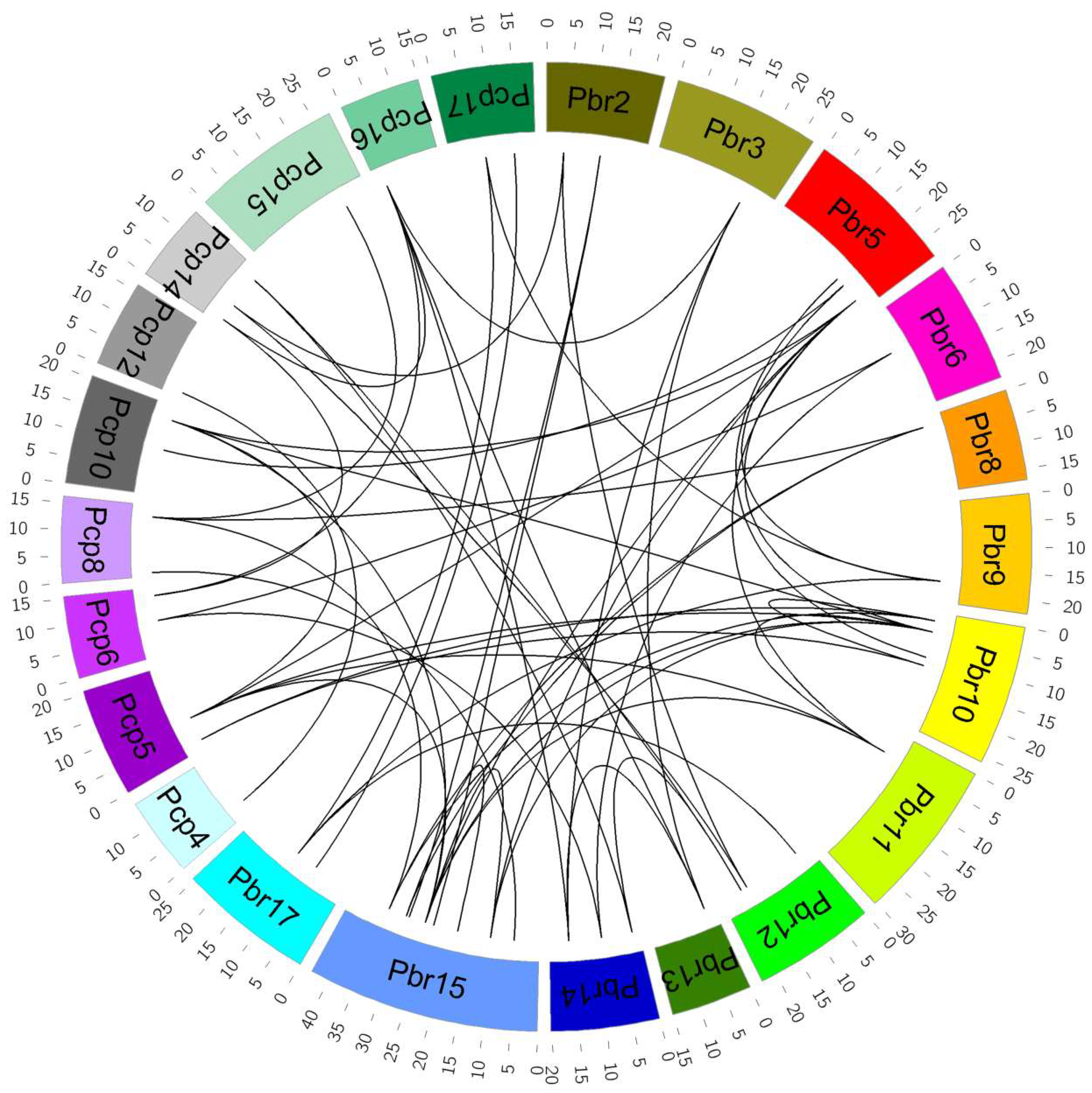
It is well-known that synteny relations between species help us to understand the conserved biological functions and genome evolution among species. To gain insight into the evolution of P. bretschneideri and P. communis VQ motif-containing genes, the syntenic blocks were searched in the chromosomes using MCScanX software. Among them, 31 orthologous pairs were identified within these pairwise syntenic blocks (Figure 4 and Table S3). Additionally, we also observed a reciprocal homology in syntenic blocks between the P. bretschneideri and P. communis chromosomes. The syntenic blocks were distributed in most chromosomes of the Pyrus genomes. For P. bretschneideri, both chromosomes 15 and 10 presented the highest number of matches with P. communis VQ motif-containing genes, each of them displaying linkage with four genes in the corresponding chromosomes. The P. communis chromosome 5 matched the highest number of P. bretschneideri VQ motif-containing genes, exhibiting homology with five sequences. Curiously, the genes distributing in the same chromosome in one species matched syntenic blocks in different chromosomes from the other species, as in chromosome 15 from P. bretschneideri and in chromosomes 5, 8, and 10 from P. communis (Figure 4).
In order to further clarify the evolutionary history of VQ motif-containing genes, we specifically analyzed the paralogous and orthologous VQ motif-containing gene pairs between P. bretschneideri and P. communis. In the present study, we identified 31 orthologous gene pairs between P. bretschneideri and P. communis (Figure 4 and Table S3). A total of 32 and 8 paralogous gene pairs were found within the P. bretschneideri and P. communis genomes, respectively. Additionally, we found that some VQ motif-containing genes were not involved in any syntenic blocks, indicating that they evolved from independent duplication events. Interestingly, we found that two or more VQ motif-containing genes from P. communis or P. bretschneideri matched one P. bretschneideri or P. communis VQ motif-containing gene, implying that these genes might have contributed to the expansion of the VQ motif-containing gene family during the course of evolution. For example, PbrVQ7, PbrVQ12, and PbrVQ28 were orthologous to PcpVQ5, and PcpVQ24 and PcpVQ24 were orthologous to PbrVQ34.
3.4. Gene Structural and Conserved Motifs Analysis of P. bretschneideri and P. communis VQ Motif-Containing Genes
Gene structural diversity provides the primary resources for the evolution of multi-gene families, as found in previously published articles [38,40,41]. The exon–intron distribution and organization maps of each VQ motif-containing gene were generated to understand the structural diversity of VQ motif-containing genes. As shown in Figure S1, the different numbers of exons (ranging from 2 to 13) were found among 69 VQ motif-containing genes. In P. bretschneideri, we found that the exon–intron number of only one homologous pair (PbrVQ10 and PbrVQ6) had changed within the 32 duplicated gene pairs. In P. communis, the exon–intron number of two homologous pairs (PcpVQ15 and PcpVQ2, PcpVQ5 and PcpVQ11) had changed. For example, PbrVQ10 and PcpVQ2 lost four introns when compared with PbrVQ6 and PcpVQ15. At the same time, we also investigated the structural diversity of orthologous genes between P. bretschneideri and P. communis. Among 31 orthologous gene pairs, we identified that the exon–intron numbers of six orthologous gene pairs (PbrVQ6 and PcpVQ12, PbrVQ7 and PcpVQ5, PbrVQ12 and PcpVQ5, PbrVQ28 and PcpVQ5, PbrVQ16 and PcpVQ5, and PbrVQ38 and PcpVQ5) had changed. Subsequent gene structure analysis suggested that 97.6% (40/41 genes) of PbrVQ genes and 78.5% (22/28 genes) of PcpVQ genes did not contain introns, with PbrVQ6 (3 introns), PcpVQ3 (12 introns), PcpVQ5 (1 intron), PcpVQ10 (2 introns), PcpVQ12 (11 introns), PcpVQ15 (8 introns), and PcpVQ2 (4 introns) being the exceptions (Figure S1). These data were consistent with the previous studies that reported 54 VQ motif-containing genes in Z. mays (88.5%) [7], 37 genes in O. sativa (92.5%) [4], 25 genes in P. edulis (86.2%) [42], and 30 genes in A. thaliana (88.2%) [3] without introns. On the contrary, 28% (7/25) of moss VQ motif-containing genes did not possess introns. These comparative analyses (higher plants P. bretschneideri, P. communis, P. edulis, A. thaliana, Z. mays, and O. sativa, and lower plants, for example, moss) indicated that most VQ motif-containing genes had lost introns during the long evolutionary period. PcpVQ3, PcpVQ12, and PcpVQ15—which include 12, 11, and 8 introns, respectively—are obviously special cases, and are similar to VQ36 in Brassica rapa, which contains 14 introns [11].
We also analyzed the conserved motif using the MEME software (Figure S1). In the present study, we identified twenty different motifs (Figure S1 and Table S4). All of the VQ motif-containing genes from P. bretschneideri and P. communis clearly possessed the same motif 1, which was encoded by the VQ domain. The motif analysis suggested that VQ motif-containing proteins belonging to the same subfamily possessed basically the same type of motif, which supported the phylogenetic analysis. Subsequently, we also obtained additional information from the analysis of paralogous gene pairs and orthologous gene pairs between P. bretschneideri and P. communis. Among these paralogous/orthologous gene pairs, the type of motif in each gene pair was basically consistent. The presence of the same conserved domain implied that the function of the homologous gene pairs was similar at the protein level.
3.5. Expression Pattern Analysis of P. bretschneideri VQ Motif-Containing Genes by Transcriptome Data
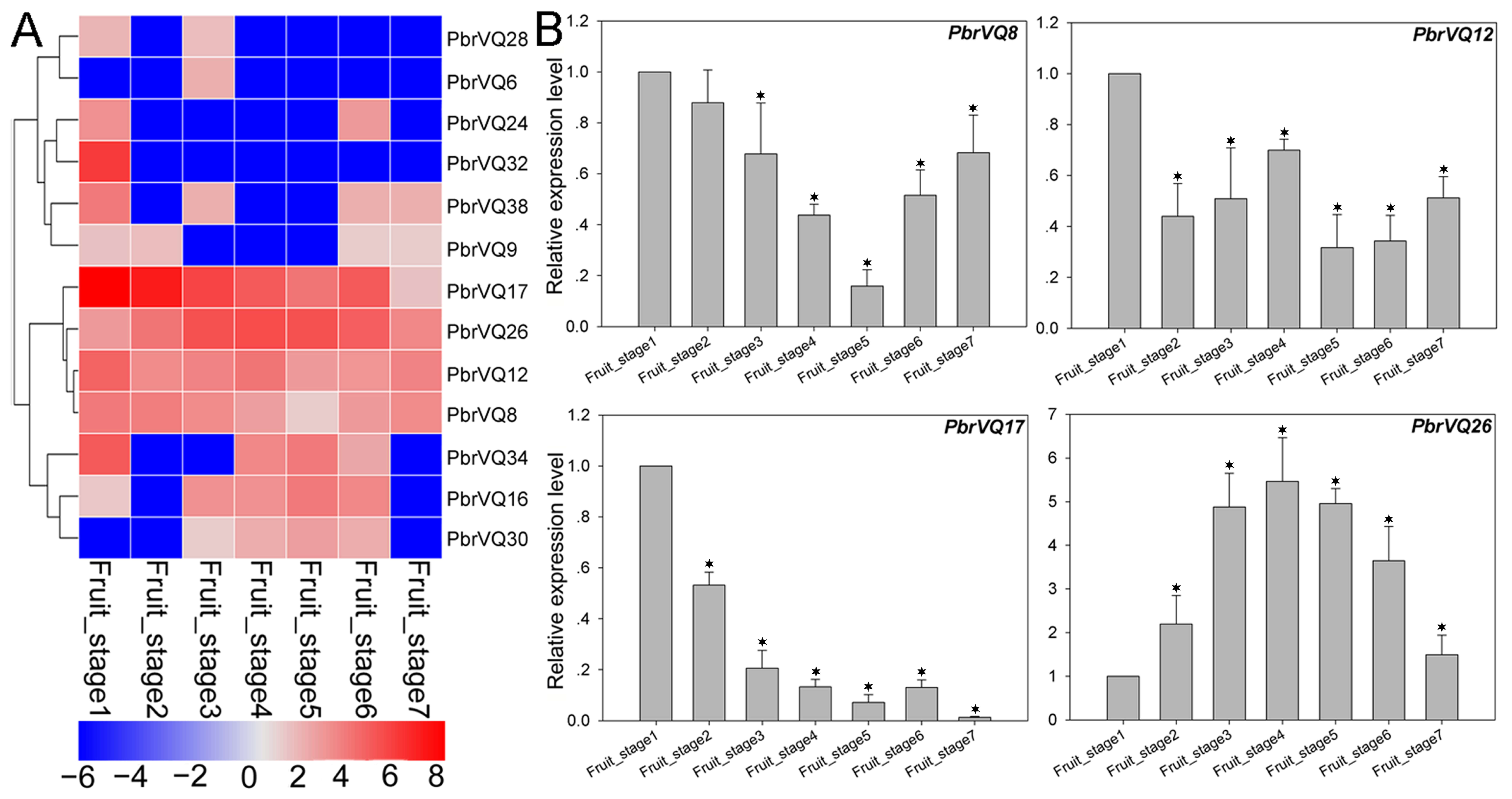
To further understand the expression patterns of VQ motif-containing genes, we analyzed publicly available RNA-seq data from the NCBI SRA (Sequence Read Archive) database for a total of eight experiments, including pollen tube development, fruit development, abiotic stress, biotic stress, and developmental biology. The sample details and treatments for these data are presented in the availability of data and materials section. In the present study, we examined the expression of P. bretschneideri VQ motif-containing genes in four growth stages of pollen (i.e., MP (mature pollen grain), HP (hydrated pollen grain), PT (growing pollen tube), and SPT (stopped-growth pollen tube)) (Figure S2). We found that only three PbrVQ genes, including PbrVQ8, PbrVQ1, and PbrVQ23, were expressed in one or several periods. Among them, the PbrVQ23 gene was expressed in all periods, indicating that this gene plays an important role in pear pollen tube development. The expression patterns of PbrVQ genes were also investigated during pear fruit development. A total of 13/41 (31.7%) PbrVQ genes were relatively highly expressed in pear fruit. Among them, four genes (PbrVQ8, PbrVQ12, PbrVQ17, and PbrVQ26) were expressed in all pear fruit development stages (Figure 5 and Table S5). These genes were mainly distributed in subfamilies I, II, IX, and X. We also used the qRT-PCR experiment to confirm the expression patterns, as shown in Figure 5B. The expression trends of these genes were similar to those of RNA-seq data (Figure 5 and Table S5).
In addition, VQ motif-containing genes have been previously reported to be involved in biotic and abiotic stresses [4,6,11]. We found that the most of the PbrVQ genes (29 genes) presented differential expression when compared with the untreated control, while the remaining genes (12 genes) were not detectable. For example, after black spot (Alternaria alternata) treatment, PbrVQ8, 28, 23, 33, 9, 30, 2, 32, 13, 5, 11, 38, 3, 15, 24, 15, 10, 39, and 20 were up-regulated, and PbrVQ12, 16, 26, 34, 41, and 6 were down-regulated (Figure S2). It is well-known that GA (Gibberellic acid) has various regulation functions in plants, such as controlling fertilization time, simulating organ growth, and early seed development. In our study, the top highly expressed gene in the P. bretschneideri sample by GA treatment was PbrVQ9 (Figure S2). Our data suggested that these genes contained the potential roles in stress response.
3.6. Comparison of the Expression Patterns of P. bretschneideri VQ Motif-Containing Genes and Their Orthologous in P. communis during Fruit Development
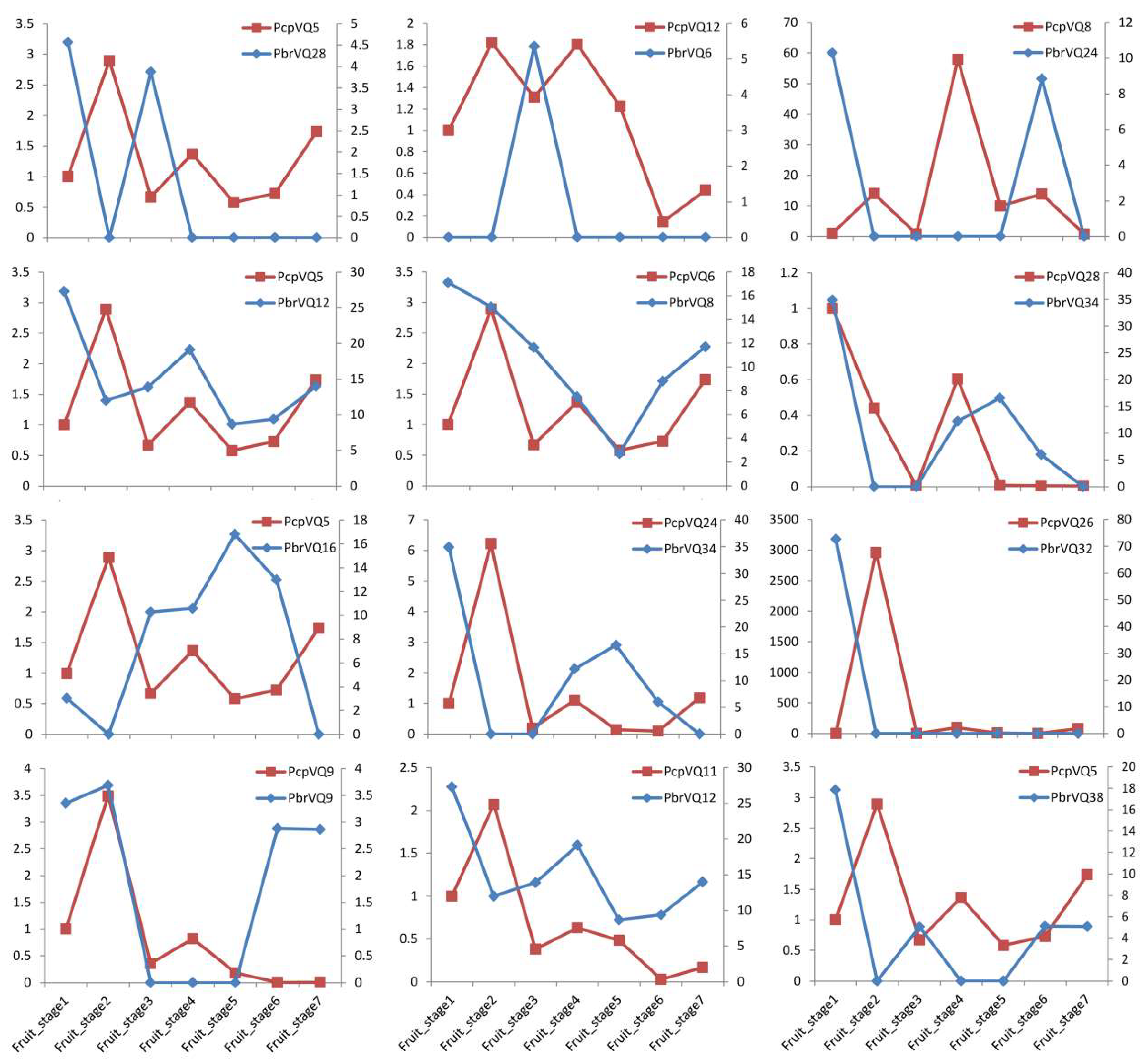
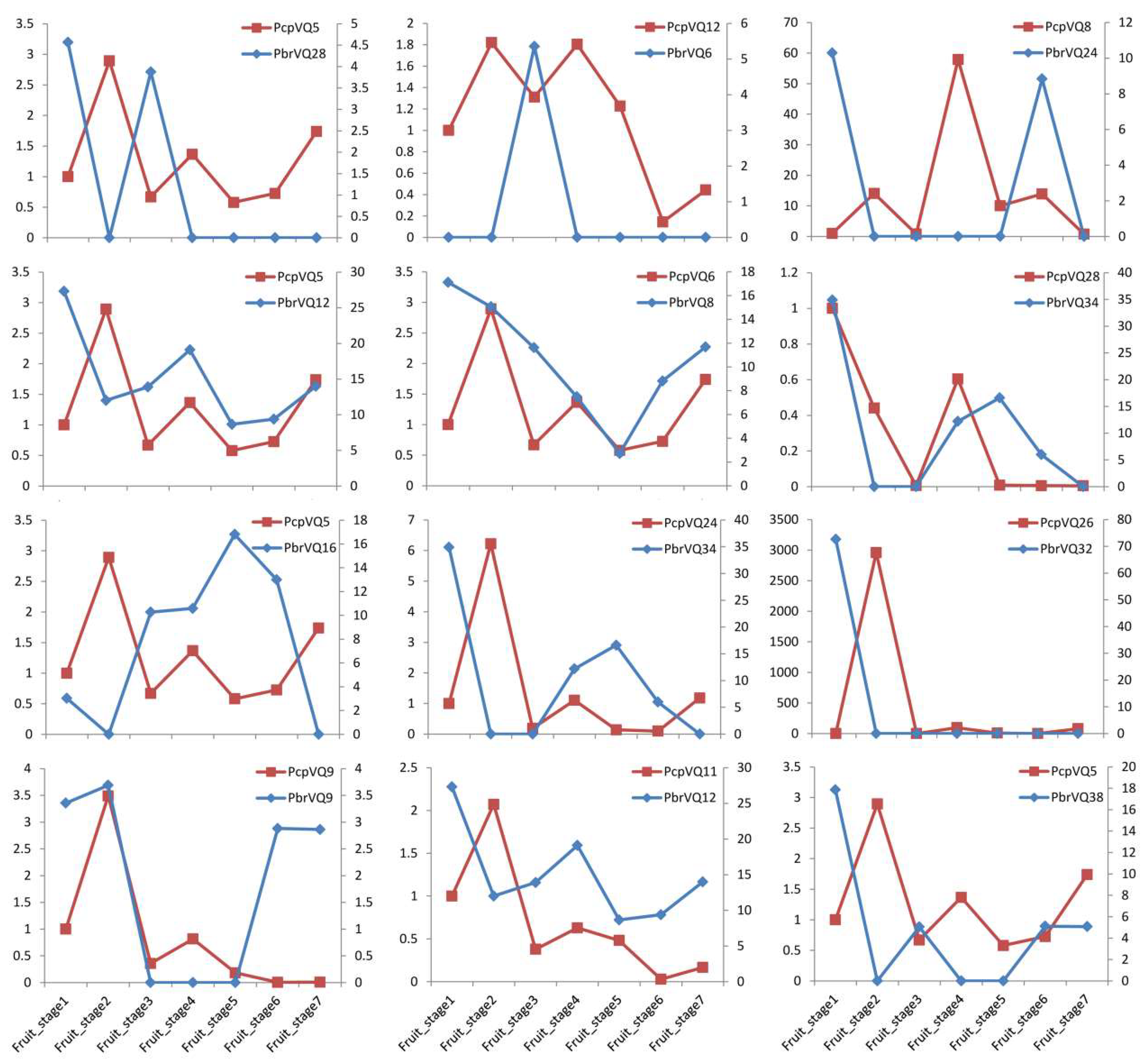
Pears are widely cultivated around the world, and the fruit is the focus of this study because of its economic value. As we know, the homologous gene pairs might possess similar expression patterns. In our study, in order to further understand the degree of expression diversity of VQ motif-containing genes between P. bretschneideri and P. communis, we estimated their expression correlations. In the present study, one orthologous gene pair (PbrVQ34–PcpVQ28) was identified to be non-divergent, two orthologous gene pairs (PbrVQ9–PcpVQ9 and PbrVQ8–PcpVQ6) were ongoing divergent, and the remaining orthologous gene pairs were divergent (Figure 6 and Table S4). These results indicated that most of the orthologous VQ motif-containing gene pairs from P. bretschneideri and P. communis have undergone functional divergence (Figure 6 and Table S6). These expression patterns differ from those previously reported between B. rapa and A. thaliana orthologous VQ motif-containing gene pairs, where it was found that most of the orthologous gene pairs presented similar expression profiles under hormone and abiotic treatment [11]. The reason for this divergence might have been that the culture conditions in the orchard of the tree of both pear species were different. Our study mainly explored the divergences of orthologous gene pairs in the developmental stages of pear fruit. At the same time, gene expression profiles in both P. bretschneideri and P. communis fruits suggested that functional redundancy derived from a common ancestry for some orthologous gene pairs, and from sub-functionalization or neo-functionalization for others.
4. Discussion
4.1. Phylogeny of P. bretschneideri and P. communis VQ Motif-Containing Gene Family
VQ motif-containing genes are involved in the regulation of various processes, such as plant growth and development, as well as the resistance to biotic and abiotic stresses. The VQ motif-containing gene family has been discovered in many species, such as O. sativa, Z. mays, A. thaliana, and others [3,4,5,6,7,9,11,42]. However, there is still lack of systematic analyses of VQ motif-containing genes in two Pyrus species (i.e., P. bretschneideri and P. communis). Subsequently, the genome-wide analyses of P. bretschneideri and P. communis VQ motif-containing genes and their regulation during pollen tube, fruit developmental processes, and/or stress responses were performed.
In the higher eukaryotes, intronless genes are very common in their genomes [43]. In our study, we found that most of the PbrVQ and PcpVQ genes are intronless, based on the gene structure analysis. Only one gene contained multiple introns in P. bretschneideri and five genes contained multiple introns in P. communis. Subsequent to the phylogenetic analysis of VQ motif-containing genes in P. bretschneideri, P. communis, A. thaliana, and O. sativa, VQ genes containing intron(s) were presented in several different subfamilies, indicating that these introns arose relatively independently and recently in P. bretschneideri, P. communis, A. thaliana, and O. sativa. The PbrVQ and PcpVQ genes identified in the present study, and those identified in other species, will facilitate further studies of plant intron evolution.
Some conserved motifs and intron–exon structures within the VQ motif-containing genes existed not only in higher plants (such as P. bretschneideri, P. communis, Arabidopsis, maize, and rice) but also in lower plants (such as moss), indicating their ancient origin in evolutionary history and their important roles in plant developmental processes. Indeed, our hypothesis was also supported by some research articles. For example, AtVQ14 mutation produces small seeds and reduces endosperm growth, indicating that this gene might be involved in seed development [44,45]. Throughout the entire life cycle, the recessive loss-of-function AtVQ8 mutant exhibits stunted-growth and pale-green phenotypes, indicating a predominant role in photosystem assembly or chloroplast development [3]. VQ motif-containing genes also mediate the responses to environmental stresses and pathogen infection in rice [4]; salicylic acid (SA), ethylene (ETH), powdery mildew infection, and drought in grapes [6]; low-nitrogen stress in soybeans; osmotic stresses and drought in maize [7]; and plant hormone treatments and different abiotic stresses in cabbage [11].
4.2. Expansion and Duplication of P. bretschneideri and P. communis VQ Motif-Containing Gene Family
The identification of duplicate genes that are retained after GWD may be challenging, as found in previously published articles [46]. Many rearrangements of the pear genome via fractionation and two GWD events can lead to loss of synteny [20,47], which causes the detection of gene pairs to be complicated. In our study, we did not distinguish between segmental duplications and GWD gene pairs, as this makes it difficult to separate these two duplication types. For pears, previous research indicated that dispersed duplication and GWD/segmental duplication were the main sources of duplicate genes [38,40,41,48]. The VQ motif-containing genes were identified in GWD/segmental duplication as a common evolutionary mechanism in P. bretschneideri and P. communis. GWD/segmental duplication events were the main driving force during the VQ motif-containing gene family expansion in pears. In the present study, 32 and 8 duplication VQ motif-containing gene pairs were identified as GWD/segmental duplication in P. bretschneideri and P. communis, respectively. However, previous researchers did not identify any duplication events in the VQ motif-containing genes of grapevine [6]. These results suggested that differences in the evolution of the VQ motif-containing gene family exist in different species. Additionally, we also explored the role of GWD events in the expansion of P. bretschneideri and P. communis VQ motif-containing gene family members. Our data suggested that the recent GWD events contributed to the expansions of P. bretschneideri VQ motif-containing genes.
4.3. Expression Analysis of P. bretschneideri VQ Motif-Containing Genes
Previous studies reported that many VQ motif-containing genes play roles in response to biotic and abiotic stresses [3,4,6,9,11,19]. In the present study, some PbrVQ genes were relatively highly expressed in P. bretschneideri samples that were treated with GA (Gibberellic acid), salt, and black spot (Alternaria alternata). For example, 17 PbrVQ genes were induced by black spot treatments. The PbrVQ17 was highly expressed in P. bretschneideri samples by GA treatment, which was consistent with B. rapa VQ genes in response to GA [11]. Additionally, increasingly more studies have shown that the transcription of VQ motif-containing genes is regulated by environmental and various endogenous signals, which is consistent with its different roles in plant growth and development. In A. thaliana, the AtVQ14 gene is mainly associated with seed development, such that its mutation produces small seeds [44,45]. Compared with wild type plants, over-expression of AtVQ22, AtVQ18, and AtVQ17 causes highly stunted growth of the transgenic plans [10]. The AtVQ8 plays an important role in photosystem assembly and chloroplast development, which is consistent with its location in plastid [3]. Recently, a study has shown that the heterologous expression of AtVQ21 in petunia and jonquil led to delayed flowering and dwarfed phenotypes in these species [49]. In our study, the expression levels of PbrVQ genes were detected during P. bretschneideri fruit development. These data indicated that the majority of the PbrVQ genes were differentially expressed during P. bretschneideri fruit development. Several PbrVQ genes were expressed in all periods of pear fruit development, indicating that these genes might be very important for the development of pear fruit. Previous studies have shown that AtVQ20 protein interacts with both AtWRKY2 and AtWRKY34 transcription factors to modulate pollen development and function [50]. Therefore, we detected the expression patterns of the PbrVQ genes during pear pollen development. The PbrVQ23, the homolog of AtVQ20, was highly expressed during pear pollen development when compared with its level in other examined tissues. Taken together, our study suggested that the VQ motif-containing genes were extensively involved in pollen tube and fruit growth and development.
5. Conclusions
In our study, a systematic study was performed on the genome-wide identification, phylogenetic relationship, characterization, and expression analysis of VQ domain-containing genes in both P. bretschneideri and P. communis. By combining a phylogenomics approach at the genome level with expression pattern analysis during fruit development, we have increased functional divergences and expansion patterns in the VQ motif-containing genes of P. bretschneideri and P. communis that evolved from the same common ancestry. Finally, this study illustrates the decisive contribution of GWD/segmental duplication events to the evolution of genes, and as a foundation for future exploration of the functional divergences of the VQ domain-containing genes in both P. bretschneideri and P. communis.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4425/9/4/224/s1, Data S1: Scripts used in Section 2.2, Section 2.3 and Section 2.4. Figure S1: Exon–intron structure and conserved motifs analysis of P. bretschneideri and P. communis VQ motif-containing genes. Figure S2: The heatmap of the expression of P. bretschneideri VQ motif-containing genes in different tissues or pollen tube development. PbrVQ gene expression levels during pollen germination and pollen tube growth, including MP (Mature pollen grains), HP (Hydrated pollen grains), PT (Pollen tube), SPT (Stop-growth pollen tube), and PbrVQ gene expression levels in different conditions, including pear leaf with inoculated distilled water (Leaf_CK), pear leaf with inoculated black spot (Alternarlia alternate) 2 (Leaf_T2), pear leaf with inoculated black spot (Alternarlia alternate) 3 (Leaf_T3), pear fruit with no any treatment (Fruit_CK), pear fruit with Gibberellin treatment (Fruit_GA), pear leaf with salt treatment (Leaf_Salt), pear fruit pericarp-russet (pericarp-russet), and pear fruit pericarp-green (pericarp-green). Table S1: Primers in this study. Table S2: Ka/Ks analysis of between paralogous VQ motif-containing gene pairs in Pyrus. Table S3: Homologous analyses of VQ motif-containing genes between P. bretschneideri and P. communis. Table S4: All MEME motif sequences in both P. bretschneideri and P. communis VQ motif-containing proteins. Table S5: The FPKM values of PbVQ genes in different tissues or treatments. Table S6: Divergence analysis of orthologous VQ motif-containing gene pairs among P. bretschneideri and P. communis.
Acknowledgments
We extend our thanks to the reviewers and editors for their careful reading and helpful comments on this manuscript. This study was supported by The National Natural Science Foundation of China (grant 31640068). The Funding bodies were not involved in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Author Contributions
Yu.C. and Yo.C. designed and performed the experiments; Yu.C. and D.M. analyzed the data; D.M., Q.J., Y.L., M.A., Yu.C. and Yo.C. contributed reagents/materials/analysis tools; Yu.C. wrote the paper. All authors reviewed and approved the final submission.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Morikawa, K.; Shiina, T.; Murakami, S.; Toyoshima, Y. Novel nuclear-encoded proteins interacting with a plastid sigma factor, Sig1, in Arabidopsis Thaliana. FEBS Lett. 2002, 514, 300–304. [Google Scholar] [CrossRef]
- Wang, A.; Garcia, D.; Zhang, H.; Feng, K.; Chaudhury, A.; Berger, F.; Peacock, W.J.; Dennis, E.S.; Luo, M. The VQ motif protein IKU1 regulates endosperm growth and seed size in Arabidopsis. Plant J. 2010, 63, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhou, Y.; Yang, Y.; Chi, Y.; Zhou, J.; Chen, J.; Wang, F.; Fan, B.; Shi, K.; Zhou, Y. Structural and functional analysis of VQ motif-containing proteins in Arabidopsis as interacting proteins of WRKY transcription factors. Plant Physiol. 2012, 159, 810–825. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Kwon, S.I.; Choi, C.Y.; Lee, H.; Ahn, I.; Park, S.R.; Bae, S.; Lee, S.; Hwang, D.J. Expression analysis of rice VQ genes in response to biotic and abiotic stresses. Gene 2013, 529, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, H.; Sun, G.; Jin, Y.; Qiu, L. Identification of active VQ motif-containing genes and the expression patterns under low nitrogen treatment in soybean. Gene 2014, 543, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.M.; Vannozzi, A.; Wang, G.G.; Zhong, Y.Y.; Corso, M.; Cavallini, E.; Cheng, Z.M. A comprehensive survey of the grapevine VQ gene family and its transcriptional correlation with WRKY proteins. Front. Plant Sci. 2015, 6, 417. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Zhao, H.; Zhang, X.; Lei, L.; Lai, J. Genome-wide identification of VQ motif-containing proteins and their expression profiles under abiotic stresses in maize. Front. Plant Sci. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Guo, C.; Chu, J.; Liu, H.; Cheng, Z.-M. Microevolution of the VQ gene family in six species of Fragaria. Genome 2017, 61, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Lin, R. The VQ motif-containing protein family of plant-specific transcriptional regulators. Plant Physiol. 2015, 169, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jing, Y.; Li, J.; Xu, G.; Lin, R. Arabidopsis VQ MOTIF-CONTAINING PROTEIN29 represses seedling deetiolation by interacting with PHYTOCHROME-INTERACTING FACTOR1. Plant Physiol. 2014, 164, 2068–2080. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Wang, F.; Li, J.; Ding, Q.; Zhang, Y.; Li, H.; Zhang, J.; Gao, J. Genome-wide identification and analysis of the VQ motif-containing protein family in Chinese Cabbage (Brassica rapa L. ssp. Pekinensis). Int. J. Mol. Sci. 2015, 16, 28683–28704. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, Y.; Pan, J.; Yu, D. Arabidopsis VQ motif-containing proteins VQ12 and VQ29 negatively modulate basal defense against Botrytis cinerea. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Weyhe, M.; Eschenlippold, L.; Pecher, P.; Scheel, D.; Lee, J. Ménage à trois: The complex relationships between mitogen-activated protein kinases, WRKY transcription factors, and VQ-motif-containing proteins. Plant Signal. Behav. 2014, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Perruc, E.; Charpenteau, M.; Ramirez, B.C.; Jauneau, A.; Galaud, J.; Ranjeva, R.; Ranty, B. A novel calmodulin-binding protein functions as a negative regulator of osmotic stress tolerance in Arabidopsis Thaliana seedlings. Plant J. 2004, 38, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Andreasson, E.; Jenkins, T.; Brodersen, P.; Thorgrimsen, S.; Petersen, N.H.T.; Zhu, S.; Qiu, J.; Micheelsen, P.O.; Rocher, A.; Petersen, M.; et al. The MAP kinase substrate MKS1 is a regulator of plant defense responses. EMBO J. 2005, 24, 2579–2589. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.; Qiu, J.; Lutje, J.; Fiil, B.K.; Hansen, S.; Mundy, J.; Petersen, M. Arabidopsis MKS1 is involved in basal immunity and requires an intact N-terminal domain for proper function. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.; Brodersen, P.; Naested, H.; Andreasson, E.; Lindhart, U.; Johansen, B.; Nielsen, H.B.; Lacy, M.; Austin, M.J.; Parker, J.E.; et al. Arabidopsis MAP kinase 4 negatively regulates systemic acquired resistance. Cell 2000, 103, 1111–1120. [Google Scholar] [CrossRef]
- Cao, Y.; Han, Y.; Meng, D.; Li, D.; Jin, Q.; Lin, Y.; Cai, Y. Genome-wide analysis suggests high level of microsynteny and purifying selection affect the evolution of EIN3/EIL family in Rosaceae. PeerJ 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Pecher, P.; Eschenlippold, L.; Herklotz, S.; Kuhle, K.; Naumann, K.; Bethke, G.; Uhrig, J.; Weyhe, M.; Scheel, D.; Lee, J. The Arabidopsis Thaliana mitogen-activated protein kinases MPK3 and MPK6 target a subclass of ‘VQ-motif’-containing proteins to regulate immune responses. New Phytol. 2014, 203, 592–606. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, Z.; Shi, Z.; Zhang, S.; Ming, R.; Zhu, S.; Khan, M.A.; Tao, S.; Korban, S.S.; Wang, H.; et al. The genome of the pear (Pyrus bretschneideri rehd.). Genome Res. 2013, 23, 396–408. [Google Scholar] [CrossRef] [PubMed]
- Chagné, D.; Crowhurst, R.N.; Pindo, M.; Thrimawithana, A.; Deng, C.; Ireland, H.; Fiers, M.; Dzierzon, H.; Cestaro, A.; Fontana, P.; et al. The draft genome sequence of European pear (Pyrus communis L.‘Bartlett’). PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.Z.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.I.; Yau, C.K.; Looseley, M.; Meyers, B.C. Effects of gene expression on molecular evolution in Arabidopsis Thaliana and Arabidopsis Lyrata. Mol. Biol. Evol. 2004, 21, 1719–1726. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Finn, R.D.; Eddy, S.R.; Bateman, A.; Punta, M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 2013, 41. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2012, 40, D302–D305. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’S conserved domain database. Nucleic Acids Res. 2014, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed]
- Punta, M.; Coggill, P.C.; Eberhardt, R.Y.; Mistry, J.; Tate, J.; Boursnell, C.; Pang, N.; Forslund, K.; Ceric, G.; Clements, J.; et al. The Pfam protein families database. Nucleic Acids Res. 2011, 40, D290–D301. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME suite. Nucleic Acids Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with Tophat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed]
- Blanc, G.; Wolfe, K.H. Widespread paleopolyploidy in model plant species inferred from age distributions of duplicate genes. Plant Cell 2004, 16, 1667–1678. [Google Scholar] [CrossRef] [PubMed]
- Yim, W.C.; Lee, B.-M.; Jang, C.S. Expression diversity and evolutionary dynamics of rice duplicate genes. Mol. Genet. Genom. 2009, 281, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Han, Y.; Li, D.; Lin, Y.; Cai, Y. MYB transcription factors in Chinese Pear (Pyrus bretschneideri Rehd.): Genome-wide identification, classification, and expression profiling during fruit development. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Han, Y.; Meng, D.; Li, D.; Jiao, C.; Jin, Q.; Lin, Y.; Cai, Y. B-BOX genes: Genome-wide identification, evolution and their contribution to pollen growth in pear (Pyrus bretschneideri Rehd.). BMC Plant Biol. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Wines, D.R.; Brady, J.M.; Southard, E.M.; MacDonald, R.J. Evolution of the rat Kallikrein gene family: Gene conversion leads to functional diversity. J. Mol. Evol. 1991, 32, 476–492. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Ding, T.; Su, B.; Jiang, H. Genome-wide identification, characterization and expression analysis of the Chalcone Synthase family in maize. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, H.; Zhu, D.; Gao, Y.; Yan, H.; Xiang, Y. Genome-wide analysis of VQ motif-containing proteins in Moso Bamboo (Phyllostachys edulis). Planta 2017, 246, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Louhichi, A.; Fourati, A.; Rebaï, A. IGD: A resource for intronless genes in the human genome. Gene 2011, 488, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Saingery, V.; Chambrier, P.; Mayer, U.; Jürgens, G.; Berger, F. Arabidopsis haiku mutants reveal new controls of seed size by endosperm. Plant Physiol. 2003, 131, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Dennis, E.S.; Berger, F.; Peacock, W.J.; Chaudhury, A. MINISEED3 (MINI3), a WRKY family gene, and HAIKU2 (IKU2), a leucine-rich repeat (LRR) kinase gene, are regulators of seed size in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 17531–17536. [Google Scholar] [CrossRef] [PubMed]
- Van de Peer, Y. Computational approaches to unveiling ancient genome duplications. Nat. Rev. Genet. 2004, 5, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, J.A.; Maere, S.; Van de Peer, Y. Plants with double genomes might have had a better chance to survive the Cretaceous–Tertiary extinction event. Proc. Natl. Acad. Sci. USA 2009, 106, 5737–5742. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Li, L.; Yin, H.; Liu, X.; Wang, D.; Wu, J.; Wu, J.; Zhang, S. Modes of gene duplication and gene family expansion and evolution in Chinese white pear. In Proceedings of the Plant and Animal Genome XXIII Conference, San Diego, CA, USA, 10–14 January 2015. [Google Scholar]
- Gargul, J.M.; Mibus, H.; Serek, M. Manipulation of MKS1 gene expression Affects Kalanchoë blossfeldiana and Petunia hybrida phenotypes. Plant Biotechnol. J. 2015, 13, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Lei, R.; Li, X.; Ma, Z.; Lv, Y.; Hu, Y.; Yu, D. Arabidopsis WRKY2 and WRKY34 transcription factors interact with VQ20 protein to modulate pollen development and function. Plant J. 2017, 91, 962–976. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Summary of Pyrus bretschneideri and Pyrus communis VQ motif-containing genes. The classification information of VQ motif-containing genes was extracted from previously published articles [9]. The VQ domain is indicated in green.
Figure 1.
Summary of Pyrus bretschneideri and Pyrus communis VQ motif-containing genes. The classification information of VQ motif-containing genes was extracted from previously published articles [9]. The VQ domain is indicated in green.
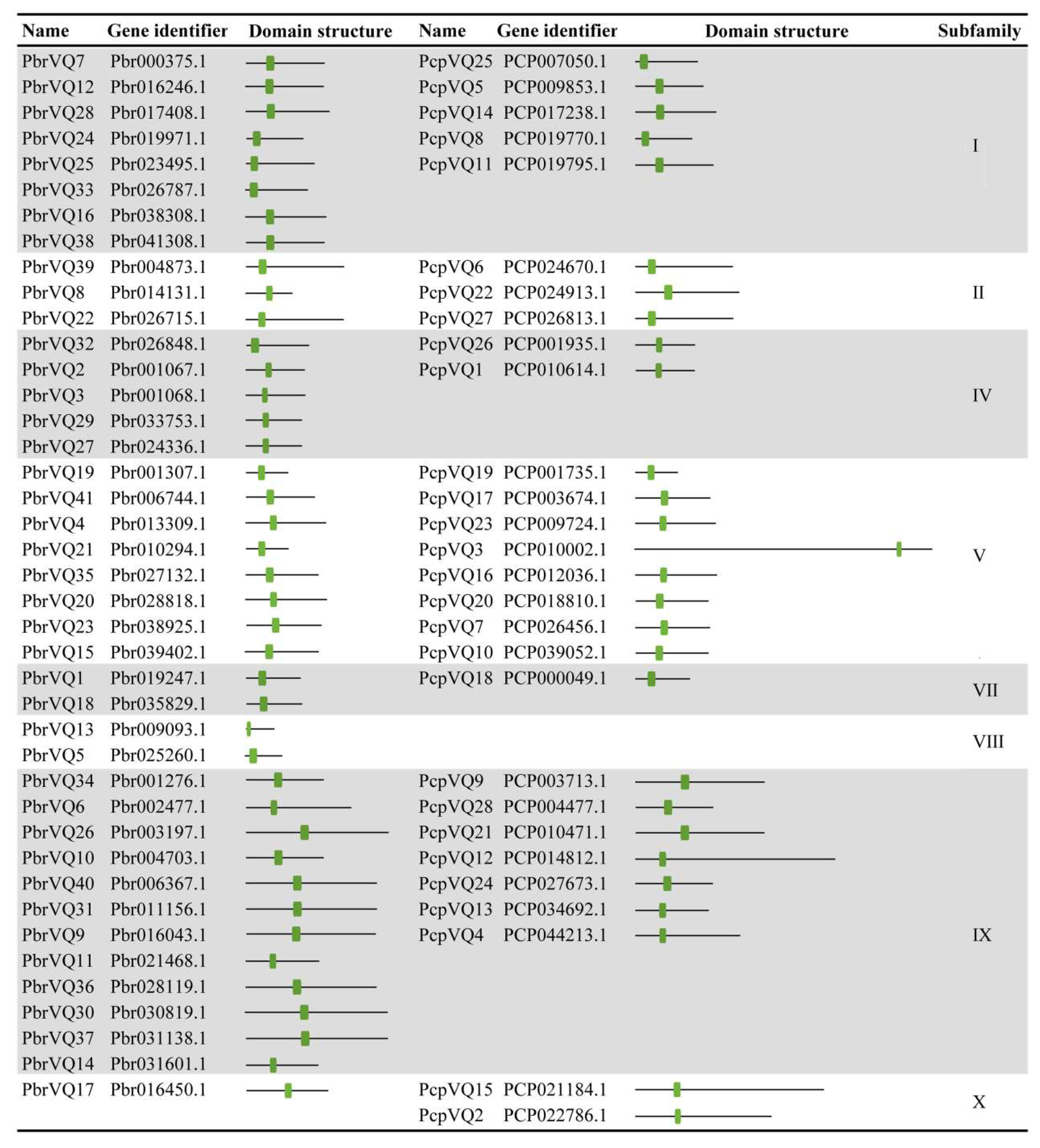
Figure 2.
Classification and phylogenetic relationships of P. bretschneideri and P. communis VQ motif-containing genes. The neighbor-joining (NJ) tree was constructed by the full-length amino acid sequences using MEGA 5 software. According to the previously published articles [9], we divided these sequences into nine different subfamilies (I-X). Proteins from P. bretschneideri, P. communis, Arabidopsis thaliana, Fragaria vesca, Vitis vinifera, Oryza sativa, and Phyllostachys edulis are denoted by red, green, pink, blue, purple, yellow, and brown circles, respectively.
Figure 2.
Classification and phylogenetic relationships of P. bretschneideri and P. communis VQ motif-containing genes. The neighbor-joining (NJ) tree was constructed by the full-length amino acid sequences using MEGA 5 software. According to the previously published articles [9], we divided these sequences into nine different subfamilies (I-X). Proteins from P. bretschneideri, P. communis, Arabidopsis thaliana, Fragaria vesca, Vitis vinifera, Oryza sativa, and Phyllostachys edulis are denoted by red, green, pink, blue, purple, yellow, and brown circles, respectively.
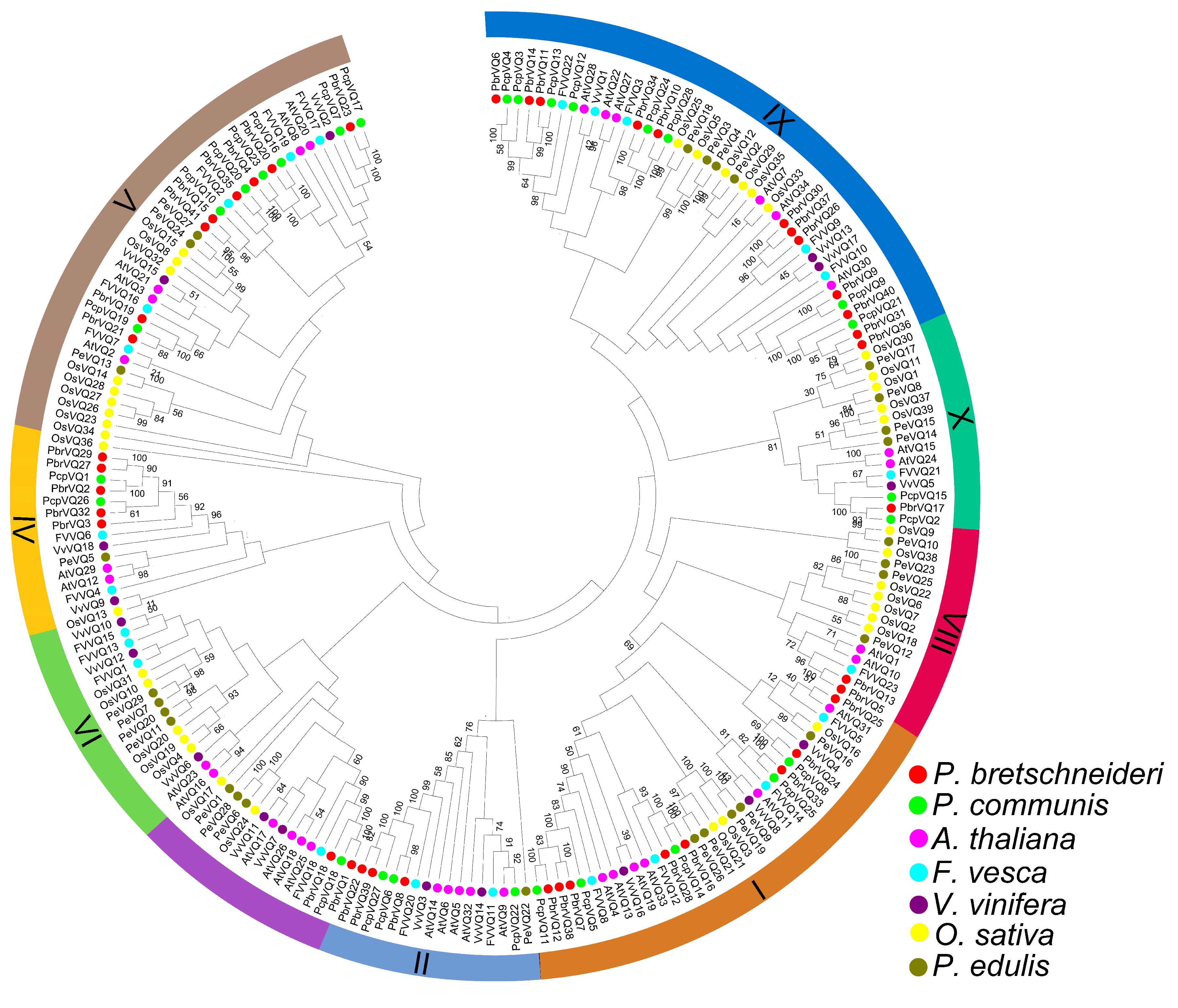
Figure 3.
Chromosome location and duplication events analysis in both P. bretschneideri (A) and P. communis (B) VQ motif-containing genes. The outermost scale represents the megabases (Mb). The location of each VQ motif-containing genes is marked with a grey line using Circos software. The duplication gene pairs were linked by black lines. Remarkably, because PbrVQ35, PbrVQ36, PbrVQ37, PbrVQ38, PbrVQ39, PbrVQ40, PbrVQ41, PcpVQ27, and PcpVQ28 were located on the scaffold, the location of these genes is not shown in Figure 3.
Figure 3.
Chromosome location and duplication events analysis in both P. bretschneideri (A) and P. communis (B) VQ motif-containing genes. The outermost scale represents the megabases (Mb). The location of each VQ motif-containing genes is marked with a grey line using Circos software. The duplication gene pairs were linked by black lines. Remarkably, because PbrVQ35, PbrVQ36, PbrVQ37, PbrVQ38, PbrVQ39, PbrVQ40, PbrVQ41, PcpVQ27, and PcpVQ28 were located on the scaffold, the location of these genes is not shown in Figure 3.

Figure 4.
Microsynteny of VQ motif-containing genes across P. bretschneideri and P. communis. The outermost scale represents the megabases (Mb). The P. bretschneideri and P. communis chromosomes are represented by different color boxes and are labeled Pbr and Pcp, respectively. The syntenic relationships are represented by black lines. Remarkably, because PbrVQ35, PbrVQ36, PbrVQ37, PbrVQ38, PbrVQ39, PbrVQ40, PbrVQ41, PcpVQ27, and PcpVQ28 were located on the scaffold, the related orthologous gene pairs are not shown in Figure 4.
Figure 4.
Microsynteny of VQ motif-containing genes across P. bretschneideri and P. communis. The outermost scale represents the megabases (Mb). The P. bretschneideri and P. communis chromosomes are represented by different color boxes and are labeled Pbr and Pcp, respectively. The syntenic relationships are represented by black lines. Remarkably, because PbrVQ35, PbrVQ36, PbrVQ37, PbrVQ38, PbrVQ39, PbrVQ40, PbrVQ41, PcpVQ27, and PcpVQ28 were located on the scaffold, the related orthologous gene pairs are not shown in Figure 4.
Figure 5.
Expression profiles of P. bretschneideri VQ motif-containing genes during seven fruit developmental stages, including fruit_stage1 (15 days after full blooming (DAB)), fruit_stage2 (30 DAB), fruit_stage3 (55 DAB), fruit_stage4 (85 DAB), fruit_stage5 (115 DAB), fruit_stage6 (mature stage), and fruit_stage7 (fruit senescence stage). (A) Heat map showing expression levels of 13 P. bretschneideri VQ motif-containing genes in seven stages of pear fruit development. As shown in the bar at the left bottom of Figure 5, gene transcript abundance is represent by different colors in the map. The FPKM (Fragments Per Kilobase of exon model per Million mapped fragments) values of PbVQ genes are presented in Table S5. (B) Gene expression of PbrVQ8, PbrVQ12, PbrVQ17, and PbrVQ26 was analyzed by real-time polymerase chain reaction (qRT-PCR). The standard error bar indicates three biological replicates. The asterisk indicates the significant difference (* p < 0.05) between treatment and control (fruit_stage1).
Figure 5.
Expression profiles of P. bretschneideri VQ motif-containing genes during seven fruit developmental stages, including fruit_stage1 (15 days after full blooming (DAB)), fruit_stage2 (30 DAB), fruit_stage3 (55 DAB), fruit_stage4 (85 DAB), fruit_stage5 (115 DAB), fruit_stage6 (mature stage), and fruit_stage7 (fruit senescence stage). (A) Heat map showing expression levels of 13 P. bretschneideri VQ motif-containing genes in seven stages of pear fruit development. As shown in the bar at the left bottom of Figure 5, gene transcript abundance is represent by different colors in the map. The FPKM (Fragments Per Kilobase of exon model per Million mapped fragments) values of PbVQ genes are presented in Table S5. (B) Gene expression of PbrVQ8, PbrVQ12, PbrVQ17, and PbrVQ26 was analyzed by real-time polymerase chain reaction (qRT-PCR). The standard error bar indicates three biological replicates. The asterisk indicates the significant difference (* p < 0.05) between treatment and control (fruit_stage1).
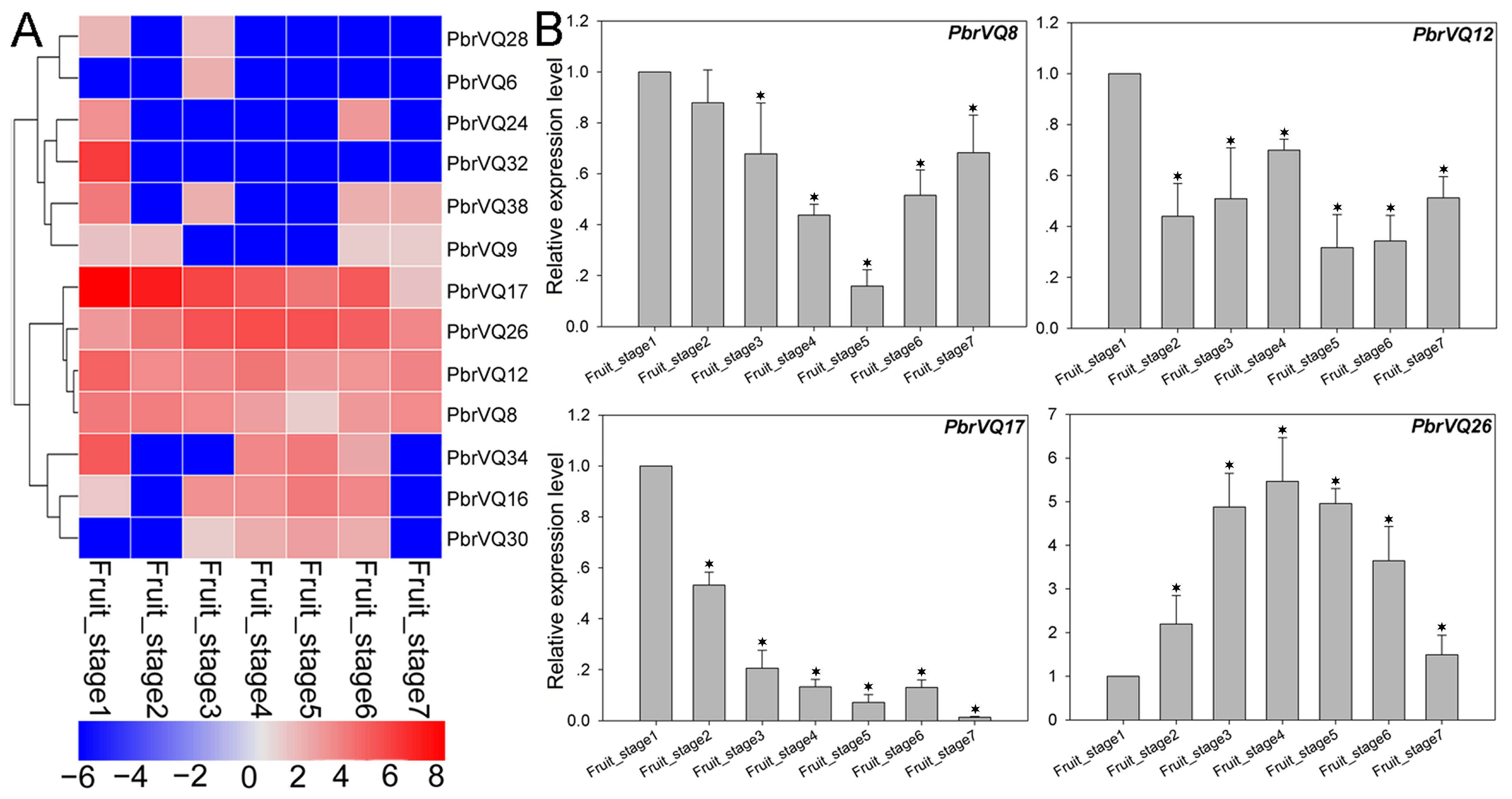
Figure 6.
Comparison of the expression patterns of P. bretschneideri VQ motif-containing genes and their orthologous in P. communis during fruit development. X-axis represents the fruit developmental stages. Y-axis on the left indicates the relative values of FPKM in P. bretschneideri VQ motif-containing genes by blue line charts, and Y-axis on the right represents the relative gene expression levels of qRT-PCR in P. communis by red line charts. The FPKM values of PbVQ genes are presented in Table S5.
Figure 6.
Comparison of the expression patterns of P. bretschneideri VQ motif-containing genes and their orthologous in P. communis during fruit development. X-axis represents the fruit developmental stages. Y-axis on the left indicates the relative values of FPKM in P. bretschneideri VQ motif-containing genes by blue line charts, and Y-axis on the right represents the relative gene expression levels of qRT-PCR in P. communis by red line charts. The FPKM values of PbVQ genes are presented in Table S5.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cao, Y.; Meng, D.; Abdullah, M.; Jin, Q.; Lin, Y.; Cai, Y. Genome Wide Identification, Evolutionary, and Expression Analysis of VQ Genes from Two Pyrus Species. Genes 2018, 9, 224. https://doi.org/10.3390/genes9040224
AMA Style
Cao Y, Meng D, Abdullah M, Jin Q, Lin Y, Cai Y. Genome Wide Identification, Evolutionary, and Expression Analysis of VQ Genes from Two Pyrus Species. Genes. 2018; 9(4):224. https://doi.org/10.3390/genes9040224
Chicago/Turabian StyleCao, Yunpeng, Dandan Meng, Muhammad Abdullah, Qing Jin, Yi Lin, and Yongping Cai. 2018. "Genome Wide Identification, Evolutionary, and Expression Analysis of VQ Genes from Two Pyrus Species" Genes 9, no. 4: 224. https://doi.org/10.3390/genes9040224
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.