Pathogenicity Islands Distribution in Non-O157 Shiga Toxin-Producing Escherichia coli (STEC)


Immunochemistry and Biotechnology Laboratory, Faculty of Veterinary Sciences, Tandil Veterinary Research Center, CONICET-CIC-UNCPBA, University Campus, Tandil 7000, Buenos Aires, Argentina
*
Author to whom correspondence should be addressed.
Genes 2018, 9(2), 81; https://doi.org/10.3390/genes9020081
Submission received: 29 December 2017
/
Revised: 6 February 2018
/
Accepted: 7 February 2018
/
Published: 10 February 2018
(This article belongs to the Special Issue Genetics and Genomics of Foodborne Pathogens)
Abstract
:Shiga toxin-producing Escherichia coli (STEC) are foodborne pathogens associated with outbreaks and hemolytic-uremic syndrome. Cattle and meat foods are the main reservoir and infection source, respectively. Pathogenicity islands (PAIs) play an important role in STEC pathogenicity, and non-locus of the enterocyte effacement(LEE) effector (nle) genes present on them encode translocated substrates of the type III secretion system. A molecular risk assessment based on the evaluation of the nle content has been used to predict which STEC strains pose a risk to humans. The goal was to investigate the distribution of the PAIs OI (O-island)-36 (nleB2, nleC, nleH1-1, nleD), OI-57 (nleG2-3, nleG5-2, nleG6-2), OI-71 (nleA, nleF, nleG, nleG2-1, nleG9, nleH1-2) and OI-122 (ent/espL2, nleB, nleE, Z4321, Z4326, Z4332, Z4333) among 204 clinical, food and animal isolates belonging to 52 non-O157:H7 serotypes. Differences in the frequencies of genetic markers and a wide spectrum of PAI virulence profiles were found. In most LEE-negative strains, only module 1 (Z4321) of OI-122 was present. However, some unusual eae-negative strains were detected, which carried other PAI genes. The cluster analysis, excluding isolates that presented no genes, defined two major groups: eae-negative (determined as seropathotypes (SPTs) D, E or without determination, isolated from cattle or food) and eae-positive (mostly identified as SPTs B, C, or not determined).
1. Introduction
Shiga toxin-producing Escherichia coli (STEC) are heterogeneous foodborne pathogens associated with outbreaks and hemolytic-uremic syndrome (HUS) [1]. Cattle are the main reservoir of STEC and human infections are acquired mainly by ingesting food or water contaminated directly or indirectly with cattle feces. E. coli O157:H7 is the serotype most associated with diseases; however, more current studies have shown that the number of non-O157 STEC infections sometimes surpasses the number of STEC O157 infections [2].
Determinants of bacterial virulence are predominantly encoded by or associated with mobile genetic elements (MGEs) such as phages, plasmids, insertion elements, or transposons. A large number of these determinants are located within pathogenicity islands (PAIs), and can be exchanged among different bacterial species, and assembled and stabilized by selective pressure, leading to emerging pathogenic variants [3]. Increasing evidence shows that differences in virulence between pathogenic and non-pathogenic bacterial strains can be attributed in part to virulence genes located in PAIs [4,5].
Some pathogenic Escherichia coli use a type III secretion system (T3SS), encoded in the locus of the enterocyte effacement (LEE) pathogenicity island, to translocate a wide repertoire of effector proteins into the host cell in order to subvert cell signaling cascades and promote bacterial colonization and survival. Genes encoding type III-secreted effectors are located in the LEE and scattered throughout the chromosome. In addition to genes located on the LEE, a large number of non-LEE effector (nle) genes located on other PAIs have been identified in strains responsible for human infections. These genes are involved in various functions within the host cell, contributing to colonization and full virulence, including anti-apoptotic activities, disruption of host innate immune responses, increase of paracellular permeability, blockage of cell division, disruption of microtubule cytoskeleton, and inhibition of phagocytosis, among others [6].
A molecular risk assessment (MRA) approach based on the evaluation of the nle gene content has been used to predict which STEC strains pose a significant risk to human health [7,8,9,10]. Besides the production of Shiga toxins, the attaching and effacing phenotype encoded by the LEE and the presence of some non-LEE genomic O-islands (OI) such as OI-57, OI-71 and OI-122, are significantly associated with STEC types that are frequently involved in outbreaks and cause hemorrhagic colitis and HUS in humans [4,7,8,10].
OI-57 contains genes nleG2-3, nleG6-2, and nleG5-2 in EDL933 [7,11]. NleG proteins are ubiquitin ligases. Although the exact functions of NleG2-3, NleG6-2, and NleG 5-2 are still unclear, similar proteins have been identified as effectors that suppress the immune response of the host [12].
OI-122 has, at least, six open reading frames (ORF), which show significant homology to known virulence genes. ORF Z4321 is homologous to pagC of Salmonella enterica serovar Typhimurium; ORF Z4326 is homologous to sen of Shigella flexneri; ORFs Z4328 and Z4329 are homologous to non-LEE effector (nle) genes nleB and nleE of Citrobacter rodentium; and ORFs Z4332 and Z4333 are homologous to the enterohemorrhagic E. coli (EHEC) factor for adherence gene cluster efa1 and efa2 found in STEC O157:H7 [4,8,13]. Wickham et al. [14] have described a modular arrangement of OI-122 genes based upon their association with each other across HUS-associated non-O157 STEC strains: module 1 contains Z4318, pagC, and Z4322; module 2 contains Z4323, sen, nleB, and nleE; and module 3 contains the efa gene cluster. The presence of putative transposases in OI-122 has led to the hypothesis that its elements are acquired or lost in a modular manner. It was shown that, while pagC, Z4322, sen, nleB, nleE, and efa1 individually were more prevalent in non-O157 STEC associated with HUS, the simultaneous presence of all of these genes strengthened the association with serious disease [15].
Karmali et al. [4] proposed to group STEC strains into five seropathotypes (SPTs), from A through E, according to their reported frequencies in human illness, their known association with outbreaks and severe disease, and the presence of MGEs such as the LEE and the OI-122 PAI. While nearly all O157:H7/H- strains carry a complete OI-122 (COI-122), a progressive decrease in the prevalence of OI-122 genes in non-O157 STEC belonging to SPT B through E with a concomitant decreasing pathogenicity was observed.
Few studies to date have investigated PAIs other than LEE in STEC. Since PAIs may serve as useful markers to distinguish highly virulent strains from less-virulent strains, the aim of this study was to investigate the prevalence and the distribution of OI-36, OI-57, OI-71 and OI-122 among clinical, food and animal STEC isolates and evaluate the possible public health significance of these PAIs in STEC seropathotypes.
2. Materials and Methods
2.1. Bacterial Strains
A total of 204 STEC isolates belonging to 52 non-O157:H7 serotypes were characterized in this study (see Table S1 in the Supplementary Material). All isolates are part of the Immunochemistry and Biotechnology Laboratory (FCV-UNCPBA, Argentina) collection and had been isolated in Argentina between 1998 and 2014. Twelve of the isolates were obtained from clinical cases (children up to six years old) (H), 73 from meat food, two from vegetables (F), and 117 from cattle (C). All isolates have been previously analyzed by polymerase chain reaction (PCR) for the presence of genes encoding for Shiga toxin 1 and 2 (stx1 and stx2), intimin (eae), and enterohaemolysin (ehxA) [16,17,18,19,20]. Isolates were classified into SPTs, A to E, according to their serotypes, criteria described by Karmali et al. [4]. The assignment of SPTs was based on previously published information [4,5,21].
2.2. Detection of Non-Locus of the Enterocyte Effacement Effector Genes Encoded in Pathogenicity Islands
PCR was used to screen all isolates included in this study. They were examined by the presence of 16 markers of virulence effector genes encoded in genomic OIs different to LEE: OI-36 (nleB2, nleC, nleH1-1, nleD); OI-57 (nleG2-3, nleG5-2, nleG6-2); OI-71 (nleA, nleF, nleG, nleG2-1, nleG9, nleH1-2); and OI-122 (ent/espL2, nleB, nleE) (Table 1). The primers and PCR conditions for the amplification of non-LEE encoded effectors (nle) and ent/espL2 were taken from Coombes et al. [7] and PCR products were visualized on a 1.5% or 2% agarose gel.
2.3. Detection of Particular Genes for the Presence of OI-122
All strains were screened for the presence of OI-122 by testing for four genes (Z4321, Z4326, Z4332, and Z4333) (Table 1). The primers and PCR conditions were taken from Karmali et al. [4]. PCR products were visualized on a 2% agarose gel.
According to Karmali´s proposal (l.c.), the presence of all four genes was considered the presence of a complete OI-122 (COI-122). The absence of one or more of the genes evidenced an incomplete OI-122, whereas the absence of all four genes indicated an absent OI-122.
2.4. Cluster Analysis
A cluster analysis of the studied isolates based on their PAI virulence-associated genes profiles was generated using the BioNumerics v.6.6 software (Applied Maths NV, Sint-Martens-Latem, Belgium).
3. Results
O-Islands are genetic regions absent from nonpathogenic E. coli and frequently contain virulence determinants. The distribution of 16 nle genes and four putative virulence genes encoded in four genomic PAIs (OI-36, OI-57, OI-71, OI-122) among non-O157 STEC strains were analyzed.
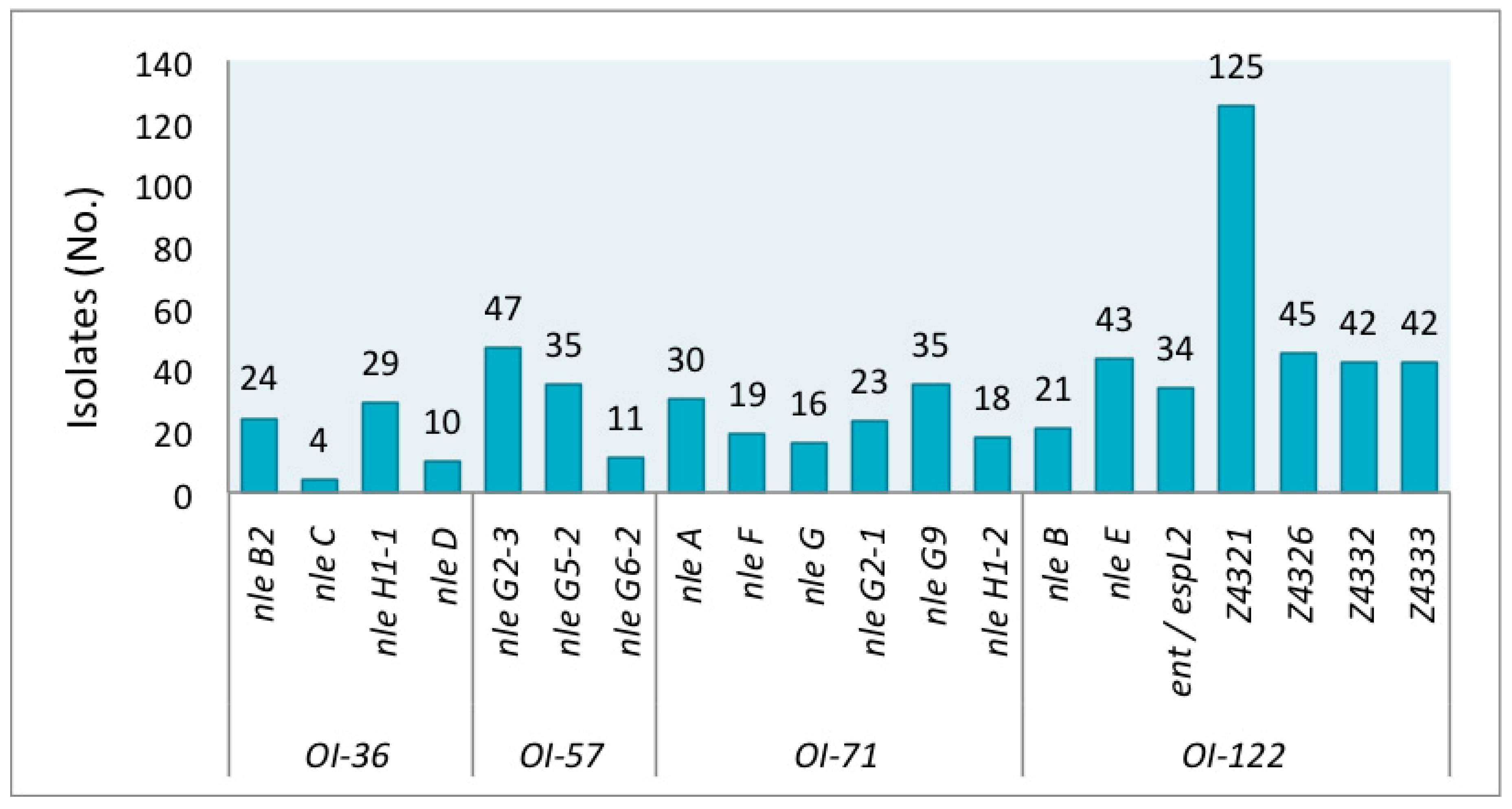
The genetic markers were detected at different frequencies in the STEC studied group. The prevalence of individual virulence genes is shown in Figure 1. Among all strains, the gene Z4321 (pagC) encoded in OI-122 was the most prevalent, resulting in both LEE-positive and LEE-negative isolates. In contrast, nleC encoded on OI-36 was the least prevalent, found in only four isolates of O26:H11 (from human), O145:NM (2 from humans), and O165:NM (from cattle).
A group of forty-six isolates presented no gene. Forty-four strains (from cattle, meat food, and vegetables) out of these 46 strains were eae-negative, as expected, but, strikingly, two were eae-positive. Both were O145:NM strains isolated from humans. The serotypes with absence of the genes studied were: O8:H19, O20:H7, O20:H19, O25:H19, O79:H19, O117:H2, O120:H19, O141:H7, O175:H8, and ONT:H19. A second group comprised of 106 isolates carried only Z4321 (Table S1). Serotypes of these Z4321-positive strains are detailed in the legend of Figure 2.
Most eae-negative strains lacked all 20 tested virulence genes; however, six exceptional strains, which carried additional genes to Z4321 were detected: nleH1-1, nleG2-3, nleG9, Z4326, Z4332, and Z4333. Gene nleH1-1 was detected in strains that belonged to serotypes O8:H16 (#7) and O39:H49 (#43, which also presented Z4332 and Z4333); nleG9 in strains which belonged to serotypes O141:H8 (#91), O171:H2 (#116), and O171:NM (#129); and nleH1-1 + nleG2-3 + nleG9 in one O162:H7 (#113) strain (Figure 2).
The O26:H11 strains are among the strains that have the largest number of virulence genes. They showed 14 to 18 of the 20 genes. Those with more detected genes (16 and 18) were from humans. The strain with 18 genes only lacked nleD and Z4321 (OI-57 and 71 were complete). Notably, two cattle isolates belonging to O118:H16 and O165:NM carried 16 of the 20 analyzed genes, with the presence of genes of each OI.
Table 2 gives details on the prevalence of PAI genes in five STEC eae-positive serotypes represented by more than one isolate. Depending on the serotype, some nle genes were never detected, for example, nleB2, nleC, nleG5-2, nleG6-2, nleF, and nleB in O5:NM or nleH1-1, nleG6-2, nleH1-2, and Z4321 in O145:NM.
In several serotypes (O5:NM, O8:H16, O20:H19, O26:H11, O103:NM, O117:H7, O141:H8, O145:NM, O171:H2, O171:NM, O174:H21, O177:NM, O178:H19) more than one virulence profile were found.
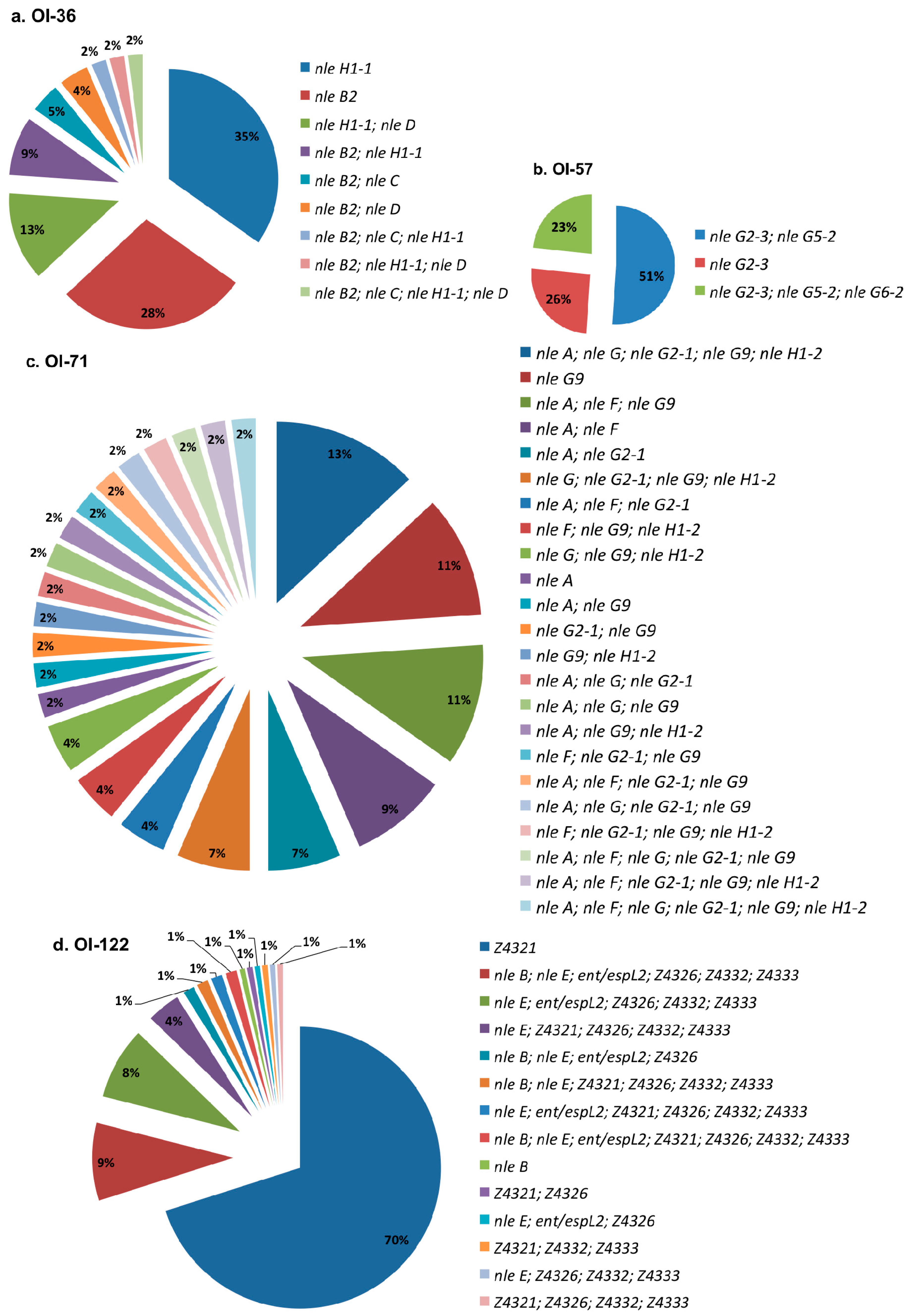
Figure 3 shows the distribution of profiles for each investigated PAI. Among the analyzed STEC, (a) OI-36 showed nine profiles, with the most prevalent being nleH1-1 and nleB2; (b) OI-57 exhibited three profiles, including each one of these profiles with one, two, or three genes. Approximately half of the strains presented nleG2-3 and nleG5-2 simultaneously; (c) OI-71 showed 23 profiles, with each one represented by 2–13% of the strains; and (d) OI-122 displayed 14 profiles, with the most prevalent, Z4321, present in 70% of the isolates.
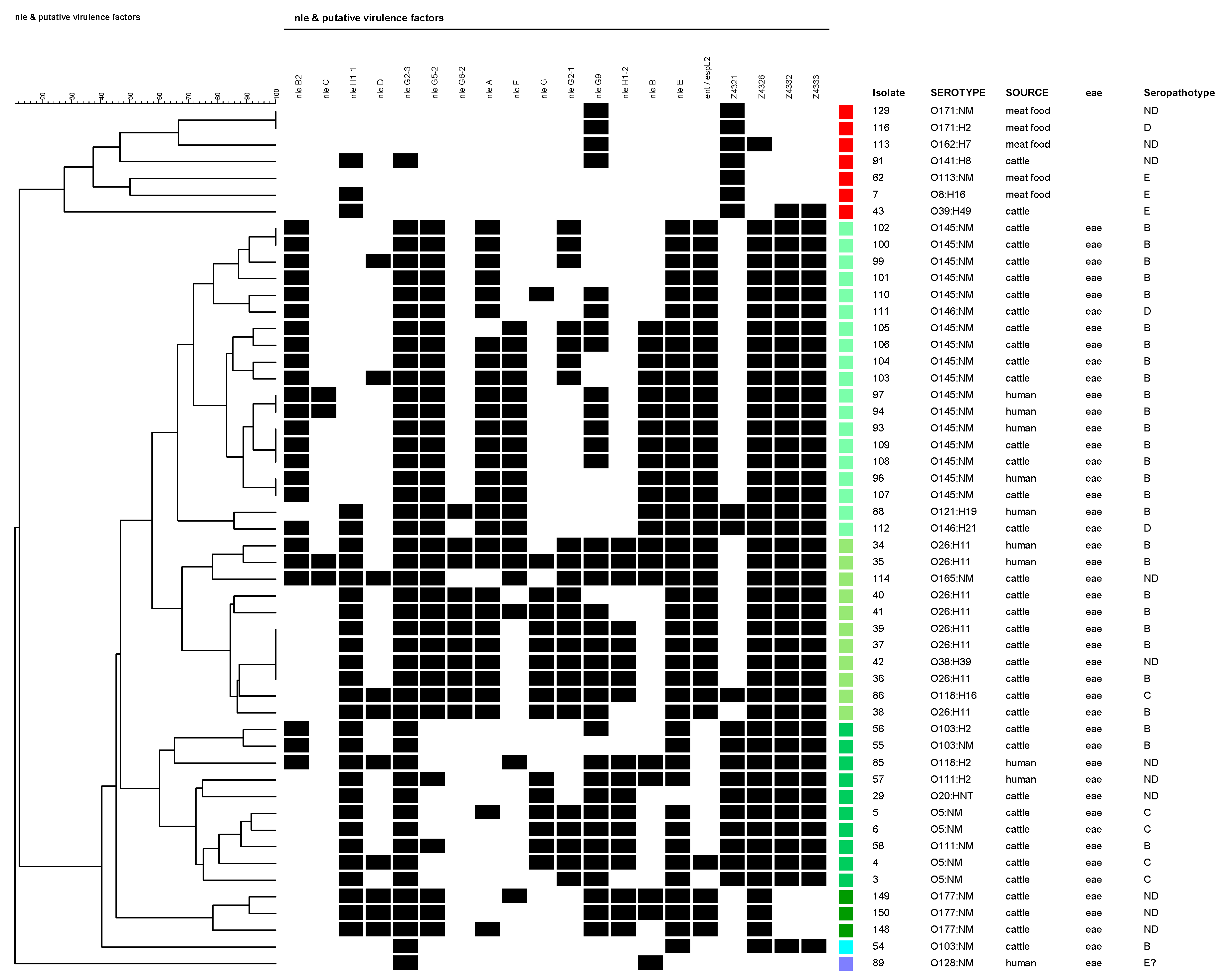
Only O26:H11 (humans -H- and cattle -C-), O38:H39 (C), O118:H16 (C), and O121:H19 (H) strains carried a complete OI-57. Meanwhile, only one O165:NM strain isolated from cattle had a complete OI-36 and one other, O26:H11 (H), presented a complete OI-71 and only three strains, one O111:H2 and one O121:H19 from a human, and one O146:H21 from cattle carried a complete OI-122. All of them were eae-positive serotypes.
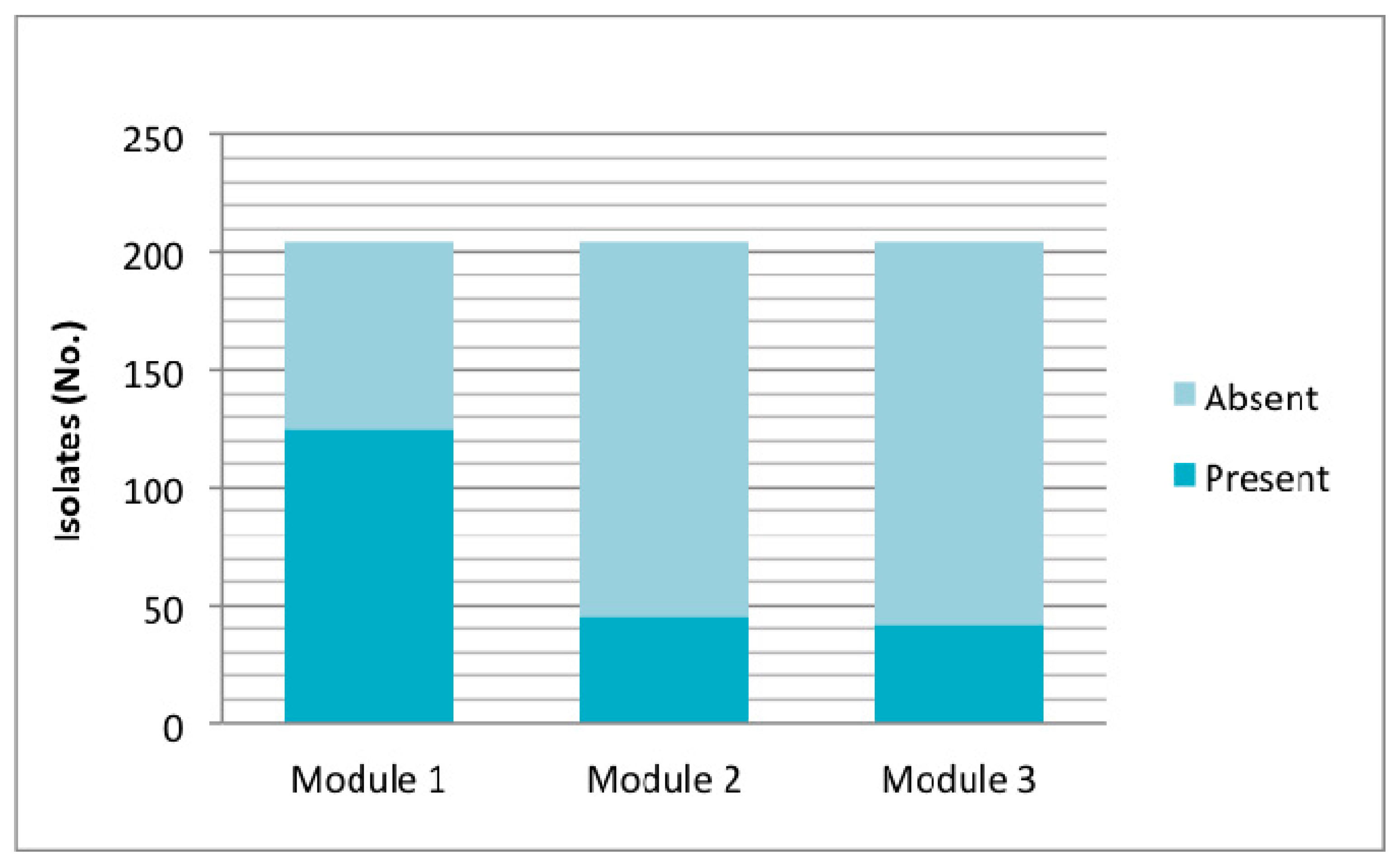
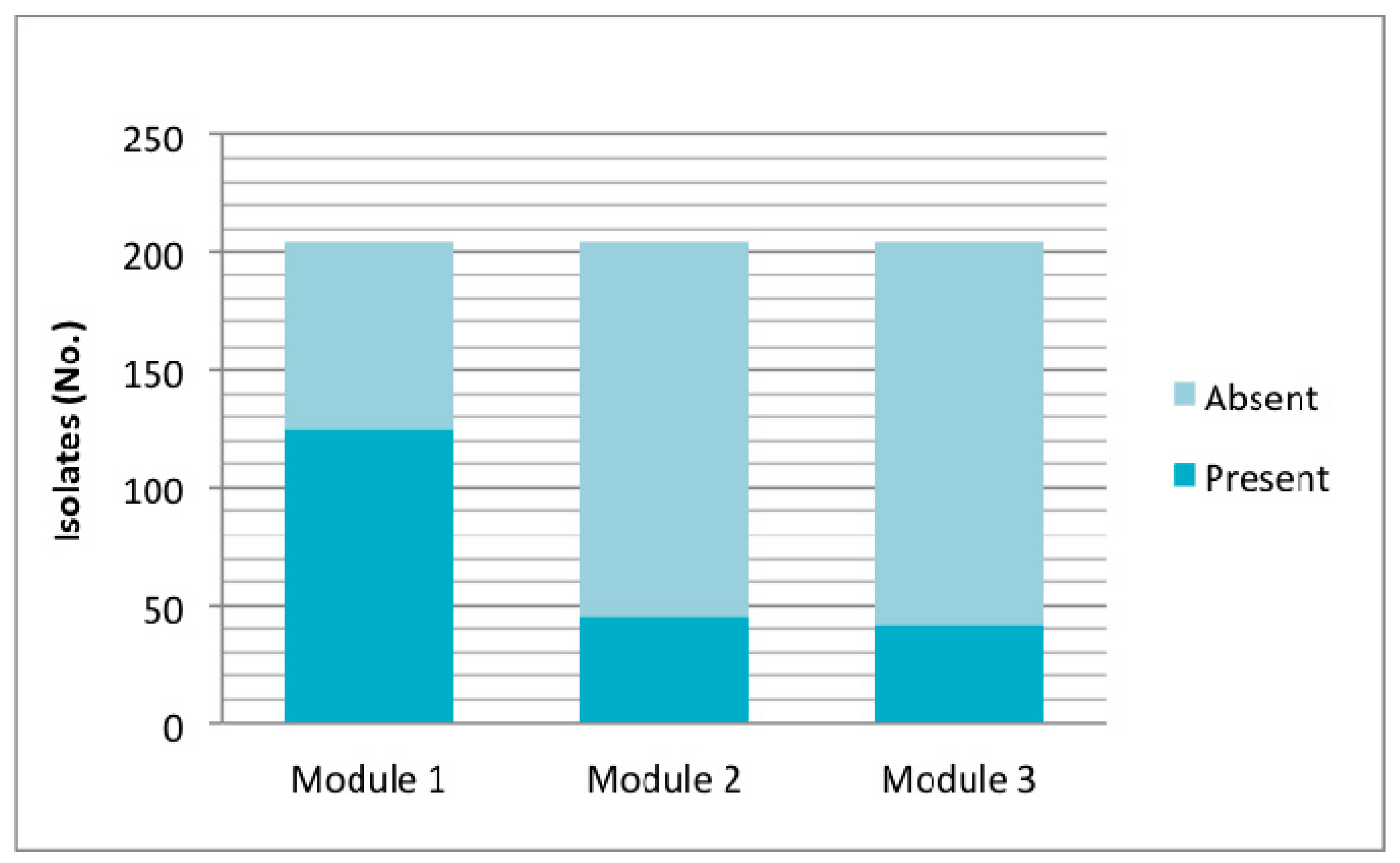
Particularly, in relation to OI-122, its presence was determined by seven marker genes located in different regions of the island. Of the 204 isolates studied, 157 (76.9%) were positive for at least one gene of OI-122, of which 45 isolates were eae+ and 112 eae−. Among the eae-isolates, three profiles were obtained: Z4321 (n = 110) (module 1); Z4321/Z4326 (n = 1) (modules 1 and 2); and Z4321/Z4332/Z4333 (modules 1 and 3) (n = 1) (Figure 4). Analyzing OI-122 in a modular way, module 1 was present (at least presence of one gene) in 61.3% of the strains; module 2 was present in 22.5%; and module 3 was present in 20.5% (Figure 4).
According to the criteria described by Karmali et al. [4] based on the detection of COI-122 that requires testing for four virulence putative genes (Z4321; Z4326; Z4332; Z4333), 13 of 204 (6.4%) isolates studied had a complete OI-122, 143 (70.1%) had an incomplete OI-122, and OI-122 was absent in 48 (23.5%) strains.
The cluster analysis (which did not include isolates that presented no genes), presented in Figure 2, divided the studied set of strains into two major groups: (A) eae-negative, and (B) eae-positive isolates. Within the eae-negative group, the isolates were determined as belonging to SPTs D, E, or without determination and were isolated from cattle or meat food. Within the eae-positive group, the isolates were mostly identified as SPTs B, C, or not determined. Exceptionally, two isolates were characterized as SPT D (O146:H21 and O146:NM, both from cattle). Among this group, four principal branches could be differentiated (represented in four different green shades in Figure 2): (B1) O145:NM (+O121:H8+O146:H21/NM) isolates; (B2) O26:H11 (+O118:H16+O165:NM) isolates; (B3) O5, O103, O111 (+O20:HNT+O118:H2) isolates; and (B4) O177:NM isolates.
4. Discussion
The presence of nle genes and the number of genes carried by an E. coli strain are important criteria for estimating its virulence potential [7]. Previous evidence shows that virulence genes located on PAIs can be used to identify new and emerging pathogenic bacteria [4,5]. In this study, 18 isolates carried more than 60% of the 20 OI-genes analyzed. These isolates belong to the O5:NM, O26:H11, O38:H39, O118:H2/H16, O121:H19, O145:NM, O146:H21, and O165:NM serotypes. From these, four isolates, belonging to O26:H11 from humans, and O118:H16 and O165:NM from cattle, were classified as “virulence top ranking”, presenting between 80 and 90% of the genes. The O26:H11 strains represent a group with a high virulence potential for humans. This is also corroborated by their serotype, which is associated with the classical EHEC serotype.
This study detected several profiles for each PAI, especially in different serotypes, and that one or more PAIs virulence genes could be absent in STEC non-O157. In agreement with Ju et al. [5,23], we postulate that PAI could be unstable in STEC. However, as highlighted in the results summarized in Table 2, the presence or absence of particular genes´ intra-serotype is not random; instead, there are specific arrays of virulence factors. Further studies using whole-genome sequencing might identify additional virulence markers and increase the understanding of their contribution to human disease.
In spite of the fact that LEE is important, it is not essential for STEC pathogenesis, and sporadic cases and small outbreaks of STEC infections have been caused by LEE-negative strains [24]. Although the absence of eae was associated with the absence of many of the virulence genes, unlike previous publications, such as Franz et al. [22], this study detected some unusual eae-negative cattle and food strains belonging to serotypes O8:H16, O39:H49, O141:H8, O162:H7, and O171:H2/NM, which carried genes nleH1-1, nleG2-3, nleG9, Z4326, Z4332, or Z4333. Such strains, unlike other LEE-negative strains, may cause human diseases. Indeed, some of these serotypes, for example, O171:H2 and O171:NM, have been associated with HUS and/or bloody diarrhea in children in Argentina [25]. Therefore, the risk represented by them to public health should be monitored. On the contrary, two eae-positive strains, O145:NM isolated from a human, lacked all of the studied PAI genes.
It has been proposed and agreed that COI-122–positive strains belong to SPTs associated with outbreaks (A and B) and SPTs associated with HUS (A, B, and C) [4,14,15]. However, in this study, some COI-122–positive strains identified with different SPTs or not determined, O20:HNT (SPT not determined), O146:H21 (D) strains from cattle, and O111:H2 and O118:H2 (SPT not determined) from humans were detected. Also, in relation to OI-122, and coincidently with the results from Konczy et al. [8], results showed that when LEE was absent, only module 1 (gene Z4321) was present. These strains that carry the pagC-like gene exclusively (modules 2 and 3 absent) would have a non-synonymous substitution in (His→Gln) [8]. Wickham et al. [14] postulated that there was a significant association between the presence of a combination of OI-122 markers, such as the pagC-like gene and sen, nleB, and efa-1, and HUS after infection in non-O157 E. coli. Four strains with this profile: three human strains (one O111:H2, one O118:H2, and one O121:H19), and, interestingly, one O146:H21 strain isolated from the bovine group were found.
Newton et al. [26] proposed that NleE and NleB (OI-122) contribute to pathogenesis by inhibiting an initial host inflammatory response (contribute to the suppression of innate signaling pathways) to allow the bacteria to persist in the early stages of infection. NleE appears to obstruct nuclear translocation of Rel family transcriptional activators, while allowing the nuclear import of a transcriptional repressor, resulting in the suppression of IL8 expression. NleB prevents translocation of the immune regulator nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB) to the cell nucleus [26]. Gene nleB was detected in O26:H11, O111:H2, O118:H2, O121:H19, O128:NM, and O145:NM strains isolated from patients, and in O145:NM, O146:H21, O165:NM, and O177:NM cattle strains. The O26:H11 strains from humans presented nleB and nleE, but cattle O26:H11 strains presented only nleE. Unlike a study of Karmali et al. [4] in which they postulated that in cases where a serotype was represented by more than one strain, all strains belonging to the same serotype had identical patterns of distribution of OI-122 genes, in this work different virulence profiles for OI-122 in isolates belonging to the same serotype were found (see Figure 2). In previous work, in which a subset of these strains was analyzed using multilocus sequence typing (MLST), different sequence types (STs) in the same serotype were detected [27]. Further studies should determine if different STs correspond to different PAI-virulence profiles.
According to the results from Ju et al. [5], some of OI-122 and OI-57 nle (nleB, nleG2-3, nleG5-2, and nleG6-2) were primarily associated with eae-positive STEC strains and associated with SPTs that cause severe diseases (SPT A, B, C), offering an important basis for STEC MRA. But, based on the MRA framework, which uses nle genes as sole markers, all eae-negative virulence STEC strains, including HUS-associated O113:H21 and O91:H21, would be categorized as harmless. Therefore, we agree that additional markers, especially for eae-negative STEC, are needed to absolutely predict the potential of an STEC strain to cause disease. Recently, a novel PAI named Locus of Adhesion and Autoaggregation (LAA), exclusively present in a subset of LEE-negative STEC strains, with the mostly clinically relevant ones including O91:H21, O113:H21, and O174:H21, has been proposed. Several virulence factors, among them Hes, which participates in colonization-associated phenotypes, and other ones participating in adhesion and autoaggregation such as Pag-C, Iha, and, Agn43 are encoded in it [28].
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4425/9/2/81/s1. Table S1: Non-O157 STEC isolates analyzed in this study and PAIs-encoded virulence genes results.
Acknowledgments
We thank M. R. Ortiz for her technical assistance. This study was supported by Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), and Secretaría de Ciencia, Arte y Tecnología (SECAT)-Universidad Nacional del Centro de la Provincia de Buenos Aires (UNICEN).
Author Contributions
J.S.C., A.V.B. and A.M.S. conceived and designed the experiments; J.S.C. performed the experiments; J.S.C., A.V.B., J.G. and A.M.S. analyzed the data; A.M.S. and J.S.C. wrote the paper. All authors have read and approved the final version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Karmali, M.A.; Gannon, V.; Sargeant, J.M. Verocytotoxin-producing Escherichia coli (VTEC). Vet. Microbiol. 2010, 140, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Gould, L.H.; Mody, R.K.; Ong, K.L.; Clogher, P.; Cronquist, A.B.; Garman, K.N.; Lathrop, S.; Medus, C.; Spina, N.L.; Webb, T.H.; et al. Increased recognition of non-O157 Shiga toxin-producing Escherichia coli infections in the United States during 2000-2010: Epidemiologic features and comparison with E. coli O157 infections. Foodborne Pathog. Dis. 2013, 10, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.; Hensel, M. Pathogenicity islands in bacterial pathogenesis. Clinical Microbiology Reviews 2004, 17, 14–56. [Google Scholar] [CrossRef] [PubMed]
- Karmali, M.A.; Mascarenhas, M.; Shen, S.; Ziebell, K.; Johnson, S.; Reid-Smith, R.; Isaac-Renton, J.; Clark, C.; Rahn, K.; Kaper, J.B. Association of genomic O Island 122 of Escherichia coli EDL 933 with Verocytotoxin-Producing Escherichia coli seropathotypes that are linked to epidemic and/or serious disease. J. Clin. Microbiol. 2003, 41, 4930–4940. [Google Scholar] [CrossRef] [PubMed]
- Ju, W.; Shen, J.; Toro, M.; Zhao, S.; Menga, J. Distribution of pathogenicity islands OI-122, OI-43/48, and OI-57 and a high-pathogenicity island in Shiga toxin-producing Escherichia coli. Appl. Environ. Microbiol. 2013, 79, 3406–3412. [Google Scholar] [CrossRef] [PubMed]
- García-Angulo, V.A.; Martínez-Santos, V.I.; Villaseñor, T.; Santana, F.J.; Huerta-Saquero, A.; Martínez, L.C.; Jiménez, R.; Lara-Ochoa, C.; Téllez-Sosa, J.; Bustamante, V.H.; et al. A distinct regulatory sequence is essential for the expression of a subset of nle genes in attaching and effacing Escherichia coli. J. Bacteriol. 2012, 194, 5589–5603. [Google Scholar] [CrossRef]
- Coombes, B.K.; Wickham, M.E.; Mascarenhas, M.; Gruenheid, S.; Finlay, B.B.; Karmali, M.A. Molecular analysis as an aid to assess the public health risk of non-O157 shiga toxin-producing Escherichia coli strains. Appl. Environ. Microbiol. 2008, 74, 2153–2160. [Google Scholar] [CrossRef] [PubMed]
- Konczy, P.; Ziebell, K.; Mascarenhas, M.; Choi, A.; Michaud, C.; Kropinski, A.M.; Whittam, T.S.; Wickham, M.; Finlay, B.; Karmali, M.A. Genomic O island 122, locus for enterocyte effacement, and the evolution of virulent verocytotoxin-producing Escherichia coli. J. Bacteriol. 2008, 190, 5832–5840. [Google Scholar] [CrossRef] [PubMed]
- Bugarel, M.; Beutin, L.; Martin, A.; Gill, A.; Fach, P. Micro-array for the identification of Shiga toxin-producing Escherichia coli (STEC) seropathotypes associated with Hemorrhagic Colitis and Hemolytic Uremic Syndrome in humans. Int. J. Food Microbiol. 2010, 142, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Bugarel, M.; Beutin, L.; Fach, P. Low-density macroarray targeting non-locus of enterocyte effacement effectors (nle genes) and major virulence factors of Shiga toxin-producing Escherichia coli (STEC): A new approach for molecular risk assessment of STEC isolates. Appl. Environ. Microbiol. 2010, 76, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Imamovic, L.; Tozzoli, R.; Michelacci, V.; Minelli, F.; Marziano, M.L.; Caprioli, A.; Morabito, S. OI-57, a genomic island of Escherichia coli O157, is present in other seropathotypes of Shiga toxin-producing E. coli associated with severe human disease. Infect. Immun. 2010, 78, 4697–4704. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Skarina, T.; Yee, A.; Jobin, M.C.; Dileo, R.; Semesi, A.; Fares, C.; Lemak, A.; Coombes, B.K.; Arrowsmith, C.H.; et al. NleG type 3 effectors from enterohaemorrhagic Escherichia coli are U-Box E3 ubiquitin ligases. PLoS Pathog. 2010, 6, e1000960. [Google Scholar] [CrossRef] [PubMed]
- Morabito, S.; Tozzoli, R.; Oswald, E.; Caprioli, A. A mosaic pathogenicity island made up of the locus of enterocyte effacement and a pathogenicity island of Escherichia coli O157:H7 is frequently present in attaching and effacing E. coli. Infect. Immun. 2003, 71, 3343–3348. [Google Scholar] [CrossRef] [PubMed]
- Wickham, M.E.; Lupp, C.; Mascarenhas, M.; Vazquez, A.; Coombes, B.K.; Brown, N.F.; Coburn, B.A.; Deng, W.; Puente, J.L.; Karmali, M.A.; Finlay, B.B. Bacterial genetic determinants of non-O157 STEC outbreaks and hemolytic-uremic syndrome after infection. J. Infect. Dis. 2006, 194, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Buvens, G.; Piérard, D. virulence profiling and disease association of verocytotoxin-producing Escherichia coli O157 and Non-O157 isolates in Belgium. Foodborne Pathog. Dis. 2012, 9, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Parma, A.E.; Sanz, M.E.; Blanco, J.E.; Blanco, J.; Viñas, M.R.; Blanco, M.; Padola, N.L.; Etcheverria, A.I. Virulence genotypes and serotypes of verotoxigenic Escherichia coli isolated from cattle and foods in Argentina: Importance in public health. Eur. J. Epidemiol. 2000, 16, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Blanco, M.; Padola, N.L.; Krüger, A.; Sanz, M.E.; Blanco, J.E.; Gonzalez, E.A.; Dahbi, G.; Mora, A.; Bernardez, M.I.; Etcheverria, A.I.; et al. Virulence genes and intimin types of Shiga-toxin-producing Escherichia coli isolated from cattle and beef products in Argentina. Int. Microbiol. 2004, 7, 269–276. [Google Scholar] [PubMed]
- Sanz, M.E.; Cristina, V.; Elida, E.; Arroyo, G. Prevalencia de Escherichia coli verotoxigénico en productos cárnicos de la ciudad de Tandil. La Ind. Cárnica Latinoam. 2007, 146, 56–58. [Google Scholar]
- Rivero, M.A.; Passucci, J.A.; Rodriguez, E.M.; Parma, A.E. Role and clinical course of verotoxigenic Escherichia coli infections in childhood acute diarrhoea in Argentina. J. Med. Microbiol. 2010, 59, 345–352. [Google Scholar] [CrossRef] [PubMed]
- González, J.; Cadona, J.S.; Sanz, M.; Bustamante, A.V.; Sanso, A.M. Molecular characterization of diarrheagenic Escherichia coli isolated from vegetables in Argentina. Int. J. Food Microbiol. 2017, 261, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Toma, C.; Espinosa, E.M.; Song, T.; Miliwebsky, E.; Chinen, I.; Iyoda, S.; Iwanaga, M.; Rivas, M. Distribution of putative adhesins in different seropathotypes of Shiga toxin-producing Escherichia coli. J. Clin. Microbiol. 2004, 42, 4937–4946. [Google Scholar] [CrossRef] [PubMed]
- Franz, E.; Van Hoek, A.H.A.M.; Wuite, M.; Van Der Wal, F.J.; De Boer, A.G.; Bouw, E.I.; Aarts, H.J.M. Molecular hazard identification of non-O157 Shiga toxin-producing Escherichia coli (STEC). PLoS ONE 2015, 10, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Ju, W.; Rump, L.; Toro, M.; Shen, J.; Cao, G.; Zhao, S.; Meng, J. Pathogenicity islands in Shiga toxin-producing Escherichia coli O26, O103, and O111 isolates from humans and animals. Foodborne Pathog. Dis. 2014, 11, 342–345. [Google Scholar] [CrossRef] [PubMed]
- Chui, L.; Li, V.; Fach, P.; Delannoy, S.; Malejczyk, K.; Patterson-Fortin, L.; Poon, A.; King, R.; Simmonds, K.; Scott, A.N.; Lee, M.C. Molecular profiling of Escherichia coli O157:H7 and non-O157 strains isolated from humans and cattle in Alberta, Canada. J. Clin. Microbiol. 2015, 53, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Rivas, M.; Miliwebsky, E.; Chinen, I.; Deza, N.; Leotta, G.A. Epidemiología del síndrome urémico hemolítico en Argentina. Diagnóstico del agente etiológico, reservorios y vías de transmisión. Medicina 2006, 66, 27–32. [Google Scholar] [PubMed]
- Newton, H.J.; Pearson, J.S.; Badea, L.; Kelly, M.; Lucas, M.; Holloway, G.; Wagstaff, K.M.; Dunstone, M.A.; Sloan, J.; Whisstock, J.C.; et al. The type III effectors NleE and NleB from enteropathogenic E. coli and Ospz from Shigella block nuclear translocation of NF-κB p65. PLoS Pathog. 2010, 6, e1000898. [Google Scholar] [CrossRef] [PubMed]
- Cadona, J.S.; Bustamante, A.V.; González, J.; Sanso, A.M. Genetic relatedness and novel sequence types of Non-O157 Shiga toxin-producing Escherichia coli strains isolated in Argentina. Front. Cell. Infect. Microbiol. 2016, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Montero, D.A.; Velasco, J.; Del Canto, F.; Puente, J.L.; Padola, N.L.; Rasko, D.A.; Farfán, M.; Salazar, J.C.; Vidal, R. Locus of adhesion and autoaggregation (LAA), a pathogenicity island present in emerging Shiga Toxin-producing Escherichia coli strains. Sci. Rep. 2017, 7, 7011. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Prevalence of individual Pathogenicity Islands (PAIs) (OI-36, OI-57, OI-71, OI-122) virulence genes in shiga toxin-producing Escherichia coli (STEC) non-O157:H7.
Figure 1.
Prevalence of individual Pathogenicity Islands (PAIs) (OI-36, OI-57, OI-71, OI-122) virulence genes in shiga toxin-producing Escherichia coli (STEC) non-O157:H7.
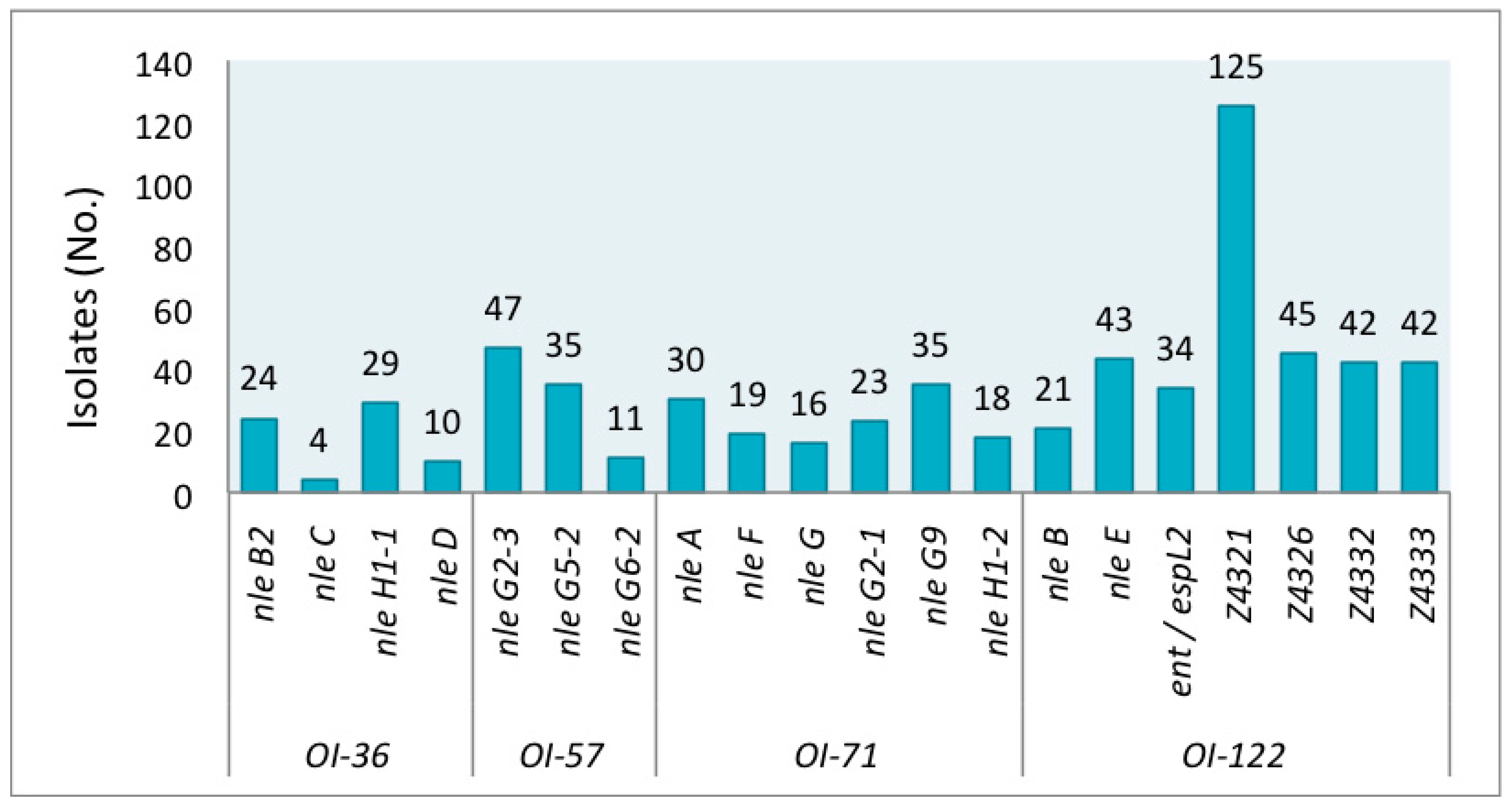
Figure 2.
Cluster analysis of PAIs OI-36, OI-57, OI-71, and OI-122 virulence-associated genes in clinical, cattle, meat food, and vegetable non-O157 STEC isolates. A black box indicates the presence of the target sequence in the strain and a white box, the absence. Isolates that presented no gene were excluded. The colour red represents all the eae-negative isolates; tree branch corresponding to isolate #62 also includes all the isolates with profile Z4321-positive, O2:H5 (1), O2:NM (1), O8:H16 (2), O15:H21 (1), O20:H19 (1), O22:H8 (3), O39:H49 (2), O88:H21 (1), O91:H21 (5), O113:NM (10: strain #62 + 9), O113:H21 (5), O116:H21 (1), O117:H7 (6), O171:HNT, O171:H2 (9), O171:NM (1), O174:H21 (14), O178:H19 (4), O185:H7 (1), ONT:H7 (5), ONT:H8 (2), ONT:H21 (13), ONT:HNT (1), and ONT:NM (16). All of them were isolated from cattle or meat food. All the studied strains belonging to serotypes in bold had the same profile. The remaining colours are eae-positive isolates.
Figure 2.
Cluster analysis of PAIs OI-36, OI-57, OI-71, and OI-122 virulence-associated genes in clinical, cattle, meat food, and vegetable non-O157 STEC isolates. A black box indicates the presence of the target sequence in the strain and a white box, the absence. Isolates that presented no gene were excluded. The colour red represents all the eae-negative isolates; tree branch corresponding to isolate #62 also includes all the isolates with profile Z4321-positive, O2:H5 (1), O2:NM (1), O8:H16 (2), O15:H21 (1), O20:H19 (1), O22:H8 (3), O39:H49 (2), O88:H21 (1), O91:H21 (5), O113:NM (10: strain #62 + 9), O113:H21 (5), O116:H21 (1), O117:H7 (6), O171:HNT, O171:H2 (9), O171:NM (1), O174:H21 (14), O178:H19 (4), O185:H7 (1), ONT:H7 (5), ONT:H8 (2), ONT:H21 (13), ONT:HNT (1), and ONT:NM (16). All of them were isolated from cattle or meat food. All the studied strains belonging to serotypes in bold had the same profile. The remaining colours are eae-positive isolates.
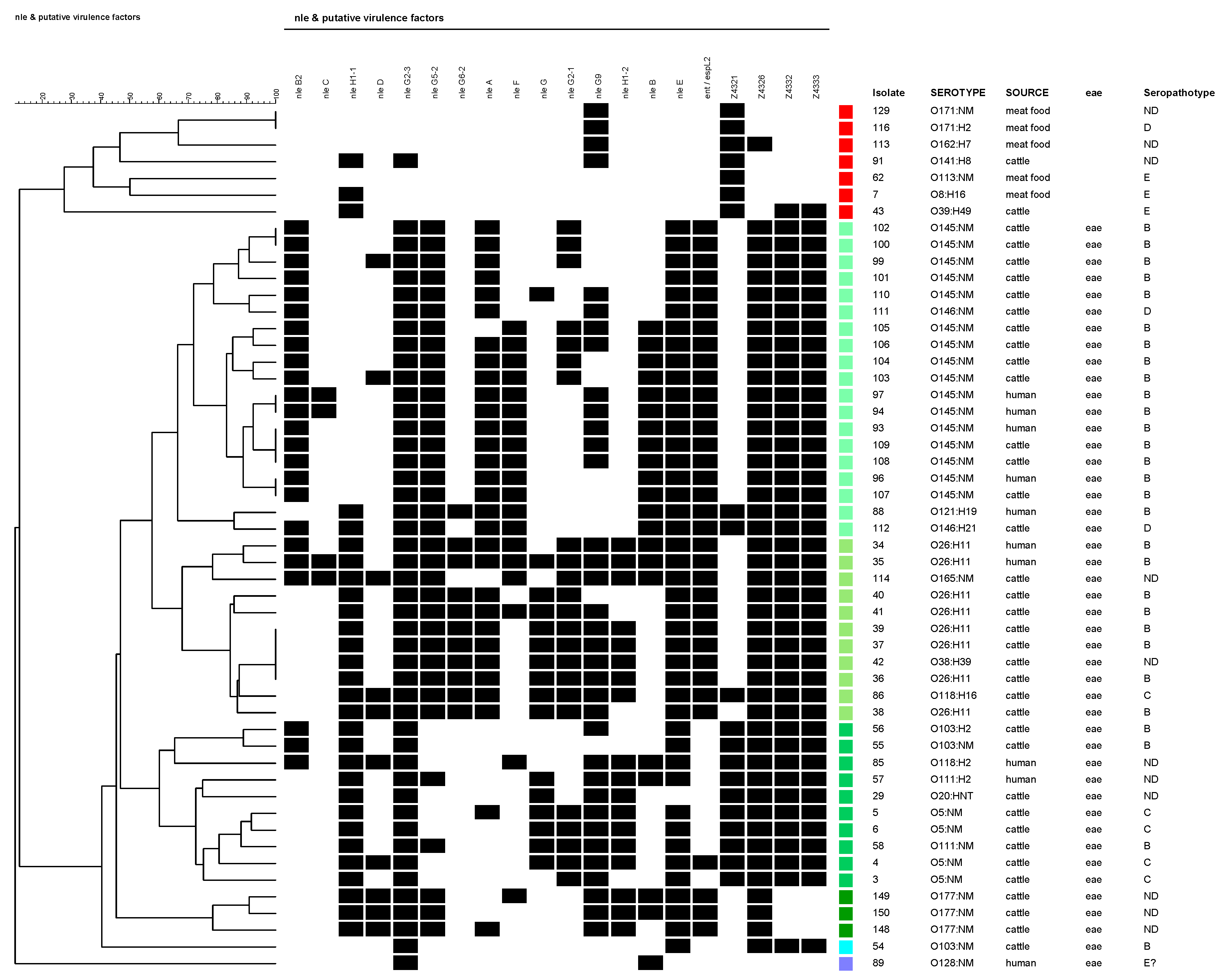
Figure 3.
Virulence profiles distribution for each studied pathogenicity island in OI-encoded genes-positive STEC non-O157:H7. (a) OI-36; (b) OI-57; (c) OI-71; (d) OI-122.
Figure 3.
Virulence profiles distribution for each studied pathogenicity island in OI-encoded genes-positive STEC non-O157:H7. (a) OI-36; (b) OI-57; (c) OI-71; (d) OI-122.
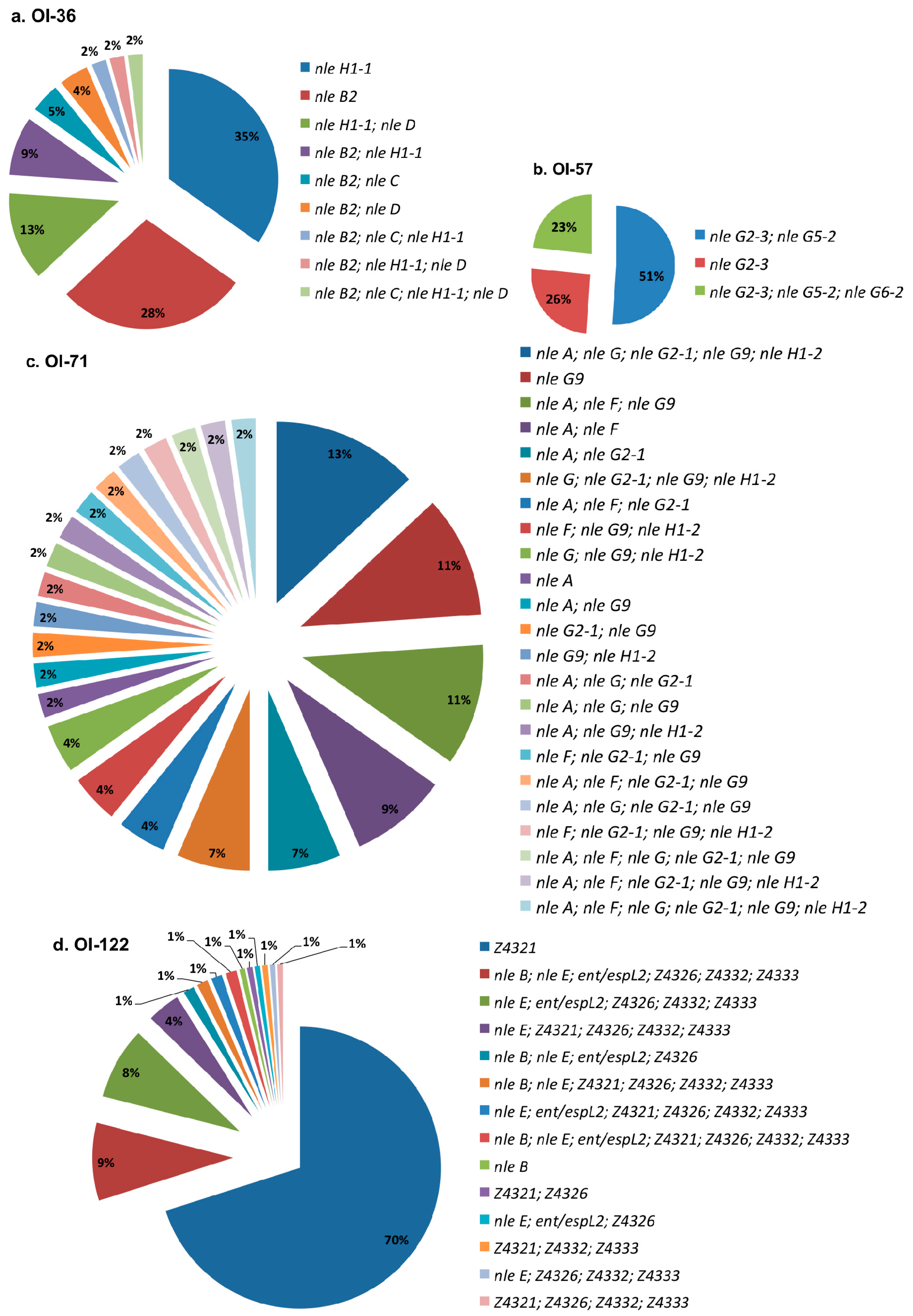
Figure 4.
Presence of OI-122 according to marker genes located in the three modules; module 1: Z4321; module 2: nleB, nleE, ent/espL2, Z4326; and module 3: Z4332, Z4333. Presence of each module was considered by detecting at least one marker of it.
Figure 4.
Presence of OI-122 according to marker genes located in the three modules; module 1: Z4321; module 2: nleB, nleE, ent/espL2, Z4326; and module 3: Z4332, Z4333. Presence of each module was considered by detecting at least one marker of it.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Virulence genes encoded in pathogenicity islands (PAIs) analyzed in this study.
| PAIs | Target | Encoded Protein or Family Effector 1 |
|---|---|---|
| O-Island 36 | nle B2 | Non-LEE encoded type III effector |
| nle C | Immunomodulation, zinc-metalloprotease | |
| nle H1-1 | Immunomodulation | |
| nle D | Immunomodulation, zinc-metalloprotease | |
| O-Island 57 | nle G2-3 | Ubiquitin ligase |
| nle G5-2 | Ubiquitin ligase | |
| nle G6-2 | Ubiquitin ligase | |
| O-Island 71 | nle A | Disruption tight junctions and protein trafficking |
| nle F | Disruption protein trafficking | |
| nle G | Ubiquitin ligase | |
| nle G2-1 | Ubiquitin ligase | |
| nle G9 | Ubiquitin ligase | |
| nle H1-2 | Immunomodulation | |
| O-Island 122 | nle B | Immunomodulation |
| nle E | Immunomodulation | |
| ent/espL2 | Microcolony formation and F-actin aggregation | |
| Z4321 (pagC) | Similarity to Salmonella enterica serovar Typhimurium PhoP-activated gene C | |
| Z4326 (sen) | Similarity to Shigella flexneri enterotoxin 2 | |
| Z4332 (efa1) | EHEC factor for adherence | |
| Z4333 (efa2) | EHEC factor for adherence |
Table 2.
Prevalence of PAI virulence genes in STEC eae-positive serotypes represented by more than one isolate: O5:NM, O26:H11, O103 (O103:NM and O103:H2), O145:NM, and O177:NM.
Table 2.
Prevalence of PAI virulence genes in STEC eae-positive serotypes represented by more than one isolate: O5:NM, O26:H11, O103 (O103:NM and O103:H2), O145:NM, and O177:NM.
| % Isolates Positive for Gene | ||||||
|---|---|---|---|---|---|---|
| PAI | Gene | O5:NM (n = 4) | O26:H11 (n = 8) | O103 (n = 3) | O145:NM (n = 18) | O177:NM (n = 3) |
| O-Island 36 | nle B2 | 0 | 25 | 66.7 | 88.9 | 0 |
| nle C | 0 | 12.5 | 0 | 11.1 | 0 | |
| nle H1-1 | 100 | 100 | 66.7 | 0 | 100 | |
| nle D | 25 | 12.5 | 0 | 11.1 | 100 | |
| O-Island 57 | nle G2-3 | 100 | 100 | 100 | 94.4 | 100 |
| nle G5-2 | 0 | 100 | 0 | 88.9 | 100 | |
| nle G6-2 | 0 | 100 | 0 | 0 | 0 | |
| O-Island 71 | nle A | 25 | 100 | 0 | 83.3 | 33.3 |
| nle F | 0 | 37.5 | 0 | 61.1 | 33.3 | |
| nle G | 75 | 87.5 | 0 | 5.5 | 0 | |
| nle G2-1 | 100 | 87.5 | 0 | 38.9 | 0 | |
| nle G9 | 100 | 87.5 | 33.3 | 44.4 | 100 | |
| nle H1-2 | 75 | 62.5 | 0 | 0 | 100 | |
| O-Island 122 | nle B | 0 | 25 | 0 | 61.1 | 66.7 |
| nle E | 100 | 100 | 100 | 88.9 | 100 | |
| ent/espL2 | 25 | 100 | 0 | 88.9 | 100 | |
| Z4321 (pagC) | 100 | 0 | 66.7 | 0 | 0 | |
| Z4326 (sen) | 100 | 100 | 100 | 88.9 | 100 | |
| Z4332 (efa1) | 100 | 100 | 100 | 88.9 | 0 | |
| Z4333 (efa2) | 100 | 100 | 100 | 88.9 | 0 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cadona, J.S.; Bustamante, A.V.; González, J.; Sanso, A.M. Pathogenicity Islands Distribution in Non-O157 Shiga Toxin-Producing Escherichia coli (STEC). Genes 2018, 9, 81. https://doi.org/10.3390/genes9020081
AMA Style
Cadona JS, Bustamante AV, González J, Sanso AM. Pathogenicity Islands Distribution in Non-O157 Shiga Toxin-Producing Escherichia coli (STEC). Genes. 2018; 9(2):81. https://doi.org/10.3390/genes9020081
Chicago/Turabian StyleCadona, Jimena Soledad, Ana Victoria Bustamante, Juliana González, and Andrea Mariel Sanso. 2018. "Pathogenicity Islands Distribution in Non-O157 Shiga Toxin-Producing Escherichia coli (STEC)" Genes 9, no. 2: 81. https://doi.org/10.3390/genes9020081
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.