
Magnetofection Enhances Lentiviral-Mediated Transduction of Airway Epithelial Cells through Extracellular and Cellular Barriers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lentiviral Vector Production and Titration
2.2. Paramagnetic Nanoparticles
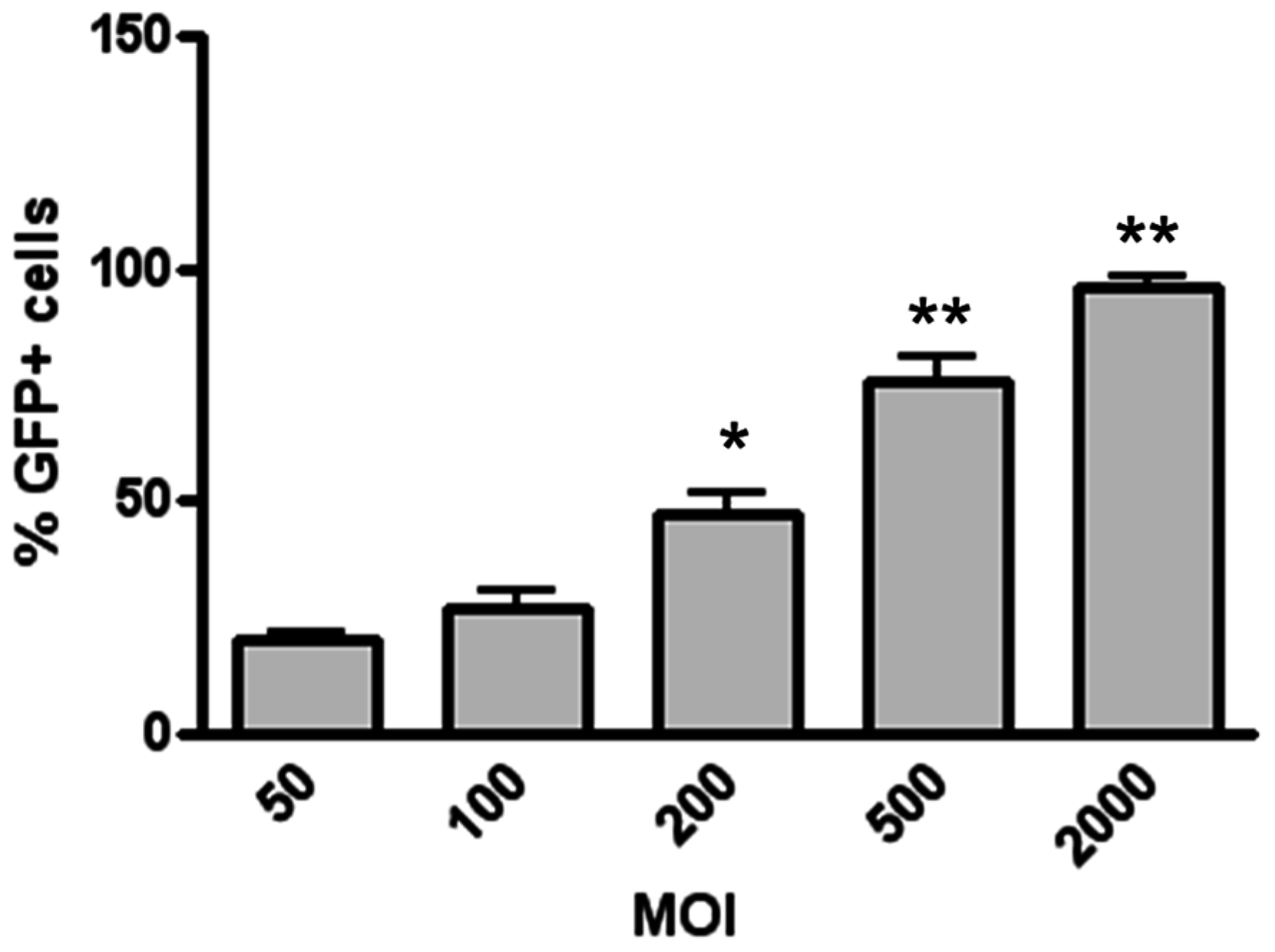
2.3. Transduction of H441 Cells with Different LV MOIs
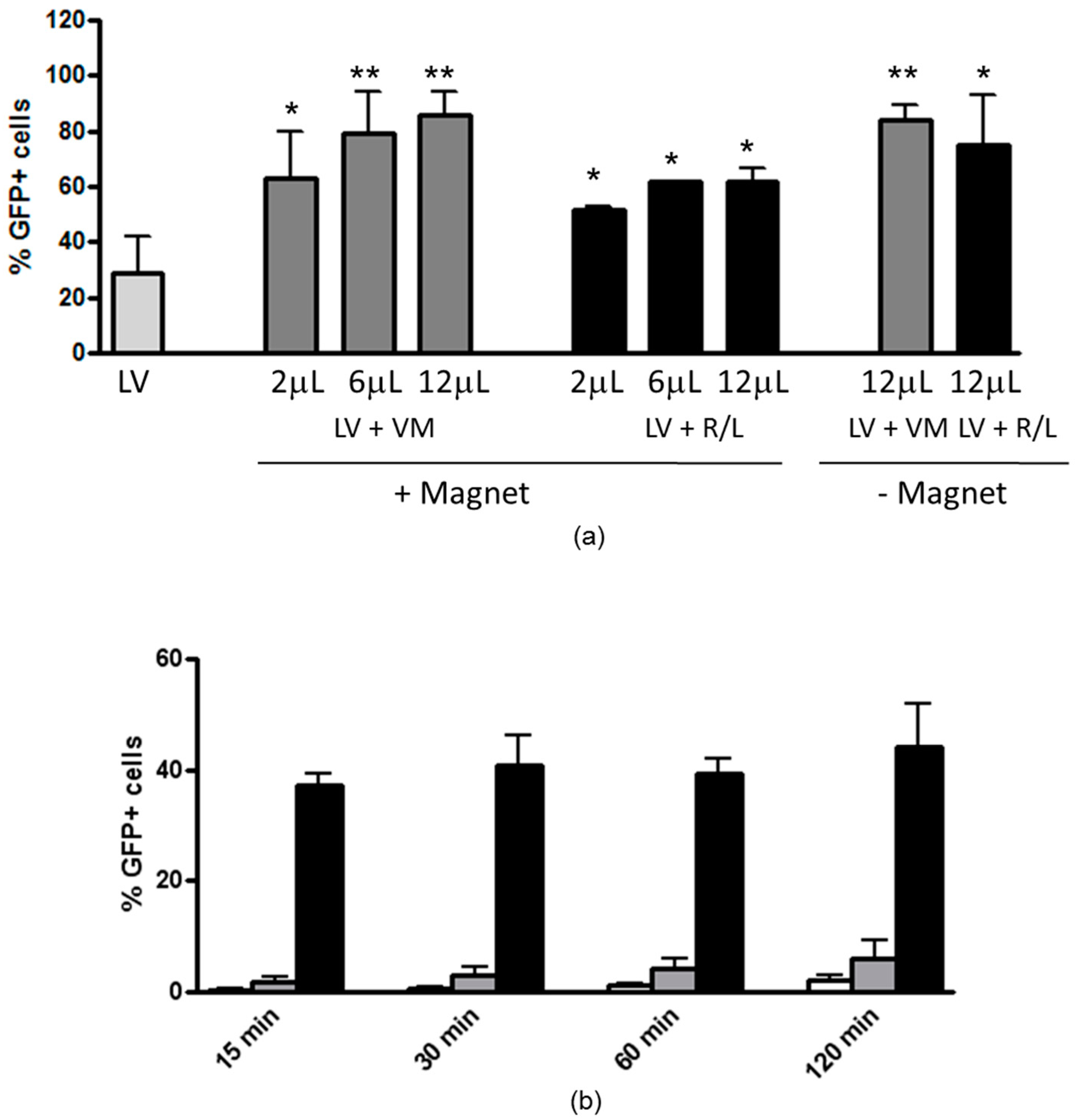
2.4. Magnetofection-Assisted Transduction of H441 Cells
2.5. Polarization of H441 Airway Epithelial Cells
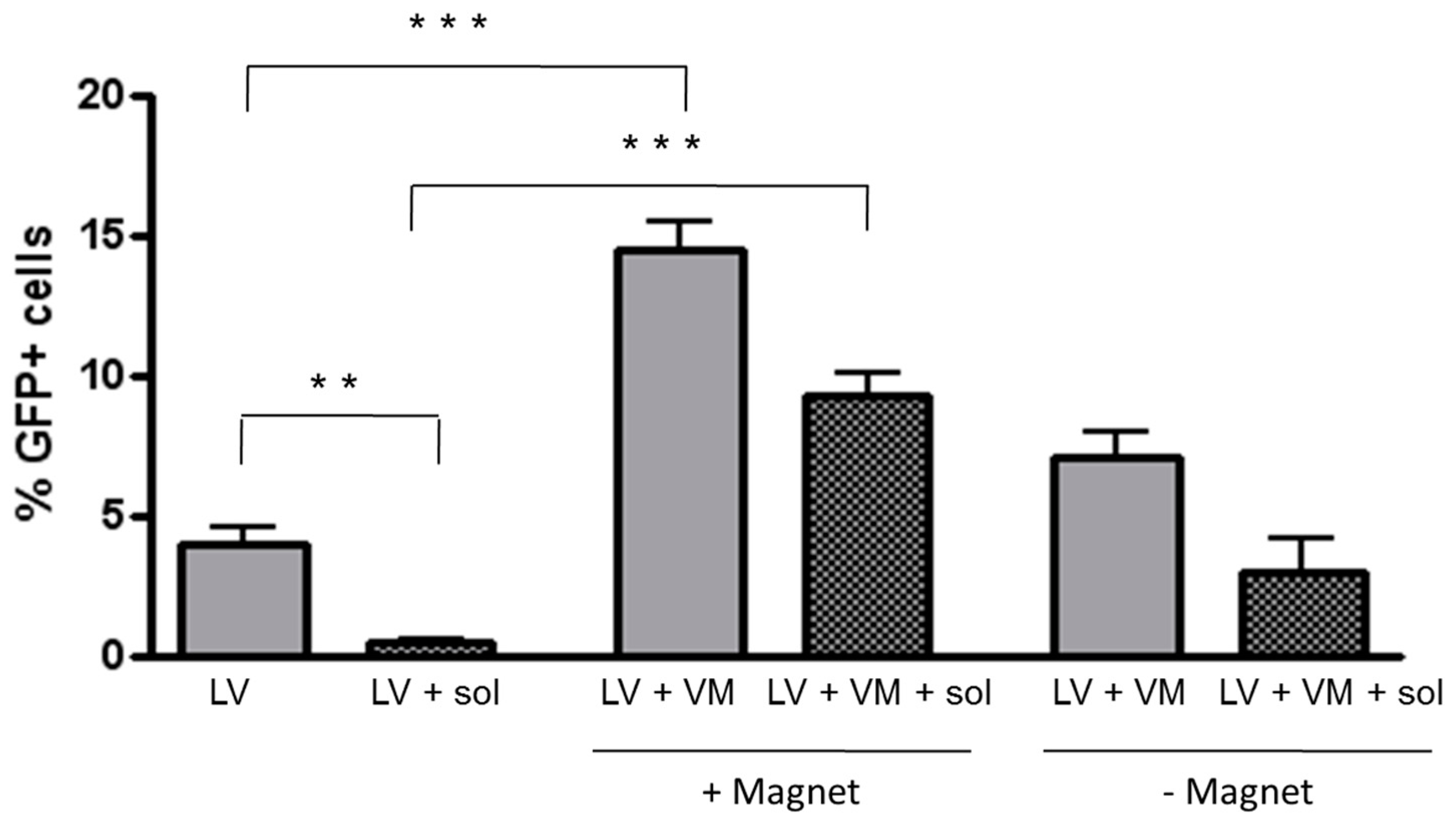
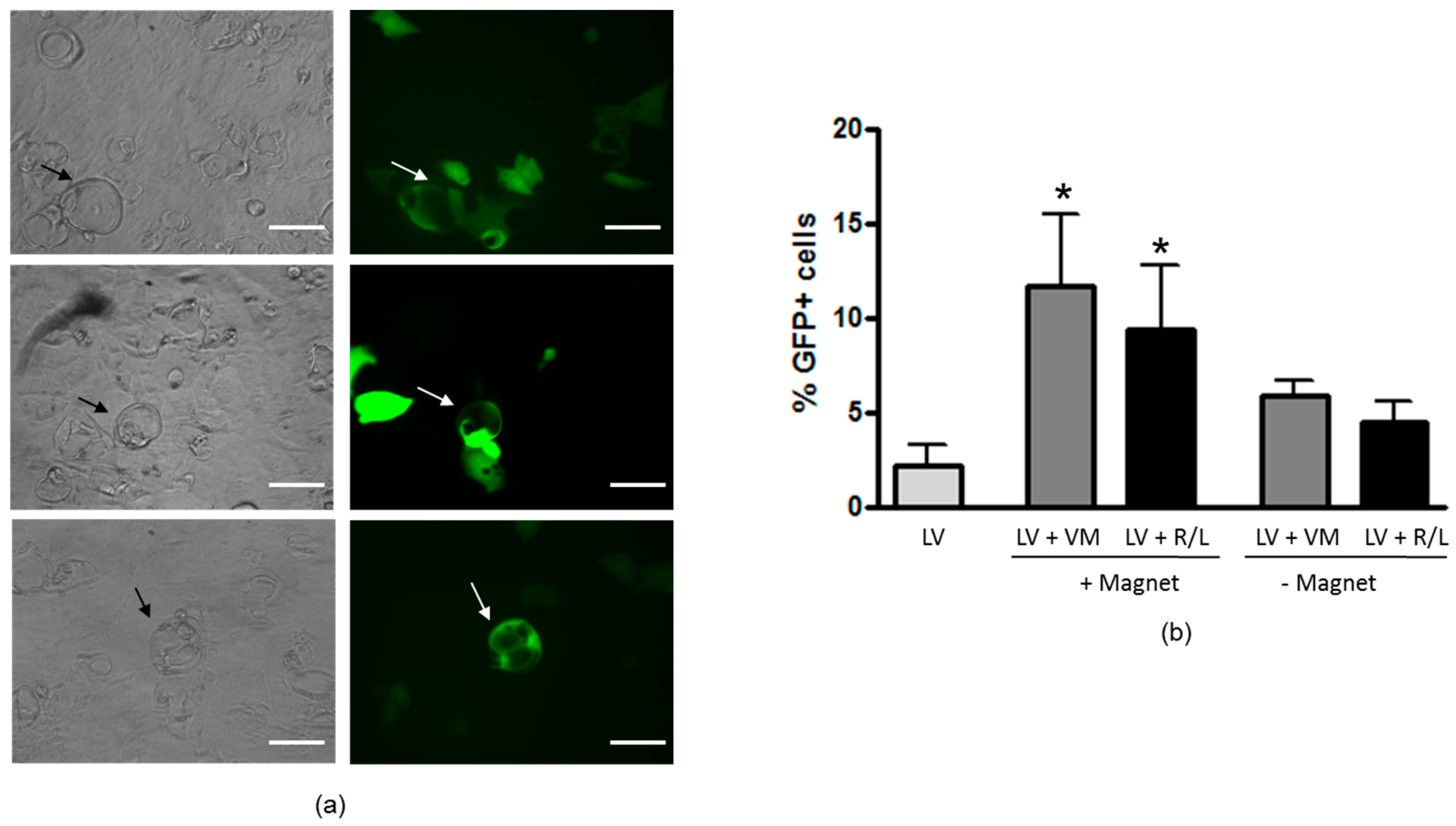
2.6. Transduction of Polarized H441 Cells in the Presence of CF Sputum Sol
2.7. Cell Culture and LV Transduction of H441 Cells Forming Domes
2.8. Zonula Occludens-1 Localization
2.9. Statistical Analysis
3. Results
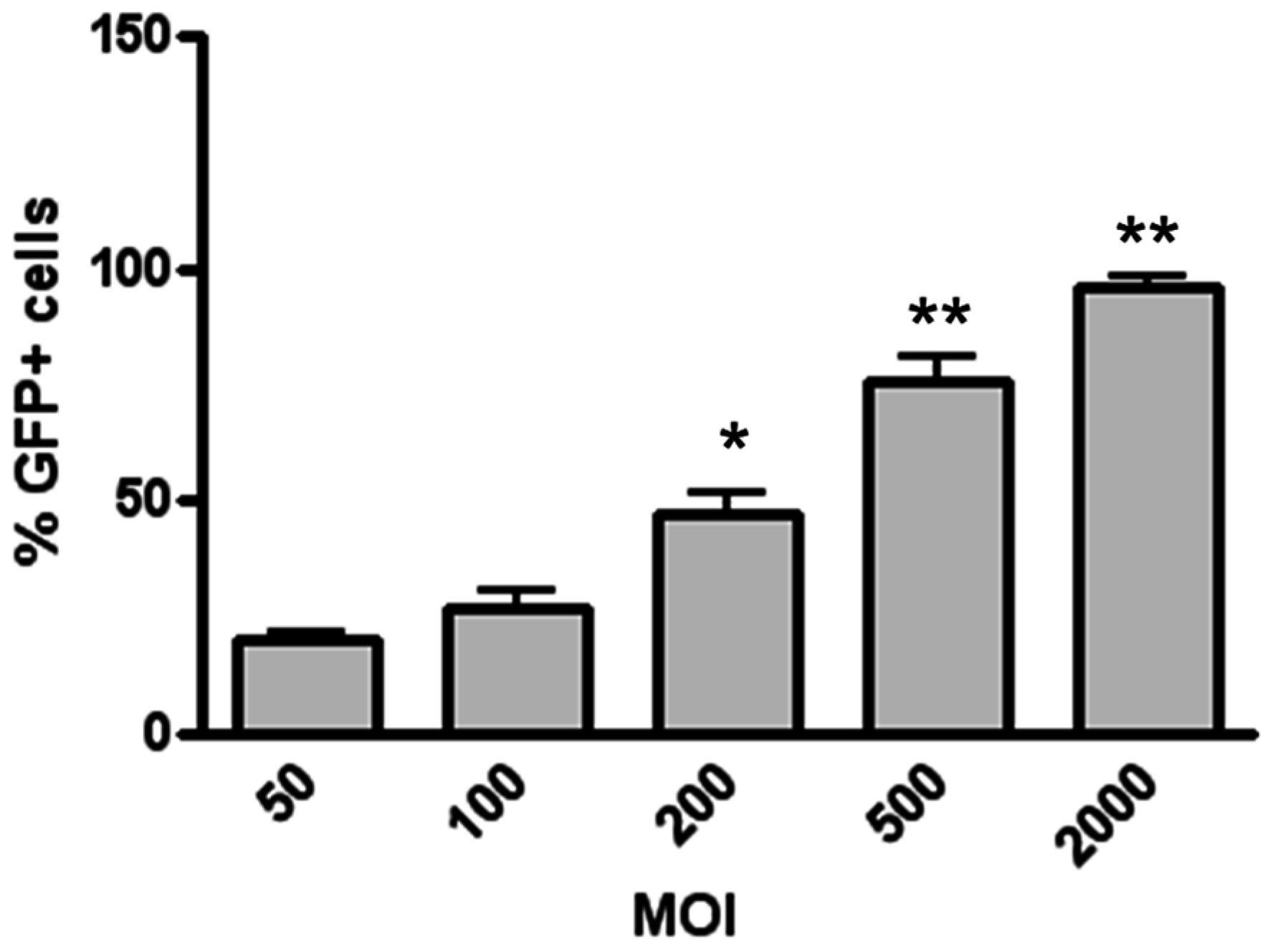
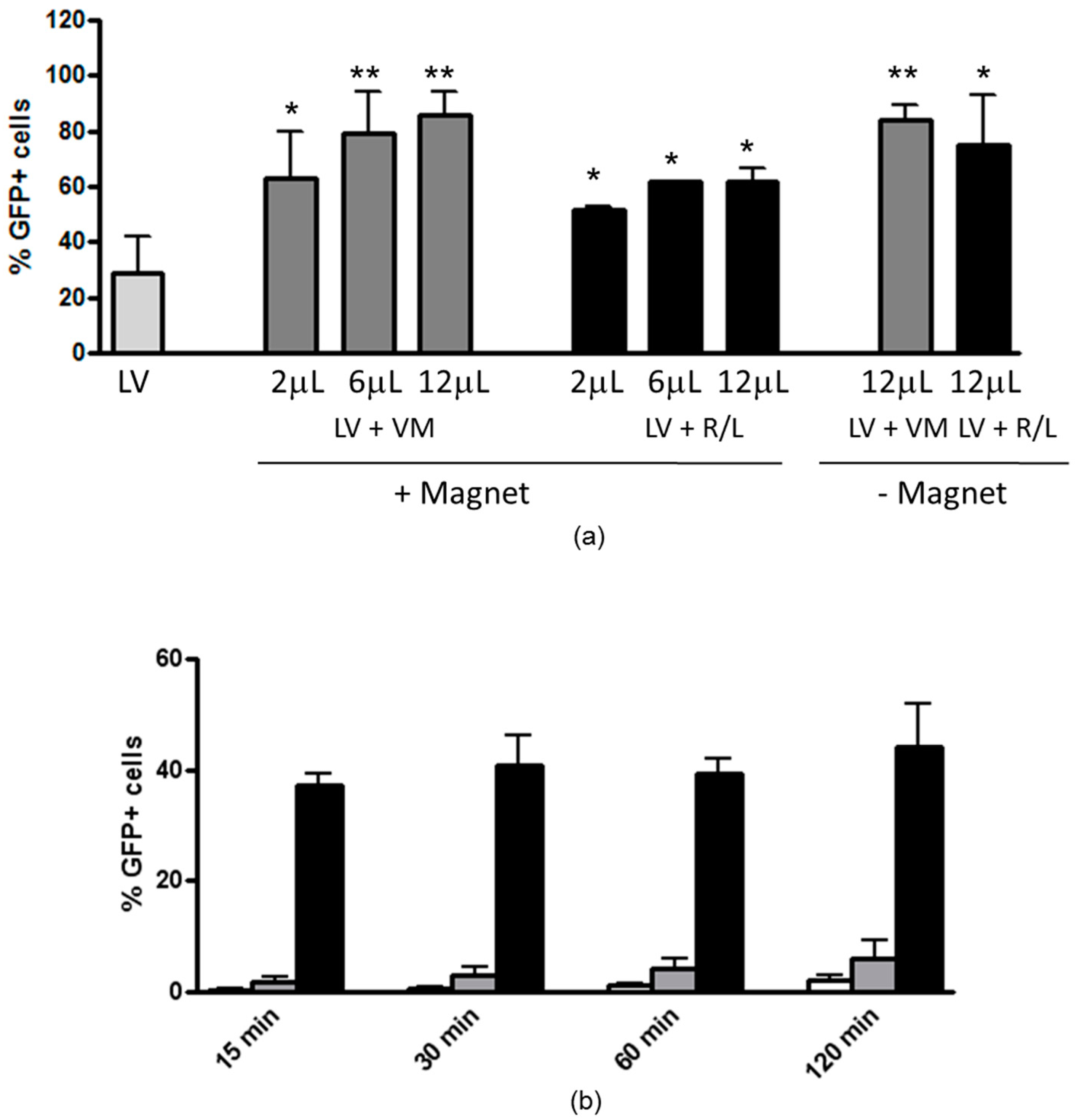
3.1. Magnetofection Enhances LV Transduction of Bronchiolar Epithelial Cells
3.2. Magnetofection Can Reduce the Time of Contact between LV and Cells to Obtain Efficient Transduction
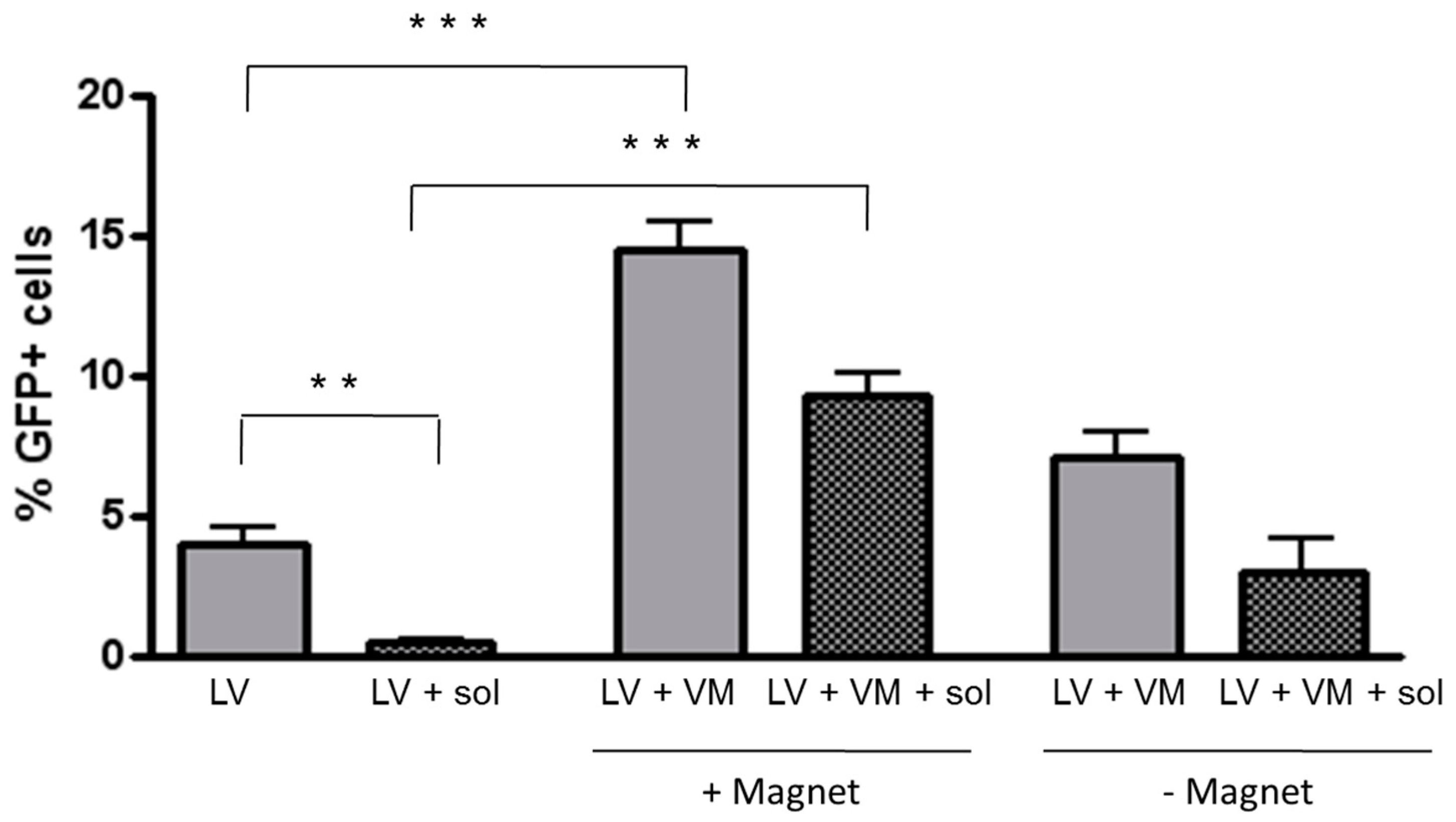
3.3. Magnetofection Enhances LV-Mediated Transduction through CF Sputum Sol
3.4. Dexamethasone Treatment Increases Tight Junction Organization and Induces Refractoriness to LV-Mediated Transduction
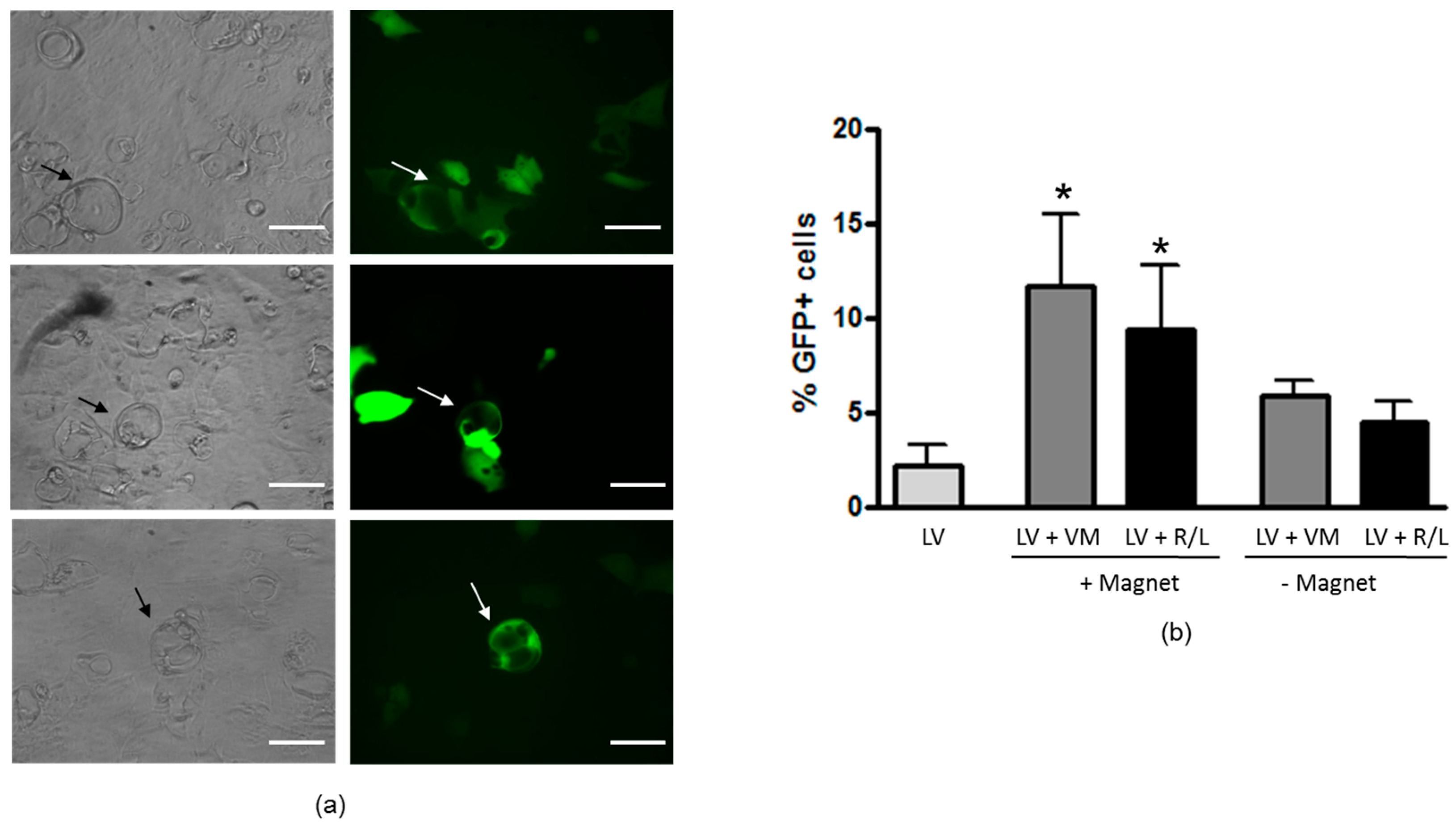
3.5. Magnetofection Increases LV Transduction of Epithelial Domes
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Griesenbach, U.; Alton, E.W. Gene Transfer to the Lung: Lessons Learned from More Than 2 Decades of CF Gene Therapy. Adv. Drug Deliv. Rev. 2009, 61, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Sanders, N.; Rudolph, C.; Braeckmans, K.; De Smedt, S.C.; Demeester, J. Extracellular Barriers in Respiratory Gene Therapy. Adv. Drug Deliv. Rev. 2009, 61, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Stern, M.; Caplen, N.J.; Browning, J.E.; Griesenbach, U.; Sorgi, F.; Huang, L.; Gruenert, D.C.; Marriot, C.; Crystal, R.G.; Geddes, D.M.; et al. The Effect of Mucolytic Agents on Gene Transfer across a CF Sputum Barrier in Vitro. Gene Ther. 1998, 5, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Kitson, C.; Farley, R.; Steel, R.; Marriott, C.; Parkins, D.A.; Scarpa, M.; Wainwright, B.; Evans, M.J.; Colledge, W.H.; et al. Mucus Altering Agents as Adjunct for Nonviral Gene Transfer to Airway Epithelium. Gene Ther. 2001, 8, 1380–1386. [Google Scholar] [CrossRef] [PubMed]
- Hida, K.; Lai, S.K.; Suk, J.S.; Won, S.Y.; Boyle, M.P.; Hanes, J. Common Gene Therapy Viral Vectors Do Not Efficiently Penetrate Sputum from Cystic Fibrosis Patients. PLoS ONE 2011, 6, e19919. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Geddes, D.M.; Alton, E.W. Barriers to and New Approaches for Gene Therapy and Gene Delivery in Cystic Fibrosis. Adv. Drug Deliv. Rev. 2002, 54, 1373–1393. [Google Scholar] [CrossRef]
- Duan, D.; Yue, Y.; Yan, Z.; Mccray, P.B.; Engelhardt, J.F. Polarity Influences the Efficiency of Recombinant Adenoassociate Virus Infection in Differentiated Airway Epithelia. Hum. Gene Ther. 1998, 9, 2761–2776. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Davidson, B.L.; Melchert, P.; Sleupushkin, V.A.; Van Hes, H.H.G.; Bodner, M.; Jolly, D.J.; Mccray, P.B., Jr. Influence of Cell Polarity on Retrovirus-Mediated Gene Transfer to Differentiated Human Airway Epithelia. J. Virol. 1998, 72, 9818–9826. [Google Scholar] [PubMed]
- Walters, R.W.; Grunst, T.; Bergelson, J.M.; Finberg, R.W.; Welsh, M.J.; Zabner, J. Basolateral Localization of Fiber Receptor Limits Adenovirus Infection from the Apical Surface of Airway Epithelia. J. Biol. Chem. 1999, 274, 10219–10226. [Google Scholar] [CrossRef] [PubMed]
- Castellani, S.; Di Gioia, S.; Carbone, A.; Conese, M. Lentiviral Vector-Mediated Gene Transfer and Applications in the Field of the Respiratory Epithelium. In Virology I—HIV and Related Issues; Iconcept Press Ltd.: Hong Kong, China, 2014. [Google Scholar]
- Oakland, M.; Sinn, P.L.; Mccray, P.B., Jr. Advances in Cell and Gene-Based Therapies for Cystic Fibrosis Lung Disease. Mol. Ther. 2012, 20, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Plank, C.; Zelphati, O.; Mykhaylyk, O. Magnetically Enhanced Nucleic Acid Delivery. Ten Years of Magnetofection-Progress and Prospects. Adv. Drug Deliv. Rev. 2011, 63, 1300–1331. [Google Scholar] [CrossRef] [PubMed]
- Orlando, C.; Castellani, S.; Mykhaylyk, O.; Copreni, E.; Zelphati, O.; Plank, C.; Conese, M. Magnetically Guided Lentiviral-Mediated Transduction of Airway Epithelial Cells. J. Gene Med. 2010, 12, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Follenzi, A.; Ailles, L.E.; Bakovic, S.; Geuna, M.; Naldini, L. Gene Transfer by Lentiviral Vectors Limited by Nuclear Translocation and Rescued by HIV-1 pol sequences. Nat. Genet. 2000, 25, 217–222. [Google Scholar] [PubMed]
- Copreni, E.; Castellani, S.; Palmieri, L.; Penzo, M.; Conese, M. Involvement of Glycosaminoglycans in Vesicular Stomatitis Virus G Glycoprotein Pseudotyped Lentiviral Vector-Mediated Gene Transfer into Airway Epithelial Cells. J. Gene Med. 2008, 10, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Aarbiou, J.; Copreni, E.; Buijs-Offerman, R.M.; Van Der Wegen, P.; Castellani, S.; Carbone, A.; Tilesi, F.; Fradiani, P.; Hiemstra, P.S.; Yueksekdag, G.; et al. Lentiviral Small Hairpin RNA Delivery Reduces Apical Sodium Channel Activity in Differentiated Human Airway Epithelial Cells. J. Gene Med. 2012, 14, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Scherer, F.; Plank, C. Magnetofection: Using Magnetic Particles and Magnetic Force to Enhance and to Target Nucleic Acid Delivery. In Gene and Cell Therapy: Therapeutic Mechanisms and Strategies; Smyth Templeton, N., Ed.; Crc Press: Boca Raton, FL, USA, 2009; pp. 379–404. [Google Scholar]
- Xenariou, S.; Griesenbach, U.; Ferrari, S.; Dean, P.; Scheule, R.K.; Cheng, S.H.; Geddes, D.M.; Plank, C.; Alton, E.W. Using Magnetic Forces to Enhance Non-Viral Gene Transfer to Airway Epithelium in Vivo. Gene Ther. 2006, 13, 1545–1552. [Google Scholar] [CrossRef] [PubMed]
- Gersting, S.W.; Schillinger, U.; Lausier, J.; Nicklaus, P.; Rudolph, C.; Plank, C.; Reinhardt, D.; Rosenecker, J. Gene Delivery to Respiratory Epithelial Cells by Magnetofection. J. Gene Med. 2004, 6, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Mykhaylyk, O.; Antequera, Y.S.; Vlaskou, D.; Plank, C. Generation of Magnetic Nonviral Gene Transfer Agents and Magnetofection in Vitro. Nat. Protoc. 2007, 2, 2391–2411. [Google Scholar] [CrossRef] [PubMed]
- Mykhaylyk, O.; Zelphati, O.; Hammerschmid, E.; Anton, M.; Rosenecker, J.; Plank, C. Recent Advances in Magnetofection and Its Potential to Deliver siRNAs in Vitro. Methods Mol. Biol. 2009, 487, 111–146. [Google Scholar] [PubMed]
- Huth, S.; Lausier, J.; Gersting, S.W.; Rudolph, C.; Plank, C.; Welsch, U.; Rosenecker, J. Insights into the Mechanism of Magnetofection using PEI-based Magnetofectins for Gene Transfer. J. Gene Med. 2004, 6, 923–936. [Google Scholar] [CrossRef] [PubMed]
- Scherer, F.; Anton, M.; Schillinger, U.; Henke, J.; Bergemann, C.; Kruger, A.; Gansbacher, B.; Plank, C. Magnetofection: Enhancing and Targeting Gene Delivery by Magnetic Force in Vitro and in Vivo. Gene Ther. 2002, 9, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Kadota, S.; Kanayama, T.; Miyajima, N.; Takeuchi, K.; Nagata, K. Enhancing of Measles Virus Infection by Magnetofection. J. Virol. Methods 2005, 128, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Antequera, Y.; Mykhaylyk, O.; Van Til, N.P.; Cengizeroglu, A.; De Jong, J.H.; Huston, M.W.; Anton, M.; Johnston, I.C.; Pojda, Z.; Wagemaker, G.; et al. Magselectofection: an Integrated Method of Nanomagnetic Separation and Genetic Modification of Target Cells. Blood 2011, 117, e171–e181. [Google Scholar] [CrossRef] [PubMed]
- Trueck, C.; Zimmermann, K.; Mykhaylyk, O.; Anton, M.; Vosen, S.; Wenzel, D.; Fleischmann, B.K.; Pfeifer, A. Optimization of Magnetic Nanoparticle-Assisted Lentiviral Gene Transfer. Pharm. Res. 2012, 29, 1255–1269. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.; Wenzel, D.; Becher, U.M.; Freitag, D.F.; Klein, A.M.; Eberbeck, D.; Schulte, M.; Zimmermann, K.; Bergemann, C.; Gleich, B.; et al. Combined Targeting of Lentiviral Vectors and Positioning of Transduced Cells by Magnetic Nanoparticles. Proc. Natl. Acad. Sci. USA 2009, 106, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Sensenig, R.; Sapir, Y.; MacDonald, C.; Cohen, S.; Polyak, B. Magnetic Nanoparticle-Based Approaches to Locally Target Therapy and Enhance Tissue Regeneration in Vivo. Nanomedicine 2012, 7, 1425–1442. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lee, M.Y.; Hogg, M.G.; Dordick, J.S.; Sharfstein, S.T. Gene Delivery in Three-Dimensional Cell Cultures by Superparamagnetic Nanoparticles. ACS Nano 2010, 4, 4733–4743. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, C.; Friedman, G.; Alamia, J.; Barbee, K.; Polyak, B. Time-Varied Magnetic Field Enhances Transport of Magnetic Nanoparticles in Viscous Gel. Nanomedicine 2010, 5, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Zahm, J.M.; Galabert, C.; Chaffin, A.; Chazalette, J.P.; Grosskopf, C.; Puchelle, E. Improvement of Cystic Fibrosis Airway Mucus Transportability by Recombinant Human DNAse is Related to Changes in Phospholipid Profile. Am. J. Respir. Crit. Care Med. 1998, 157, 1779–1784. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.R.; Svitacheva, N.; Lannefors, L.; Kornfalt, R.; Carlstedt, I. Identification of MUC5B, MUC5AC and Small Amounts of MUC2 Mucins in Cystic Fibrosis Airway Secretions. Biochem. J. 1999, 344, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Perricone, M.A.; Rees, D.D.; Sacks, C.R.; Smith, K.A.; Kaplan, J.M.; St George, J.A. Inhibitory Effect of Cystic Fibrosis Sputum on Adenovirus-Mediated Gene Transfer in Cultured Epithelial Cells. Hum. Gene Ther. 2000, 11, 1997–2008. [Google Scholar] [CrossRef] [PubMed]
- Dames, P.; Gleich, B.; Flemmer, A.; Hajek, K.; Seidl, N.; Wiekhorst, F.; Eberbeck, D.; Bittmann, I.; Bergemann, C.; Weyh, T.; et al. Targeted Delivery of Magnetic Aerosol Droplets to the Lung. Nat. Nanotechnol. 2007, 2, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Báez, C.A.A.; Cruz, I.E.L.; Padilla, M.C.R.; González, J.M.A. Magnetic Nanoparticles of Chitosan for Targeted Delivery System of Plasmids to the Lungs. J. Nanotechnol. 2014, 2014. [Google Scholar] [CrossRef]
- Kirch, J.; Guenther, M.; Doshi, N.; Schaefer, U.F.; Schneider, M.; Mitragotri, S.; Lehr, C.M. Mucociliary Clearance of Micro- and Nanoparticles is Independent of Size, Shape and Charge—An Ex Vivo and in Silico Approach. J. Control. Release 2012, 159, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Broughton-Head, V.J.; Smith, J.R.; Shur, J.; Shute, J.K. Actin Limits Enhancement of Nanoparticle Diffusion through Cystic Fibrosis Sputum by Mucolytics. Pulm. Pharmacol. Ther. 2007, 20, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Shlyonsky, V.; Goolaerts, A.; Van Beneden, R.; Sariban-Sohraby, S. Differentiation of Epithelial Na+ Channel Function. An in Vitro Model. J. Biol. Chem. 2005, 280, 24181–24187. [Google Scholar] [CrossRef] [PubMed]
- Woollhead, A.M.; Baines, D.L. Forskolin-Induced Cell Shrinkage and Apical Translocation of Functional Enhanced Green Fluorescent Protein-Human AlphaENaCin H441 Lung Epithelial Cell Monolayers. J. Biol. Chem. 2006, 281, 5158–5168. [Google Scholar] [CrossRef] [PubMed]
- Sekiyama, A.; Gon, Y.; Terakado, M.; Takeshita, I.; Kozu, Y.; Maruoka, S.; Matsumoto, K.; Hashimoto, S. Glucocorticoids Enhance Airway Epithelial Barrier Integrity. Int. Immunopharmacol. 2012, 12, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Fanning, A.S.; Van Itallie, C.M.; Anderson, J.M. Zonula Occludens-1 and -2 Regulate Apical Cell Structure and The Zonula Adherens Cytoskeleton in Polarized Epithelia. Mol. Biol. Cell 2012, 23, 577–590. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castellani, S.; Orlando, C.; Carbone, A.; Di Gioia, S.; Conese, M. Magnetofection Enhances Lentiviral-Mediated Transduction of Airway Epithelial Cells through Extracellular and Cellular Barriers. Genes 2016, 7, 103. https://doi.org/10.3390/genes7110103
Castellani S, Orlando C, Carbone A, Di Gioia S, Conese M. Magnetofection Enhances Lentiviral-Mediated Transduction of Airway Epithelial Cells through Extracellular and Cellular Barriers. Genes. 2016; 7(11):103. https://doi.org/10.3390/genes7110103
Chicago/Turabian StyleCastellani, Stefano, Clara Orlando, Annalucia Carbone, Sante Di Gioia, and Massimo Conese. 2016. "Magnetofection Enhances Lentiviral-Mediated Transduction of Airway Epithelial Cells through Extracellular and Cellular Barriers" Genes 7, no. 11: 103. https://doi.org/10.3390/genes7110103